
Growth Performance of Photoperiod-Sensitive Rice (Oryza sativa L.) Varieties in Different Soil Types under Rainfed Condition in Cambodia


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.1.1. Trial in 2019
2.1.2. Trial in 2020
2.2. Environments and Experimental Design
2.2.1. Trial in 2019
2.2.2. Trial in 2020
2.3. Measurements
2.3.1. Trial in 2019
2.3.2. Trial in 2020
2.4. Statistical Analysis
3. Results and Discussion
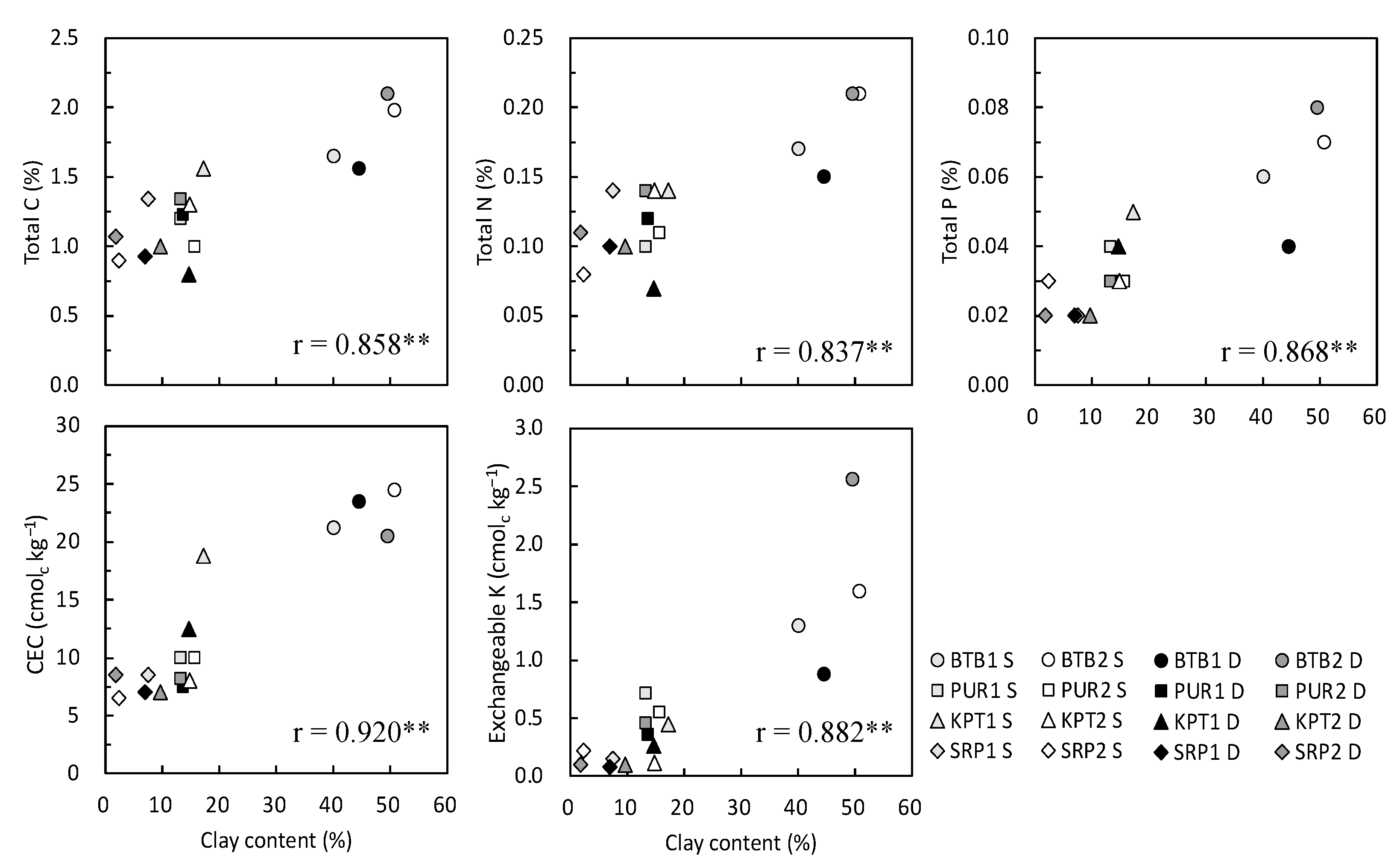
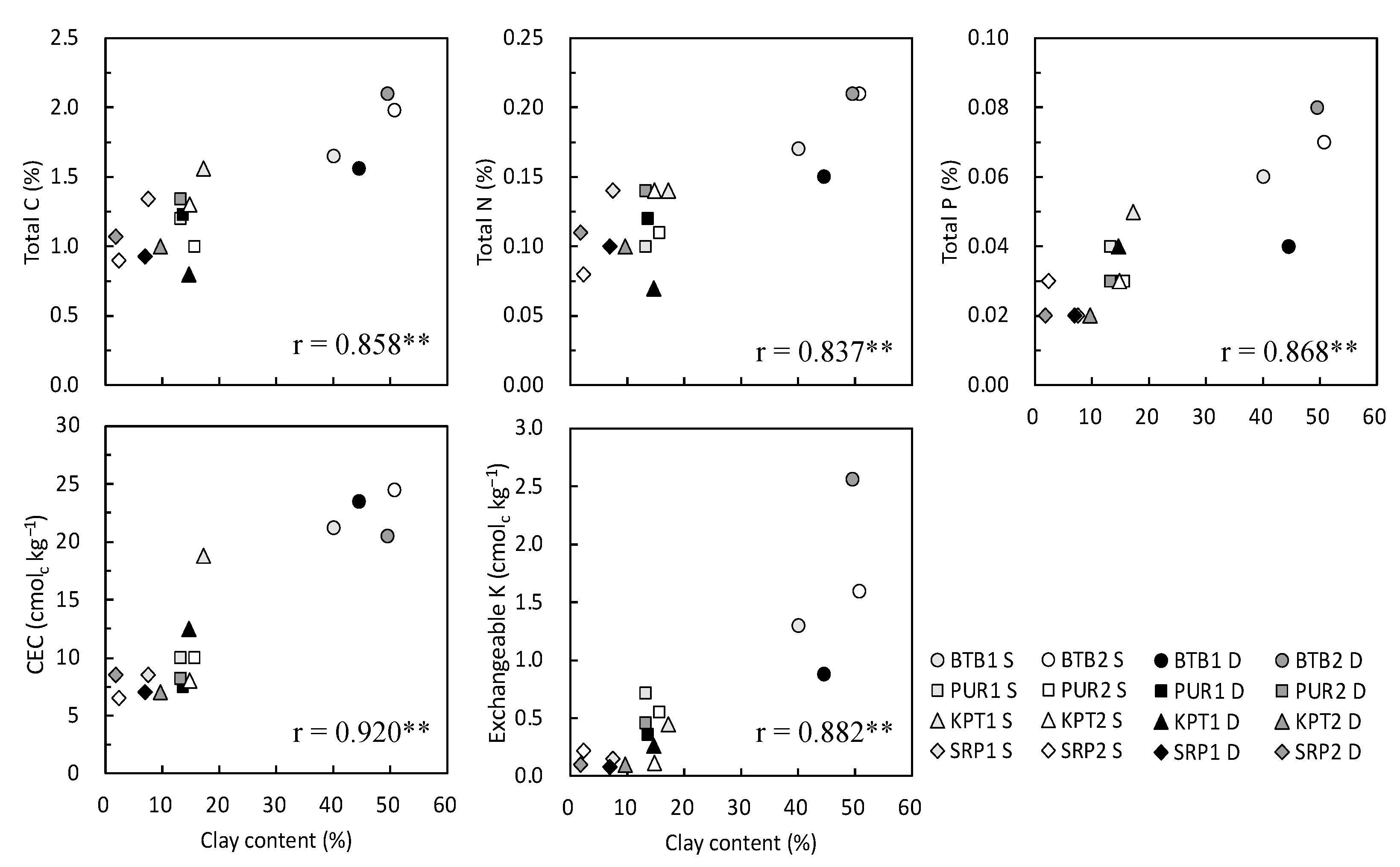
3.1. Soil Properties at the Experimental Sites
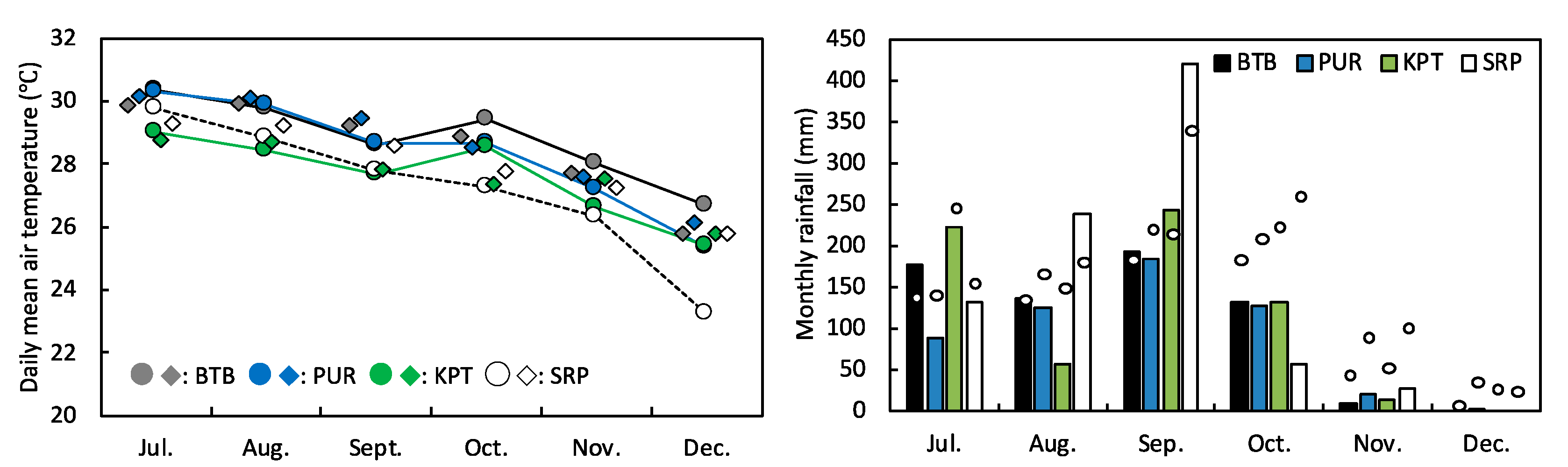
3.2. Climatic Conditions of the Experimental Sites in 2019
3.3. Yield-Related Traits in 2019
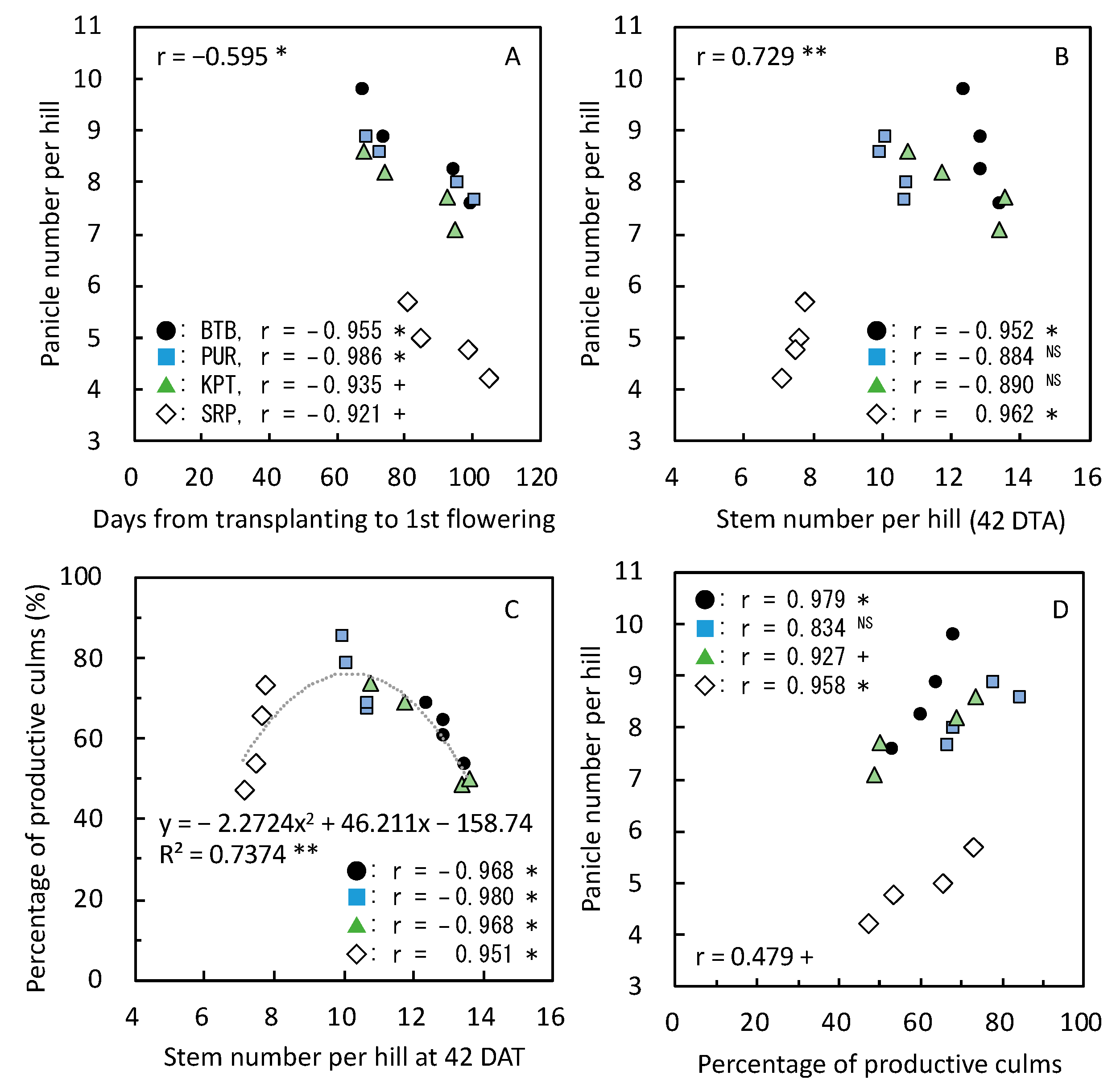
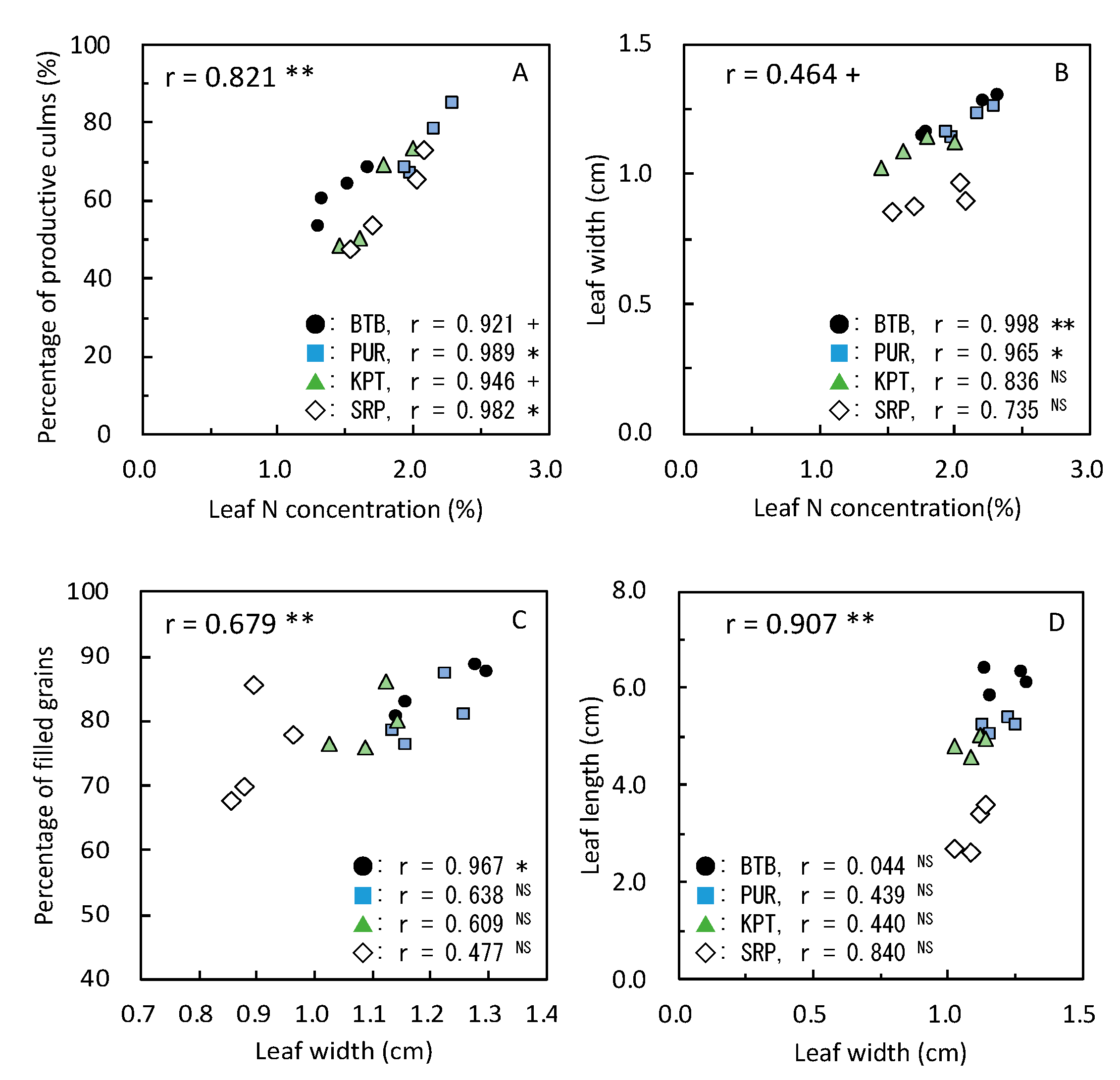
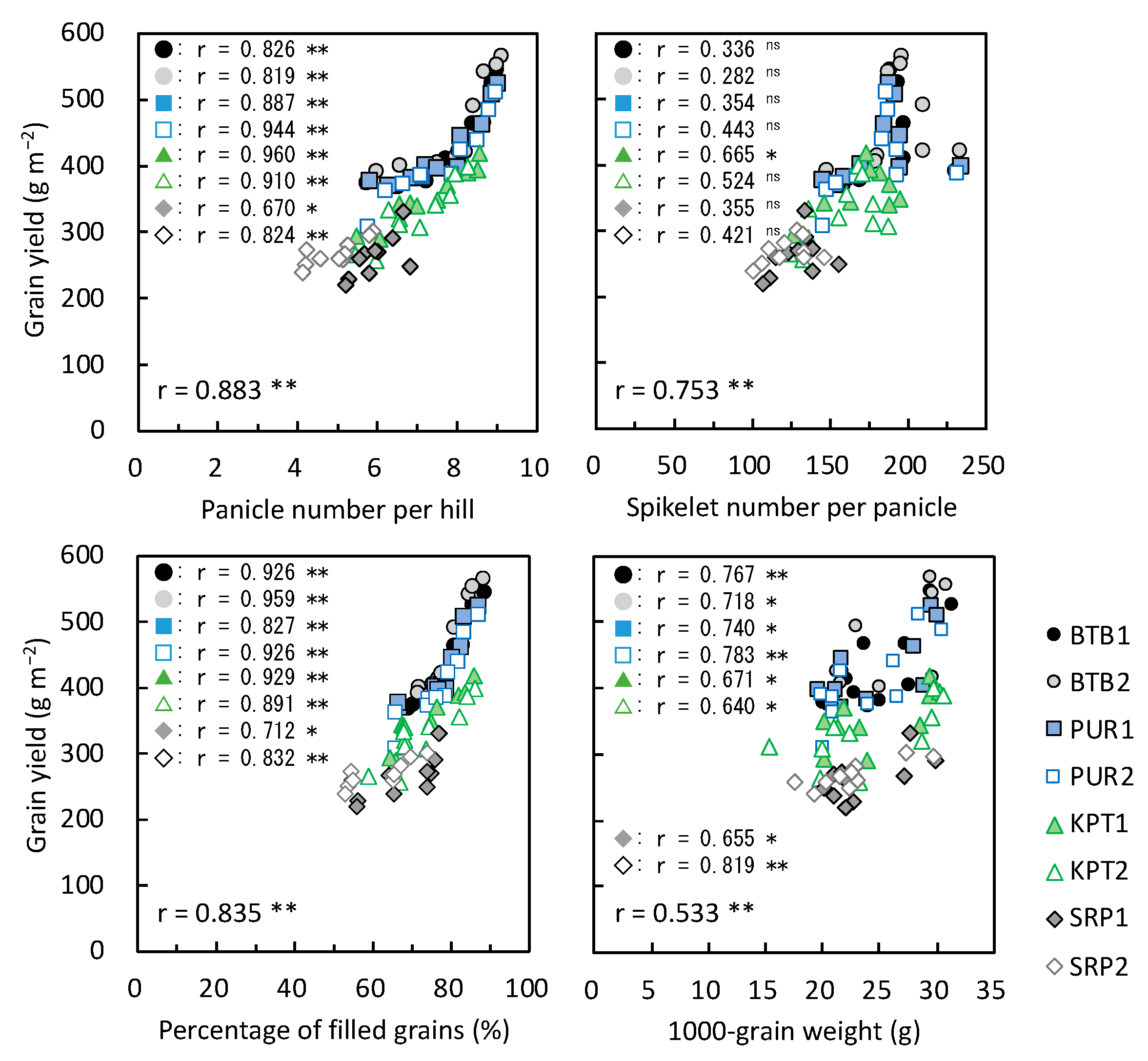
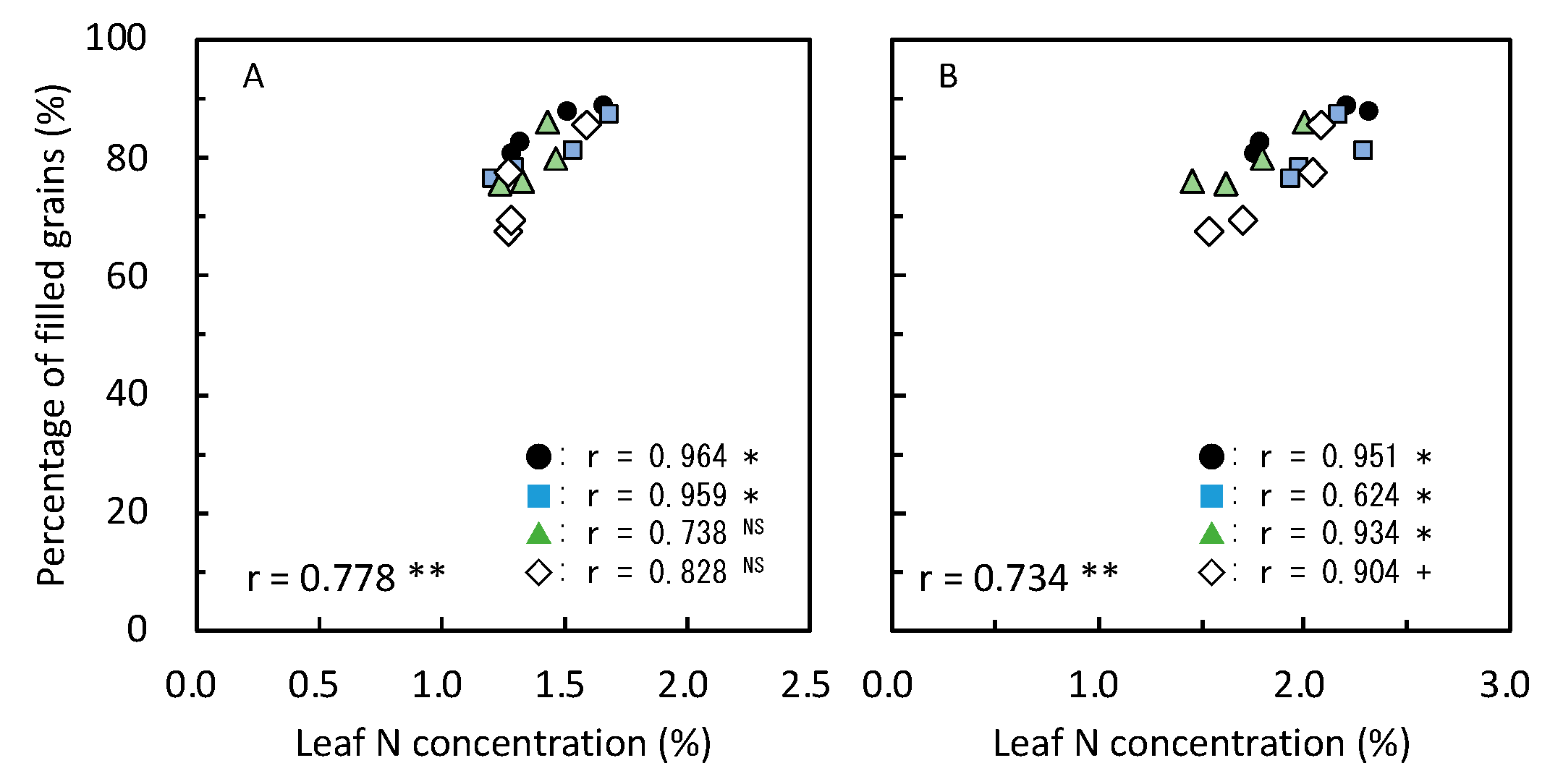
3.4. Yield-Determining Factors in 2019
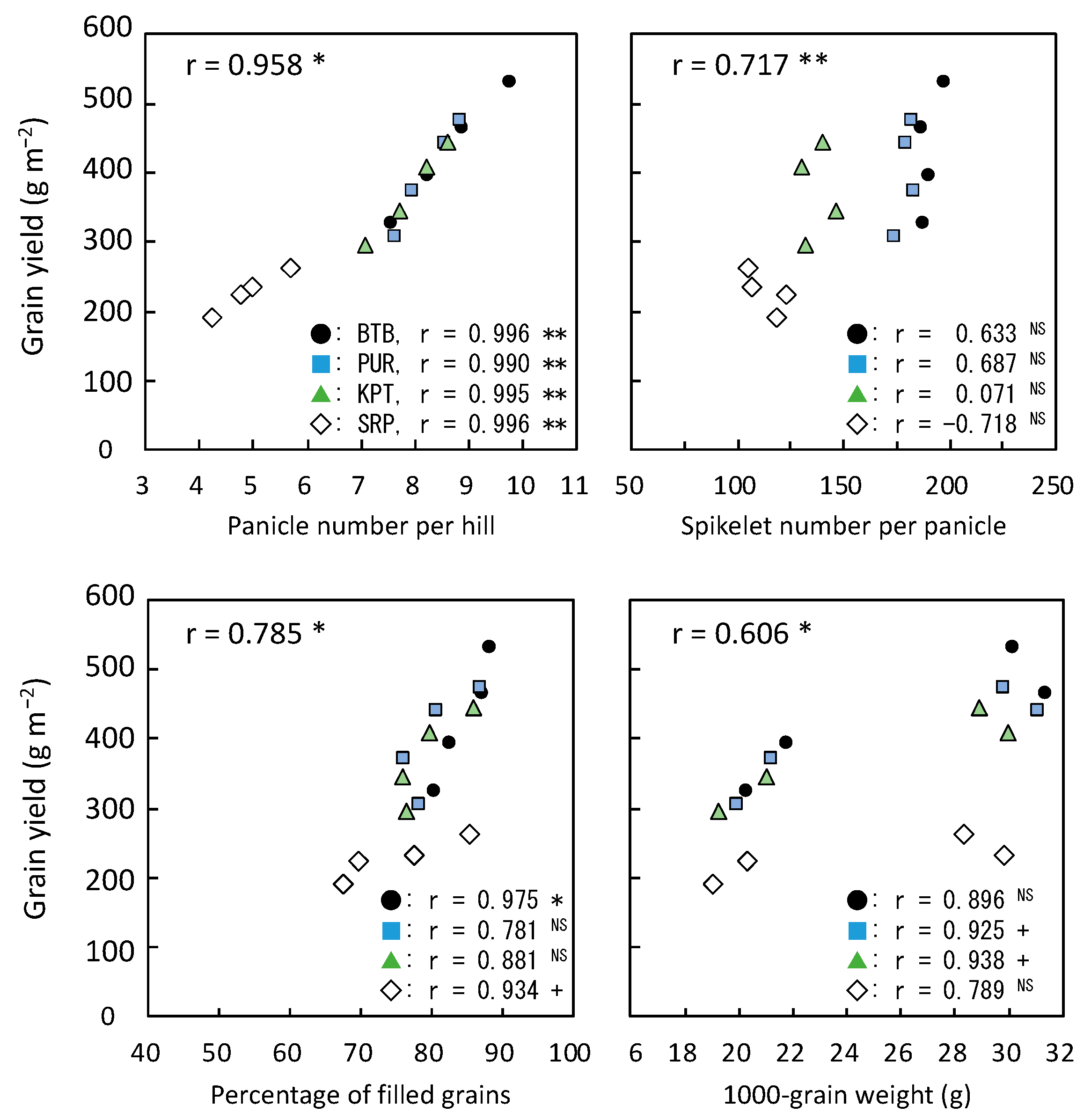
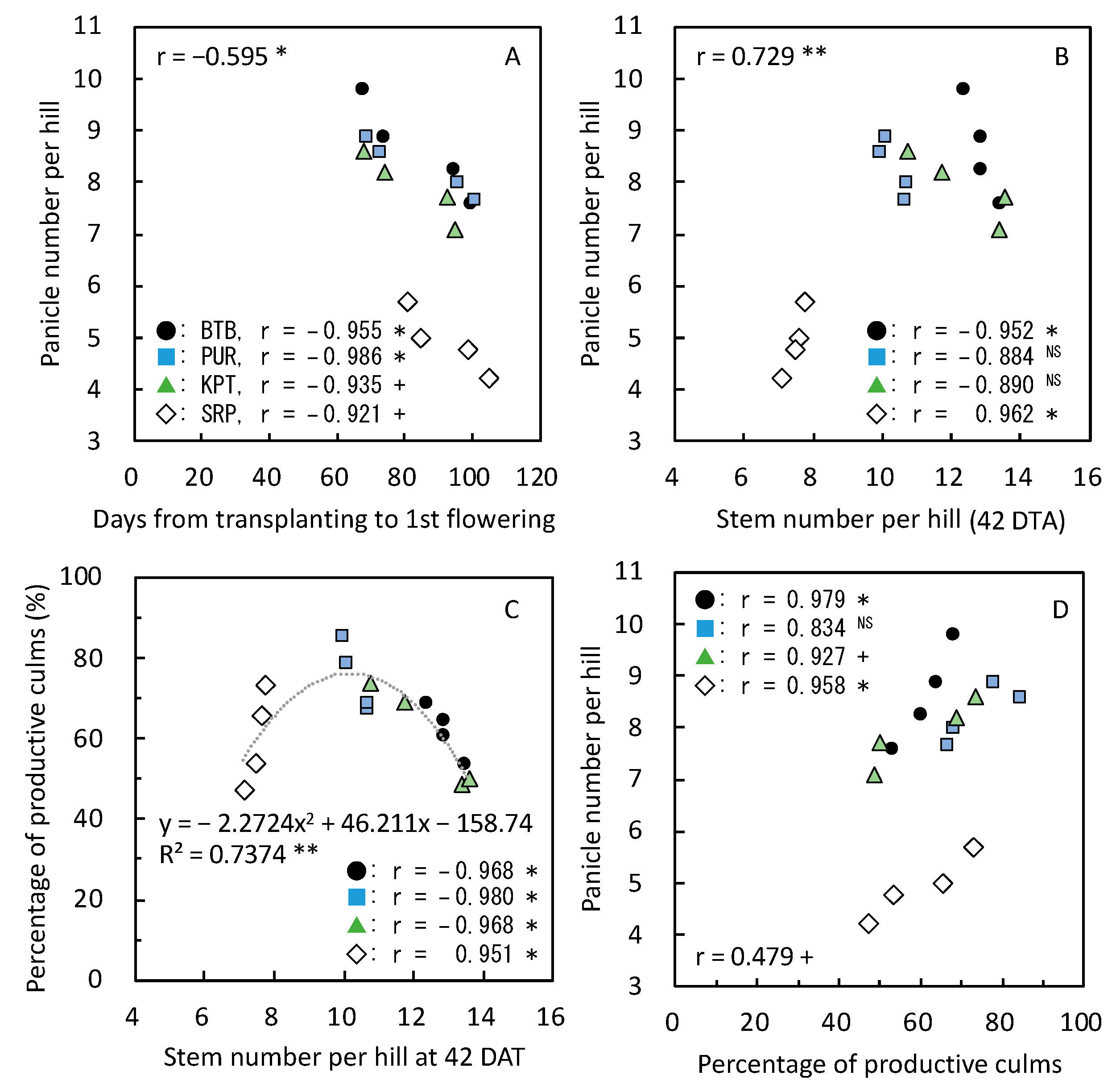
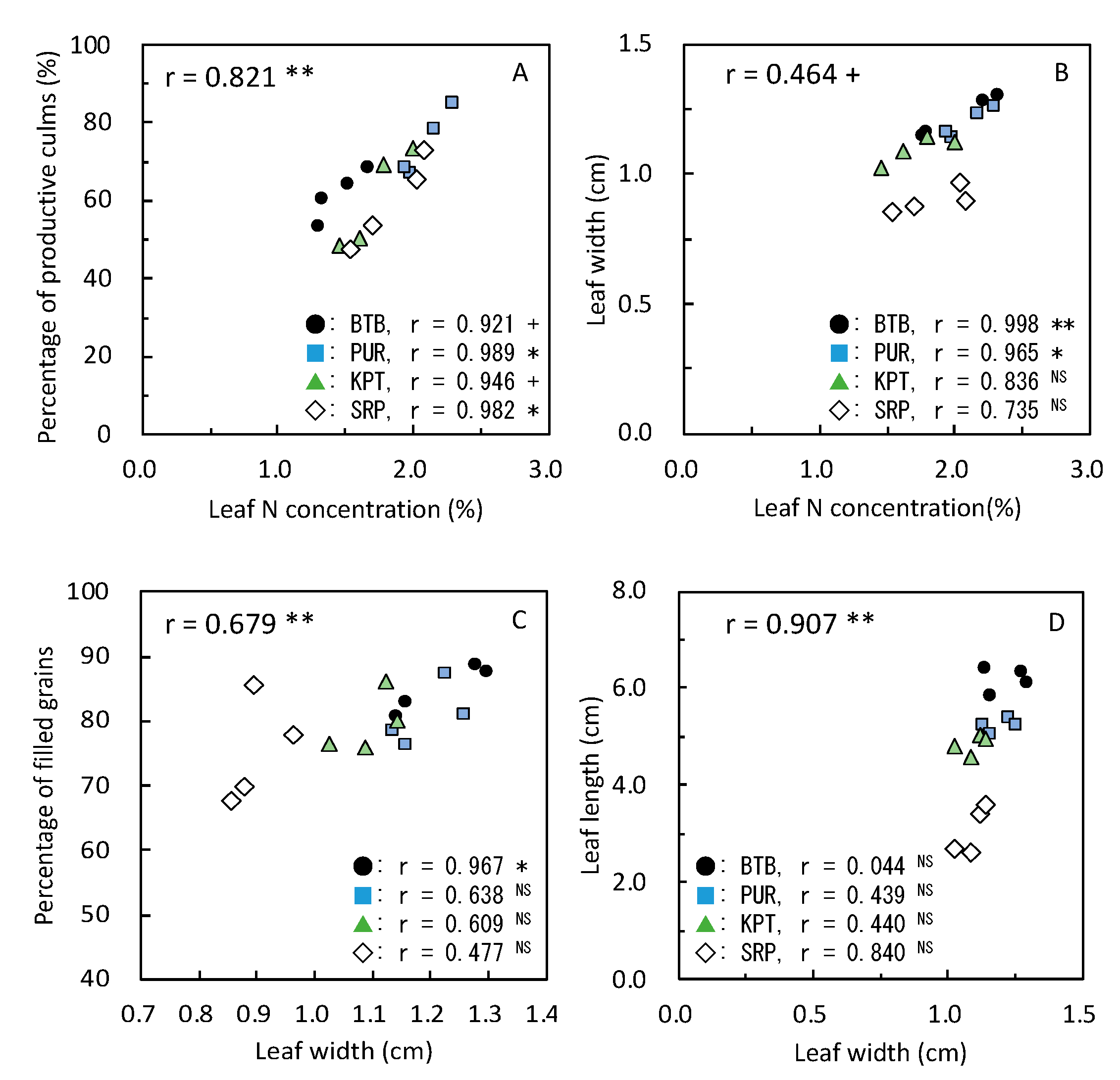
3.5. Yield and Its Components with Relating Parameters in 2020
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oladosu, Y.; Rafii, M.; Abdullah, N.; Hussin, G.; Ramli, A.; Rahim, H.A.; Miah, G.; Usman, M. Priciple and application of plant mutagenesis in crop improvement: A review. Biotechnol. Biotechnol. Equip. 2016, 30, 1–16. [Google Scholar]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; et al. Rational design of high-yield and superior-quality rice. Nat. Plants 2017, 3, 17031. [Google Scholar] [CrossRef]
