Abstract
Soybean is an important oil crop with multiple uses. Soybeans can grow in various soil types and climates; however, salt stress reduces their yield. Plant growth-promoting microorganisms are an environmentally benign way to combat stress and boost plant tolerance. In the present study, we have identified plant growth-promoting bacteria that can produce indole acetic acid (IAA) and induce distinct growth characteristics in soybean plants under salt stress. The YNA40 isolate was identified as Acinetobacter pittii through 16S rRNA sequencing and phylogenetic analysis. A pure culture of Acinetobacter pittii YNA40 was subjected to chromatographic and mass spectrometry selected-ion monitoring (GC-MS/SIM) for IAA quantification. The results revealed that the YNA40 bacterial strain showed a significantly higher IAA concentration (473.88 ng/mL) at 4% sodium chloride (NaCl). Moreover, in a salt-stress condition, inoculation with Acinetobacter pittii YNA40 was able to induce increased shoot length (23.48%), shoot weight (24%), root length (2.47%), and root weight (44.82%) compared to the uninoculated control. Therefore, soybean seedlings were inoculated with YNA40 to examine their potential for promoting growth and reprogramming after salt stress. Inoculation with YNA40 isolates mitigated the salt stress and significantly improved the growth of the plant, enhanced the chlorophyll contents, and improved the quantum efficiency of chlorophyll fluorescence, total phenolic content, flavonoid content, the diphenyl-1-picrylhydrazyl (DPPH) activity, and antioxidant activities of soybean plants during and after salt stress. The present research demonstrated that the application of the YNA40 isolate is promising for reducing salt stress in soybean plants and helps plants grow better in a salt-stressed environment.
1. Introduction
Urbanization and industrialization have led to a reduction in the amount of land that can be used for agriculture. Several abiotic stresses in the agricultural system also affect crop yield. Abiotic stresses such as soil salinity can stunt plant development and crop productivity. Extreme salinity is usually caused by high levels of sodium (Na+) and chloride (Cl−) ions in the soil. Soil with an electrical conductivity > 4 dS m−1 is considered salt-affected because it results in increased osmotic pressure (to about 0.2 MPa) that prevents plants from efficiently absorbing water and minerals [1]. The Food and Agriculture Organization (FAO) estimates that the worldwide area of salt-affected soils spans 424 million hectares of topsoil (0–30 cm) and 833 million hectares of subsoil (30–100 cm) based on 73% of the land identified so far [2]. Several physiological and biochemical indices, including protein synthesis, photosynthesis, water status, leaf area, lipid metabolism, and membrane integrity, are affected by high concentrations of sodium chloride (NaCl) in all phases of plant development [3].
Several biotic and abiotic factors interact in the soil to promote plant development. The rhizosphere is a thin layer of soil around a plant’s root that is important for the root’s metabolism and activity [4]. There are also fungi, protozoa, and algae, though the bacterial population far outweighs the rest. Plants choose bacteria that help them grow by releasing certain organic compounds from their roots [4]. This creates a highly selective environment where only a few species of bacteria can survive, hence the diversity is limited [5,6]. This occurs as a result because microbes have the capability of producing and utilizing these compounds [7]. Moreover, the rhizosphere serves as a special ecological niche for each plant. The beneficial bacteria that live in and on plants are known as plant growth-promoting bacteria. Bacteria belonging to the genera Azospirillum [8], Azotobacter [8], Alcaligenes [9], Arthrobacter [10], Bacillus [11], Burkholderia, [12], Bradyrhizobium [13], Enterobacter [14], Pseudomonas [14], Rhizobium [15], and Serratia [16] have all been found in the rhizosphere and have been shown to promote plant growth.
IAA has been found to be a plant hormone, which is an important finding. This has led to much research on how it affects plant growth and development. It has been found that IAA plays a crucial role in plant life. It is involved in leaf formation, embryonic development, root initiation and development, abscission (leaf falling), phototropism, geotropism, and fruit development. Furthermore, IAA produces more root hairs, root laterals, and root branches, all of which help the plant absorb nutrients from its surroundings [17]. Several bacteria, fungi, and algae have been found to generate IAA in physiologically active amounts. However, the tryptophan-dependent pathway is the best-known way for plants and bacteria to make IAA [18]. Furthermore, amino acids, sugars, and organic acids are just some of the water-soluble substances that plants secrete as they expand. As a result, the resources required for the production of secondary metabolites such as IAA are readily available to the bacteria in the rhizosphere [18].
Plant growth-promoting rhizobacteria (PGPR) can alleviate salt stress by improving photosynthesis, producing reactive oxygen scavengers, phytohormones, and exopolysaccharides. Salinity can induce oxidative stress, which results in the accumulation of reactive oxygen species (ROS) that cause oxidative damage to membrane lipids, proteins, and nucleic acids. PGPR under salt-stress conditions produces antioxidant enzymes such as catalase (CAT), peroxidase (POD), superoxide dismutase (SOD), and ascorbate peroxidase, which function as a defense system against ROS. Under salt-stress conditions, exopolysaccharides (EPS) produced by PGPR bind Na+ ions and reduce their toxicity in the root region. PGPR increases the growth of plants and induces systemic resistance to environmental stresses through the antioxidant activity of enzymes, the accumulation of inorganic solutes such as sodium (Na+), magnesium (Mg2+), and potassium (K+) [19], and the reduction of ethylene levels by ACC deaminase activity [20]. Many functional groups in EPS bind with soil particles and help in soil aggregation, water retention, and the chelation of metal ions under salinity [21].
The nutritional value of soybeans and their oil-yielding properties have led to their widespread cultivation [22]. Soybean plants are very adaptable; however, they are sensitive to salt and can lose up to 40% of their seed germination, growth after germination, photosynthesis, and yield [23]. High salinity is a major cause of soybean plants’ ionic toxicity and osmotic stress [24]. Several conventional and non-conventional strategies, such as seed priming, growth regulators, signaling molecules, and salt-tolerant varieties, have been used to mitigate the negative impacts of salinity on soybeans [25]. Studies have shown that many strains of bacteria from different families can help reduce the negative effects of salinity stress [26,27,28,29]. The purpose of this research was to evaluate the effectiveness of plant growth-promoting rhizobacteria (PGPR) isolates, specifically Acinetobacter pittii YNA40, which produces IAA, in mitigating the abiotic stress caused by salt in soybean plants, as well as the oxidative damage and endogenous hormones (abscisic acid and salicylic acid) in plants subjected to salt stress.
