Abstract
The yellow spot disease of Pleurotus ostreatus, caused by the bacterium Pseudomonas tolaasii, poses a threat to both the yield and quality of oyster mushrooms due to the limited availability of effective bactericides and the insufficient comprehensive safety assessments for the chemicals applied in the field. In this study, we performed greenhouse toxicity tests to identify highly effective bactericides against P. tolaasii, while simultaneously assessing their safety for P. ostreatus. Subsequently, we verified the field efficacy of the selected bactericide and its impact on mushroom yield through field experiments. Additionally, we established detection methods for the selected highly effective and safe bactericides in P. ostreatus and measured their residue levels during the evaluation. Our findings indicated that kasugamycin and seboctylamine acetate displayed potent inhibitory effects against P. tolaasii. The field application results showed that 1.8% seboctylamine acetate AS with concentrations of 3 mg/L exhibited good disease control, achieving 83.00%, which is comparable to the efficacy of the registered kasugamycin. Furthermore, the application of seboctylamine acetate did not adversely affect the yield of P. ostreatus, and the detected chemical residues were below the maximum residue limit. Seboctylamine acetate is anticipated to serve as a viable substitute for the currently registered fungicides, holding the promise of offering comparable or superior performance while potentially addressing concerns related to environmental impact and sustainability.
1. Introduction
Edible fungi are known for their exceptional taste and considerable nutritional and medicinal benefits [1], and they are enjoyed by people worldwide [2]. The edible fungi industry has uniquely contributed to the restructuring of agricultural industries [3], enhancing agricultural productivity, and promoting circular agriculture practices. Consequently, it has developed into a distinctive and innovative sector within China’s agricultural landscape [4,5]. Pleurotus ostreatus, commonly referred to as the oyster mushroom [6], belongs to the class Basidiomycetes, order Agaricales, family Pleurotaceae, and genus Pleurotus [7]. It is a widely cultivated and commercially significant edible mushroom in many countries [8], with China being a leading producer and exporter [9,10]. Oyster mushrooms are not only delicious, but also rich in nutritional value [11]. They are well-known for their anti-thrombotic [12,13], cholesterol-lowering, anti-aging, and liver-protective properties [5,14,15].
With the steady increase in the diversity of oyster mushrooms and the expansion of their cultivation areas in China, the incidence of oyster mushroom diseases has been rising annually [16]. These diseases include a variety of conditions, such as oyster mushroom white mold, soft rot, yellow spot disease (also known as brown spot disease), and yellow pus disease [17,18,19,20]. Among them, bacterial blotch is one of the most severe bacterial diseases affecting oyster mushrooms [6]. Caused by the pathogen Pseudomonas. tolaasii, it poses a significant threat to both the quality and yield of the mushrooms [21]. Also known as bacterial spot disease, rust spot disease, or brown spot disease, bacterial blotch can develop at any stage of the oyster mushroom fruiting body’s growth [22]. The pathogen specifically targets the surface tissues of the fruiting body. Initially, affected oyster mushroom fruiting bodies may secrete yellow droplets, with yellow to tan lesions appearing on their surfaces [21]. This leads to stunted growth and the development of malformed mature mushrooms. In severe cases, the fruiting bodies may stop growing entirely, resulting in decay and death [23]. If the disease is not addressed promptly, and the management conditions are not improved, it can further affect subsequent mushroom flushes, potentially halting mushroom production altogether.
Currently, strategies for managing diseases in edible mushrooms involve a multifaceted approach, including the breeding of disease-resistant varieties, stringent control over strain quality, careful management of cultivation environments, and the induction and application of disease resistance mechanisms, as well as chemical and biological control methods [24]. Among these strategies, regulating environmental conditions plays a crucial role in mitigating disease severity [25], although it is a challenging task often associated with significant costs. Reports have highlighted the use of biocontrol methods, such as the application of essential oils, antagonistic microorganisms, and specific bacteriophages, for managing bacterial blotch in P. ostreatus [6,26]. Nevertheless, the mechanisms underlying the action of certain biological pesticides remain only partially understood, and their effectiveness in controlling yellow spot disease in oyster mushrooms in field conditions remains inferior to that of chemical fungicides. Consequently, chemical control may continue to be the primary strategy for managing this disease in the foreseeable future [23,27]. To date, no chemical control agents have been identified as particularly effective against brown spot disease in oyster mushrooms. From the 1930s to the 1980s, sodium hypochlorite, commonly in the form of bleaching powder solution, was recommended for the management of bacterial diseases. Additionally, domestic researchers have suggested the use of streptomycin as a control measure [28]. Currently, the primary pesticide formulations used to control brown spot disease in oyster mushrooms include agricultural streptomycin, bleaching powder solution, and bronopol. However, their effectiveness in practical agricultural applications often falls short of expectations [29]. However, according to information from the China Pesticide Information Network, kasugamycin is currently the only registered bactericide used for controlling bacterial blotch in oyster mushrooms. This highlights the urgent need to identify and screen additional chemical bactericides that can effectively combat bacterial blotch in oyster mushrooms.
Seboctylamine acetate is an eco-friendly amino acid polymer bactericide developed in China. It represents a new generation of high-tech, environmentally sustainable, efficient, and broad-spectrum bactericides with low toxicity. The structural formula of seboctylamine acetate can be found in Supplementary Figure S1. The bactericidal mechanism of this bactericide is distinctive, as follows: In an aqueous solution, it undergoes ionization, with its hydrophilic segment acquiring a strong positive charge [30]. This positive charge attracts and adsorbs negatively charged fungi, bacteria, and viruses, effectively inhibiting their reproduction. Additionally, it coagulates the proteins of pathogens and disrupts their enzymatic systems. The polymeric film formed by seboctylamine acetate blocks the ion channels of these microorganisms, causing them to suffocate and perish rapidly, thus achieving optimal bactericidal efficacy. Notably, prolonged use of this bactericide does not lead to resistance and may even promote crop growth [31].