- Gruneberge, W.J.; Manrique, K.; Zhang, D.; Hermann, M. Genotype x environment interactions for a diverse set of sweet potato clones evaluated across varying ecogeographic conditions in Peru. Crop Sci. 2005, 45, 2160–2171. [Google Scholar] [CrossRef]
- Horn, L.; Shimelis, H.; Sarsu, F.; Mwadzingeni, L.; Laing, M.D. Genotype-by-environment interaction for grain yield among novel cowpea (Vigna unguiculata L.) selections derived by gamma irradiation. Crop J. 2018, 6, 306–313. [Google Scholar] [CrossRef]
- Chandra, B.S.; Reddy, T.D.; Ansari, N.A.; Kumar, S.S. Correlation and path analysis for yield and yield components in rice (Oryza sativa L.). Agric. Sci. Dig. 2019, 29, 45–47. [Google Scholar]
- Chung, S.O.; Sudduth, K.A.; Chang, Y.C. Path analysis of factors limiting crop yield in rice paddy and upland corn fields. J. Biosyst. Eng. 2005, 39, 377–388. [Google Scholar]
- Huang, M.; Zou, Y.B.; Jiang, P.; Xia, B.; Md, I.; Ao, H.J. Relationship between grain yield and yield components in super hybrid rice. Agric. Sci. China 2011, 10, 1537–1544. [Google Scholar]
- Li, R.; Li, M.; Liu, S.; Zhang, J. Exploring the relationships between yield and yield-related traits for rice varieties released in China from 1978 to 2017. Front. Plant Sci. 2019, 10, 543. [Google Scholar]
- Yang, W.; Peng, S.; Laza, R.C.; Visperas, R.M. Yield gap analysis between dry and wet season rice crop grown under high-yielding management conditions. Agron. J. 2008, 100, 1390–1395. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.S. Quantitative genetics, genomics and plant breeding. Ann. Bot. 2004, 93, 225. [Google Scholar] [CrossRef] [Green Version]
- Horn, L.; Shimelis, H.; Sarsu, F.; Mwadzingeni, L.; Laing, M.D. Genotype by environment (G × E) interaction study on yield traits in different maturity group of rice. J. Crop Sci. Biotechnol. 2019, 22, 425–449. [Google Scholar]