2. Materials and Methods
2.1. Isolation, Screening, and Identification of Isolates
Rhizospheric bacteria were isolated from highland soil in Daehaw-myeon, Gangwon-do, Republic of Korea (37° 32′ 20.3″ N, 128° 27′ 05.8″ E) for this study. Plant growth-promoting traits such as the production of EPS, solubilization of phosphate, and IAA production, as well as their resilience to 0, 2, 4, 6, 8, and 10% NaCl stress, were screened. The Salkowski reagent was used to measure the production of IAA for all the isolates [30]. Briefly, the broth culture of the bacteria was centrifuged at 2500× g for 10 min at 4 °C to separate the cells from the supernatant. Subsequently, 0.45 μm cellulose acetate filters (DISMIC; Frisenette ApS, Knebel, Denmark) were used to remove debris from the resulting supernatant, and the clear supernatant was used for IAA analysis. The bacterial supernatant (2 mL) was mixed with 1 mL of Salkowski’s reagent (50 mL, 35% perchloric acid; 1 mL, 0.5 M ferric chloride). The EPS formation for all the isolates was measured using the Congo red assay, as described in the literature [31]. Furthermore, the siderophore production was measured using chromeazurol “S” agar medium, which was used for all the isolates as described in the literature [32]. Briefly, 1 mL of culture aliquots was added to 10 mL of sterilized LB broth and incubated at 28 °C. The absorbance was measured at 600 nm every eight hours (8 h, 16 h, 24 h, and 48 h) using a spectrophotometer.
Acinetobacter pittii YNA40 was also chosen for further study because it showed the most remarkable performance and survival trajectory when exposed to NaCl, EPS, siderophore, phosphate solubilizing, and IAA production. Acinetobacter pittii YNA40 was found by amplifying its genomic DNA with a 16S rRNA-specific primer, as explained in detail in [3]. The accession number is MZ596237. MEGAx was used for evolutionary research.
2.2. Growth Conditions and Treatments
The experiment was carried out at the Crop Physiology Lab of Kyungpook National University on 4th April 2022. Daewon soybean seeds were planted in 50-hole trays of autoclaved horticultural soil [33,34]. We used specific soil prepared by Punong Co., Ltd., Gyeongju, Korea, which comprised perlite (11%), cocopeat (68%), zeolite (8%), NO3− (~0.205 mg g−1), NH4+ (~0.09 mg g−1), P2O5 (~0.35 mg g−1), and K2O (~0.1 mg g−1). The soil pH was 5–7, and the EC (S/m) was ≤1.2. The seedlings were transferred into pots (10 cm × 9 cm) on the 10th day. The seedlings were cultivated in a plant growth chamber at 28 ± 0.5 °C for 16 h and 25 °C for 8 h, with 55–65% relative humidity and a light intensity of 200 µmol m−2 s−1 under long-day conditions (16 h of daytime and 8 h of dark time). The soybean seedlings were treated with 100 µM IAA (Sigma Aldrich, Burlington, MA, USA) and an inoculation with YNA40 isolate for 5 days. On day 18, 100 mM NaCl was used after 3 days of IAA and the application of the microbes [34]. We divided our treatments into two groups: (a) no-stress: (i) distilled water; (ii) 100 µM IAA; (iii) YNA40; and (b) salinity stress: (i) distilled water + 100 mM NaCl (1 S/m); (ii) 100 M IAA + 100 mM NaCl (1 S/m); (iii) YNA40 + 100 mM NaCl (1 S/m). Furthermore, to assess the defense action of plants with an isolate of Acinetobacter pittii YNA40 under salt stress, 100 mL of the bacteria was inoculated in each pot via a soil drench. In contrast, the control plants were watered with distilled water for two weeks. The growth parameters were regularly monitored after the total doses of salt stress had been administered. On the 30th day, the plants were harvested and collected in liquid nitrogen and stored at −80 °C until further biochemical analysis. A CCM-300 chlorophyll meter (ADC Bio Scientific Ltd., Herts, UK) was used to measure the chlorophyll content in the plants before harvesting. The instant chlorophyll concentration on the final day of harvesting was measured using the Minolta SPAD-502 (Konica Minolta, Tokyo, Japan) [34].
2.3. Estimation of Peroxidase
Peroxidase levels were measured in the Acinetobacter pittii YNA40-inoculated plants growing under both unstressed and salinity-stressed conditions, according to the method described by Bilal et al. [35]. After immediate harvesting, 500 mg of plant sample was centrifuged at 5000 rpm for 15 min at 4 °C to collect the supernatant. The sample was first homogenized in a 0.1 M potassium phosphate solution (pH 6.8). Next, 100 μL of the supernatant was mixed with a reaction mixture (0.1 M potassium phosphate buffer (pH 6.8), 50 μL of pyrogallol, and 50 μL of H2O2) and incubated for 5 min at room temperature (25 °C) to start the reaction. Adding a 5% H2SO4 solution halted the newly started process. Purpurogallin production was estimated by recording the absorbance change at 420 nm using a spectrophotometer.
2.4. Estimation of Catalase
Catalase activity was measured in the Acinetobacter pittii YNA40-inoculated plants grown in 100 mM NaCl environments under both unstressed and salinity-stressed conditions according to the method described by Bilal et al. [35]. Briefly, the supernatant was extracted by grinding 200 mg of plant leaves in a buffer containing 50 mM Tris HCl (pH 7.0), 3 mM magnesium chloride, 1 mM EDTA, and 1.0% polyvinylpyrrolidone. Later, 0.2 M H2O2 in a 10 mM phosphate solution (pH 7.0) was added to 0.5 mL of the supernatant, and the absorbance was recorded at 240 nm using a spectrophotometer. Standard curves determined the CAT.
2.5. Measurement of DPPH Radical Scavenging Activity
We measured the capability of the leaf samples to scavenge free radicals using the DPPH test [36]. Briefly, methanol (MeOH) was used to extract the leaf samples, and 5 mg of DPPH was dissolved in 50 mL of MeOH to create the reaction mixture. The reaction combination took 30 min at room temperature and dark incubation to reach equilibrium after adding the MeOH extract (1:1). Sample absorption was measured spectrophotometrically at 517 nm. Finally, the radical scavenging efficiency was calculated.
