Fertilization with Olive Mill Pomace Compost Can Moderate Pest Damage in a Superintensive Olive Grove


Abstract
:1. Introduction
2. Materials and Methods
2.1. Location
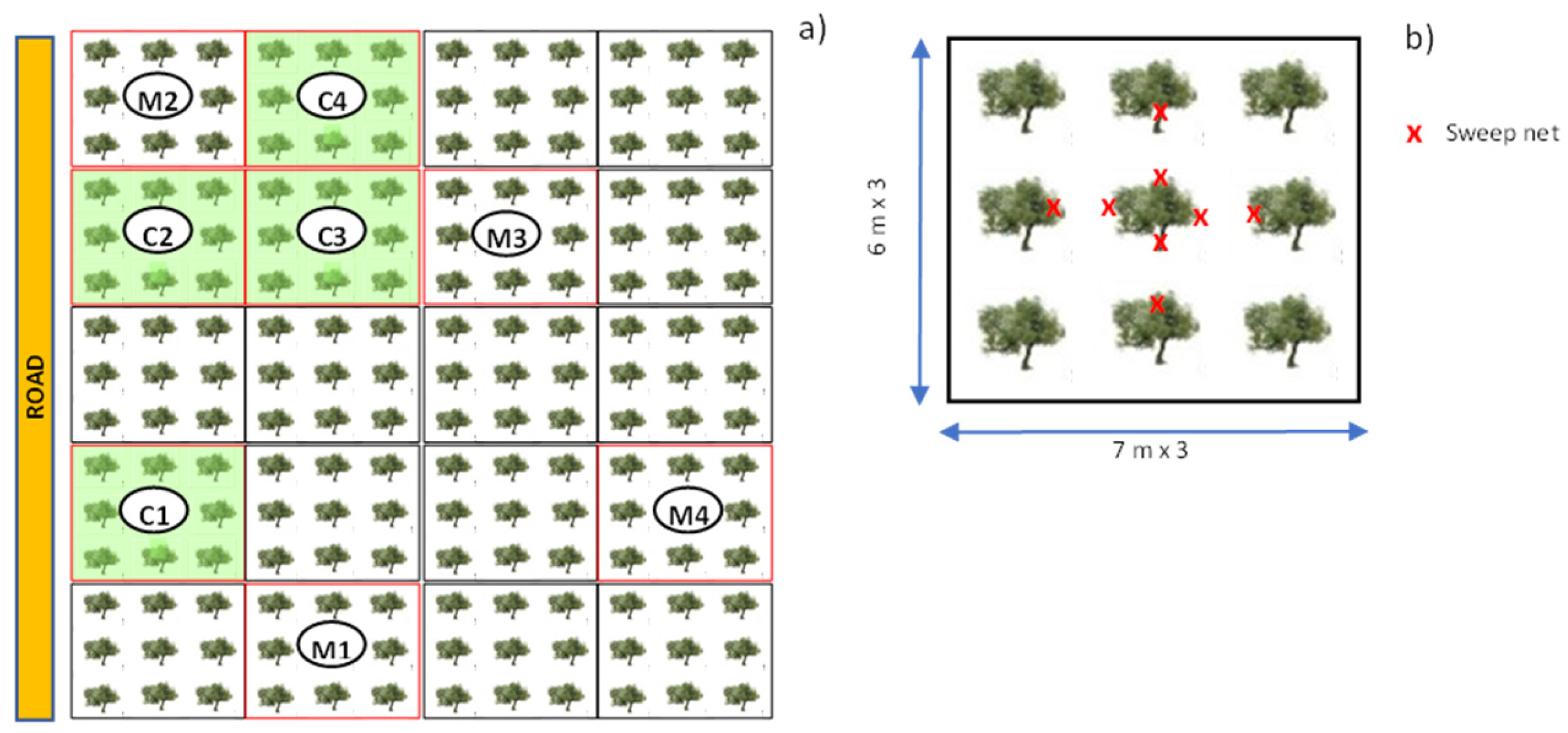
2.2. Experimental Design
2.3. Sampling
2.4. Data Analysis
3. Results
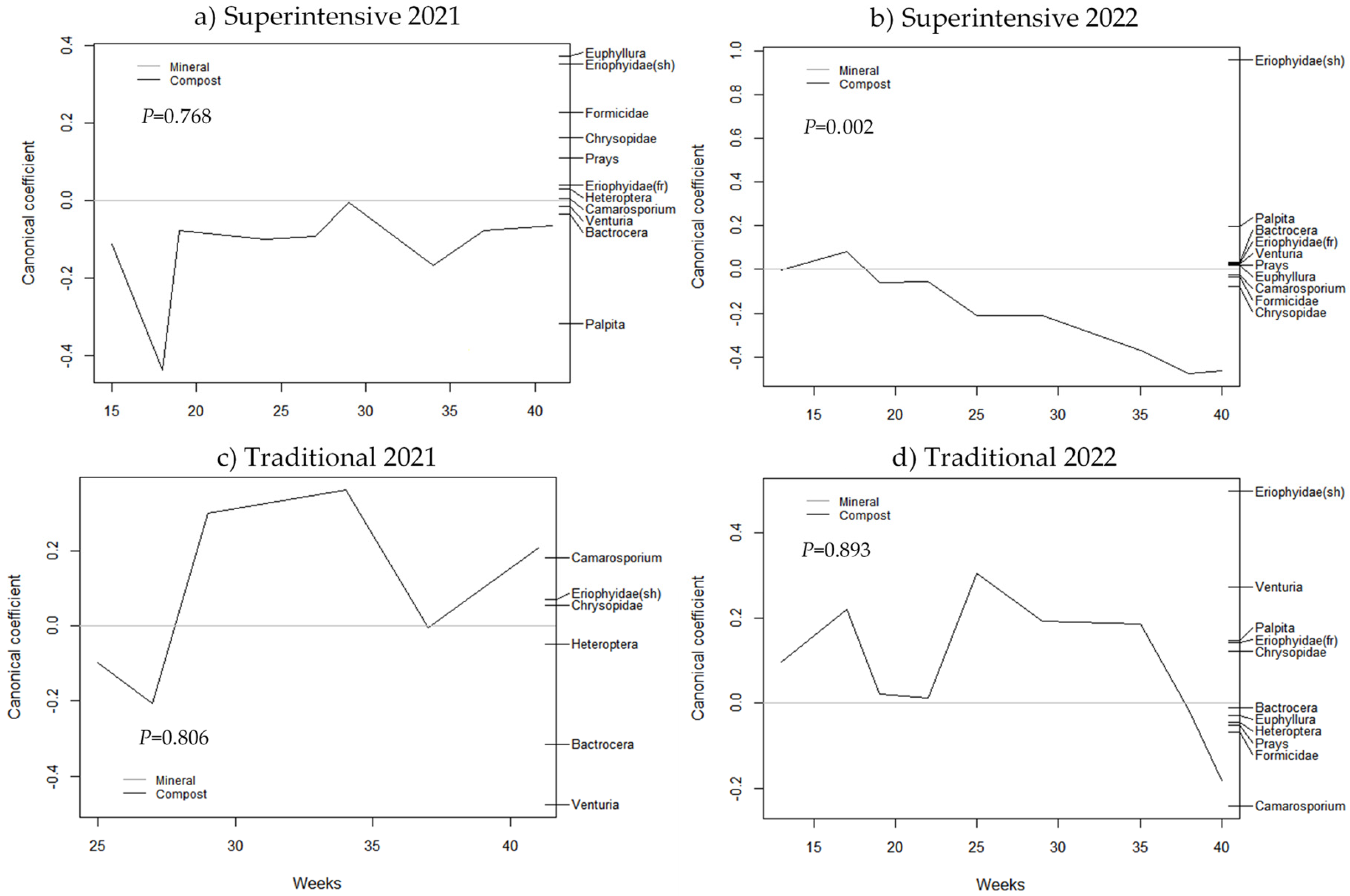
3.1. Principal Response Curves
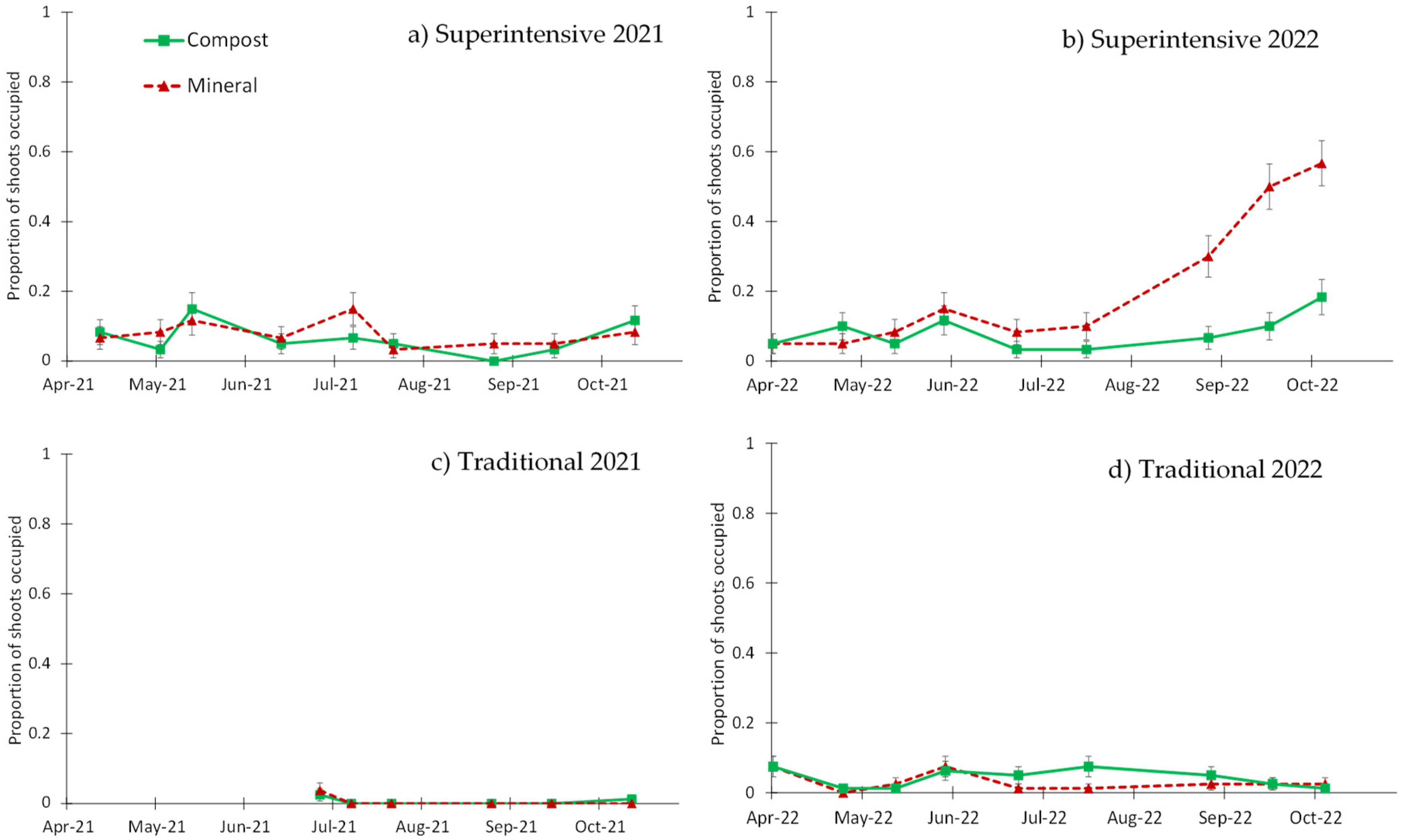
3.2. Visual Sampling
3.3. Sweep Net Sampling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Units | Mean | SD |
|---|---|---|---|
| Density | g cm−3 | 0.49 | 0.03 |
| Humidity | % | 21.5 | 0.02 |
| Organic Materia | % | 58.5 | 1.34 |
| pH | 9.18 | 0.09 | |
| CE | mS cm−1 | 5.87 | 0.25 |
| N | % | 2.50 | 0.26 |
| N-NH4+ | mg kg−1 | 56.8 | 2.68 |
| N-NO3− | mg kg−1 | 26.2 | 0.77 |
| P2O5 | % | 2.53 | 0.21 |
| CaO | % | 9.40 | 1.04 |
| MgO | % | 2.34 | 0.01 |
| K2O | % | 3.38 | 0.07 |
| SO3 | % | 2.14 | 0.06 |
| Na | % | 0.93 | 0.01 |
| Fe | mg kg−1 | 5628 | 642 |
| Mn | mg kg−1 | 332 | 35 |
| Cu | mg kg−1 | 70.2 | 7.80 |
| Zn | mg kg−1 | 301 | 21.3 |
| As | mg kg−1 | <0.10 | 0.00 |
| Cd | mg kg−1 | 0.10 | 0.03 |
| Co | mg kg−1 | 1.70 | 0.11 |
| Ni | mg kg−1 | 19.0 | 1.06 |
| Pb | mg kg−1 | 5.48 | 0.64 |
| Hg | mg kg−1 | 1.00 | 0.05 |
| Cr | mg kg−1 | 38.6 | 2.65 |
| Superintensive | Traditional | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 a | 2021 a | ||||||||||||||
| Mineral | Compost b | Mineral | Compost c | ||||||||||||
| Foliar | Irrigation | Total | Foliar | Compost | Total | Foliar | Mineral | Total | Foliar | Compost | Total | ||||
| N | 6.4 | -- | 6.4 | 6.4 | 14.1 | 20.5 | 6.4 | -- | 6.4 | 6.4 | 14.1 | 20.5 | |||
| P | -- | -- | -- | -- | 93.8 | 93.8 | -- | -- | -- | -- | 93.8 | 93.8 | |||
| K | 22.4 | -- | 22.4 | 22.4 | 477.0 | 499.4 | 22.4 | -- | 22.4 | 22.4 | 477.0 | 499.4 | |||
| 2022 | 2022 | ||||||||||||||
| Mineral d | Compost b | Mineral d | Compost c | ||||||||||||
| Foliar | Irrigation | Total | Foliar | Compost | Total | Foliar | Mineral | Total | Foliar | Compost | Total | ||||
| N | 6.4 | 119.0 | 125.4 | 6.4 | 14.1 | 20.5 | 6.4 | 72.9 | 79.3 | 6.4 | 14.1 | 20.5 | |||
| P | -- | 59.5 | 59.5 | -- | 93.8 | 93.8 | -- | 24.3 | 24.3 | -- | 93.8 | 93.8 | |||
| K | 22.4 | 178.6 | 201.0 | 22.4 | 477.0 | 499.4 | 22.4 | 97.1 | 119.5 | 22.4 | 477.0 | 499.4 | |||
| Sampling Dates | Agricultural Operations | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Visual | Sweep Net | |||||||||||
| Superintensive | Traditional | Superintensive | Traditional | Superintensive | Traditional | |||||||
| 12 April 2021 | 12 April 2021 | 1–8 April 2021 | Streets mowed | April 2021 | Plots tilled | |||||||
| 3 May 2021 | 3 May 2021 | 20–25 April 2021 | Trees pruned | |||||||||
| 14 May 2021 | 14 May 2021 | 1–3 July 2021 | Compost addition 1 | |||||||||
| 14 June 2021 | 28 June 2021 | 14 June 2021 | 28 June 2021 | 17 September 2021 | Harvest | November 2021 | Harvest | |||||
| 9 July 2021 | 9 July 2021 | 12 July 2021 | 12 July 2021 | 15 October 2021 | Streets mowed | |||||||
| 23 July 2021 | 23 July 2021 | 26 July 2021 | 26 July 2021 | |||||||||
| 27 August 2021 | 27 August 2021 | 30 August 2021 | 30 August 2021 | |||||||||
| 17 September 2021 | 17 September 2021 | 20 September 2021 | 20 September 2021 | |||||||||