- Yan, W. GGE biplot- A windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical analysis of yield trail. Agron. J. 1988, 80, 388–393. [Google Scholar]
- Nesbitt, H.J. Rice Production in Cambodia; International Rice Research Institute: Manila, Philippines, 1997; pp. 1–112. Available online: http://books.irri.org/9712201007_content.pdf (accessed on 22 May 2023).
- Jones, M.; Leng, V.; Ong, S.; Mean, M. Gender Technology Assessment: Appropriate Scale Mechanization Consortium. Feed the Future. Available online: https://agrilinks.org/sites/default/files/resources/jones_leng_ong_mean_2019_ca_service_provision_tech_assessment_final.pdf (accessed on 22 May 2023).
- Ministry of Agriculture, Forestry and Fisheries. Annual Conference Report on Agriculture Forestry and Fisheries; Ministry of Agriculture, Forestry and Fisheries: Phnom Penh, Cambodia, 2019; pp. 1–137. [Google Scholar]
- Fukai, S.; Ouk, M. Increased productivity of rainfed lowland rice cropping systems of the Mekong region. Crop Pasture Sci. 2012, 63, 944–973. [Google Scholar] [CrossRef]
- Kong, K.; Hin, S.; Seng, V.; Ismail, A.M.; Vergara, G.; Choi, I.R.; Ehara, H.; Kato, Y. Importance of phosphorus and potassium in soil-specific nutrient management for wet-season rice in Cambodia. Exp. Agric. 2019, 56, 204–217. [Google Scholar] [CrossRef]
- Kong, K.; Hin, S.; Seng, V.; Ismail, A.M.; Vergara, G.; Choi, I.-R.; Ehara, H.; Kato, Y. Potential yield and nutrient requirements of direct-seeded, dry-season rice in Cambodia. Exp. Agric. 2019, 56, 255–264. [Google Scholar] [CrossRef]