2.6. Quantification of Superoxide Dismutase
SOD activity in Acinetobacter pittii YNA40 under no-stress and salinity-stressed plants grown under 100 mM NaCl toxicity was measured [37]. Briefly, 1.3 mL of buffer solution containing 50 mM Tris HCl and 10 mM EDTA (pH 8.5) was used to grind the leaf tissues. Later, 100 μL of pyrogallol (7.2 mM) was added to 100 μL of the extracted sample and incubated at 25 °C for 10 min. The reaction was stopped by adding 50 μL of 1 N hydrochloric acid, and the absorbance was measured spectrophotometrically at 420 nm. The percentage of SOD activity was determined using the following equation:
where A is the extracted sample containing pyrogallol, B is the extracted sample without pyrogallol, and C is the control with a buffer solution containing pyrogallol.
2.7. Estimation of Total Polyphenol Content
The total polyphenol content was calculated using the Folin–Ciocalteu colorimetric method [38]. Leaf tissues were extracted with 80% MeOH, and 50 μL of the extract was combined at room temperature for 30 min with 1 mL of 2% sodium carbonate and 50 μL of 1 N Folin–Ciocalteu reagent. The absorbance of the mixture was recorded spectrophotometrically at 750 nm. The total polyphenol content was calculated using gallic acid as the standard.
2.8. Measurement of the Total Flavonoid Content
The total flavonoid content was measured by the method described previously in the literature [39]. Briefly, 80% MeOH was used to extract the leaves, and 100 μL of crude extract was added to a reaction mixture containing 500 μL of MeOH, 50 μL of 10% aluminum chloride, 50 μL of 1 M sodium hydroxide, and 300 μL of distilled water at room temperature for 30 min. The total content of flavonoids was measured spectrophotometrically at 510 nm using a quercetin standard.
2.9. Determination of Abscisic Acid (ABA)
The extraction and measurement of the endogenous ABA level of the Acinetobacter pittii YNA40-inoculated plants cultivated in no-stress and salinity-stressed environments were performed following the experimental protocol described by Shahzad et al. [40]. In brief, 0.5 g of freeze-dried plant materials was extracted with 95% isopropanol and 5% glacial acetic acid and augmented with 20 ng of [(±)3,5,5,7,7,7,7-d6]-ABA as an internal ABA standard. Gas chromatography–mass spectrometry (GC-MS) analysis was performed using a SIM 6890N GC instrument and a SIM 5973 network mass-selective detector on the diazomethane-treated, extracted materials (Agilent Technologies, Palo Alto, CA, USA). The measured responses to ions at m/z 190 and 162 for Me-ABA and 194 and 166 for Me-[2H6]-ABA were acquired using the Lab-Base (ThermoQuest, Manchester, UK) data system software.
2.10. Determination of Salicylic Acid (SA)
According to the method outlined by Shahzad et al., [41], we were able to extract and quantify the endogenous SA content of Acinetobacter pittii YNA40 in plants grown in 100 mM NaCl toxicity, both in the absence of stress and in the presence of salt. Extraction and high-performance liquid chromatography (HPLC) were carried out on 0.2 g of freeze-dried plant sample using a Shimadzu instrument fitted with a fluorescence indicator (Shimadzu RF-10AxL) having excitation and emission wavelengths of 305 and 365 nm, respectively, and a C18 reverse-phase HPLC column (HP Hypersil ODS, particle size 5 m, pore size 120, Waters). The pace of one milliliter per minute was kept constant.
2.11. Statistical Analysis
All the tests used a completely randomized design to apply the culture to the plants to determine the impact of PGPR. All the experiments were performed with three replicates. Duncan’s multiple range tests (DMRTs) at p < 0.05 found mean value differences. The statistical analysis system (SAS 9.1) was used for DMRT analysis, and GraphPad Prism (version 5.0, San Diego, CA, USA) was used for graphs.
3. Results
3.1. Screening for Salt Tolerance
Isolated bacterial strains such as YNA40 and MO1 have been trained to withstand greater NaCl concentrations, which might have a biological role in increasing salt stress. In the present study, the optimal bacterial growth and their level of tolerance at various concentrations (0, 2, 4, 6, 8, and 10%) of NaCl were monitored at 24 h and 48 h. Figure 1 demonstrates that the OD600 nm of the YNA40 isolate was the highest compared to the MO1 isolates. The isolated strain of YNA40 could grow at a stable optical density under 0–4% NaCl stress, showing a maximum OD at 600nm at 4% NaCl concentration. A further increase in the salt content resulted in a decrease in the growth of YNA40. Despite this, the isolated YNA40 strain maintained a sustained growth rate. Because it showed maximum growth at 4% NaCl concentration, the YNA40 isolate was used for the present investigation.
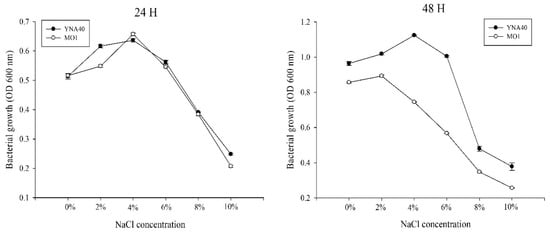
Figure 1.
Growth curve of YNA40 isolate at different salinity levels.
3.2. Quantification of IAA
The IAA concentration of the YNA40 bacterial strain was 473.88 ng/mL at 4% NaCl, which was significantly higher compared to 3% NaCl (382.06 ng/mL) and 0% NaCl (295.8 ng/mL) (Figure 2).
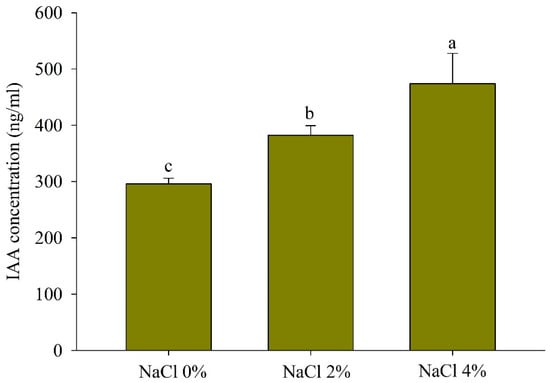
Figure 2.
