

Genetic Variability of Oil Palm in Mexico: An Assessment Based on Microsatellite Markers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Isolation
2.3. SSR and RAM Markers Analysis
2.4. Data Analysis
3. Results
3.1. Elaeis Guineensis Plant and DNA Samples
3.2. SSR and RAM Markers Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 4th ed.; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 1–50. [Google Scholar]
- Adam, H.; Collin, M.; Richaud, F.; Beulé, T.; Cros, D.; Omore, A.; Nodichao, L.; Nouy, B.; Tregear, J.W. Environmental regulation of sex determination in oil palm: Current knowledge and insights from other species. Ann. Bot. 2011, 108, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.G.; Martin, S.M. Palm oil. In The Cambridge World History of Food; Kiple, K.F., Ornelas, K.C., Eds.; Cambridge University Press: New York, NY, USA, 2000; Volume 1, pp. 397–411. [Google Scholar]
- Umaña, C.H. Desarrollo del cultivo de la palma de aceite en Centroamérica. Rev. Palmas 1998, 19, 266–272. [Google Scholar]
- Bastidas, S.; Peña, E.A.; Reyes, R. Genealogía del germoplasma de palma de aceite (Elaeis guineensis Jacq.) del proyecto de mejoramiento genético de Corpoica. Rev. Palmas 2003, 24, 21–29. [Google Scholar]
- Hardon, J.J. Interspecific hybrids in the genus Elaeis II. Vegetative growth and yield of F1 hybrids E. guineensis x E. oleifera. Euphytica 1969, 18, 380–388. [Google Scholar] [CrossRef]
- Cochard, B.; Adon, B.; Rekima, S.; Billotte, N.; de Chenon, R.D.; Koutou, A.; Nouy, B.; Omore, A.; Purba, A.R.; Glazsmann, J.C.; et al. Geographic and genetic structure of African oil palm diversity suggests new approaches to breeding. Tree Genet. Genomes 2009, 5, 493–504. [Google Scholar] [CrossRef]
- Sulaiman, O.; Salim, N.; Nordin, N.A.; Hashim, R.; Ibrahim, M.; Sato, M. The potential of oil palm trunk biomass as an alternative source for compressed wood. BioResources 2012, 7, 2688–2706. [Google Scholar] [CrossRef]
- Federación Mexicana de Palma de Aceite (FEMEXPALMA). Anuario Estadístico 2021; Villahermosa: Tabasco, Mexico, 2021; pp. 20–51. [Google Scholar]
- USDA. Available online: https://ipad.fas.usda.gov/cropexplorer/cropview/commodityView.aspx?cropid=4243000 (accessed on 4 April 2022).