In this study, we conducted indoor toxicity tests to screen for bactericides with high efficacy against the bacterial spot disease of P. ostreatus. Indoor toxicity tests refer to experimental methods conducted under controlled conditions (such as in a laboratory or simulated environment) to evaluate the toxic effects of chemical agents, biological formulations, or other treatments on specific organisms (such as pests, pathogens, or weeds). We investigated the safety of seboctylamine acetate and tetramycin on P. ostreatus and further assessed their field control efficacy against the disease in the mushroom. Field control effects refer to the evaluation of the efficacy of chemical agents, biological formulations, or other management measures in controlling target organisms (such as diseases, pests, or weeds) under actual field conditions. Field efficacy trials aim to validate the practical effectiveness of these treatments in real-world production environments, providing scientific evidence for agricultural practices and broader applications. For oyster mushrooms, which are cultivated using a grid-layer rack system, field control efficacy tests are typically conducted in greenhouse settings. Additionally, we examined the residue levels of seboctylamine acetate in P. ostreatus and its impact on crop yield. The objective of this study is to provide valuable insights and guidance for managing P. ostreatus yellow spot disease and other bacterial diseases in edible mushrooms.
2. Materials and Methods
2.1. Strain for Experimentation and Bactericides
After conducting an investigation into the primary diseases affecting edible fungi in Shandong Province, the Tai’an Academy of Agricultural Sciences isolated, purified, and identified the fruiting bodies of P. ostreatus collected from a mushroom shed in Jining. The test strain was subsequently identified as P. tolaasii and stored at 4 °C for future use. The P. ostreatus strains used in this study were provided by the Tai’an Academy of Agricultural Sciences.
Three bactericides were utilized in this experiment, as follows: 12% zhongshengmycin TC, sourced from Fujian Kaili Bio-Product Co., Ltd. in Zhangzhou, China; 20% seboctylamine acetate AS, acquired from Weifang Voelsing Biopesticide Co., Ltd. in Weifang, China; and 15% tetramycin TC, obtained from Liaoning Micro Biotechnology Co., Ltd. in Chaoyang, China.
For the conduct of this experiment, LB medium and PDA medium were prepared using the following specific formulations:
LB medium: It consisted of 10 g of NaCl, 10 g of peptone, 5 g of yeast extract, and 1 L of distilled water. The pH was adjusted to 7.0, and the medium was sterilized at 121 °C for 30 min.
PDA medium: It was prepared by combining 200 g of potatoes, 20 g of glucose, 20 g of agar, and distilled water to a final volume of 1 L. The natural pH was maintained, and the medium was sterilized at 121 °C for 30 min.
2.2. Isolation and Identification of the Pathogens
To investigate the bacterial pathogens associated with yellow spot disease in P. ostreatus, ten fruiting bodies exhibiting varying degrees of the disease were collected from a mushroom shed located in Jining, China. These fruiting bodies were selected to represent a range of symptoms to ensure diverse pathogen sampling. Each fruiting body was processed under sterile conditions on an ultra-clean bench to minimize the risk of contamination. A 1 cm² section of surface tissue was aseptically excised from each fruiting body using a sterile blade. To remove surface contaminants and non-adherent microorganisms, the tissue samples were washed three times with sterile distilled water. After washing, the samples were individually placed into 10 mL of Luria–Bertani (LB) liquid medium, a general-purpose nutrient medium conducive to bacterial growth. The tubes were incubated at 25 °C with shaking at 160 rpm for 24 h to allow for bacterial enrichment. Following incubation, a 10-fold serial dilution series was prepared from each enrichment culture, diluting the bacterial suspension up to 10⁶-fold. Aliquots (100 µL) from each dilution were plated onto LB agar plates to isolate individual colonies. The plates were incubated at 25 °C under static, dark conditions for 36 h to facilitate bacterial growth. Colonies with distinct morphological characteristics were selected, purified by repeated streaking on fresh LB agar plates, and subsequently stored in 20% (v/v) sterile glycerol at −80 °C in an ultra-low temperature freezer for long-term preservation [32]. From the collected samples, three bacterial strains (designated HF, JPX4-2, and NKY3) were identified as potential pathogens based on their consistent isolation from symptomatic fruiting bodies. These strains were selected for further pathogenicity testing and molecular identification. To confirm the pathogenicity of the isolated strains, the bacteria were activated by culturing in LB liquid medium at 30 °C with shaking at 160 rpm for 24 h. The bacterial cultures were then diluted with sterile distilled water to achieve a final concentration of 1 × 10⁶ colony-forming units (cfu)/mL. Each bacterial suspension (200 µL) was evenly spread onto the surface of healthy, young mushroom pilei under sterile conditions. The inoculated mushrooms were placed in an environment maintained at room temperature (25 °C) and 85% relative humidity for 24 h to promote infection development. After visible symptoms appeared on the fruiting bodies, the pathogens were re-isolated from the symptomatic tissues using the same methods described earlier, confirming Koch’s postulates. The re-isolated strains (HF, JPX4-2, and NKY3) were streaked onto LB agar plates to ensure purity and subjected to further analysis [33]. For morphological identification, single colonies from each strain were analyzed using Gram staining and examined using scanning electron microscopy (SEM) to observe their structural characteristics. Genomic DNA was then extracted from the bacterial isolates using a commercial bacterial DNA extraction kit. The 16S rRNA gene of each strain was amplified using specific primers under the following PCR conditions: an initial denaturation step at 94 °C for 5 min; 32 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 90 s; followed by a final extension at 72 °C for 10 min. The amplified PCR products were sequenced by Sangon Biotech (Shanghai) Co., Ltd. in Shanghai, China. The sequencing data were analyzed using the BLAST program from the NCBI database to determine sequence homology. Based on the BLAST results, preliminary classification and identification of the bacterial strains were performed. The relevant 16S rRNA gene sequences from closely related bacterial species were retrieved from the GenBank database. A phylogenetic tree was constructed using MEGA 7.0 software, employing the neighbor-joining method with 1000 bootstrap replications to ensure the robustness of the analysis. This phylogenetic analysis provided insights into the evolutionary relationships and species-level identification of the isolated strains.