| 15 October 2021 | 15 October 2021 | 18 October 2021 | 18 October 2021 | |||||||||
| 1 April 2022 | 1 April 2022 | 1 April 2022 | 1 April 2022 | March 2022 | Nitrofoska Perfect (15–5–20) | March 2022 | Nitrofoska Perfect (15–5–20) | |||||
| 25 April 2022 | 25 April 2022 | 25 April 2022 | 25 April 2022 | 4 March 2022 | Compost addition 1 | 28–31 March 2022 | Compost addition 2 | |||||
| 13 May 2022 | 13 May 2022 | 13 May 2022 | 13 May 2022 | 13–17 May 2022 | Streets mowed | 20–22 April 2022 | Plots tilled | |||||
| 30 May 2022 | 30 May 2022 | 30 May 2022 | 30 May 2022 | 15 September 2022 | Harvest | November 2022 | Harvest | |||||
| 24 June 2022 | 24 June 2022 | 24 June 2022 | 24 June 2022 | |||||||||
| 18 July 2022 | 18 July 2022 | 15 July 2022 | 15 July 2022 | |||||||||
| 29 August 2022 | 29 August 2022 | 26 August 2022 | 26 August 2022 | |||||||||
| 19 September 2022 | 19 September 2022 | 16 September 2022 | 16 September 2022 | |||||||||
| 7 October 2022 | 7 October 2022 | 7 October 2022 | 7 October 2022 | |||||||||
| Superintensive Grove | ||
|---|---|---|
| Date | Product Used | Used Against |
| 1 January 2021 | Glyphosate 36% | Mono and Dicotyledoneae |
| MCPA 40% | Dicotyledoneae | |
| 15 February 2021 | Copper oxychloride 52% | Venturia oleaginea—Other diseases |
| 19 April 2021 | Lambda-cyhalothrin 10% | Prays oleae |
| 19 May 2021 | Deltamethrin 2.5% | Bactrocera oleae—Prays oleae |
| 22 June 2021 | Glyphosate 36% | Mono and Dicotyledoneae |
| MCPA 40% | Dicotyledoneae | |
| 23 June 2021 | Phosmet 50% | Bactrocera oleae—Prays oleae—Palpita unionalis |
| 5 November 2021 | Copper oxychloride 52% | Venturia oleaginea—Other diseases |
| Kresoxim methyl 50% | ||
| 16 February 2022 | Copper oxychloride 52% | Venturia oleaginea |
| 20 April 2022 | Glyphosate 36% | Mono and Dicotyledoneae |
| 26 April 2022 | Deltamethrin 2.5% | Prays oleae |
| 27 May 2022 | Kresoxim methyl 50% | Venturia oleaginea |
| Lambda-cyhalothrin 10% | Prays oleae | |
| 2 June 2022 | Glyphosate 36% | Mono and Dicotyledoneae |
| 22 June 2022 | Sulfur 80% | Eriophyidae |
| 13 July 2022 | Phosmet 50% | Bactrocera oleae—Prays oleae—Palpita unionalis |
| 4 August 2022 | Phosmet 50% | Bactrocera oleae—Prays oleae—Palpita unionalis |
| 26 October 2022 | Copper oxychloride 52% | Venturia oleaginea |
| Traditional grove | ||
| Date | Product used | Used against |
| 5 November 2021 | Copper oxychloride 52% | Venturia oleaginea |
| 26 April 2022 | Deltamethrin 2.5% | Prays oleae |
| 30 May 2022 | Deltamethrin 2.5% | Prays oleae |
| Difenoconazole 23.5% | Venturia oleaginea | |
| 13 July 2022 | Phosmet 50% | Bactrocera oleae—Prays oleae—Palpita unionalis |
| 4 August 2022 | Phosmet 50% | Bactrocera oleae—Prays oleae—Palpita unionalis |
| 27 October 2022 | Copper oxychloride 52% | Venturia oleaginea |
References
- International Olive Council. Olive Oil Production. Available online: https://www.internationaloliveoil.org/what-we-do/economic-affairs-promotion-unit/#figures (accessed on 8 May 2023).