- Mazariegos-Sánchez, A.; Águila-González, J.M.; Martínez-Chávez, J.; Arévalo-Lozano, O. La industria de la palma de aceite en Acapetahua, Chiapas: El caso de Propalma. Rev. Mex. Agronegocios 2014, 35, 1052–1064. [Google Scholar]
- Velázquez-Cágal, M.; Pérez-Castañeda, G.; Corona-Sáez, T. Oil palm (Elaeis guineensis Jacq.) plantations diagnosis in the South of the Veracruz State, Mexico. In Proceedings of the 2001 PIPOC International Palm Oil Congress, Agriculture Conference, Kuala Lumpur, Malaysia, 20–22 August 2001. [Google Scholar]
- Ordoñez-Frías, E.J.; Azamar-Barrios, J.A.; Mata-Zayas, E.; Silván-Hernández, O.; Pampillón-González, L. Bioenergy potential and technical feasibility assessment of residues from oil palm processing: A case study of Jalapa, Tabasco, Mexico. Biomass Bioenergy 2020, 142, 105668. [Google Scholar] [CrossRef]
- John Martin, J.J.; Yarra, R.; Wei, L.; Cao, H. Oil Palm Breeding in the Modern Era: Challenges and Opportunities. Plants 2022, 11, 1395. [Google Scholar] [CrossRef]
- Okoye, M.; Uguru, M.; Bakoumé, C.; Singh, R.; Okwuagwu, C. Assessment of Genetic Diversity of NIFOR Oil Palm Main Breeding Parent Genotypes Using Microsatellite Markers. Am. J. Plant Sci. 2016, 7, 218–237. [Google Scholar] [CrossRef]
- Owolarafe, O.K.; Olabige, M.T.; Faborode, M.O. Physical and mechanical properties of two varieties of fresh oil palm fruit. J. Food Eng. 2007, 78, 1228–1232. [Google Scholar]
- Washid, M.B.; Abdullah, S.N.A.; Henson, I.E. Oil palm-achievements and potential. Plant Prod. Sci. 2005, 8, 288–297. [Google Scholar]
- Trujillo-Castillo, L.F.; Velázquez-Martínez, J.R.; López-Hernández, E.; Gómez-Vázquez, A. Palma de aceite africana. In Palma africana en Tabasco. Resultados de investigación; Velázquez, M.J.R., Gómez, V.A., Eds.; UJAT: Villahermosa, Tabasco, Mexico, 2011; pp. 100–225. [Google Scholar]
- Sandoval-García, A.M.; Altamirano-Cárdenas, J.R.; Aguilar-Ávila, J.; García-Muñiz, J.G. Caracterización química del aceite obtenido por métodos artesanales de tres variedades de palma africana (Elaeis guineensis Jacq.). Rev. Fitotec. Mex. 2016, 39, 317–322. [Google Scholar]
- Mayes, S.; Jack, P.L.; Corley, R.H.V. The use of molecular markers to investigate the genetic structure of an oil palm breeding programme. Heredity 2000, 85, 288–293. [Google Scholar] [CrossRef]