2.3. Toxicity Determination
Using the indoor toxicity determination method, 12% zhongshengmycin, 15% tetramycin, and 20% seboctylamine were dissolved in sterile water and subsequently diluted to create a stock solution with a concentration of 1 × 104 mg/L. This solution was temporarily stored in a refrigerator at 4 °C for future use, as described in [34]. Based on the preliminary experimental findings, seven concentration series were prepared, as follows: seboctylamine concentrations ranged from 1 mg/L to 64 mg/L in gradients of 1, 2, 4, 8, 16, 32, and 64 mg/L; tetramycin concentrations ranged from 0.05 mg/L to 3.2 mg/L in gradients of 0.05, 0.1, 0.2, 0.4, 0.8, 1.6, and 3.2 mg/L; and zhongshengmycin concentrations ranged from 0.5 mg/L to 32 mg/L in gradients of 0.5, 1, 2, 4, 8, 16, and 32 mg/L. These concentrations were prepared by dilution with sterile water, as outlined in [35].
In this study, the K-B paper method was employed to evaluate the antimicrobial activity of the liquid medications. Small, round pieces of qualitative filter paper, each with a diameter of 5 mm, were punched using a standard puncher [36]. These filter paper discs were then sterilized with high-pressure steam and subsequently dried by baking at 165 °C for 2 h to eliminate any potential microbial contamination [37]. Under sterile conditions on an ultra-clean bench, potato dextrose agar (PDA) medium was prepared and poured into Petri dishes, allowing the medium to solidify. After solidification, a bacterial suspension of P. tolaasii was prepared. Specifically, 100 µL of the suspension was pipetted onto the surface of each agar plate, and the plates were allowed to dry completely under the ultra-clean bench to ensure even bacterial distribution and prevent excess moisture. Next, the sterilized filter paper discs were immersed in various concentrations of the liquid medications and sterile water, respectively. After immersion, the filter paper discs were allowed to dry completely before being placed onto the surface of the agar plates that were inoculated with P. tolaasii. This ensured that the filter paper discs contained the desired concentration of the test substances. Each treatment was replicated on three separate plates, with the filter paper discs soaked solely in sterile water serving as the control [38]. Following the placement of the filter paper discs, the agar plates were incubated in a constant-temperature incubator at 28 °C. The growth of the bacteria was monitored over time to observe the formation of inhibition zones around the filter paper discs. Once inhibition zones became clearly visible, the diameters of these zones were measured using a vernier caliper, applying the cross method for accuracy. The average diameter of the inhibition zones was then calculated for each treatment, and the effective concentration that resulted in 50% inhibition (EC50) was determined from the dose–response curve constructed using the measured data [39]. This method enabled a quantitative assessment of the antimicrobial efficacy of each liquid medication.
2.4. Greenhouse Safety Evaluation of Bactericides on P. ostreatus
The safety of the agents was assessed using the mycelial growth rate method. Following the indoor toxicity determination protocol, 12% zhongshengmycin, 15% tetramycin, and 20% seboctylamine were dissolved in sterile water and diluted to create a stock solution with a concentration of 1 × 104 mg/L. This solution was temporarily stored in a refrigerator at 4 °C for future use. For the toxicity tests, specific concentration gradients were prepared for each agent, as follows: tetramycin concentrations were set at 0.4 mg/L, 0.8 mg/L, and 1.6 mg/L; seboctylamine concentrations were 12 mg/L, 24 mg/L, and 48 mg/L; and zhongshengmycin concentrations were 14 mg/L, 28 mg/L, and 56 mg/L [40].
The preparation of the drug-containing medium was carried out by first adding 45 mL of the prepared potato dextrose agar (PDA) medium into a pre-sterilized, dried 100 mL flask. To this flask, 5 mL of the drug solution was added sequentially, starting from the lowest concentration and progressing to the highest concentration. The solution was thoroughly mixed to ensure homogeneity before being poured into three 9 cm diameter Petri dishes. This procedure was followed to ensure consistent distribution of the drug in each Petri dish. To establish a concentration range of the bactericides that would affect the growth of P. ostreatus mycelium, three treatment concentrations were prepared based on the methodology described by Du et al. [41]. These concentrations were selected based on the EC50 value of the pathogen and included the EC50 value itself, twice the EC50 value, and four times the EC50 value. Additionally, blank control plates, which did not contain the drug, were included to serve as negative controls. Each concentration and control was replicated three times to ensure the reproducibility of the results. Under aseptic conditions, actively growing P. ostreatus mycelium was harvested, and 5 mm diameter mycelial discs were excised using a sterilized hole puncher. These mycelial discs were then carefully placed at the center of the prepared drug-containing PDA plates. The plates were sealed and placed in a constant-temperature incubator at 25 ± 1 °C for dark incubation to promote mycelial growth [42]. The growth of the mycelium was monitored, and once the mycelium on the blank control plates had covered approximately three-quarters of the Petri dish, the colony diameter was measured using the cross method. The results were then compared with those of the blank control plates that did not contain the drug. This comparison allowed for the determination of the inhibitory effect of each drug concentration on mycelial growth. The growth inhibition rate of each bactericide on mycelial growth was calculated using the following formula [43]:
Growth inhibition rate (%) = [1 − (diameter of treatment colony − diameter of mycelial disc)/(diameter of control colony − diameter of mycelial disc)] × 100
2.5. Field Control Effects
The field control efficacy experiment was conducted in mushroom sheds at the Shandong Agricultural University’s Resource Utilization Practice Base, with trials carried out in October 2020 and March 2021. The experimental sites were purposefully selected from mushroom sheds that had been continuously used for edible mushroom cultivation, ensuring alignment with local agricultural practices. This selection method guaranteed uniform and consistent cultivation conditions across all experimental sites.