IAA production by the YNA40 bacterial strain with different NaCl concentrations. Values sharing different letters are significantly different at p < 0.05.
3.3. Molecular Identification of YNA40 Bacterial Isolate
The amplification and sequencing of the 16S rRNA gene, followed by comparison with known database 16S rRNA gene sequences using the BLAST search engine of the NCBI database/EzTaxon, allowed for the molecular identification and phylogenetic study of the YNA40 isolate. The present study demonstrated that YNA40 isolate shares a high degree of sequence identity (99%) with Acinetobacter pittii, and its sequence was submitted to GenBank under the accession number MZ596237 (Figure 3).

Figure 3.
Phylogenetic tree constructed using the neighbor-joining method and maximum likelihood techniques. Based on the 16S rRNA gene sequences, showing the taxonomic position of YNA40.
3.4. Assessment of Plant-Growth Parameters under Salinity
Salt stress greatly reduced the root length, length of shoot, shoot weight, root weight, and stem diameter of the soybean seedlings. In the no-stress conditions, the application of YNA40 significantly increased the shoot length (9.39%) and shoot weight (17.24%) in comparison with the application of 100 µM IAA and the uninoculated control, whereas root length, root weight, and stem diameter were unaffected. (Table 1).

Table 1.
Inoculation with YNA40 isolate on the growth characteristics of soybean seedlings under no-stress and salt-stress environment. Data points are the mean of three replicates ± SEM. Values sharing the same letter are statistically equal at p < 0.05, whereas those sharing different letters are significantly different.
Contrarily, the application of YNA40 significantly increased shoot length (23.48%), shoot weight (24%), root length (2.47%), and root weight (44.82%) under salinity compared to the no-treatment plants. Moreover, the application of 100 µM IAA significantly increased shoot length (22.14%), shoot weight (16%), root length (1.24%), and root weight (37.93%) compared to the control. However, the stem diameter was unchanged in all the cases. Among all the treatments, the YNA40 bacterial strain produced the best results for the growth parameters of the soybean seedlings (Figure 4).

Figure 4.
Changes in soybean seedling development characteristics with inoculation with YNA40 under no stress or NaCl.
3.5. Effect on Chlorophyll Content and Chlorophyll Fluorescence
Salt stress lowers the amount of chlorophyll and the efficiency of photosynthesis system II (fv/fm) in plants. Figure 5a demonstrates that the applications of bacterial strains YNA40 and 100 µM IAA responded similarly to the chlorophyll content. They did not significantly change under salt stress compared with no stress. Contrarily, inoculation with YNA40 and 100 µM IAA under salinity resulted in statistically significant differences in chlorophyll content compared with the no-treatment condition. YNA40 and 100 µM IAA increased the chlorophyll content by 11.03% and 10.20%, respectively, compared to the control.
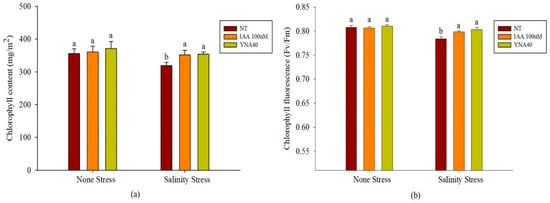
Figure 5.
Effect of inoculation with YNA40 isolate on (a) chlorophyll content and (b) chlorophyll fluorescence in a no-stress and salt-stress environment. Data points are the means of three replicates ± SEM. Values sharing the same letter are statistically equal at p < 0.05, whereas those sharing different letters are significantly different.
Similarly, the application of the bacterial strains YNA40 and 100 µM IAA responded identically to chlorophyll fluorescence in a no-stress environment. As opposed to this, under the salt-stress condition, the application of YNA40 and 100 µM IAA responded with a statistically significant difference in fv/fm compared to the no-treatment condition. The YNA40 and 100 µM IAA increased the fv/fm content by 2.42% and 1.78%, respectively, compared to their controls (Figure 5a).
3.6. Quantification of Total Phenolic Content, Total Flavonoid Content, and DPPH Activities in Soybean Seedlings
Total phenolic content, total flavonoid content, and DPPH activities are greatly influenced under salt-stress conditions. Our investigation demonstrated that the inoculation with the YNA40 isolate under no-stress conditions showed significantly increased total phenolic content compared to the application of 100 µM IAA; however, it was statistically similar to the control. The application of the YNA40 isolate increased the total phenolic content by 9.96% compared to the control (Figure 6a).
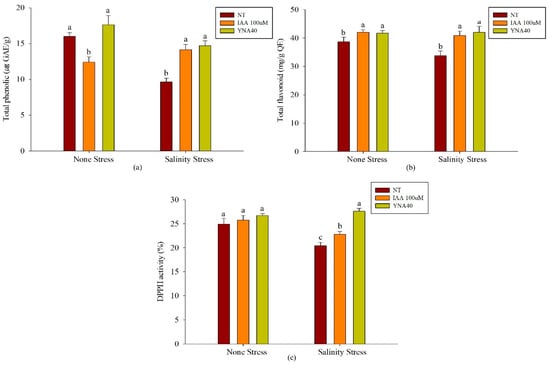
Figure 6.
Effect of inoculation with the YNA40 isolate on (a) total phenolic content, (b) total flavonoid content, and (c) DPPH activity of soybean seedlings under no-stress and salt-stress conditions. Data points are the means of three replicates ± SEM. Values sharing the same letter are statistically equal at p < 0.05, whereas those sharing different letters are significantly different.
Whereas, the application of 100 µM IAA reduced the total phenolic content by 22.62% under no-stress conditions. Under the salt-stress conditions, inoculation with the YNA40 isolate showed significantly increased total phenolic content in contrast to the control. Inoculation with the YNA40 isolate showed significantly increased total phenolic content, by 52.63%, in contrast to the uninoculated control. Conversely, the application of 100 µM IAA significantly increased the total phenolic content by 46.60% compared to the control, but it significantly decreased the total phenolic content compared with the application of the YNA40 isolate.