- Xia, W.; Luo, T.; Zhang, W.; Mason, A.S.; Huang, D.; Huang, X.; Tang, W.; Dou, Y.; Zhang, C.; Xiao, Y. Development of high-density SNP markers and their application in evaluating genetic diversity and population structure in Elaeis guineensis. Front. Plant Sci. 2019, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Arif, I.A.; Bakir, M.A.; Khan, H.A.; Al Farhan, A.H.; Al Homaidan, A.A.; Bahkali, A.H.; Al Saadon, M.; Shobrak, M. A brief review of molecular techniques to assess plant diversity. Int. J. Mol. Sci. 2010, 11, 2079–2096. [Google Scholar] [CrossRef] [PubMed]
- Mayes, S.; James, C.; Price, Z.; Groves, C.M.; Jack, P.L.; Corley, R.H.V. The application of restriction fragment length polymorphism for the genetic fingerprinting of oil palm (Elaeis guineensis). Mol. Breed. 1996, 2, 175–180. [Google Scholar]
- Billotte, N.; Rusterucci, A.M.; Barcelos, E.; Noyer, J.L.; Amblard, P.; Baurens, F.C. Development, characterization, and across-taxa utility of oil palm (Elaeis guineensis Jacq.) microsatellite markers. Genome 2001, 44, 413–425. [Google Scholar]
- Barcelos, E.; Amblard, P.; Berthaud, J.; Seguin, M. Genetic diversity and relationship in American and African oil palm as revealed by RFLP and AFLP molecular markers. Pesqui. Agropecu. Bras. 2002, 37, 1105–1114. [Google Scholar]
- Rocha, P.J. Marcadores moleculares, una herramienta útil para la selección genética de palma de aceite. Rev. Palmas 2003, 24, 11–25. [Google Scholar]
- Mayes, S.; Jack, P.L.; Marshall, D.F.; Corley, R.H.V. Construction of RFLP genetic linkage map for oil palm (Elaeis guineensis Jacq.). Genome 1997, 40, 116–122. [Google Scholar] [PubMed]
- Billotte, N.; Frances, L.; Amblard, P.; Durand-Gasselin, T.; Noyer, J.L.; Courtois, B. Search for AFLP and microsatellite molecular markers of the SH gene in oil palm (Elaeis guineensis Jacq) by bulk segregant analysis (BSA) and by genetic mapping. In Cutting-edge Technologies for Sustained Competitiveness, Proceedings of the 2001 PIPOC International Palm Oil Congress, Agriculture Conference, Kuala Lumpur, Malaysia, 20–22 August 2001; Malaysian Palm Oil Board (MPOB): Kuala Lumpur, Malaysia, 2000. [Google Scholar]