- Morillo, J.A.; Antizar-Ladislao, B.; Monteoliva-Sánchez, M.; Ramos-Cormenzana, A.; Russell, N.J. Bioremediation and biovalorisation of olive-mill wastes. Appl. Microbiol. Biotechnol. 2009, 82, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Peña, D.; Fernández, D.; Albarrán, A.; Gómez, S.; Martín, C.; Sánchez-Terrón, J.; Vicente, L.; López-Piñeiro, A. Using olive mill waste compost with sprinkler irrigation as a strategy to achieve sustainable rice cropping under Mediterranean conditions. Agron. Sustain. Dev. 2022, 42, 36. [Google Scholar] [CrossRef]
- Sánchez-Monedero, M.A.; Cayuela, M.L.; Sánchez-García, M.; Vandecasteele, B.; D’Hose, T.; López, G.; Martínez-Gaitán, C.; Kuikman, P.J.; Sinicco, T.; Mondini, C. Agronomic Evaluation of Biochar, Compost and Biochar-Blended Compost across Different Cropping Systems: Perspective from the European Project FERTIPLUS. Agronomy 2019, 9, 225. [Google Scholar] [CrossRef]
- Leone, A.; Romaniello, R.; Tamborrino, A.; Beneduce, L.; Gagliardi, A.; Giuliani, M.; Gatta, G. Composting of Olive Mill Pomace, Agro-Industrial Sewage Sludge and Other Residues: Process Monitoring and Agronomic Use of the Resulting Composts. Foods 2021, 10, 2143. [Google Scholar] [CrossRef]
- Diacono, M.; Montemurro, F. Olive Pomace Compost in Organic Emmer Crop: Yield, Soil Properties, and Heavy Metals’ Fate in Plant and Soil. J. Soil Sci. Plant Nutr. 2019, 19, 63–70. [Google Scholar] [CrossRef]
- Bargougui, L.; Guergueb, Z.; Chaieb, M.; Braham, M.; Mekki, A. Agro-physiological and biochemical responses of Sorghum bicolor in soil amended by olive mill wastewater. Agric. Water Manag. 2019, 212, 60–67. [Google Scholar] [CrossRef]
- Mamkagh, A.; AL-Zyoud, F.; Al-Atiyat, R. Olive Mill Pomace Compost as Soil Amendment, and Sources of Biopesticides and Animal Feed: A Review. Int. J. Agric. Biol. 2022, 28, 257–268. [Google Scholar]
- De Sosa, L.; Panettieri, M.; Moreno, B.; Benítez, E.; Madejón, E. Compost application in an olive grove influences nitrogen dynamics under Mediterranean conditions. Appl. Soil Ecol. 2022, 175, 104462. [Google Scholar] [CrossRef]
- Panettieri, M.; Moreno, B.; de Sosa, L.L.; Benítez, E.; Madejón, E. Soil management and compost amendment are the main drivers of carbon sequestration in rainfed olive trees agroecosystems: An evaluation of chemical and biological markers. Catena 2022, 214, 106258. [Google Scholar] [CrossRef]
- Madejón, P.; Alaejos, J.; García-Álbala, J.; Fernández, M.; Madejón, E. Three-year study of fast-growing trees in degraded soils amended with composts: Effects on soil fertility and productivity. J. Environ. Manag. 2016, 169, 18–26. [Google Scholar] [CrossRef]
- Podgornik, M.; Bučar-Miklavčič, M.; Levart, A.; Salobir, J.; Rezar, V.; Butinar, B. Chemical Characteristics of Two-Phase Olive-Mill Waste and Evaluation of Their Direct Soil Application in Humid Mediterranean Regions. Agronomy 2022, 12, 1621. [Google Scholar] [CrossRef]
- Hrameche, O.; Tul, S.; Manolikaki, I.; Digalaki, N.; Kaltsa, I.; Psarras, G.; Koubouris, G. Optimizing Agroecological Measures for Climate-Resilient Olive Farming in the Mediterranean. Plants 2024, 13, 900. [Google Scholar] [CrossRef] [PubMed]
- Proietti, P.; Federici, E.; Fidati, L.; Scargetta, S.; Massaccesi, L.; Nasini, L.; Regni, L.; Ricci, A.; Cenci, G.; Gigliotti, G. Effects of amendment with oil mill waste and its derived-compost on soil chemical and microbiological characteristics and olive (Olea europaea L.) productivity. Agric. Ecosyst. Environ. 2015, 207, 51–60. [Google Scholar] [CrossRef]