Figure 6b illustrates that under no-stress conditions, the application of YNA40 isolate and the application of 100 µM IAA demonstrated a significant increment in the total flavonoid content compared to the control. Furthermore, the application of YNA40 isolates to the plant increased the flavonoid content by 7.84% compared to the uninoculated control. In contrast, the application of 100 µM IAA increased the flavonoid content of the soybean plant by 8.62% compared to the control plant. Similarly, under the salt-stressed conditions, the applications of YNA40 isolate and 100 µM IAA significantly increased the total flavonoid content compared to the non-treated plants, i.e., 24.65% and 21.06%, respectively.
Furthermore, the DPPH activity on the applications of YNA40 isolate and 100 µM IAA under the no-stress conditions was statistically identical to the control. Nevertheless, there was a significant difference under salt-stressed conditions compared to the control Figure 6c. The application of YNA40 isolate increased the DPPH activity of the soybean plant by 35.07% compared to the uninoculated control, whereas the application of 100 µM IAA also increased the DPPH activity by 11.58%.
3.7. Impact of YNA40 Isolate on Different Antioxidant Enzymes
Soybean seedlings were subjected to salt stress, and their antioxidant enzyme levels were measured. Under the no-stress conditions, the application of 100 µM IAA significantly increased the amount of POD (6.32%) compared to the control (Figure 7a). Contrarily, the inoculations with the YNA40 isolate and 100 µM IAA significantly increased the POD by 28.50% and 26.30%, respectively, compared to the control under the NaCl stress.
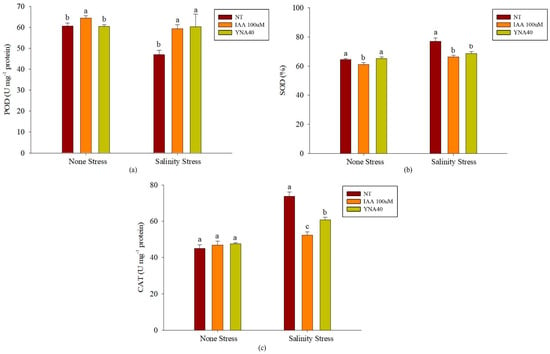
Figure 7.
Effect of YNA40 bacterial isolate on different antioxidants: (a) peroxidase (POD); (b) superoxide dismutase (SOD); and (c) catalase (CAT) content under no-stress and salt-stress conditions in soybean plants. Data points are the means of three replicates ± SEM. Values sharing the same letter are statistically equal at p < 0.05, whereas those sharing different letters are significantly different.
Furthermore, under the no-stress conditions, the application of 100 µM IAA significantly reduced the amount of SOD, among others. In contrast, the inoculation with the YNA40 isolate showed the highest increamentiof SOD by 1.29% compared to the uninoculated control (Figure 7b). However, under salinity conditions, the amount of SOD was significantly reduced by the applications of the YNA40 isolate and 100 µM IAA by 10.98% and 13.95%, respectively, compared to the control. Furthermore, under the no-stress conditions, the amount of CAT remained unchanged by the applications of the YNA40 isolate and 100 µM IAA. Contrarily, under salinity, the applications of YNA40 isolate and 100 µM IAA reduced the CAT by 17.78% and 28.99%, respectively. Moreover, the 100 µM IAA application significantly reduced the amount of catalase in the soybean plants compared to the control under salt stress (Figure 7c).
3.8. Analysis of ABA and SA in Soybean Plants
As shown in Figure 8a, the applications of YNA40 and 100 µM IAA remained statistically identical to their controls under the no-stress conditions. Under salinity, the applications of YNA40 and 100 µM IAA significantly reduced the abscisic acid content by 15.65% and 37.89% compared to the control. Between them, the application of 100 µM IAA greatly reduced the amount of abscisic acid. Furthermore, in the case of SA quantification, the soybean plants inoculated with YNA40 and 100 µM IAA have significantly reduced the SA, by 23.11% in both, compared with the control under the no salt-stress conditions. In contrast, under salt-stress conditions, the inoculation with YNA40 significantly increased the amount of SA by 72.42% compared to the uninoculated control (Figure 8b).
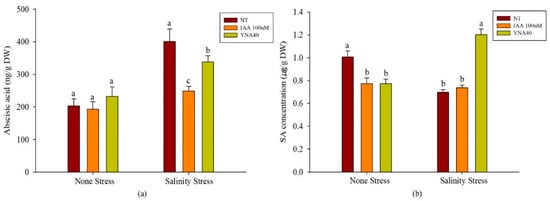
Figure 8.
Endogenous (a) abscisic acid (ABA), and (b) salicylic acid (SA) quantification in soybean plants inoculated with YNA40 under normal and salt stress. Data points are means of three replicates ± SEM. Values sharing the same letter are statistically equal at p < 0.05, whereas those sharing different letters are significantly different.
4. Discussion
Salt is a common abiotic stressor that reduces crop yield because it interferes with the ability of plants to grow and mature. Plants have a variety of defense mechanisms that have evolved to help them survive periods of high salt concentration. Rhizobacteria promote plant growth by colonizing the roots of plants and facilitating the uptake of iron and phosphorus by producing siderophores and phosphate solubilizers, respectively. Furthermore, they also help the growth and development of plants by producing biofilms, 1-aminocyclopropyl-1-carboxylate deaminase, fixing nitrogen, and phytohormones such as IAA, SA, and GA [42].
In the present study, the application of salt stress caused a significant decline in plant traits such as length of the shoot (17.6%), root length (3.2%), shoot weight (16%), and root weight (30.9%) compared to the control plant in a non-stressed condition (Table 1). Similar results of decreased growth indices have been reported in several plant species when cultivated in saline conditions [30]. The elevated salt concentrations in the soil result in the inability of the soybean plants to take up water and nutrients. This could be the result of salt toxicity or excessive osmotic pressure. Furthermore, cells often produce an excessive amount of ROS in response to salinity stress, which can be toxic, damage cell membranes, and cause cell death [43]. The present study revealed that inoculation with Acinetobacter pittii YNA40 significantly increased the growth attributes of soybean seedlings.