- Tapia-Tussell, R.; Quijano-Ramayo, A.; Rojas-Herrera, R.; Larque-Saavedra, A.; Perez-Brito, D. A fast, simple, and reliable high-yielding method for DNA extraction from different plant species. Mol. Biotechnol. 2005, 31, 137–139. [Google Scholar]
- Trejo-Saavedra, D.L. Desarrollo y validación de metodología para la detección de transgenes en organismos genéticamente modificados y sus subproductos. Ph.D. Thesis, Centro de Investigación y Estudios Avanzados del Instituto Politécnico Nacional (CINVESTAV), Irapuato, Guanajuato, Mexico, 2002. [Google Scholar]
- Castañeda-Cardona, C.C.; Murillo-Coronado, Y.; Murillo-Coronado, A.C.; Ochoa, I. Genetic diversity in oil palm (Elaeis guineensis Jacq) using RAM (Random Amplified Microsatellites). Bragantia 2018, 77, 546–556. [Google Scholar] [CrossRef]
- Hoisington, D.; Khairallah, M.; Gonzalez-de-Leon, D. Laboratory Protocols: CIMMYT Applied Molecular Genetics Laboratory, 2nd ed.; Cimmyt: Mexico, Mexico, 1994; pp. 51–52. [Google Scholar]
- Jaccard, P. Nouvelles recherché sur la distribution florale. Bull. Soc. Vaudoise Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Exeter Software: Setauket, NY, USA, 2000. [Google Scholar]
- Anderson, J.A.; Churcill, G.A.; Autrique, J.E.; Tanksley, S.D.; Sorrels, M.E. Optimizing parental selection for genetic linkage maps. Genome 1992, 36, 181–186. [Google Scholar]
- Billotte, N.; Marseillac, N.; Risterucci, A.M.; Adon, B.; Brottier, P.; Baurens, F.C.; Singh, R.; Herrán, A.; Asmady, H.; Billot, C.; et al. Microsatellite-based high density linkage map in oil palm (Elaeis guineensis Jacq.). Theor. Appl. Genet. 2005, 110, 754–765. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Yusop, M.R.; Ithnin, M.; Saleh, G.; Latif, M.A. Genetic variability of oil palm parental genotypes and performance of its’ progenies as revealed by molecular markers and quantitative traits. C. R. Biol. 2011, 334, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.S.; Murphy, J.P. The effect of parental divergence on F2 heterosis in winter wheat crosses. Theor. Appl. Genet. 1990, 79, 241–250. [Google Scholar]
- Okwuagwu, C.O.; Okoye, M.N.; Okolo, E.C.; Ataga, C.D.; Uguru, M.I. Genetic variability of fresh fruit bunch yield in Deli/dura x tenera breeding populations of oil palm (Elaeis guineensis Jacq.) in Nigeria. J. Trop. Agric. 2008, 46, 52–57. [Google Scholar] [CrossRef]