- Kato, Y.; Katsura, K. Panicle architecture and grain number in irrigated rice, grown under different water management regimes. Field Crops Res. 2010, 117, 237–244. [Google Scholar] [CrossRef]
- Seng, V.; Ros, C.; Bell, R.W.; White, P.F.; Hin, S. Nutrient requirements of rainfed lowland rice in Cambodia. In Increased Lowland Rice Production in the Mekong Region; Fukai, S., Basnayake, J., Eds.; ACIAR Proceedings No. 101; Australian Centre for International Agricultural Research: Canberra, Australia, 2001; pp. 170–178. [Google Scholar]
- Dou, F.; Soriano, J.; Tabien, R.E.; Chen, K. Soil Texture and Cultivar Effects on Rice (Oryza sativa L.) Grain Yield, Yield Components and Water Productivity in Three Water Regimes. PLoS ONE 2016, 11, e0150549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, M.; Li, X.; Dai, Q.; Xu, K.; Guo, B.; Hu, Y.; Wang, W.; Huo, Z. Effects of Soil Types and Irrigation Modes on Rice Root Morphophysiological Traits and Grain Quality. Agronomy 2021, 11, 20. [Google Scholar] [CrossRef]
- Blair, G.; Blair, N. Nutrient Status of Cambodian Soils, Rationalisation of Fertiliser Recommendations and the Challenges Ahead for Cambodian Soil Science. Curr. Agric. Res. J. 2014, 2, 5–13. [Google Scholar]
- Becker, H.C.; Leon, J. Stability analysis in plant breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Dobermann, A.; Fairhurst, T.H. Rice: Nutrient Disorders and Nutrient Management; Potash and Phosphate Institute, International Rice Research Institute: Los Baños, Philippines, 2000; pp. 1–254. [Google Scholar]