Conversely, all the morphological attributes displayed improvement in the plants treated with the YNA40 bacterial strains, either in the presence or absence of salt treatments (Table 1). Similar results were reported in [44], which displayed an increase in bean growth characteristics when inoculated with Pseudomonas putida and P. fluorescens under salt stress. A similar observation was found in [45], where a meaningful increment in the fresh weight of the shoot and stem length was recorded in plants treated with P. fluorescens. The mechanism indicates the production of phytohormones is responsible for the increase in roots, which help to take up nutrients and improve plant growth under salinity [46]. In the present investigation, the production of IAA is highest at a 4% concentration of NaCl (Figure 2). PGPR produces auxin and other growth-promoting substances when a plant is under salt stress. The plant’s incorporation of bacterial IAA-enhanced ACC synthase activity [47] subsequently increased ethylene, which suppresses root growth [48]. In addition, the root surface area and length are both increased by bacterial IAA, giving the plant better access to the nutrients in the soil. Furthermore, bacterial IAA relaxes plant cell walls, allowing for a greater volume of root exudation, which supplies extra nutrients to support the proliferation of rhizosphere bacteria. These chemicals help to maintain root growth and can help maintain the rate of leaf growth [49]. These microorganisms can also inhibit the movement of harmful ions, such as sodium (Na+) and chloride (Cl−) ions from plant roots to shoots [50].
Chlorophyll plays a vital role in the photosynthetic systems of plants. Beneficial microbes can maintain photosynthesis and protective mechanisms, creating a growth equilibrium in a stressful salt environment. Hence, plant biomass and grain productivity increase under stress. The present observation showed that under salt stress, the inoculation with bacterial strain YNA40 and application of 100 µM IAA significantly increased the content of chlorophyll (Figure 5a) and fluorescence compared to the control (Figure 5b). The present findings are in line with those reported by Golpayegani et al. [51].
In times of stress, plants release ROS, and an excess of these may cause oxidative stress, which is the most detrimental aspect to plants subjected to various environmental stresses. Beneficial microorganisms for plants also improve phosphorylation efficiency and eliminate free radicals. PGPR boosts the antioxidant enzymes Peroxidase (POD), Superoxide dismutase (SOD), Glutathione Reductase (GR), and Catalase (CAT) [42]. The present study described that total phenolic content (Figure 6a), total flavonoid content (Figure 6b), and DPPH activity significantly (Figure 6c) increased after inoculation with the YNA40 bacterial strain under salt stress. The present findings are consistent with those of others [52] who found a similar result after inoculation with Enterobacter sp. (S16-3) and Pseudomonas sp. (C16-20) in canola under salinity. In this study, the addition of YNA40 isolate decreased antioxidant activities, such as SOD and CAT, but increased the amount of POD (Figure 7). This could be because PGPR can enhance the growth of plants under salt stress by lowering ROS production, which, in turn, lowers the demand for antioxidant production.
Moreover, it is well known that phytohormones control various cellular processes in plants, such as how plants respond to different stresses. Hormones in this group include jasmonic acid, salicylic acid (SA), and abscisic acid (ABA) [52]. The “stress hormone,” known as ABA, actively participates in physiological processes such as active chemical cues that close stomata and conserve water [53,54,55]. However, the application of the YNA40 isolate decreased ABA accumulation in soybean plants. In [56], similar results were discovered. SA plays a crucial role in sending defense signals when plants are exposed to abiotic stresses. In the present study, the amount of SA increased significantly with the inoculation with the YNA40 isolate during salt stress (Figure 7). The results are in line with previous findings [57] demonstrating that, under salt stress, the content of SA was considerably greater in tomato plants inoculated with just the LK11 strain.
5. Conclusions
Crop failure due to salt stress is a major problem that threatens the global food supply. Climate change-induced salt stress may have long-lasting negative consequences on plants because it activates many pathways that regulate the stress-responsive expression of genes, ROS (reactive oxygen species) scavenging, and proteins and membranes of cell integrity. PGPR could be able to save crops by making phytohormones, EPS, and osmolytes, maintaining ionic homeostasis, reducing osmotic and oxidative stress, and changing antioxidative defense systems. The present study shows that IAA-producing Acinetobacter pittii YNA40 could reduce the problem caused by salt and maintain the growth characteristics of plants even when they are under salt stress. Using this microbe in our crop production system can be an excellent strategy by allowing crops to adjust to changing situations.
Author Contributions
S.-M.K. and J.-I.W. generally conducted the experimental analysis, methodology, and data interpretation. M.I.U.H. undertook the writing, conceptualization, and interpretation of the data. I.-J.L. designed and supervised the experiment. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1I1A1A01065443).
Institutional Review Board Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- FAO. Global Map of Salt-Affected Soils; FAO: Rome, Italy, 2021; pp. 1–20. [Google Scholar]
- Hmaeid, N.; Wali, M.; Mahmoud, O.M.B.; Pueyo, J.J.; Ghnaya, T.; Abdelly, C. Efficient rhizobacteria promote growth and alleviate NaCl-induced stress in the plant species Sulla carnosa. Appl. Soil. Ecol. 2019, 133, 104–113. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial Bacteria and Their Role in Plant Growth Promotion: A Review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Barriuso, J.; Solano, B.R.; Lucas, J.A.; Lobo, A.P.; García-Villaraco, A.; Mañero, F.J.G. Ecology, Genetic Diversity and Screening Strategies of Plant Growth Promoting Rhizobacteria (PGPR); Wiley-VCH Verlag GmbH and Co. KGaA: Weinheim, Germany, 2008; pp. 1–17. [Google Scholar]