- Mairech, H.; López-Bernal, Á.; Moriondo, M.; Dibari, C.; Regni, L.; Proietti, P.; Villalobos, F.J.; Testi, L. Sustainability of olive growing in the Mediterranean area under future climate scenarios: Exploring the effects of intensification and deficit irrigation. Eur. J. Agron. 2021, 129. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- Murrell, E.G. Can agricultural practices that mitigate or improve crop resilience to climate change also manage crop pests? Curr. Opin. Insect Sci. 2017, 23, 81–88. [Google Scholar] [CrossRef]
- Lin, P.-A.; Paudel, S.; Lai, P.-C.; GC, R.K.; Yang, D.-H.; Felton, G.W. Integrating water and insect pest management in agriculture. J. Pest Sci. 2024, 97, 521–538. [Google Scholar] [CrossRef]
- Han, P.; Lavoir, A.-V.V.; Rodriguez-Saona, C.; Desneux, N. Bottom-Up Forces in Agroecosystems and Their Potential Impact on Arthropod Pest Management. Annu. Rev. Entomol. 2022, 67, 239–259. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Barzman, M.; Booij, K.; Boonekamp, P.; Desneux, N.; Huber, L.; Kudsk, P.; Langrell, S.R.H.; Ratnadass, A.; Ricci, P.; et al. Robust cropping systems to tackle pests under climate change. A review. Agron. Sustain. Dev. 2015, 35, 443–459. [Google Scholar] [CrossRef]
- Baldovino Sanjuan, A.; Castellanos Gonzalez, L.; Prado, R.d.M. Edaphic mesofauna, some studies done: A review. INGE CUC 2022, 18, 197–208. [Google Scholar] [CrossRef]
- Miller, J.J.; Battigelli, J.P.; Beasley, B.W.; Drury, C.F. Response of Soil Mesofauna to Long-Term Application of Feedlot Manure on Irrigated Cropland. J. Environ. Qual. 2017, 46, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Rowen, E.; Tooker, J.F.; Blubaugh, C.K. Managing fertility with animal waste to promote arthropod pest suppression. Biol. Control 2019, 134, 130–140. [Google Scholar] [CrossRef]
- Goncharov, A.A.; Glebova, A.A.; Chernov, T.I.; Karpukhin, M.M.; Kuznetsova, N.A.; Leonov, V.D.; Makarova, O.L.; Maygurova, V.N.; Shesteperov, A.A.; Taranets, I.P.; et al. Detrital subsidy alters the soil invertebrate community and reduces infection of winter wheat seedlings by Fusarium wilt. Appl. Soil Ecol. 2021, 163, 103914. [Google Scholar] [CrossRef]
- Hata, F.T.; Ventura, M.U.; de Souza, M.S.d.J.; de Sousa, N.V.; Oliveira, B.G.; da Silva, J.B. Mineral and organic fertilization affects Tetranychus urticae, pseudofruit production and leaf nutrient content in strawberry. Phytoparasitica 2019, 47, 513–521. [Google Scholar] [CrossRef]
- Sousa, V.; Ventura, M.U.; Hoshino, A.T.; Hata, F.T.; Constantino, L.V. Development and population growth of the two-spotted spider mite (Tetranychus urticae Koch) on strawberry fertilized with different doses and sources of organic fertilizers. Int. J. Acarol. 2021, 47, 528–535. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Wright, D.J.; Leather, S.R. The effects of farming system and fertilisers on pests and natural enemies: A synthesis of current research. Agric. Ecosyst. Environ. 2011, 141, 261–270. [Google Scholar] [CrossRef]
- Martín Gil, A.; Ruiz Torres, M. Guía de Gestión Integrada de Plagas-Olivar; Ministerio de Agricultura, Pesca y Alimentacion: Madrid, Spain, 2014; ISBN 9788449113871. [Google Scholar]