In this study, the cultivation of P. ostreatus (oyster mushroom) was carried out using a substrate composed of fermented materials, following a specific formulation: 60% soybean straw, 28% cottonseed hulls, 5% wheat bran, 2% gypsum, and 5% lime. These raw materials were selected in accordance with the safety technical requirements for edible fungus cultivation substrates outlined in the NY 5099-2002 standards, ensuring that no additional additives were incorporated [44]. The soybean straw was first shredded into pieces approximately 1 cm in length and pre-wetted one day before use, maintaining a material-to-water ratio of 1:1.6 to facilitate fermentation. After the raw materials were thoroughly mixed, they underwent pile fermentation. The fermentation process was initiated by stacking the mixture into piles, and the pile temperature was monitored throughout. Once the temperature of the pile exceeded 60 °C, it was maintained at this level for 24 h to ensure proper pasteurization, after which the pile was turned. This fermentation process lasted a total of nine days, with the pile being turned four times. During the final turn, the test agents were introduced at three different concentrations—0.05%, 0.1%, and 0.2% (based on the dry weight of the culture material). A control treatment (CK) with no test agent was included for comparison. After fermentation, P. ostreatus mycelium was inoculated into polyethylene cultivation bags (23 cm × 35 cm), with each bag containing 1 kg of the prepared dry substrate. The cultivation was carried out under controlled conditions at a constant temperature of 25 °C. To assess the effects of bacterial yellow spot disease, a bacterial suspension of the pathogen strain responsible for the disease in P. ostreatus was prepared. The suspension was adjusted to a concentration of 3 × 10⁸ cfu/mL. Once the fruiting bodies reached a diameter of approximately 1 to 2 cm, 5 mL of the bacterial suspension was evenly sprayed onto the fruiting bodies. A water control group was also established for comparison. The inoculated fruiting bodies were then kept under high humidity for 24 h to allow optimal conditions for pathogen development. The experiment included ten treatments, as follows: 0.3% tetramycin AS with active ingredient dosages of 0.05, 0.08, and 0.11 mg/L; 6% kasugamycin WP with dosages of 0.08, 0.12, and 0.16 mg/L; 1.8% seboctylamine AS with dosages of 1, 2, and 3 mg/L; and a blank control. Each treatment was applied at a rate of 250 mL/m2. Disease incidence was assessed according to the “Pesticide Field Efficacy Test Guidelines,” with data collected on the number of diseased mushrooms in each plot and the overall incidence of bacterial yellow spot disease in P. ostreatus. The bactericide’s control efficacy was then calculated. The harvesting of P. ostreatus began approximately 12 days after treatment application, and the results were analyzed accordingly.
The efficacy calculation method: P = (CK − PT)/CK × 100
- where P = control effect, in percentage (%); CK = number of diseased mushrooms (colonies) in the blank control area; PT = number of diseased mushrooms (colonies) in the agent-treated area.
2.6. Effects of Bactericides on Yield of P. ostreatus
In compliance with the “Pesticides for Crop Safety Evaluation Criteria” (NY/T 1965.1-2010) [45], a bactericide trial was conducted to evaluate the efficacy of various treatments on the control of bacterial yellow spot disease in Pleurotus ostreatus. The dosages of the bactericides were applied at 1, 2, and 4 times that of the recommended field dosage, based on the active ingredient concentration. These treatments were administered at the stage when mushroom buds had just emerged from the covering soil, which is considered a critical phase for pathogen infection. The specific bactericide concentrations were as follows: 0.3% tetramycin aqueous solution at effective ingredient concentrations of 0.05, 0.08, and 0.11 mg/L; 1.8% seboctylamine acetate AS at concentrations of 1, 2, and 3 mg/L; and 6% kasugamycin wettable powder at concentrations of 0.08, 0.12, and 0.16 mg/L. A water-only control treatment, without any bactericide, was included to assess the baseline level of disease incidence. All treatments were applied at a uniform rate of 250 mL per square meter, ensuring consistent coverage across the experimental plots. The experimental design followed a randomized block arrangement, with each treatment being replicated three times. Each experimental plot covered an area of 2 m², allowing for sufficient space to monitor the treatment effects. Data collection was conducted using the Five-Point Sampling Method, which involved randomly selecting 10 fruiting bodies from each experimental plot. These fruiting bodies were then assessed for signs of bacterial yellow spot disease, and disease incidence was recorded. The outcomes were evaluated 12 days after the initial treatment application to determine the bactericide’s efficacy in controlling the disease.