- Qiao, Q.; Wang, F.; Zhang, J.; Chen, Y.; Zhang, C.; Liu, G.; Zhang, H.; Ma, C.; Zhang, J. The Variation in the Rhizosphere Microbiome of Cotton with Soil Type, Genotype and Developmental Stage. Sci. Rep. 2017, 7, 3940. [Google Scholar] [CrossRef]
- Ahmed, T.; Shahid, M.; Noman, M.; Hussain, S.; Khan, M.A.; Zubair, M.; Ismail, M.; Manzoor, N.; Shahzad, T.; Mahmood, F. Plant Growth-Promoting Rhizobacteria as Biological Tools for Nutrient Management and Soil Sustainability. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability; Kumar, A., Meena, V., Eds.; Springer: Singapore, 2019; pp. 95–110. [Google Scholar]
- Sonia, Y.; Davoud, K.; Mohamad, B.; Reza, N. Effect of Azospirillum lipoferum and Azotobacter chroococcum on germination and early growth of hopbush shrub (Dodonaea viscosa L.) under salinity stress. J. Sustain. For. 2017, 36, 107–120. [Google Scholar]
- Fatima, T.; Mishra, I.; Verma, R.; Arora, N.K. Mechanisms of halotolerant plant growth promoting Alcaligenes sp. involved in salt tolerance and enhancement of the growth of rice under salinity stress. Biotech 2020, 10, 361. [Google Scholar] [CrossRef]
- Deepti, B.; Nidhi, B.; Deepamala, M.; Chandan, S.C.; Alok, K. ACC deaminase-containing Arthrobacter protophormiae induces NaCl stress tolerance through reduced ACC oxidase activity and ethylene production resulting in improved nodulation and mycorrhization in Pisum sativum. J. Plant Physiol. 2014, 171, 884–894. [Google Scholar]
- Khan, M.A.; Hamayun, M.; Asaf, S.; Khan, M.; Yun, B.W.; Kang, S.M.; Lee, I.J. Rhizospheric Bacillus spp. Rescues Plant Growth Under Salinity Stress via Regulating Gene Expression, Endogenous Hormones, and Antioxidant System of Oryza sativa L. Front. Plant Sci. 2021, 12, 665590. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Fu, Q.; Naveed, M.; Iqbal, S.; Roitsch, T.; Jacobsen, S.-E. Burkholderia Phytofirmans Psjn Stimulate Growth and Yield of Quinoa Under Salinity Stress. Plants 2020, 9, 672. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Schwinghamer, T.D.; Smith, D.L.; Smith, D.L. Rhizobacteria from Root Nodules of an Indigenous Legume Enhance Salinity Stress Tolerance in Soybean. Front. Sustain. Food Syst. 2021, 4, 617978. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Arshad, M. Rhizobacteria containing ACC-deaminase confer salt tolerance in maize grown on salt-affected fields. Can. J. Microbiol. 2009, 55, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, Z.; Faizan, S.; Gulzar, B.; Hakeem, K.R. Inoculation of Rhizobium Alleviates Salinity Stress Through Modulation of Growth Characteristics, Physiological and Biochemical Attributes, Stomatal Activities and Antioxidant Defence in Cicer arietinum L. J. Plant Growth Regul. 2021, 40, 2148–2163. [Google Scholar] [CrossRef]
- El-Esawi, M.; Alaraidh, I.; Alsahli, A.; Alzahrani, S.; Ali, H.; Alayafi, A.; Ahmad, M. Serratia Liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [PubMed]
- Datta, C.; Basu, P.S. Indole acetic acid production by a Rhizobium species from root nodules of a leguminous shrub, Cajanus cajan. Microbiol. Res. 2000, 155, 123–127. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant. Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef]
- Egamberdiyeva, D.; Islam, K.R. Salt-tolerant rhizobacteria: Plant growth promoting traits and physiological characterization within ecologically stressed environments. In Plant-Bacteria Interactions: Strategies and Techniques to Promote Plant Growth; Springer: New York, NY, USA, 2008; pp. 257–281. [Google Scholar]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Priya, M.; Jitendra, M.; Naveen, K.A. Plant growth promoting bacteria for combating salinity stress in plants—Recent developments and prospects: A review. Microbiol. Res. 2021, 252, 126861. [Google Scholar]
- Murithi, H.M.; Beed, F.; Tukamuhabwa, P.; Thomma, B.P.H.J.; Joosten, M.H.A.J. Soybean production in eastern and southern Africa and threat of yield loss due to soybean rust caused by Phakopsora Pachyrhizi. Plant Pathol. 2016, 65, 176–188. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K.; Miransari, M. Plant Growth Promoting Rhizobacteria to Alleviate Soybean Growth under Abiotic and Biotic Stresses. In Abiotic and Biotic Stresses in Soybean Production; Academic Press: San Diego, MA, USA, 2016; pp. 131–155. [Google Scholar]
- Adhikari, B.; Dhungana, S.K.; Kim, I.D.; Shin, D.H. Effect of foliar application of potassium fertilizers on soybean plants under salinity stress. J. Saudi Soc. Agric. Sci. 2019, 19, 261–269. [Google Scholar] [CrossRef]
- Chen, H.T.; Liu, X.Q.; Zhang, H.M.; Yuan, X.X.; Gu, H.P.; Cui, X.Y. Advances in salinity tolerance of soybean: Genetic diversity heredity and gene identification contribute to improving salinity tolerance. J. Integr. Agric. 2018, 17, 2215–2221. [Google Scholar] [CrossRef]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y.; Alananbeh, K.M.; Al-Sayaydeh, R. Isolation and characterization of halotolerant plant growth promoting rhizobacteria from Durum Wheat (Triticum Turgidum subsp. durum) cultivated in saline areas of the dead sea region. Front. Microbiol 2019, 10, 1639. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Iqbal, A.; Ahmed, F.; Ahmad, M. Phytobeneficial and salt stress mitigating efficacy of IAA producing salt tolerant strains in Gossypium hirsutum. Saudi J. Biol. Sci. 2021, 28, 5317–5324. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, P.; Khan, A. Optimization of PGPR and silicon fertilization using response surface methodology for enhanced growth, yield and biochemical parameters of French bean (Phaseolus vulgaris L.) under saline stress. Biocatal. Agric. Biotechnol. 2020, 23, 101463. [Google Scholar] [CrossRef]
- Maria, J.F.; Helena, S.; Angela, C. Siderophore-Producing Rhizobacteria as a Promising Tool for Empowering Plants to Cope with Iron Limitation in Saline Soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar]