- Rodrigues, M.Â.; Coelho, V.; Arrobas, M.; Gouveia, E.; Raimundo, S.; Correia, C.M.; Bento, A. The effect of nitrogen fertilization on the incidence of olive fruit fly, olive leaf spot and olive anthracnose in two olive cultivars grown in rainfed conditions. Sci. Hortic. 2019, 256, 108658. [Google Scholar] [CrossRef]
- Roca, L.F.; Romero, J.; Bohórquez, J.M.; Alcántara, E.; Fernández-Escobar, R.; Trapero, A. Nitrogen status affects growth, chlorophyll content and infection by Fusicladium oleagineum in olive. Crop Prot. 2018, 109, 80–85. [Google Scholar] [CrossRef]
- González-Zamora, J.E.; Alonso-López, M.T.; Gómez-Regife, Y.; Ruiz-Muñoz, S. Decreased water use in a super-intensive olive orchard mediates arthropod populations and pest damage. Agronomy 2021, 11, 1337. [Google Scholar] [CrossRef]
- González-Zamora, J.E.; Gamero-Monge, J.M.; Pérez-de la Luz, R. The use of olive mill pomace compost increases the population of certain ground/soil organisms in olive groves. Eur. J. Soil Biol. 2024, 122, 103668. [Google Scholar] [CrossRef]
- González-Zamora, J.E.; Gamero-Monge, J.M.; Pérez-de la Luz, R. Ant Community Is Not Influenced by the Addition of Olive Mill Pomace Compost in Two Different Olive Crop Managements. Insects 2023, 14, 783. [Google Scholar] [CrossRef] [PubMed]
- De Sosa, L.L.; Martín-Palomo, M.J.; Castro-Valdecantos, P.; Madejón, E. Agricultural use of compost under different irrigation strategies in a hedgerow olive grove under Mediterranean conditions—A comparison with traditional systems. Soil 2023, 9, 325–338. [Google Scholar] [CrossRef]
- González, M.I.; Alvarado, M.; Durán, J.M.; De la Rosa, A.; Serrano, A. Los eriófidos (Acarina, Eriophidae) del olivar de la provincia de Sevilla. Problemática y control. Boletín Sanid. Veg. 2000, 26, 203–214. [Google Scholar]
- Barrientos, A.J. (Ed.) Curso Práctico de Entomología; Manuals; 41; Asociación Española de Entomología; CIBIO.; Universidad Autónoma de Barcelona: Alicante, Bellaterra, Spain, 2004; ISBN 8449023831. [Google Scholar]
- Chinery, M. Guía de Campo de los Insectos de España y de Europa; Omega: Barcelona, Spain, 2005; ISBN 84-282-0469-1. [Google Scholar]
- R CoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. Available online: https://www.R-project.org/ (accessed on 8 May 2023).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. vegan: Community Ecology Package. R Package Version 2.6-8. 2024. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 November 2024).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Huber, D.; Römheld, V.; Weinmann, M. Relationship between Nutrition, Plant Diseases and Pests. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 283–298. ISBN 9780123849052. [Google Scholar]
- Cárdenas, M.; Pascual, F.; Campos, M.; Pekar, S. The Spider Assemblage of Olive Groves Under Three Management Systems. Environ. Entomol. 2015, 44, 509–518. [Google Scholar] [CrossRef]
- Gonçalves, M.F.; Pereira, J.A. Abundance and Diversity of Soil Arthropods in the Olive Grove Ecosystem. J. Insect Sci. 2012, 12, 20. [Google Scholar] [CrossRef]
- Álvarez, H.A.; Jiménez-Muñoz, R.; Morente, M.; Campos, M.; Ruano, F. Ground cover presence in organic olive orchards affects the interaction of natural enemies against Prays oleae, promoting an effective egg predation. Agric. Ecosyst. Environ. 2021, 315, 107441. [Google Scholar] [CrossRef]