- Gan, S.T.; Teo, C.J.; Manirasa, S.; Wong, W.C.; Wong, C.K. Assessment of genetic diversity and population structure of oil palm (Elaeis guineensis Jacq.) field genebank: A step towards molecular-assisted germplasm conservation. PLoS ONE 2021, 16, e0255418. [Google Scholar] [CrossRef]
- Bakoumé, C.; Wickneswari, R.; Siju, S.; Rajanaidu, N.; Kushairi, A.; Billotte, N. Genetic diversity of the world’s largest oil palm (Elaeis guineensis Jacq.) field genebank accessions using microsatellite markers. Genet. Resour. Crop. Evol. 2015, 62, 349–360. [Google Scholar] [CrossRef]

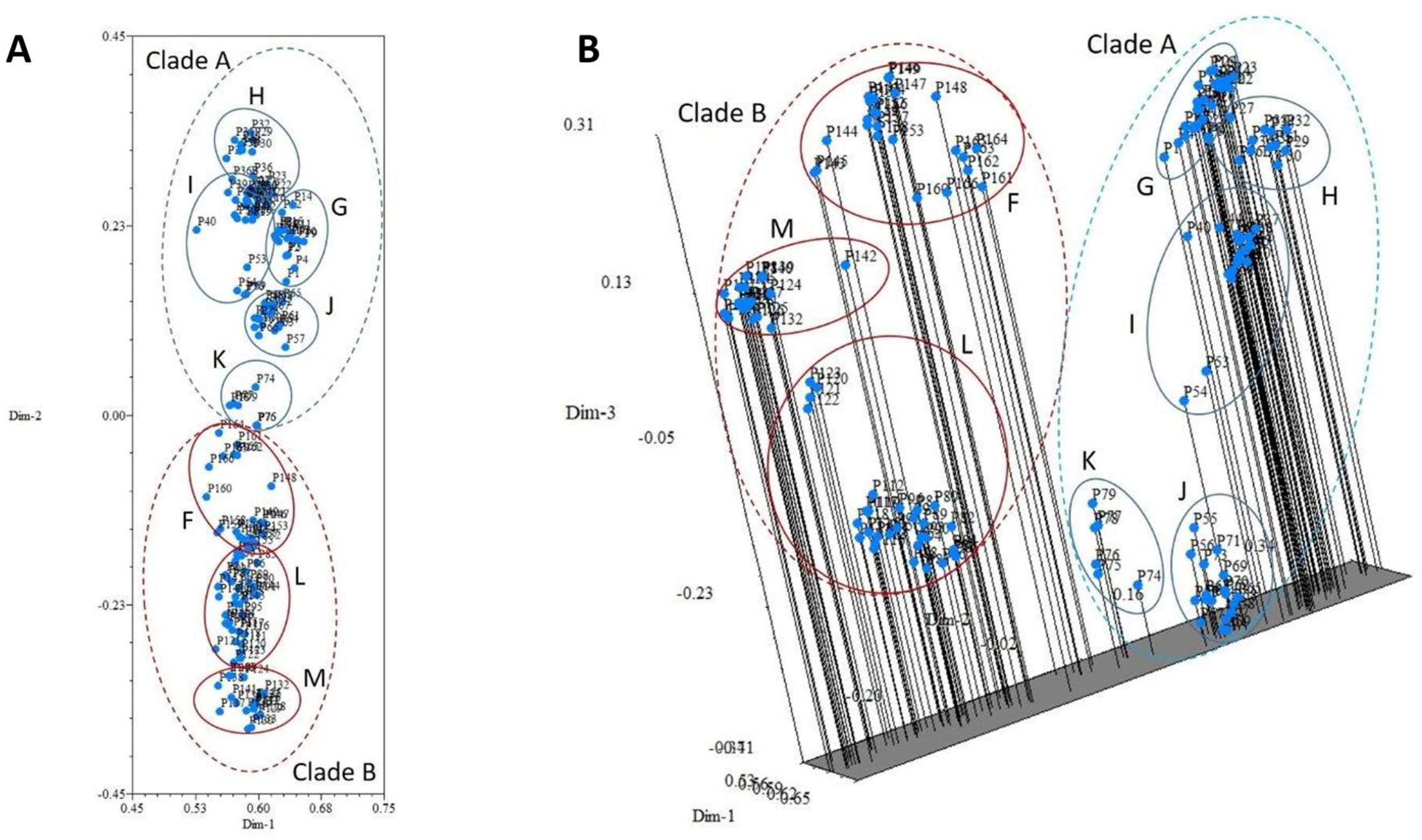

{kind=link}
{kind=link}
{kind=link}
| Primer Sequences 5′-3′ | |||
|---|---|---|---|
| SSR Locus | Motif | Forward Primer | Reverse Primer |
| mEgCIR0008 | (GA)18 | CGGAAAGAGGGAAGATG | ACCTTGATGATTGATGTGA |
| mEgCIR0009 | (GA)20 | CAGTCTTTAAGTACGGCTATGAT | GAATTTTTAGTTCAACCAGGTAGA |
| mEgCIR0018 | (GA)18 | CCTTATTTTCTTTGCTTACC | TTCTATTTTATTTTCTTCCT |
| mEgCIR0046 | (GA)19 | AGCCTTAGTATTTTGTTGAT | CCTCTGATTTGTCCTTTTGG |
| mEgCIR0067 | (GA)17 | TACACAACCCATGCACAT | AAAAACATCCAGAAATAAAA |
| mEgCIR0219 | (GA)17 | TTTGCTCGGCGGATACAT | CTCACTGGCCTCTTTCTT |
| mEgCIR0221 | (GA)11 | TGCCATGTTCCAGAGAGC | TTCAGATTTTTCCGACTTC |
| mEgCIR0230 | (TA)6GAG(GA)19 | CCCTGGCCCCGTTTTTC | AGCGCTATATGTGATTCTAA |
| mEgCIR0254 | (GA)18 | CCTTTTGTGCTTTCTTC | GCTGTGCACTAGGTTTC |
| mEgCIR0304 | (GT)4(GC)2(GT)2 | CCACAAACAATCCAAGCAAGT | TGGCATACACGAAAGCATAA |
| mEgCIR0326 | (GT)9 | GCTAACCACAGGCAAAAACA | AAGCCGCACTAACATACACATC |
| mEgCIR0332 | (GT)8 | ATTTCGTAAGGTGGGTGT | CCTCCAAACTCCTCTGT |
| mEgCIR0350 | (GT)8 | AAATCCTAAATCCTAAACTC | TCTACCTGTACTGGTGACAA |
| mEgCIR0353 | (GT)11(GA)15 | AGAGAGAGAGAGTGCGTATG | GTCCCTGTGGCTGCTGTTTC |
| mEgCIR0377 | (GT)6(GC)4 | GTCTGCTAAAACATCAACTG | GAGGAGGAGGGGAACGATAA |
| mEgCIR1772 | (GT)22 | ACCTTGTATTAGTTTGTCCA | CTTCCATTGTCTCATTATTCTCTTA |
| mEgCIR0391 | (CCG)5 | TTCATCGCCTTCCCCTCTG | CCCGACCTAATCCAACATC |
| mEgCIR0437 | (CCG)6 | CCAACCCAACCCAACATAAA | GGTCCCGATCCCGTCTACT |
| mEgCIR0465 | (CCG)6 | TCCCCCACGACCCATTC | GGCAGGAGAGGCAGCATTC |
| mEgCIR0476 | (CCG)7 | TTCCTCGGCCCCTTCTC | TCGCCGACCTTCCACTG |
| mEgCIR0134 | (GA)15 | AGTTTGGGGCTTACCTG | CTTCCACGCACCCTCTC |
| Marker | Primer Sequences 5′-3′ |
|---|---|
| CCA | DDB(CCA)5 |
| CGA | DHB(CGA)5 |