- Kang, S.-M.; Asaf, S.; Khan, A.L.; Lubna; Khan, A.; Mun, B.-G.; Khan, M.A.; Gul, H.; Lee, I.-J. Complete Genome Sequence of Pseudomonas Psychrotolerans CS51, a Plant Growth-promoting Bacterium, Under Heavy Metal Stress Conditions. Microorganisms 2020, 8, 382. [Google Scholar] [CrossRef]
- Kim, Y.N.; Khan, M.A.; Kang, S.M.; Hamayun, M.; Lee, I.J. Enhancement of Drought-Stress Tolerance of Brassica oleracea var. italica L. by Newly Isolated Variovorax sp. YNA59. J. Microbiol. Biotechnol. 2020, 30, 1500–1509. [Google Scholar] [CrossRef]
- Khan, M.A.; Sahile, A.A.; Jan, R.; Asaf, S.; Hamayun, M.; Imran, M.; Adhikari, A.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Halotolerant bacteria mitigate the effects of salinity stress on soybean growth by regulating secondary metabolites and molecular responses. BMC Plant Biol. 2021, 21, 176. [Google Scholar] [CrossRef]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Imran, Q.M.; Yun, B.W.; Lee, I.J. Osmoprotective functions conferred to soybean plants via inoculation with Sphingomonas sp. LK11 and exogenous trehalose. Microbiol. Res. 2017, 205, 135–145. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3 acetic-acid and ACC Deaminase Producing Leclercia Adecarboxylata MO1 Improves Solanum lycopersicum L. Growth and Salinity Stress Tolerance by Endogenous Secondary Metabolites Regulation. BMC Microbiol. 2019, 19, 80. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Khan, A.L.; Kang, S.M.; Imran, Q.M.; AlHarrasi, A.; Yun, B.W.; Lee, I.J. Endophytic microbial consortia of phytohormones-producing fungus Paecilomyces formosus LHL10 and bacteria Sphingomonas sp. LK11 to Glycine max L. regulates physio-hormonal changes to attenuate aluminum and zinc stresses. Front. Plant Sci. 2018, 9, 1273. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free-radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Adhikari, A.; Khan, M.; Lee, K.-E.; Kang, S.-M.; Dhungana, S.; Bhusal, N.; Lee, I.-J. The Halotolerant Rhizobacterium—Pseudomonas Koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.). Microorganisms 2020, 8, 1256. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Dhungana, S.K.; Ali, M.W.; Adhikari, A.; Kim, I.-D.; Shin, D.-H. Resveratrol, Total Phenolic andFlavonoid Contents, and Antioxidant Potential of Seeds and Sprouts of Korean Peanuts. Food Sci. Biotechnol. 2018, 27, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.J.; Liu, W.; Xiong, S.H.; Tang, J.; Lou, Z.H.; Xie, M.X.; Xia, B.H.; Lin, L.M.; Liao, D.F. Determination of Total Flavonoids Contents and Antioxidant Activity of Ginkgo Biloba Leaf by Near-infrared Reflectance Method. Int. J. Anal. Chem. 2018, 2018, 8195784. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Asaf, S.; Khan, M.A.; Kang, S.M.; Yun, B.W.; Lee, I.J. Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol. Biochem. 2016, 106, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Asaf, S.; Lee, I.J. Plant growth-promoting endophytic bacteria versus pathogenic infections: An example of Bacillus amyloliquefaciens RWL-1 and Fusarium oxysporum f. sp. Lycopersici in tomato. PeerJ 2017, 5, e3107. [Google Scholar] [CrossRef]
- Asif, M.; Pervez, A.; Ahmad, R. Role of Melatonin and Plant-Growth-Promoting Rhizobacteria in the Growth and Development of Plants. CLEAN—Soil Air Water 2019, 47, 1800459. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A.L. Alleviation of salt stress-induced adverse effects on maize plants by exogenous application of indoleacetic acid (IAA) and inorganic nutrients-A field trial. Aust. J. Crop Sci. 2013, 7, 249–254. [Google Scholar]
- Mo, W.; Tang, W.; Du, Y.; Jing, Y.; Bu, Q.; Lin, R. PHYTOCHROME-INTERACTING FACTOR-LIKE14 and SLENDER RICE1 interaction controls seedling growth under salt stress. Plant Physiol. 2020, 184, 506–517. [Google Scholar] [CrossRef]
- Metwali, E.M.; Abdelmoneim, T.S.; Bakheit, M.A.; Kadasa, N.M. Alleviation of salinity stress in faba bean (‘Vicia faba’ L.) plants by inoculation with plant growth promoting rhizobacteria (PGPR). Plant Omics 2015, 8, 449–460. [Google Scholar]
- Abulfaraj, A.A.; Jalal, R.S. Use of plant growth-promoting bacteria to enhance salinity stress in soybean (Glycine max L.) plants. Saudi J. Biol. Sci. 2021, 28, 3823–3834. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.B. Ethylene in root growth and development. In The Plant Hormone Ethylene; CRC Press: Boca Raton, FL, USA, 2018; pp. 159–181. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Mishra, S.; Dixit, V.; Kumar, M.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [PubMed]
- Albacete, A.; Ghanem, M.E.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Martínez, V.; Pérez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef] [PubMed]
- Golpayegani, A.; Tilebeni, H.G. Effect of biological fertilizers on biochemical and physiological parameters of basil (Ocimum basilicum L.) medicine plant. Am.-Eurasian J. Agric. Environ. Sci. 2011, 11, 445–450. [Google Scholar]
- Neshat, M.; Abbasi, A.; Hosseinzadeh, A.; Sarikhani, M.R.; Dadashi Chavan, D.; Rasoulnia, A. Plant Growth Promoting Bacteria (PGPR) Induce Antioxidant Tolerance Against Salinity Stress Through Biochemical and Physiological Mechanisms. Physiol. Mol. Biol. Plants 2022, 28, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Anmol, G.; Smita, R.; Ambreen, B.; Afreen, K.; Swati, S.; Neelam, P. Comparative Evaluation of Different Salt-tolerant Plant Growth-promoting Bacterial Isolates in Mitigating the Induced Adverse Effect of Salinity in Pisum Sativum. Biointerface Res. Appl. Chem. 2021, 11, 13141–13154. [Google Scholar] [CrossRef]
- Asif, M.; Pervez, A.; Irshad, U.; Mehmood, Q.; Ahmad, R. Melatonin and plant growth-promoting rhizobacteria synergistically alleviate the cadmium and arsenic stresses and increase the growth of Spinacia oleracea L. Plant Soil. Environ. 2020, 66, 234–241. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.M.; Kim, K.M.; Lee, I.J. Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PLoS ONE 2020, 15, e0232228. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.M.; Kim, K.M.; Lee, I.J. Thermotolerance effect of plant growth-promoting Bacillus cereus SA1 on soybean during heat stress. BMC Microbiol. 2020, 20, 175. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).