- Cárdenas, M.; Ruano, F.; García, P.; Pascual, F.; Campos, M. Impact of agricultural management on spider populations in the canopy of olive trees. Biol. Control 2006, 38, 188–195. [Google Scholar] [CrossRef]
- Anđelić Dmitrović, B.; Gajski, D.; Kos, T.; Jelić, M.; Šerić Jelaska, L. Insight into Trophic Interactions of Spiders in Olive Groves with Integrated and Ecological Pest Management Using DNA Metabarcoding. Diversity 2023, 15, 976. [Google Scholar] [CrossRef]
- Morente, M.; Ruano, F. Understanding the trophic relationships amongst arthropods in olive grove by δN 15 and δC 13 stable isotope analysis. J. Appl. Entomol. 2022, 146, 372–384. [Google Scholar] [CrossRef]
- Alvarado, M.; Durán, J.M.; González, M.I.; Montes, F.; Páez, J.; Sánchez, A.; Serrano, A.; Vega, J.M. Plagas y Enfermedades del Olivo; Junta de Andalucía, Consejería de Agricultura y Pesca: Sevilla, Spain, 2002. [Google Scholar]
- Sobreiro, J.; Patanita, M.I.; Patanita, M.; Tomaz, A. Sustainability of High-Density Olive Orchards: Hints for Irrigation Management and Agroecological Approaches. Water 2023, 15, 2486. [Google Scholar] [CrossRef]
- Scriber, J.M. Nitrogen Nutrition of Plants and Insect Invasion. In Nitrogen in Crop Protection; Hauck, R.D., Ed.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 441–460. [Google Scholar]
- Hasken, K.H.; Poehling, H.M. Effects of different intensities of fertilisers and pesticides on aphids and aphid predators in winter wheat. Agric. Ecosyst. Environ. 1995, 52, 45–50. [Google Scholar] [CrossRef]
- Kaneshiro, L.N.; Johnson, M.W. Tritrophic Effects of Leaf Nitrogen on Liriomyza trifolii (Burgess) and an Associated Parasitoid Chrysocharis oscinidis (Ashmead) on Bean. Biol. Control 1996, 6, 186–192. [Google Scholar] [CrossRef]
- Bi, J.L.; Ballmer, G.R.; Hendrix, D.L.; Henneberry, T.J.; Toscano, N.C. Effect of cotton nitrogen fertilization on Bemisia argentifolii populations and honeydew production. Entomol. Exp. Appl. 2001, 99, 25–36. [Google Scholar] [CrossRef]
- Nevo, E.; Coll, M. Effect of Nitrogen Fertilization on Aphis gossypii (Homoptera: Aphididae): Variation in Size, Color, and Reproduction. J. Econ. Entomol. 2001, 94, 27–32. [Google Scholar] [CrossRef]
| Superintensive | Traditional | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | 2021–2022 | 2021 | 2022 | 2021–2022 | ||||||||||||||
| Treatment | Tr × SD | Treatment | Tr × SD | Treatment | Year | Tr × Y | Treatment | Tr × SD | Treatment | Tr × SD | Treatment | Year | Tr × Y | ||||||
| Prays oleae | 0.435 | 0.624 | - 1 | - 1 | 0.488 | 0.244 | 0.160 | - 2 | - 2 | - 1 | - 1 | - | - | - | |||||
| Palpita unionalis 3 | 0.067 | 0.021 (*) | 0.664 | 0.261 | 0.321 | 0.706 | 0.357 | - 1 | - 1 | 0.580 | 0.878 | 1 | 0.993 | 1 | |||||
| Euphyllura olivina | 0.015 (*) | 0.026 (*) | 0.041 (*) | 0.285 | 0.035 (*) | 0.652 | 0.004 (*) | - 2 | - 2 | 0.839 | 0.347 | - | - | - | |||||
| Formicidae | 0.033 (*) | 0.758 | 0.834 | 0.471 | 0.273 | 0.101 | 0.782 | - 1 | - 1 | 0.134 | 0.489 | 1 | 1 | 0.997 | |||||
| Neuroptera | 0.011 (*) | 0.598 | 0.116 | 0.497 | 0.016 (*) | 0.549 | 0.991 | 0.940 | 0.430 | 0.675 | 0.69 | - 4 | - 4 | - 4 | |||||