| AG | HBH(AG)7A |
| CT | DYD(CT)7C |
| TG | HVH(TG)7T |
| CA | DBDA(CA)7 |
| ACA | BDB(ACA)5 |
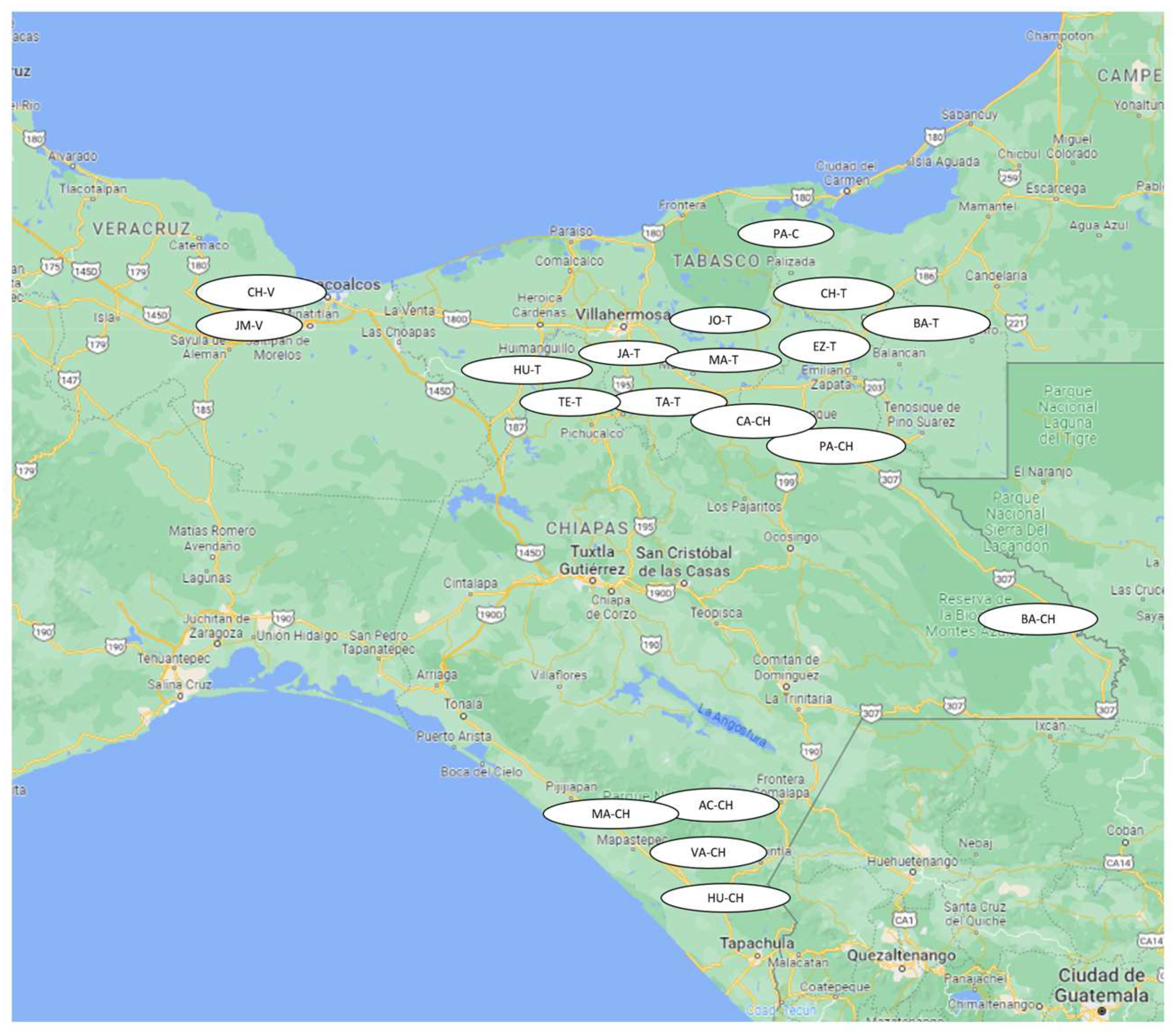
| Location Key | State | Municipality | Number of Samples |
|---|---|---|---|
| PA-C | Campeche | Palizada | 18 |
| EZ-T | Tabasco | Emiliano Zapata | 3 |
| HU-T | Huimanguillo | 18 | |
| JA-T | Jalapa | 2 | |
| JO-T | Jonuta | 3 | |
| TA-T | Tacotalpa | 8 | |
| TE-T | Teapa | 6 | |
| MA-T | Macuspana | 3 | |
| BA-T | Balancan | 3 | |
| CH-T | Chable | 2 | |
| PA-CH | Chiapas | Palenque | 18 |
| CA-CH | Catazaja | 3 | |
| BA-CH | Benemerito de las Americas | 21 | |
| AC-CH | Acapetahua | 8 | |
| HU-CH | Huixtla | 4 | |
| MA-CH | Mapaspetec | 6 | |
| VA-CH | Villa Comaltitlan | 17 | |
| CH-V | Veracruz | Chinameca | 4 |
| JM-V | Jaltipán de Morelos | 4 | |
| Total | 151 |
| Primer | Number of Amplified Bands | Number of Polymorphic Bands | Allele Size | Highest Frequency Alelle | PIC | |
|---|---|---|---|---|---|---|
| Range (bp) | Size (bp) | Frequency (%) | ||||
| RAM | ||||||
| CCA | 161 | 161 | 185–1800 | 525 | 75.5 | 0.94 |
| CGA | 183 | 179 | 250–4000 | 290 | 100.0 | 0.96 |
| 350 | 100.0 | |||||
| 400 | 100.0 | |||||
| 450 | 100.0 | |||||
| AG | 132 | 132 | 160–990 | 500 | 60.2 | 0.93 |
| CT | 203 | 203 | 230–1800 | 340 850 | 62.2 | 0.97 |
| TG | 167 | 163 | 200–7000 | 225 | 100.0 | 0.95 |
| 260 | 100.0 | |||||
| 290 | 100.0 | |||||
| 500 | 100.0 | |||||
| CA | 218 | 218 | 300–3000 | 530 | 50.3 | 0.97 |
| ACA | 154 | 154 | 230–12,000 | 575 | 69.5 | 0.95 |
| Total | 1218 | 1210 | 160–12,000 | - | - | - |
| Average | 174 | 173 | - | - | 85.9 | - |
| Overall | - | - | - | - | - | 0.96 |
| SSR | ||||||
| mEgCIR0008 | 80 | 80 | 160–3000 | 220 | 59.6 | 0.96 |
| mEgCIR0009 | 35 | 35 | 150–490 | 180 | 64.2 | 0.92 |
| mEgCIR0018 | 12 | 10 | 160–410 | 180 | 99.3 | 0.77 |
| 340 | ||||||
| mEgCIR0067 | 64 | 64 | 140–370 | 295 | 30.4 | 0.98 |
| mEgCIR0134 | 60 | 60 | 125–440 | 175 | 69.5 | 0.95 |
| mEgCIR0219 | 72 | 72 | 165–920 | 800 | 82.7 | 0.96 |
| mEgCIR0221 | 93 | 91 | 190–850 | 190 | 100 | 0.95 |
| 265 | 100 | |||||
| mEgCIR0230 | 51 | 51 | 225–960 | 345 | 73.5 | 0.88 |
| mEgCIR0254 | 31 | 31 | 140–450 | 160 | 54.3 | 0.94 |
| mEgCIR0304 | 3 | 0 | 230–270 | 230 | 100.0 | 0.00 |
| 250 | 100.0 | |||||
| 270 | 100.0 | |||||