2.7. Drug Residue Testing
The residues of seboctylamine acetate and kasugamycin in P. ostreatus (oyster mushrooms) were analyzed using liquid chromatography. The mushrooms were cultivated in the mushroom shed at the Mushroom Resources and Utilization Training Base of Shandong Agricultural University. After the mushrooms were covered with soil, they were sprayed with the bactericide treatments three days later. The experiment consisted of the following three treatments: a 6% kasugamycin wettable powder (WP) with an active ingredient dosage of 43.2 mg/L, 1.8% seboctylamine acetate AS with an effective component dosage of 216 mg/L, and a blank control. Each treatment was applied at a rate of 250 mL per square meter of cultivation area. The experimental plots were arranged in a randomized block design, with three replicates per treatment, and each plot covered an area of 2 m². This design ensured the validity of comparisons between treatments. Fresh samples of P. ostreatus were collected at specific time intervals following application, including 2, 12, 24, 30, 36, and 48 h after spraying. A minimum of 1 kg of fresh mushrooms was harvested from each plot at each time point. The mushrooms were then chopped and divided into two equal portions of 150 g each, which were promptly stored in a low-temperature refrigerator to preserve their integrity until analysis. The residual dynamics and final concentrations of kasugamycin and seboctylamine acetate in P. ostreatus were determined using liquid chromatography, following established analytical methods [46,47]. This enabled the assessment of how these chemicals were metabolized or degraded over time in the mushroom fruiting bodies.
Pretreatment: For the residue analysis of kasugamycin and seboctylamine acetate in P. ostreatus, the following sample preparation procedure was employed: A 5 g sample of fresh P. ostreatus was weighed accurately and placed into a 50 mL plastic centrifuge tube. To the sample, 25 mL of acetonitrile was added as the extraction solvent. The mixture was then subjected to oscillation for 5 min to ensure thorough mixing and initial extraction of the residues. Following the initial extraction, a salt packet containing sodium chloride (NaCl) and other additives was added to the mixture to facilitate phase separation and improve extraction efficiency. The solution was oscillated for an additional 5 min to ensure proper dissolution and mixing of the salts. After mixing, the sample was centrifuged at 4000 rpm for 5 min to separate the solid particulates from the liquid phase. The supernatant was carefully collected and filtered through a 0.45 μm membrane filter to remove any remaining particulates, ensuring a clean sample for subsequent analysis. The filtered supernatant was then ready for injection into the liquid chromatography system for the determination of kasugamycin and seboctylamine acetate residues.
Chromatographic conditions: A Poroshell 120 EC-C18 column was used, with the column temperature set at 40 °C. The injection volume was 5.0 μL, and the flow rate was 0.8 mL/min. Mobile phase A consisted of 0.1% formic acid in water (pH ≈ 3.5), while mobile phase B was acetonitrile.
The mass spectrometer parameters were as follows: the ion source was an electrospray ionization (ESI) source, operating in positive ion scanning mode with multiple reaction monitoring (MRM) as the mass spectrometry scanning method. The ion source temperature (TEM) was set to 600 °C, and the ion spray voltage (IS) was 4500 V. The curtain gas (CUR) flow rate was 30 psi, the nebulizer gas (Gas1) flow rate was 55 psi, the auxiliary gas (Gas2) flow rate was 60 psi, and the collision gas (CAD) pressure was 7 psi. Additional mass spectrometry parameters are provided in Table 1.

Table 1.
Residual test parameters.
2.8. Statistical Analysis
SPSS 25.0 software was used to calculate the biological activity regression equation, EC50 value, and 95% confidence intervals for the different bactericides against bacterial macular disease in P. ostreatus. For the safety evaluation and field efficacy tests, Duncan’s multiple range test was applied to compare significant differences between treatments in each test. A p-value of < 0.05 was considered statistically significant, and the data were expressed as mean ± standard deviation.
3. Results
3.1. Molecular Identification of Pathogens and Construction of Phylogenetic Tree