| Heteroptera 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |||||
| Eriophyidae (Shoot) | 0.662 | 0.399 | 0.023 (*) | 0.046 (*) | 0.596 | 0.496 | 0.057 | - 1 | - 1 | 0.473 | 0.559 | 1 | 0.004 | 0.732 | |||||
| Venturia oleaginea | 0.670 | 0.721 | 0.678 | 0.374 | 0.736 | 0.062 | 0.595 | 0.632 | 0.081 | 0.301 | 0.231 | 0.767 | 0.018 | 0.319 | |||||
| Camarosporium dalmaticum (Fruit) | 0.919 | 0.422 | 0.374 | 0.374 | 0.798 | 0.037 | 0.993 | 0.720 | 0.359 | 1 | 0.020 (*) | 0.449 | 0.208 | 0.648 | |||||
| Eriophyidae (Fruit) | 0.329 | 0.741 | 0.643 | 0.260 | 0.655 | 0.785 | 0.876 | - 1 | - 1 | 0.356 | 0.284 | - 4 | - 4 | - 4 | |||||
| Bactrocera oleae (Fruit) | 0.653 | 0.909 | 0.374 | 0.683 | 0.573 | 0.245 | 0.275 | 0.039 (*) | 0.875 | 0.190 | 0.446 | 0.036 (*) | 0.228 | 0.021 (*) | |||||
| Superintensive | Traditional | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | TOTAL | 2021 | 2022 | TOTAL | |||||||||||||||||||||
| Compost | Mineral | Total | Compost | Mineral | Total | Compost | Mineral | Total | Compost | Mineral | Total | |||||||||||||||
| Mean | s.e. | Mean | s.e. | Mean | s.e. | Mean | s.e. | Mean | s.e. | Mean | s.e. | Mean | s.e. | Mean | s.e. | |||||||||||
| DIPTERA | 2.11 | (0.66) | 2.07 | (0.79) | 113 | 4.78 | (2.40) | 3.44 | (1.89) | 222 | 335 | 2.83 | (1.21) | 3.17 | (0.98) | 144 | 3.31 | (0.61) | 3.08 | (0.54) | 230 | 374 | ||||
| Nematocera | 2.00 | (0.69) | 1.89 | (0.81) | 105 | 4.37 | (2.44) | 3.22 | (1.91) | 205 | 310 | 2.79 | (1.23) | 3.08 | (1.02) | 141 | 2.58 | (0.70) | 2.42 | (0.62) | 180 | 321 | ||||
| Brachycera | 0.11 | (0.08) | 0.19 | (0.15) | 8 | 0.41 | (0.12) | 0.22 | (0.12) | 17 | 25 | 0.04 | (0.04) | 0.08 | (0.05) | 3 | 0.72 | (0.39) | 0.67 | (0.26) | 50 | 53 | ||||
| LEPIDOPTERA | 0.78 | (0.40) | 1.26 | (1.06) | 55 | 0.07 | (0.05) | 0.04 | (0.04) | 3 | 58 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.14 | (0.06) | 0.14 | (0.06) | 10 | 10 | ||||
| Prays oleae | 0.63 | (0.39) | 1.22 | (1.07) | 50 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 50 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0 | ||||
| Palpita unionalis 1 | 0.15 | (0.11) | 0.04 | (0.04) | 5 | 0.04 | (0.04) | 0.00 | (0.00) | 1 | 6 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.03 | (0.03) | 0.03 | (0.03) | 2 | 2 | ||||
| HYMENOPTERA | 3.15 | (0.95) | 3.74 | (1.23) | 186 | 3.81 | (1.40) | 7.07 | (2.53) | 294 | 480 | 0.17 | (0.08) | 0.17 | (0.08) | 8 | 0.83 | (0.25) | 0.89 | (0.31) | 62 | 70 | ||||
| Formicidae | 3.04 | (0.93) | 3.63 | (1.24) | 180 | 3.48 | (1.40) | 5.48 | (2.26) | 242 | 422 | 0.04 | (0.04) | 0.04 | (0.04) | 2 | 0.36 | (0.14) | 0.36 | (0.25) | 26 | 28 | ||||
| Ichneumonoidea | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.04 | (0.04) | 0.30 | (0.19) | 9 | 9 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.11 | (0.08) | 0.11 | (0.04) | 8 | 8 | ||||
| Chalcidoidea | 0.04 | (0.04) | 0.11 | (0.08) | 4 | 0.30 | (0.07) | 1.15 (*) | (0.46) | 39 | 43 (*) | 0.13 | (0.09) | 0.13 | (0.09) | 6 | 0.33 | (0.09) | 0.36 | (0.13) | 25 | 31 | ||||
| HEMIPTERA | 0.37 | (0.14) | 1.37 (*) | (0.42) | 47 | 1.07 | (0.37) | 1.15 | (0.43) | 60 | 107 (*) | 1.88 | (0.29) | 1.88 | (0.25) | 90 | 3.19 | (1.10) | 2.69 | (0.82) | 212 | 302 | ||||
