1. Introduction
Amaranth (
Amaranthus spp.) is a grain crop similar to cereals with desirable agronomic peculiarities and elevated nutritional value suitable for animal nutrition [
1]. The genus
Amaranthus consists of several species, from annual to short-lived perennial plants, and can be grouped into grain and vegetable types. The most important species are
A. cruentus,
A. hypochondriacus,
A. hybridus, and
A. caudatus, which are mainly used for grain production, while
A. tricolor,
A. dubius, and
A. lividus are used for vegetable production [
2]. Recently, grain amaranth has gained popularity as a feed and food crop [
3] due to its excellent nutritional value [
4]. The protein content is higher (13.0–19.85%) than other cereal grains, with a favorable amino acid profile [
5], more lysine than soybean, and relatively abundant sulfur-containing amino acids, which are usually limited in pulse crops [
6,
7]. Moreover, unlike other cereals, amaranth proteins consist of albumins (about 40%), glutenins (25–30%), and globulins (20%) and contain very small amounts of prolamins (2–3%). Amaranth prolamins are richer in glutamic acid and essential amino acids than albumins and globulins, which makes amaranth a good protein source closer to the composition of animal proteins [
8]. The lipid content (5.5–16.7%) is also higher than conventional cereals [
9], and it is characterized by high levels of unsaturated fatty acids [
10], with a saturated/unsaturated fatty acid ratio ranging from 0.12 to 0.50 [
2,
11,
12]. Starch is the main component of amaranth grain, although it is slightly lower (48–69%) than in common cereals [
2,
3]. The total dietary fiber in amaranth grains is slightly lower (7.1–16.4%) than in wheat [
2]; nonetheless, dietary fiber has been reported to be comparable to quinoa and other cereals [
5,
8,
13]. Furthermore, the high crude fiber content in amaranth grains has been associated with monogastric animals’ gut health [
14].
Currently, limited literature exists on the use of amaranth grains in pig nutrition. Zralý et al. [
15], tested diets containing 10% amaranth grain on fattening pigs and obtained some remarkable results in growth efficiency, live body weight gains, and health status of pigs during the entire experiment (pre-fattening, fattening stages I and II). Shilov and Zharkovskii [
16] reported an increase in digestibility when amaranth was added to pig diets, and Manyelo et al. [
3] demonstrated positive results in pig growth performance. Unlike conventional cereals, the proteins and dietary fiber contents in amaranth grains meet the pigs’ nutritional requirements [
4]. Furthermore, it can be expected that the high content of essential fatty acids could positively modify the lipid composition of the tissues producing healthy meat [
17].
A comparison between eight accessions of
Amaranthus cruentus grown in the Mediterranean area highlighted large genotypic variability of agronomic traits [
18], with yields comparable to barley and oat [
11]. Taking into account our previous study [
19], where
A. cruentus grains were evaluated as a component of the feeding plan for dairy cows for productive, qualitative, and in vitro fermentation traits, the present study addresses the agronomic traits, proximate composition, in vitro degradability, and fatty acid content of twelve accessions of amaranth, belonging to
A. cruentus,
A. hypochondriacus,
A. hybridus, and
A. tricolor species, grown in a field trial in a semiarid Mediterranean area. The hypothesis of the investigation is that the nutritional characteristics of amaranth (energy density, protein, starch, and fat content) are useful for pig diets; in fact, the main objective was to explore the quantitative and qualitative profile of amaranth grains as a feed ingredient in swine nutrition.
4. Discussion
Across the average of investigated accessions,
A. tricolor and
A. hypochondriacus showed the significantly highest thousand seed weight (TSW), with an accession range of 0.79–0.90 g for
A. tricolor and 0.68–0.85 g for
A. hypochondriacus. The present TSW range of
A. hypochondriacus is slightly higher than that found by Pospišil et al. [
25], who reported 0.69–0.73 g in
A. hypochondriacus across the average of nitrogen fertilization levels in a three-year field trial in Zagreb, Croatia. The
A. tricolor TSW range was also wider as compared with 10 genotypes of
A. tricolor (0.73–0.85 g) grown in Raipur, India [
26]. Given the high polymorphism of accessions within the same amaranth species, differences might be ascribed to the genotype effects. However, growing conditions cannot be ruled out as demonstrated by Pospišil et al. [
25], who pointed out a lower TSW of
A. hypochondriacus under drier than more favorable years. In this study,
A. cruentus showed lower TSW than the previous two species (0.43–0.75 g), and this somehow contrasts with results found in the literature of similar TSW between
A. hypochondriacus and
A. cruentus [
25,
27]. Nonetheless, our TSW with
A. cruentus is within the range reported by Rivelli et al. [
28] in a comparison of five different accessions of
A. cruentus tested in South Italy. The
A. hybridus accessions showed the lowest TWS (0.27–0.34 g) and it is similar to the lowest range found by Rivelli et al. [