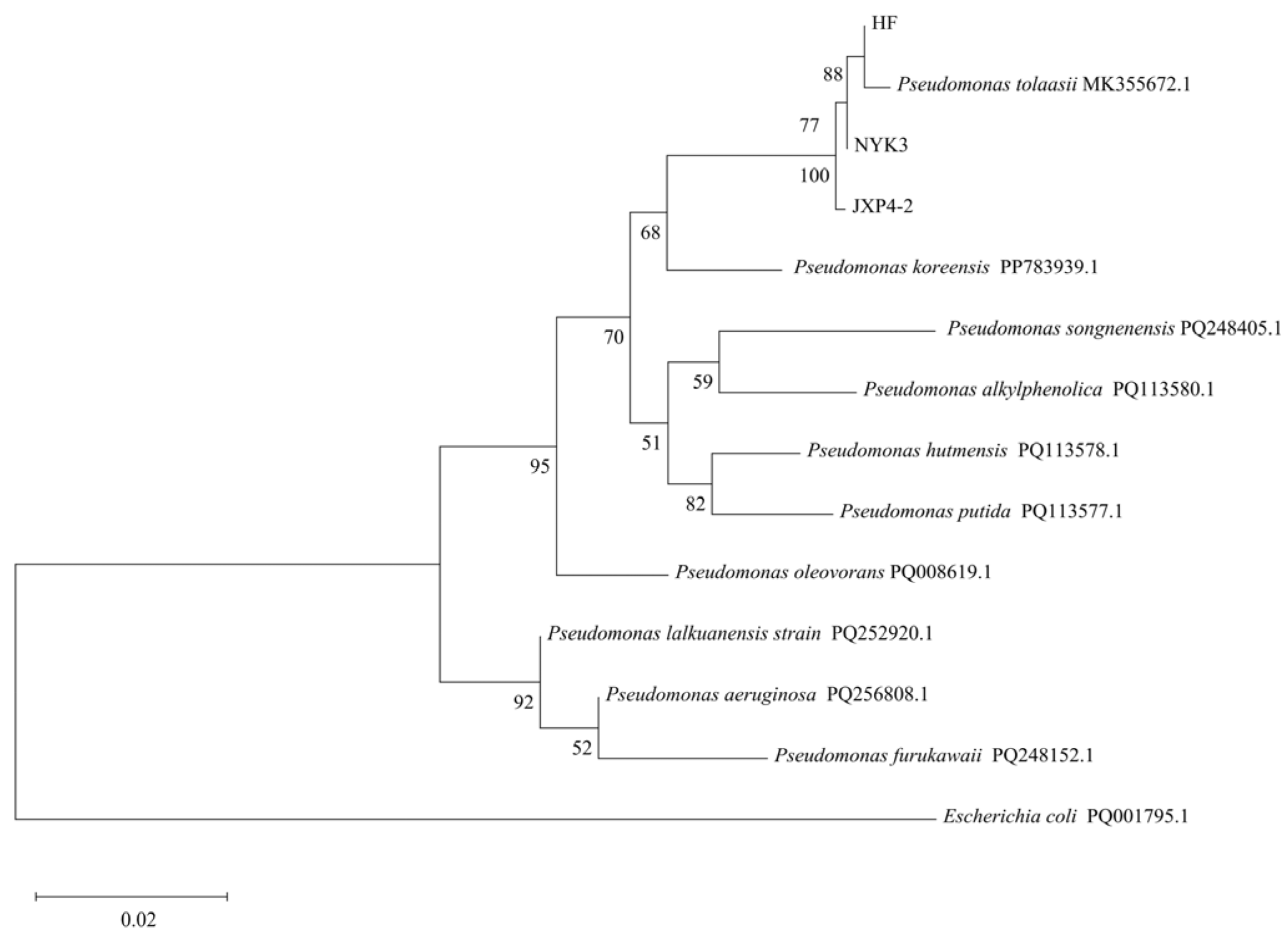
The initial isolates of the pathogen strains HF, JPX4-2, and NKY3, responsible for yellow blotch disease in oyster mushrooms, were inoculated onto the caps of healthy oyster mushrooms. After 24 h of incubation at 25 °C and 85% relative humidity, typical symptoms of yellow blotch disease appeared on the mushroom caps as irregular yellow spots. These results were consistent with the symptoms observed in conventional cultivation settings, indicating that all three pathogen strains possess significant pathogenicity, causing yellow blotches on oyster mushroom fruiting bodies. Further examination with scanning electron microscopy revealed that the pathogen cells were short-rod shaped, consistent with the morphology of the original infectious agent. BLAST comparison revealed that the 16S rRNA sequences of strains HF, JPX4-2, and NKY3 exhibited high coverage and similarity to P. tolaasii 135 (HK21), reaching 100%. As shown in Figure 1, in the 16S rRNA gene phylogenetic tree, strains HF, JPX4-2, and NKY3 were distantly related to E. coli and clustered together on the same branch as P. tolaasii 135 (HK21). Based on these findings, strains HF, JPX4-2, and NKY3 were identified as P. tolaasii.

Figure 1.
Phylogenetic analysis based on sequencing of the 16S ribosomal RNA gene.
3.2. The Toxicity of Bactericides on P. tolaasii
This experiment utilized the K-B paper disc method to assess the indoor toxicity of various bactericides against P. tolaasii strains HF, JPX4-2, and NKY3. Through screening, this study identified bactericides with relatively superior efficacy. The results showed that seboctylamine acetate, tetramycin, and zhongshengmycin exhibited stronger inhibitory effects on P. tolaasii HF. Among these, tetramycin demonstrated the most potent bacteriostatic effect, with an EC50 value of 0.395 mg/L, followed by seboctylamine acetate (EC50 = 12.014 mg/L) and zhongshengmycin (EC50 = 14.033 mg/L) (Table 2). However, zhongshengmycin demonstrated limited inhibitory effects on strains JPX4-2 and NKY3, with EC50 values of 52.731 mg/L and 32.098 mg/L, respectively. Tetramycin showed a strong inhibitory effect on P. tolaasii strain JPX4-2, with an EC50 value of 0.395 mg/L, while seboctylamine acetate exhibited an EC50 value of 12.035 mg/L against this strain (Table 3). For P. tolaasii strain NKY3, tetramycin exhibited an EC50 value of 0.398 mg/L, and seboctylamine acetate had an EC50 value of 11.942 mg/L (Table 4). Additionally, the indoor toxicity of other bactericides against the pathogen P. tolaasii was also assessed, but the antibacterial effects were not satisfactory. Specifically, 1000 mg/L concentrations of copper, copper acetate, ethylicin, and copper hydroxide did not exhibit a significant inhibitory effect on P. tolaasii. Additionally, the antibacterial effects of 64 mg/L ethylicin and 100 mg/L zinc thiazole on P. tolaasii were minimal.
Table 2.
Toxicity of bactericides to P. tolaasii (HF).
Table 3.
Toxicity of bactericides to P. tolaasii (JPX4-2).
Table 4.
Toxicity of bactericides to P. tolaasii (NKY3).
3.3. Greenhouse Safety Test of Test Agents on P. ostreatus
Based on the results from the indoor toxicity tests, seboctylamine acetate, tetramycin, and zhongshengmycin were selected for further experiments. The concentration gradients for each bactericide against P. tolaasii were set at EC50, 2EC50, and 4EC50, with the specific concentrations listed in Table 5. The results indicated that all three concentrations of zhongshengmycin significantly inhibited the growth of P. ostreatus mycelium, which posed a risk to its growth. This finding suggests that zhongshengmycin is unsuitable for use on this pathogen. Additionally, the three concentrations of tetramycin exhibited a certain, albeit weaker, inhibitory effect on P. ostreatus mycelium. In contrast, the three concentrations of seboctylamine acetate showed no significant inhibitory effect on the mycelium of P. ostreatus 62 but displayed a mild inhibitory effect on the mycelium of P. ostreatus 129 and P. ostreatus 28. Notably, no necrosis, deformities, or other signs of drug-induced damage were observed. The relevant experimental images can be found in Supplementary Figure S2. Therefore, the application of seboctylamine acetate is considered safe for the growth of P. ostreatus.
Table 5.
Indoor safety evaluation of three bactericides on P. ostreatus.