- Sumi, A.; Sugata, S.; Yahiro, I.; Odawara, M. Effect of fertilizer and fixed nitrogen on the water use efficiency of genge (Astragalus sinicus L.). Plant Prod. Sci. 2015, 18, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Skyer, J.K.; Wiliams, J.D.H.; Walker, T.W. The determination of total phosphorus in soils and parent materials. New Z. J. Agric. Res. 1968, 11, 757–762. [Google Scholar]
- Schollenberger, C.J.; Dreibelbis, E.R. Analytical methods in base-exchange investigations on soils. Soil Sci. 1930, 30, 161–173. [Google Scholar]
- Homma, K.; Horie, T.; Shiraiwa, T.; Supapoj, N.; Matsumoto, N.; Kabaki, N. Toposequential variation in soil fertility and rice productivity of rainfed lowland paddy fields in mini-watershed (Nong) in Northeast Thailand. Plant Prod. Sci. 2003, 6, 147–153. [Google Scholar] [CrossRef]
- Sharma, S.; Panneerselvam, P.; Castillo, R.; Manohar, S.; Raj, R.; Ravi, V.; Buresh, R.J. Web-based tool for calculating field-specific nutrient management for rice in India. Nutr. Cycl. Agroecosystems 2019, 113, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.; Dobermann, A.; Oberthur, T.; Ros, S. The rice soils of Cambodia, I. Soil classification for agronomists using the Cambodian Agronomic Soil Classification system. Soil Use Manag. 2000, 16, 12–19. [Google Scholar] [CrossRef]
- Tsubo, M.; Fukai, S.; Basnayake, J.; Tuong, T.P.; Bouman, B.; Harnpichitvitaya, D. Effects of soil clay content on water balance and productivity in rainfed lowland rice ecosystem in Northeast Thailand. Plant Prod. Sci. 2007, 10, 232–241. [Google Scholar]
- FAOSTAT. Electronic Database; Food and Agricultural Organization: Rome, Italy, 2020; Available online: http://www.fao.org/faostat/ (accessed on 30 June 2023).