| Cicadomorpha | 0.11 | (0.08) | 0.19 | (0.13) | 8 | 0.04 | (0.04) | 0.07 | (0.07) | 3 | 11 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.08 | (0.06) | 0.03 | (0.03) | 4 | 4 | ||||
| Fulgoromorpha | 0.04 | (0.04) | 0.00 | (0.00) | 1 | 0.04 | (0.04) | 0.07 | (0.05) | 3 | 4 | 0.17 | (0.08) | 0.08 | (0.05) | 6 | 0.50 | (0.26) | 0.33 | (0.14) | 30 | 36 | ||||
| Sternorrhyncha | 0.11 | (0.08) | 0.85 (*) | (0.32) | 26 | 0.89 | (0.31) | 0.89 | (0.44) | 48 | 74 (*) | 1.50 | (0.25) | 1.71 | (0.23) | 77 | 1.72 | (0.77) | 1.69 | (0.65) | 123 | 200 | ||||
| Heteroptera | 0.11 | (0.06) | 0.33 | (0.18) | 12 | 0.11 | (0.08) | 0.11 | (0.06) | 6 | 18 | 0.21 | (0.10) | 0.08 | (0.05) | 7 | 0.89 | (0.41) | 0.64 | (0.31) | 55 | 62 | ||||
| ARANEAE | 0.59 (*) | (0.13) | 0.19 | (0.06) | 21 | 1.15 (.) | (0.34) | 0.78 | (0.22) | 52 | 73 (*) | 0.25 | (0.06) | 0.25 | (0.09) | 12 | 0.67 | (0.12) | 0.69 | (0.17) | 49 | 61 | ||||
| NEUROPTERA | 0.30 | (0.12) | 0.59 | (0.20) | 24 | 0.19 | (0.10) | 0.41 | (0.12) | 16 | 40 (*) | 0.08 | (0.08) | 0.17 | (0.12) | 6 | 0.81 | (0.20) | 0.92 | (0.29) | 62 | 68 | ||||
| PSOCOPTERA | 0.04 | (0.04) | 0.00 | (0.00) | 1 | 0.48 | (0.36) | 0.63 | (0.43) | 30 | 31 | 0.08 | (0.08) | 0.21 | (0.21) | 7 | 0.61 | (0.31) | 0.58 | (0.28) | 43 | 50 | ||||
| THYSANOPTERA | 0.04 | (0.04) | 0.07 | (0.05) | 3 | 0.22 | (0.18) | 0.15 | (0.08) | 10 | 13 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.25 | (0.10) | 0.47 | (0.19) | 26 | 26 | ||||
| COLEOPTERA | 0.26 | (0.11) | 0.37 | (0.22) | 17 | 0.22 | (0.10) | 0.26 | (0.07) | 13 | 30 | 0.50 | (0.28) | 0.50 | (0.22) | 24 | 1.17 | (0.52) | 0.64 | (0.17) | 65 | 89 | ||||
| COLLEMBOLA | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.04 | (0.04) | 0.00 | (0.00) | 1 | 1 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0 | ||||
| OTHER | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.15 | (0.08) | 0.00 | (0.00) | 4 | 4 | 0.00 | (0.00) | 0.00 | (0.00) | 0 | 0.03 | (0.03) | 0.06 | (0.06) | 3 | 3 | ||||
| TOTAL | 7.63 | (1.03) | 9.67 | (2.02) | 467 | 12.19 | (3.32) | 13.93 | (4.22) | 705 | 1172 | 5.79 | (0.99) | 6.33 | (0.94) | 291 | 11.00 | (1.56) | 10.17 | (1.60) | 762 | 1053 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Zamora, J.E.; Gamero-Monge, J.M.; Pérez-de la Luz, R. Fertilization with Olive Mill Pomace Compost Can Moderate Pest Damage in a Superintensive Olive Grove. Agriculture 2024, 14, 2005. https://doi.org/10.3390/agriculture14112005
González-Zamora JE, Gamero-Monge JM, Pérez-de la Luz R. Fertilization with Olive Mill Pomace Compost Can Moderate Pest Damage in a Superintensive Olive Grove. Agriculture. 2024; 14(11):2005. https://doi.org/10.3390/agriculture14112005
Chicago/Turabian StyleGonzález-Zamora, José E., José M. Gamero-Monge, and Rosa Pérez-de la Luz. 2024. "Fertilization with Olive Mill Pomace Compost Can Moderate Pest Damage in a Superintensive Olive Grove" Agriculture 14, no. 11: 2005. https://doi.org/10.3390/agriculture14112005
APA StyleGonzález-Zamora, J. E., Gamero-Monge, J. M., & Pérez-de la Luz, R. (2024). Fertilization with Olive Mill Pomace Compost Can Moderate Pest Damage in a Superintensive Olive Grove. Agriculture, 14(11), 2005. https://doi.org/10.3390/agriculture14112005