3.4. Field Efficacy of the Tested Agents Against Yellow Spot Disease in P. ostreatus
The findings reveal that, across the three concentrations tested, tetramycin, seboctylamine acetate, and kasugamycin exhibited significant variations in the quality of oyster mushroom fruiting bodies compared to the control, consistently promoting an increase in yield. The influence of the tested agents on the yield of P. ostreatus is detailed in Table 6 and Table 7. Notably, no inhibitory effects were observed, and there were no signs of necrosis, deformities, or other phytotoxic symptoms. The field control efficacy of the tested agents against yellow spot disease in P. ostreatus is shown in Table 8 and Table 9. The results indicated that seboctylamine acetate and kasugamycin demonstrated equivalent control efficacy against the disease. In October 2020, the highest control efficacy, 84.00%, was observed at a concentration of 0.16 mg/L. Seboctylamine acetate at a concentration of 3 mg/L also exhibited notable efficacy, with a control rate of 83.00%. Although tetramycin’s field control performance was relatively lower, it still achieved a control rate of 30.67% at a concentration of 0.11 mg/L. In March 2021, kasugamycin at 0.16 mg/L provided the best control efficacy, reaching 81.87%, while seboctylamine acetate at 3 mg/L performed similarly with a control effectiveness of 82.11%. Despite the relatively lower field performance of tetramycin, it achieved a control rate of 33.77% at a concentration of 0.11 mg/L.
Table 6.
Effects of three bactericides on the yield of P. ostreatus in 2020.
Table 7.
Effects of three bactericides on the yield of P. ostreatus in 2021.
Table 8.
The field control effects of three bactericides on P. ostreatus yellow spot disease in 2020.
Table 9.
The field control effects of three bactericides on P. ostreatus yellow spot disease in 2021.
3.5. The Impact of the Tested Agents on the Yield of P. ostreatus
As shown in Table 6 and Table 7, 0.05, 0.08, and 0.11 mg/L of tetramycin; 1, 2, and 3 mg/L of seboctylamine acetate; and 0.08, 0.12, and 0.16 mg/L of kasugamycin had significant differences in the yield of P. ostreatus, which promoted the yield increase in P. ostreatus. No necrosis or deformity symptoms were found in the treatment of the tested agents or the control group.
3.6. Effects of Kasugamycin and Seboctylamine Acetate on Residues in P. ostreatus
Bactericide pesticide residues have become a significant concern regarding the food safety of agricultural products. The illegal and excessive use of bactericides in pest management poses a serious threat to human health [48]. To date, there has been limited research on the investigation and monitoring of bactericide pesticide residues in edible fungi in other countries. The experimental results for determining bactericide residues indicate that kasugamycin and seboctylamine acetate decompose relatively quickly in the P. ostreatus fruiting body. Specifically, when the active ingredient dosages of 1.8% seboctylamine acetate AS and 6% kasugamycin WP were set at 216 mg/L and 43.2 mg/L, respectively, the final residues measured 0.033 mg/kg and 0.045 mg/kg after 48 h of water application at a rate of 250 mL/m2, as shown in Figure 2. According to the Maximum Residue Limits for Bactericides in Food Safety National Standards (GB 2763-2021) [49], while no specific maximum residue limit for seboctylamine acetate in mushrooms is provided, the limit for vegetables is set at 0.5 mg/kg. Importantly, the results of this experiment showed that the residue levels were below this threshold, indicating that seboctylamine acetate can be safely used on P. ostreatus.
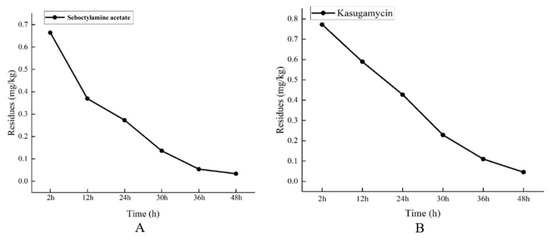
Figure 2.
Digestion curve of chemical agents in P. ostreatus fruiting body. (A) Digestion curves of seboctylamine acetate in the fruiting body of P. ostreatus at 216 mg/L; (B) Digestion curves of kasugamycin in the fruiting body of P. ostreatus at 43.2 mg/L.
4. Discussion
The growth of the mushroom industry is closely tied to the increasing consumer demand for healthy diets, with mushrooms emerging as a prominent category of edible fungi [50]. However, as the oyster mushroom industry expands and production methods advance, disease-related challenges have increasingly affected the sector, hindering its growth [51]. P. ostreatus, a crucial species in mushroom production, is particularly vulnerable to a variety of diseases, including fungal, bacterial, and viral infections [24]. These diseases not only stunt the growth and yield of P. ostreatus, but can also compromise the quality and safety of the final product [52]. Among these, bacterial yellow spot disease, caused by P. tolaasii, represents one of the most severe bacterial threats to oyster mushrooms, jeopardizing both their quality and yield [16]. Currently, chemical control measures are predominantly used to manage these diseases in P. ostreatus. However, the range of available bactericides is limited, and the safety of these chemicals in practical applications has not been thoroughly evaluated. For example, Nurziya et al. found that 72% agricultural streptomycin effectively inhibited the indoor growth of P. tolaasii, with an EC50 of 0.04 mg/L. Although new bactericide formulations offer a promising alternative, their residual efficacy requires careful evaluation [53]. According to the China Pesticide Information Network, kasugamycin is currently the only registered bactericide used for controlling bacterial yellow spot disease in oyster mushrooms. In this study, we confirmed kasugamycin’s inhibitory effect on bacterial yellow spot disease in P. ostreatus through indoor virulence tests. The results revealed an EC50 value of 0.104 mg/L. However, an over-reliance on a single agent can lead to the development of resistance [54]. To mitigate the risk of resistance, we also assessed the inhibitory effects of seven additional chemical bactericides on P. tolaasii. Notably, seboctylamine acetate, a domestically developed product, demonstrated a strong inhibitory effect on P. tolaasii, with an EC50 value of 11.942 mg/L. This finding presents a promising alternative for the future prevention and control of bacterial yellow spot disease in oyster mushrooms.