- Pheakdey, D.V.; Xuan, T.D.; Khanh, T.D. Influence of climate factors on rice yields in Cambodia. Geosciences 2017, 3, 561–575. [Google Scholar]
- Bishwas, K.C.; Poudel, M.R.; Regmi, D. AMMI and GGE biplot analysis of yield of different elite wheat line under terminal heat stress and irrigated environment. Heliyon 2021, 7, e07206. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S. GGE Biplot Analysis, A Graphical Tool for Breeders, Genetics, and Agronomist; CRC Press: Washington, DC, USA, 2003; pp. 1–225. [Google Scholar]
- Zewdu, Z.; Dessie, A.; Worede, F.; Atinaf, M.; Berie, A.; Tahir, Z.; Kinfe, H.; Bitew, M. Agronomic performance evaluation and yield stability analysis of upland rice (Oryza sativa L.) varieties using AMMI and GGE biplot. Plant 2020, 8, 87–92. [Google Scholar]
- Gauch, H.G.; Zobel, R.W. Identifying mega-environments and targeting genotypes. Crop Sci. 1997, 37, 311–326. [Google Scholar] [CrossRef]
- Khema, S.; Rin, S.; Fujita, A.; Kong, K.; Ngin, C.; Kano-Nakata, M.; Yamauchi, A.; Ehara, H. Grain yield and gross return above fertilizer cost with parameters relating to the quality of white rice cultivated in rainfed paddy fields in Cambodia. Sustainability 2022, 14, 10708. [Google Scholar] [CrossRef]
- Banayo, N.P.M.C.; Haefele, S.M.; Desamero, N.V.; Kato, Y. Site-specific nutrient management enhances sink size, a major yield constraint in rainfed lowland rice. Field Crops Res. 2018, 224, 76–79. [Google Scholar] [CrossRef]
- Ehara, H.; Komada, C.; Goto, M.; Morita, O. Relationship between nitrogen content in leaf dry weight basis and absorption capacity of ammonium and nitrate of rice seedlings grown under low nutrient concentration. Jap. J. Crop Sci. 1997, 66, 329–330. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Okajima, H.; Takashiro, S.; Honda, T. Tiller emergence mechanizm in rice plant: I. Tiller emergence of rice seedlings cultivated under nitrogen deficient condition. Bull. Inst. Agric. Res. Tohoku Univ. 1956, 8, 91–117. [Google Scholar]
- Oshima, M. Study on nitrogen nutrition in rice plant: IV. The effect of nitrogen nutrition on tillering. Jpn. Soc. Soil Sci. Plan Nutr. 1962, 33, 243–246. [Google Scholar]
- Ehara, H.; Tushiya, M.; Naito, H.; Ogo, T. Effect of the high nitrogen treatment priot to transplanting on the growth and yield in rice. Jpn. J. Crop Sci. 1992, 61, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ehara, H.; Tushiya, M.; Hirai, Y.; Ogo, T. Varietal difference on morphological change of leaf blade by different nutrient concentration of culture solution in rice seedlings. Jpn. J. Crop Sci. 1992, 61, 10–15. [Google Scholar]

 ≧ 450, 450 >
≧ 450, 450 >  ≧ 400, 400 >
≧ 400, 400 >  ≧ 350, 350 >
≧ 350, 350 >  ≧ 300, and 300 >
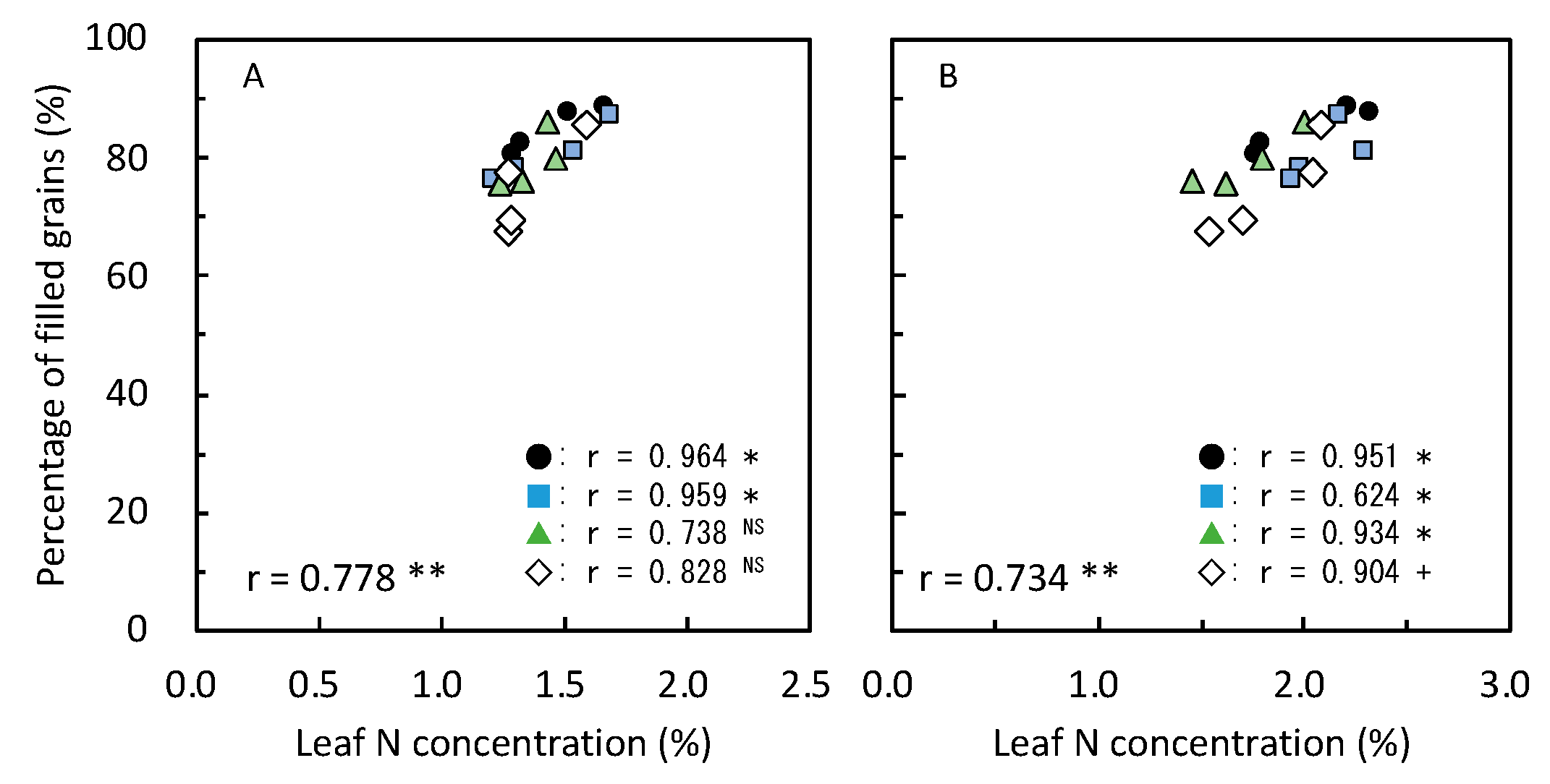
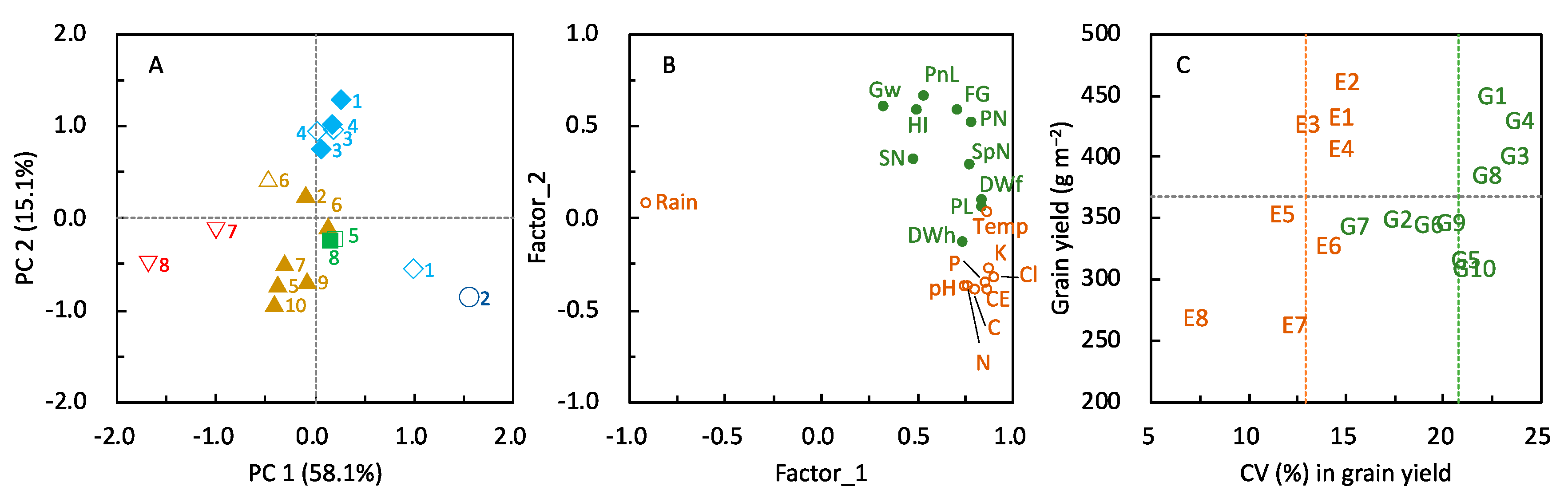
≧ 300, and 300 >  ≧ 250. SN: maximum stem number per hill; PL: plant length at harvesting; DWh: shoot dry matter weight at heading; DWf: shoot dry matter weight at harvesting; PnL: panicle length; PN: panicle number per hill; SpN: spikelet number per panicle; FG: percentage of filled grains; GW: 1000-grain weight; HI: harvest index; Cl: clay content; pH: pH(HCl); C: total C; N: total N; P: total P; CE: CEC; K: exchangeable K; Temp: average daily mean air temperature; Rain: accumulated precipitation during growth period of each variety.
≧ 450, 450 > ≧ 400, 400 > ≧ 350, 350 > ≧ 300, and 300 > ≧ 250. SN: maximum stem number per hill; PL: plant length at harvesting; DWh: shoot dry matter weight at heading; DWf: shoot dry matter weight at harvesting; PnL: panicle length; PN: panicle number per hill; SpN: spikelet number per panicle; FG: percentage of filled grains; GW: 1000-grain weight; HI: harvest index; Cl: clay content; pH: pH(HCl); C: total C; N: total N; P: total P; CE: CEC; K: exchangeable K; Temp: average daily mean air temperature; Rain: accumulated precipitation during growth period of each variety.