28]. However, this contrasts with Parveen et al. [
29], who reported a TSW of 0.55 g averaging eight
A. hybridus genotypes.
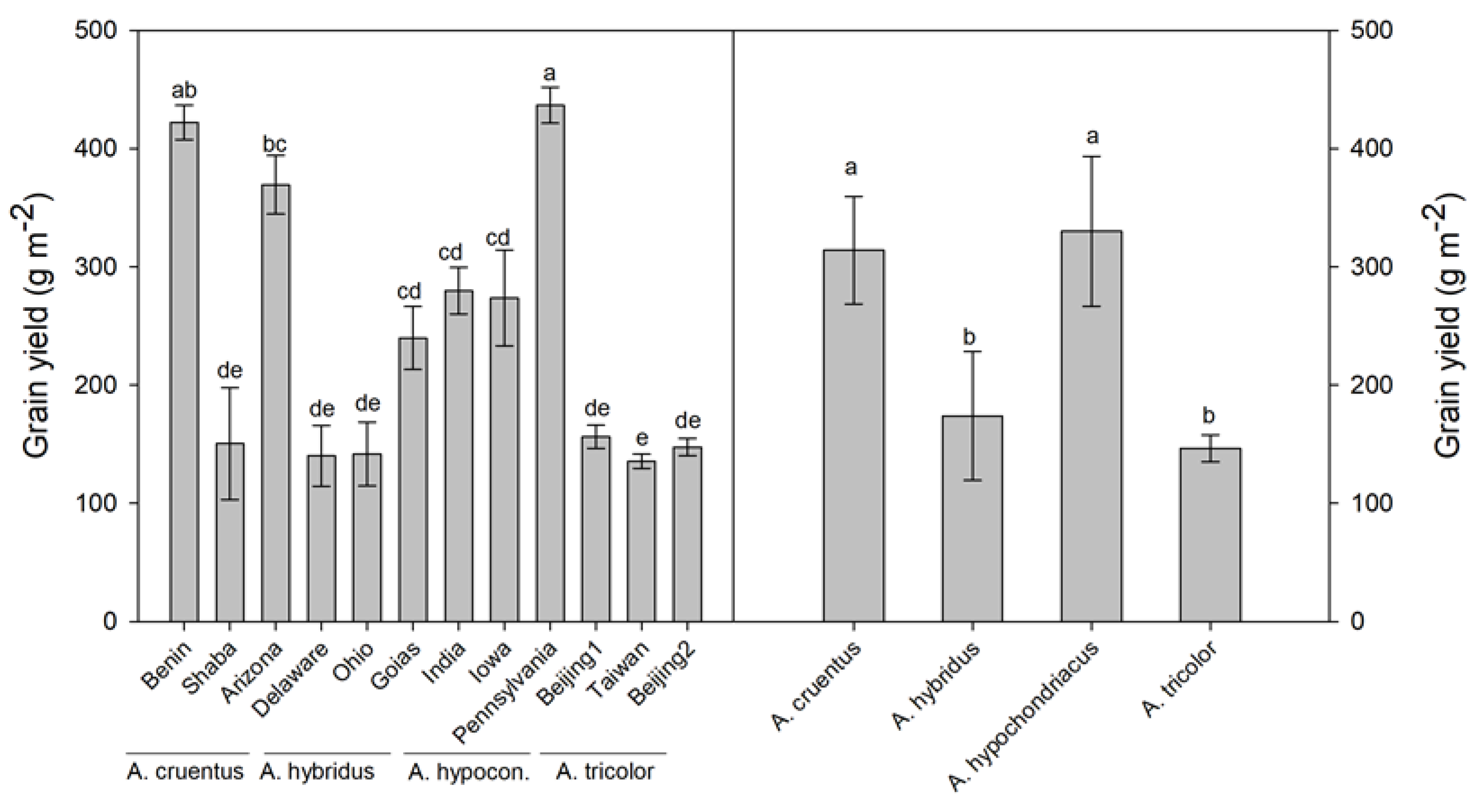
Seed yield was similar between
A. cruentus and
A. hypochondriacus, which agrees with the findings of Pospišil et al. [
25], who demonstrated no genotypic differences between
A. cruentus and
A. hypochondriacus in two out of three growing seasons. However, we found a larger accession influence on grain yield for
A. cruentus than for
A. hypochondriacus, as the coefficient of variation of accessions was 46% and 28%, respectively.
A. hybridus and
A. tricolor were both less productive, and the coefficient of variation of accessions was quite narrow for
A. tricolor (7.1%) as compared to
A. hybridus (32.8%). Grain yields of the best accessions (Pennsylvania, Benin, Shaba) were well comparable or even higher than most cereals grown in semiarid Mediterranean environments, such as durum wheat [
30,
31] and oat [
32].