| mEgCIR0326 | 4 | 0 | 190–700 | 190 | 100.0 | 0.00 |
| 220 | 100.0 | |||||
| 280 | 100.0 | |||||
| 700 | 100.0 | |||||
| mEgCIR0332 | 9 | 9 | 125–280 | 270 | 60.0 | 0.88 |
| mEgCIR0350 | 37 | 37 | 260–800 | 330 | 51.0 | 0.94 |
| 710 | 51.0 | |||||
| mEgCIR0353 | 38 | 38 | 60–875 | 80 | 80.7 | 0.94 |
| mEgCIR0377 | 12 | 9 | 110–450 | 110 | 100.0 | 0.69 |
| 160 | 100.0 | |||||
| 170 | 100.0 | |||||
| mEgCIR0391 | 16 | 15 | 150–750 | 300 | 100.0 | 0.80 |
| mEgCIR0437 | 5 | 2 | 180–330 | 180 | 100.0 | 0.39 |
| 250 | 100.0 | |||||
| 275 | 100.0 | |||||
| mEgCIR0465 | 13 | 13 | 130–220 | 130 | 72.1 | 0.91 |
| mEgCIR0476 | 54 | 49 | 150–6000 | 175 | 100.0 | 0.86 |
| 190 | 100.0 | |||||
| mEgCIR1772 | 19 | 19 | 180–350 | 195 | 84.7 | 0.82 |
| Total | 708 | 685 | 60–6000 | - | - | - |
| Average | 35 | 34 | - | - | 85.4 | - |
| Overall | - | - | - | - | - | 0.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magaña-Álvarez, A.; Pérez-Brito, D.; Cortés-Velázquez, A.; Nexticapan-Garcéz, Á.; Ortega-Ramírez, M.E.; García-Cámara, I.; Sánchez-Rodríguez, Y.; Martín-Mex, R. Genetic Variability of Oil Palm in Mexico: An Assessment Based on Microsatellite Markers. Agriculture 2023, 13, 1772. https://doi.org/10.3390/agriculture13091772
Magaña-Álvarez A, Pérez-Brito D, Cortés-Velázquez A, Nexticapan-Garcéz Á, Ortega-Ramírez ME, García-Cámara I, Sánchez-Rodríguez Y, Martín-Mex R. Genetic Variability of Oil Palm in Mexico: An Assessment Based on Microsatellite Markers. Agriculture. 2023; 13(9):1772. https://doi.org/10.3390/agriculture13091772
Chicago/Turabian StyleMagaña-Álvarez, Anuar, Daisy Pérez-Brito, Alberto Cortés-Velázquez, Ángel Nexticapan-Garcéz, Marynor E. Ortega-Ramírez, Isabel García-Cámara, Yasmín Sánchez-Rodríguez, and Rodolfo Martín-Mex. 2023. "Genetic Variability of Oil Palm in Mexico: An Assessment Based on Microsatellite Markers" Agriculture 13, no. 9: 1772. https://doi.org/10.3390/agriculture13091772
APA StyleMagaña-Álvarez, A., Pérez-Brito, D., Cortés-Velázquez, A., Nexticapan-Garcéz, Á., Ortega-Ramírez, M. E., García-Cámara, I., Sánchez-Rodríguez, Y., & Martín-Mex, R. (2023). Genetic Variability of Oil Palm in Mexico: An Assessment Based on Microsatellite Markers. Agriculture, 13(9), 1772. https://doi.org/10.3390/agriculture13091772