≧ 250. SN: maximum stem number per hill; PL: plant length at harvesting; DWh: shoot dry matter weight at heading; DWf: shoot dry matter weight at harvesting; PnL: panicle length; PN: panicle number per hill; SpN: spikelet number per panicle; FG: percentage of filled grains; GW: 1000-grain weight; HI: harvest index; Cl: clay content; pH: pH(HCl); C: total C; N: total N; P: total P; CE: CEC; K: exchangeable K; Temp: average daily mean air temperature; Rain: accumulated precipitation during growth period of each variety.
≧ 450, 450 > ≧ 400, 400 > ≧ 350, 350 > ≧ 300, and 300 > ≧ 250. SN: maximum stem number per hill; PL: plant length at harvesting; DWh: shoot dry matter weight at heading; DWf: shoot dry matter weight at harvesting; PnL: panicle length; PN: panicle number per hill; SpN: spikelet number per panicle; FG: percentage of filled grains; GW: 1000-grain weight; HI: harvest index; Cl: clay content; pH: pH(HCl); C: total C; N: total N; P: total P; CE: CEC; K: exchangeable K; Temp: average daily mean air temperature; Rain: accumulated precipitation during growth period of each variety.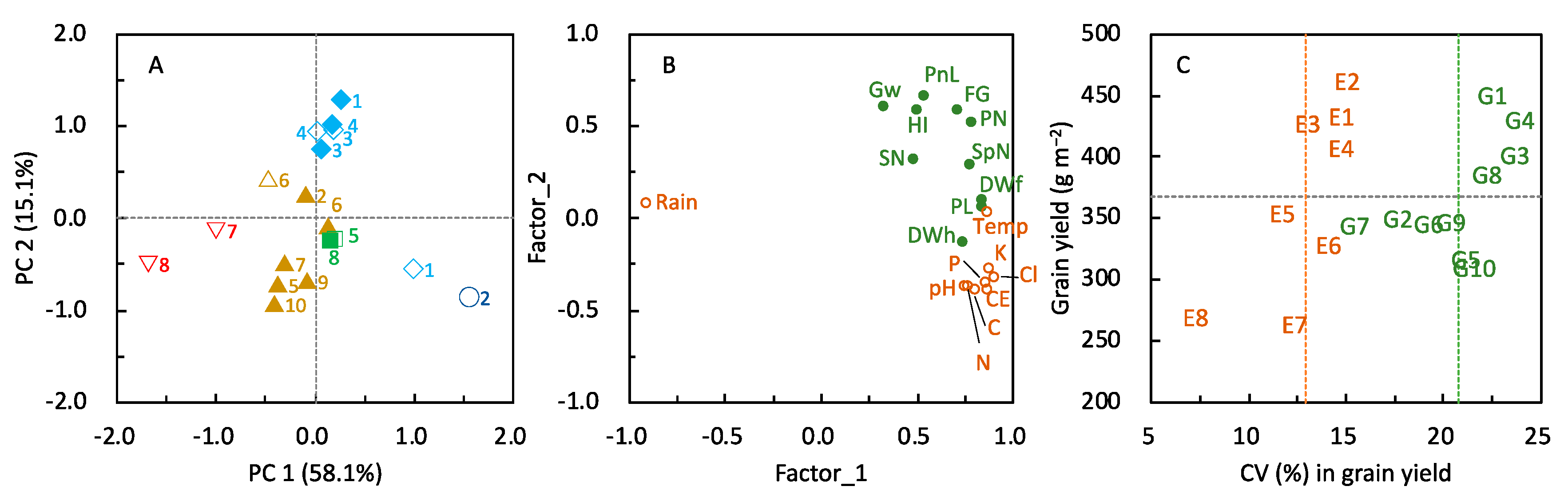



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location Environment | Province | Village | Latitude, Longitude | Soil Classification |
|---|---|---|---|---|
| E1 (BTB1) | Battambang | Thmei | 12°55.039′ N, 103°8.094′ E | Tuol Samroung |
| E2 (BTB2) | Battambang | Chong Samnay | 12°47.324′ N, 103°27.400′ E | Tuol Samroung |
| E3 (PUR1) | Pursat | Sen Pen | 12°29.467′ N, 104°2.771′ E | Krakor |
| E4 (PUR2) | Pursat | Tbaeng Chum | 12°30.025′ N, 104°2.743′ E | Krakor |
| E5 (KPT1) | Kampong Thom | Sakream Cheung | 13°11.379′ N, 104°45.690′ E | Prateah Lang |
| E6 (KPT2) | Kampong Thom | Boeng Lvea | 12°34.229′ N, 105°8.665′ E | Prateah Lang |
| E7 (SRP1) | Siem Reap | Teuk Thla | 13°23.766′ N, 103°47.594′ E | Prey Khmer |
| E8 (SRP2) | Siem Reap | Prey Thmei | 13°24.344′ N, 103°47.511′ E | Prey Khmer |
| Parameter | Soil Depth | E1 BTB1 | E2 BTB2 | E3 PUR1 | E4 PUR2 | E5 KPT1 | E6 KPT2 | E7 SRP1 | E8 SRP2 |
|---|---|---|---|---|---|---|---|---|---|
| pH (HCl) | 0–20 | 6.08 | 6.79 | 5.52 | 5.10 | 6.59 | 4.97 | 5.50 | 4.92 |
| 20–40 | 6.82 | 5.23 | 5.81 | 5.12 | 6.01 | 4.96 | 5.53 | 5.41 | |
| Total C (%) | 0–20 | 1.65 | 1.98 | 1.20 | 1.00 | 1.56 | 1.30 | 1.34 | 0.90 |
| 20–40 | 1.56 | 2.10 | 1.23 | 1.34 | 0.80 | 1.00 | 0.93 | 1.07 | |
| Total N (%) | 0–20 | 0.17 | 0.21 | 0.10 | 0.11 | 0.14 | 0.14 | 0.14 | 0.08 |
| 20–40 | 0.15 | 0.21 | 0.12 | 0.14 | 0.07 | 0.10 | 0.10 | 0.11 | |
| Total P (%) | 0–20 | 0.06 | 0.07 | 0.04 | 0.03 | 0.05 | 0.03 | 0.02 | 0.03 |
| 20–40 | 0.04 | 0.08 | 0.03 | 0.03 | 0.04 | 0.02 | 0.02 | 0.02 | |
| CEC (cmolc kg−1) | 0–20 | 21.2 | 24.5 | 10.0 | 10.0 | 18.8 | 8.0 | 8.5 | 6.5 |
| 20–40 | 23.5 | 20.5 | 7.5 | 8.2 | 12.5 | 7.0 | 7.0 | 8.5 | |
| Exchangeable K (cmolc kg−1) | 0–20 | 1.30 | 1.60 | 0.72 | 0.55 | 0.45 | 0.11 | 0.15 | 0.22 |
| 20–40 | 0.88 | 2.56 | 0.36 | 0.46 | 0.26 | 0.10 | 0.08 | 0.10 | |
| Clay (%) | 0–20 | 40.1 | 50.8 | 13.1 | 15.7 | 17.3 | 14.9 | 7.5 | 2.3 |
| 20–40 | 44.6 | 49.6 | 13.6 | 13.2 | 14.7 | 9.7 | 7.0 | 1.8 | |
| Silt (%) | 0–20 | 29.8 | 17.3 | 27.4 | 34.2 | 15.7 | 11.5 | 1.3 | 2.2 |
| 20–40 | 23.7 | 19.2 | 27.1 | 32.1 | 30.3 | 13.0 | 6.0 | 6.6 | |
| Sand (%) | 0–20 | 13.1 | 17.6 | 19.3 | 19.7 | 33.6 | 40.0 | 25.3 | 39.7 |
| 20–40 | 15.7 | 14.1 | 23.5 | 22.5 | 30.0 | 41.8 | 24.6 | 34.6 |
| Factor | Panicle Number Per Hill | Spikelet Number Per Panicle | Percentage of Filled Grains | 1000-Grain Weight (g) | Grain Yield (g m−2) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variety/genotype | ||||||||||
| G1 Phka Rumduol | 8.17 | a | 169.6 | d | 84.1 | a | 28.9 | b | 449.8 | a |
| G2 Phka Romeat | 6.67 | g | 155.5 | g | 69.5 | g | 27.5 | c | 349.7 | e |
| G3 Phka Rumdeng | 7.71 | c | 166.1 | f | 79.6 | c | 26.8 | c | 401.1 | c |
| G4 Phka Mealdei | 8.00 | b | 172.1 | c | 81.2 | b | 30.5 | a | 430.4 | b |
| G5 Phka Chan Sen Sar | 5.86 | i | 138.9 | i | 64.4 | i | 23.0 | de | 316.4 | f |
| G6 Raing Chey | 7.33 | e | 201.2 | a | 75.1 | e | 20.6 | f | 345.8 | e |
| G7 CAR3 | 6.33 | h | 145.6 | h | 67.9 | h | 23.1 | d | 343.7 | e |
| G8 CAR4 | 7.39 | d | 178.4 | b | 76.3 | d | 22.1 | e | 384.9 | d |
| G9 CAR6 | 6.82 | f | 168.5 | e | 70.4 | f | 20.2 | f | 345.8 | e |
| G10 CAR11 | 5.42 | j | 130.0 | j | 63.2 | j | 20.5 | f | 309.1 | f |
| Location/environment | ||||||||||
| E1 (BTB1) | 7.76 | b | 182.0 | b | 78.1 | b | 25.4 | ab | 433.2 | b |
| E2 (BTB2) | 7.92 | a | 187.9 | a | 78.8 | a | 25.5 | a | 461.6 | a |
| E3 (PUR1) | 7.59 | c | 180.0 | c | 77.1 | c | 23.6 | bc | 427.2 | b |
| E4 (PUR2) | 7.48 | d | 177.5 | d | 76.4 | d | 24.0 | cd | 406.7 | c |
| E5 (KPT1) | 7.24 | e | 166.6 | e | 73.6 | e | 23.6 | abc | 353.2 | d |
| E6 (KPT2) | 6.92 | f | 157.3 | f | 73.0 | f | 24.0 | cd | 327.9 | e |
| E7 (SRP1) | 5.92 | g | 127.7 | g | 67.1 | g | 23.6 | d | 262.8 | f |
| E8 (SRP2) | 4.93 | h | 121.8 | h | 61.2 | h | 22.6 | e | 268.8 | f |
| F value, Probability | ||||||||||
| Genotype (G) | 7428.6 | <0.001 | 7428.6 | <0.001 | 3281.4 | <0.001 | 286.1 | <0.001 | 417.6 | <0.001 |