In contrast to Pospišil et al. [
25], who found an increase in seed yield associated with the increase in TSW in a dry growing season, our findings did not show a significant correlation between seed yield and TSW (
p = 0.667, data not shown). However, the same author pointed out no correlation between seed yield and TSW with more favorable growing conditions. It is worth mentioning that in the present study, the air temperature was typical of the experimental area and rainfall was well distributed [
11]; this suggests that traits other than TSW, such as the seed number per panicle or the number of panicles per plant, might have influenced seed yield of investigated accessions.
The organic matter degradability (OMD) of tested
Amaranthus spp. was quite low for all samples. In particular, for
A. tricolor and
A. hybridus species, the OMD was less than 50%. These results could be ascribed to the presence of insoluble dietary fiber in the tested amaranth, as suggested by the significant negative correlations (
Table 7) between the insoluble fiber and OMD and OMCV values (−0.8577 and −0.8389, respectively).
On the other hand, insoluble fiber helps to maintain normal gut function but might decrease feed intake and nutrient digestibility [
33], increasing the rate of gut passage [
34]. Moreover, Acosta et al. [
35] tested the addition of distillers dried grains in pig diet, observing that starch digestibility can be affected by insoluble fiber level. Despite the lack of an enzymatic digestion test in this study, the starch content positively affected the fermentation parameters (0.6964 and 0.6613 for OMD and OMCV, respectively).
Starchy feeds as a source of energy and raw cereal grains, along with legume grains and potato starch, constitute the main dietary starch source in pig rations [
36]. The concentration of SCFA was quite low for all tested samples; the presence of insoluble dietary fiber could affect the fermentation pathway, as demonstrated by the limited production of SCFA of
A. hybridus and some accessions of
A. tricolor (Beijing1 and Beijing2).
A. hypochondriacus showed higher production of SCFA and butyrate probably due to the higher proportion of soluble dietary fiber, which is readily fermentable [
37]. In this regard, the correlations between chemical composition and fermentation parameters demonstrated a positive correlation between starch, SDF vs. SCFA. As suggested by Weaver et al. [
38], SDF produced more SCFA compared to IDF. Furthermore, starch feeds that bypass digestion in the stomach and enzymatic hydrolysis in the small intestine are fermented in the large intestine, producing SCFA [
39]. Furthermore, the content of crude protein positively affected the fermentation process, particularly butyrate production (0.7671). The high proportion of butyrate could be useful for the colonic epithelium as a main energy source for cell growth and differentiation [
40], suggesting a potential pre-biotic role of
A. hypochondriacus.
In swine nutrition, it is well known that the dietary FA composition and the molecular structures (chain length and number of double bonds) influence digestion, absorption, and metabolism, and the bioactivity of the FA [
41]. The manipulation of lipids in pig diet, especially in the chain length of dietary FA, may be a strategic tool to improve animal performance [
41], as explained by the complexity of digestion and absorption of these molecules. The addition of lipids to diets can, in turn, enhance protein digestibility by slowing the passage rate in the intestine by lipids, which contrasts with the effect of fiber [
42]. However, still, little focus has been devoted to the impact of lipids, in terms of fatty acids, particularly on gut health and the development of early nutrition of pigs [
43].
In general, pigs digest unsaturated dietary lipids more than saturated lipids [
44]. Hence, lipase activity and lipid digestion can be positively influenced by unsaturated long-chain fatty acids (LCFA) and negatively by saturated LCFA [
41]. Moreover, among unsaturated LCFA, those of the n3 series are believed to affect the gut microbiota mainly through regulation of the type and number of bacteria in the gut, and regulation of SCFA concentrations [
45]. This could explain the results obtained in this study where oleic and alpha-linolenic acids as well as total MUFA and n3-PUFA showed positive correlations with OMD (0.680, 0.532, 0.601, 0.532, respectively) and OMCV (0.709, 0.550, 0.627, 0.550, respectively). Conversely, among unsaturated LCFA, linoleic acid and n6-PUFA showed negative correlations with OMD (−0.409) or were not significantly correlated with OMCV. To further confirm these observations, a positive correlation for alpha-linolenic acid and n3-PUFA with SCFA (0.469) and a non-significant correlation between n6-PUFA and SCFA was found. Among the varieties,
A. hypochondriacus, with the highest content of alpha-linolenic acid and n3-PUFA (0.54 g 100g
−1), presented the highest OMD (77.6%), OMCV (182 mL g
−1), and SCFA production (52.6 mmol g
−1) and the highest levels of propionate and butyrate (24.2% and 20.7% SCFA, respectively). From a nutritional point of view, it is noteworthy that the metabolites produced by the gut microbiota significantly influence the host’s metabolism and health [
46]. The proportion of SCFA, produced by bacterial fermentation in the gut, exerts several effects on the host’s metabolism and immune system [
47]. SCFA, produced via microbial fermentation of non-digestible carbohydrates and digestible starch in the hindgut, contribute with an energy supply for the host and the colonocytes [
48] and have antimicrobial properties limiting the risk of infectious diseases in the gut [
41].



 ,
, 






{kind=link}
{kind=link}
{kind=link}