| Environment (E) | 13,717.4 | <0.001 | 13,717.4 | <0.001 | 2980.2 | <0.001 | 22.8 | <0.001 | 1367.2 | <0.001 |
| G × E | 89.8 | <0.001 | 89.8 | <0.001 | 39.4 | <0.001 | 5.2 | <0.001 | 10.5 | <0.001 |
| Genotype | E1 (BTB1) | E2 (BTB2) | E3 (PUR1) | E4 (PUR2) | E5 (KPT1) | E6 (KPT2) | E7 (SRP1) | E8 (SRP2) | CV (%) |
|---|---|---|---|---|---|---|---|---|---|
| 1. Phka Rumduol | 546.1 a B | 565.9 a A | 525.0 a C | 511.5 a C | 418.8 a D | 399.0 a E | 331.0 a F | 301.3 a G | 22.5 |
| 2. Phka Romeat | 403.5 cd A | 415.0 de A | 402.0 c A | 384.3 d B | 344.5 d C | 320.8 bc D | 267.5 bcd E | 260.0 de E | 17.7 |
| 3. Phka Rumdeng | 465.8 b B | 542.8 b A | 463.3 b B | 440.0 c B | 389.0 b C | 355.8 ab D | 270.5 bc E | 281.3 bc E | 23.7 |
| 4. Phka Mealdei | 526.0 a AB | 554.0 ab A | 509.3 a BC | 485.5 b C | 393.8 b D | 387.5 a D | 291.3 ab E | 295.8 ab E | 24.0 |
| 5. Phka Chan Sen Sar | 369.5 e B | 401.5 ef A | 370.8 d AB | 363.0 e B | 290.0 f C | 257.3 e D | 228.5 cd D | 250.5 ef D | 21.2 |
| 6. Raing Chey | 391.5 cde AB | 422.8 d A | 398.3 cd A | 388.8 d AB | 349.0 cd BC | 307.3 cd C | 248.8 bcd D | 259.3 de D | 19.3 |
| 7. CAR3 | 378.5 de B | 406.3 def A | 383.0 cd B | 373.8 de B | 342.0 d C | 333.5 bc C | 260.0 bcd D | 272.8 cd D | 15.5 |
| 8. CAR4 | 464.3 b AB | 491.8 c A | 445.8 b BC | 424.8 c C | 371.3 bc E | 341.5 bc E | 272.5 bc E | 268.0 cde E | 22.3 |
| 9. CAR6 | 411.3 c AB | 423.3 d A | 397.5 cd AB | 386.3 d B | 339.8 d C | 311.0 bc C | 238.3 cd D | 259.5 de D | 20.4 |
| 10. CAR11 | 375.5 de A | 393.0 f A | 377.5 cd A | 309.0 f B | 294.0 f BC | 265.5 de CD | 219.5 d E | 239.0 f DE | 21.6 |
| CV (%) | 14.8 | 15.1 | 13.0 | 14.8 | 11.7 | 14.1 | 12.4 | 7.3 |
| E1 (BTB1) | E2 (BTB2 | E3 (PUR1) | E4 (PUR2) | E5 (KPT1) | E6 (KPT2) | E7 (SRP1) | E8 (SRP2) | |
|---|---|---|---|---|---|---|---|---|
| E1 (BTB1) | − | 0.952 ** | 0.964 ** | 0.939 ** | 0.939 ** | 0.855 ** | 0.976 ** | 0.915 ** |
| E2 (BTB2) | 0.952 ** | − | 0.952 ** | 0.988 ** | 0.891 ** | 0.830 ** | 0.952 ** | 0.867 ** |
| E3 (PUR1) | 0.964 ** | 0.952 ** | − | 0.976 ** | 0.891 ** | 0.855 ** | 0.915 ** | 0.915 ** |
| E4 (PUR2) | 0.939 ** | 0.988 ** | 0.976 ** | − | 0.879 ** | 0.842 ** | 0.915 ** | 0.879 ** |
| E5 (KPT1) | 0.939 ** | 0.891 ** | 0.891 ** | 0.879 ** | − | 0.927 ** | 0.939 ** | 0.952 ** |
| E6 (KPT2) | 0.855 ** | 0.830 ** | 0.855 ** | 0.842 ** | 0.927 ** | − | 0.867 ** | 0.976 ** |
| E7 (SRP1) | 0.976 ** | 0.952 ** | 0.915 ** | 0.915 ** | 0.939 ** | 0.867 ** | − | 0.915 ** |
| E8 (SRP2) | 0.915 ** | 0.867 ** | 0.915 ** | 0.879 ** | 0.952 ** | 0.976 ** | 0.915 ** | − |
| Province | Applied Variable | Adjusted R Square | F = | p = | Unstandard Coefficient | Standardized | |
|---|---|---|---|---|---|---|---|
| Constant | Variable | ||||||
| E1 (BTB1) | Percentage of filled grains | 0.861 | 56.546 | <0.001 | −306.949 | 9.478 | 0.936 |
| E2 (BTB2) | Percentage of filled grains | 0.908 | 90.103 | <0.001 | −460.519 | 11.704 | 0.958 |
| E3 (PUR1) | Panicle number per hill | 0.755 | 28.744 | <0.001 | 85.805 | 44.927 | 0.885 |
| E4 (PUR2) | Panicle number per hill | 0.877 | 64.993 | <0.001 | 26.297 | 50.856 | 0.944 |
| E5 (KPT1) | Panicle number per hill | 0.907 | 88.623 | <0.001 | 74.089 | 38.501 | 0.958 |
| E6 (KPT2) | Panicle number per hill | 0.801 | 37.119 | <0.001 | 14.141 | 45.278 | 0.907 |
| E7 (SRP1) | Percentage of filled grains | 0.447 | 8.265 | 0.021 | 89.767 | 2.577 | 0.713 |
| E8 (SRP2) | Percentage of filled grains | 0.654 | 18.018 | 0.003 | 141.538 | 2.079 | 0.832 |
| Factor | d.f. | Sum of Squared Deviation | Unbiased Variance | F = | p = |
|---|---|---|---|---|---|
| All | 63 | 60.532 | 0.961 | ||
| Genotype (G) | 3 | 19.996 | 6.665 | 1484.468 | >0.001 |
| Province (E) | 3 | 37.830 | 12.610 | 2808.403 | >0.001 |
| G × E | 9 | 2.490 | 0.277 | 61.624 | >0.001 |
| Error | 48 | 0.216 | 0.005 |
| Variety | BTB | PUR | KPT | SRP | CV (%) |
|---|---|---|---|---|---|
| Phka Rumduol | 531.0 a A | 473.0 a B | 445.0 a C | 263.0 a D | 27.0 |
| Phka Mealdei | 464.0 b A | 405.0 b B | 409.0 b C | 234.0 b D | 26.4 |
| Raing Chey | 325.0 d A | 305.0 d B | 296.0 d B | 192.0 c C | 21.3 |
| CAR 4 | 394.0 c A | 372.0 c B | 346.0 c C | 224.0 b D | 22.7 |
| CV (%) | 20.7 | 18.0 | 17.7 | 12.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uch, C.; Roeurn, S.; Ro, S.; Kano-Nakata, M.; Yamauchi, A.; Ehara, H. Growth Performance of Photoperiod-Sensitive Rice (Oryza sativa L.) Varieties in Different Soil Types under Rainfed Condition in Cambodia. Agriculture 2023, 13, 1602. https://doi.org/10.3390/agriculture13081602
Uch C, Roeurn S, Ro S, Kano-Nakata M, Yamauchi A, Ehara H. Growth Performance of Photoperiod-Sensitive Rice (Oryza sativa L.) Varieties in Different Soil Types under Rainfed Condition in Cambodia. Agriculture. 2023; 13(8):1602. https://doi.org/10.3390/agriculture13081602
Chicago/Turabian StyleUch, Chanthol, Siranet Roeurn, Sophoanrith Ro, Mana Kano-Nakata, Akira Yamauchi, and Hiroshi Ehara. 2023. "Growth Performance of Photoperiod-Sensitive Rice (Oryza sativa L.) Varieties in Different Soil Types under Rainfed Condition in Cambodia" Agriculture 13, no. 8: 1602. https://doi.org/10.3390/agriculture13081602