When applying bactericides to control diseases in edible fungi, several factors must be carefully considered, particularly the complex interactions between the edible fungi and the pathogens. Notably, while these bactericides effectively eliminate pathogens, their toxicity to the host organisms should be minimal [24]. As a result, when using chemical agents to manage pathogens, it is essential to assess their potential harm to P. ostreatus strains and ensure that such harm is minimized or eliminated [55]. Zhu and colleagues have demonstrated that carbendazim has a relatively weak inhibitory effect on the growth of P. ostreatus. Specifically, at a concentration of 10 mg/L, carbendazim inhibits the growth of Trichoderma by 97.80%, while its inhibitory effect on P. ostreatus is only 52.08% [56]. Therefore, carbendazim can be safely included as a component in the production of P. ostreatus. Ye conducted a study to investigate the inhibitory effects of four biological bactericides on the mycelial growth of Neurospora and three common edible fungi. The results indicated that zhongshengmycin, at a concentration of 40 mg/kg, completely inhibited the mycelial growth of Neurospora, but had minimal inhibitory effects (less than 10%) on the mycelial growth of edible fungi [57]. Our findings are consistent with those of Zhu and Ye et al., as we observed that the application of seboctylamine acetate had no detrimental effects on the normal growth of P. ostreatus. Field experiments further revealed that, at a concentration of 3 mg/L, seboctylamine acetate achieved a control efficacy of 82.11% without compromising the yield of P. ostreatus. Additionally, our results showed that tetramycin exhibited high toxicity towards P. tolaasii, the causative agent of bacterial yellow spot disease in P. ostreatus. However, its field control effectiveness was unsatisfactory, rendering it unsuitable for managing this disease.
Seboctylamine is a lipophilic macromolecular compound and a broad-spectrum bactericide. Initially introduced in 1990 as a disinfectant and bactericide in agriculture, it was designed to protect plants from viral and bacterial infections. Seboctylamine acetate has been approved by the pesticide standardization management authority of the People’s Republic of China for production and is typically formulated as 1.8% AS [58]. It is registered in China for use on various crops, including apples, peppers, tomatoes, cotton, tobacco, and rice [31], and is effective against a range of diseases caused by fungi, bacteria, and viruses [59,60]. Due to its unique bactericidal mechanism, seboctylamine acetate has a low risk of resistance development with long-term use and may even promote crop growth. Research by Zang et al. has demonstrated that seboctylamine acid is particularly effective against bacterial pathogens, exhibiting highly specific toxicity toward Xanthomonas species. Its bactericidal action is achieved through a novel mechanism involving interference with cellular iron metabolism, making it an ideal choice for controlling bacterial diseases [61]. Jin et al. reported that seboctylamine inhibits ATP synthesis by disrupting bacterial oxidative phosphorylation and TCA cycle pathways, thereby achieving its antibacterial effect. As an eco-friendly amino acid polymer bactericide developed in China, seboctylamine acetate shows great market potential [62]. This study confirmed that seboctylamine acetate effectively controls yellow spot disease in oyster mushrooms and is safe for mushroom growth, indicating its potential as a viable alternative to existing bactericides with similar or superior performance. While elucidating its antibacterial mechanism is crucial for the efficient and safe use of this compound, unfortunately, this paper does not delve into its specific mode of action, which requires further investigation in future studies. Another notable advantage of seboctylamine acetate is its environmental friendliness. Jin et al. studied the residue of seboctylamine in tobacco and soil, showing that it degrades rapidly in both under field conditions. Jin et al. further examined the factors influencing its degradation in soil under laboratory conditions, finding that the rate of degradation is directly related to the organic matter and moisture content in the soil [63]. Unfortunately, there is limited research on the toxicity of seboctylamine acetate to non-target organisms. However, to comprehensively assess its environmental safety, future studies should focus on the impact of seboctylamine acetate on soil microbial communities, plant growth, and aquatic ecosystems. Furthermore, given its potential for broad application, it is also necessary to conduct in-depth research on its effectiveness against different crops and pathogens. Such studies would enhance our understanding of the environmental behavior and ecological risks of seboctylamine acetate, thereby providing a scientific basis for its rational use and management.
In the field of food safety, the issue of pesticide residues has garnered widespread global attention. Due to their susceptibility to environmental influences during growth, edible mushrooms are particularly prone to pesticide absorption and accumulation. Currently, there is a significant shortage of specialized agents for the prevention and control of diseases in edible mushrooms, with heavy reliance on formulations designed for vegetables and major crops [64]. Research by Wen et al. revealed the prevalence of multiple pesticide residues in edible mushroom samples from the Beijing area. Among these samples, Agrocybe aegerita had a pesticide detection rate as high as 85.7%, while P. ostreatus followed closely with a detection rate of 26.9%. Among the detected pesticides, the insecticide methomyl and the fungicide carbendazim had the highest detection rates [65]. Considering the rapid growth characteristics of P. ostreatus, the extensive use of chemical bactericides for disease control can easily lead to excessive residue levels, thus posing food safety risks. Given the importance of addressing bactericide residues on oyster mushrooms, this study underscores the necessity of research in this area. As a novel bactericide independently developed in China, studies on the environmental behavior of seboctylamine acetate remain relatively limited both domestically and internationally. Jin’s research on seboctylamine acetate residues in tobacco found that the residue levels at harvest ranged from <0.05 to 0.71 mg/kg, with a median value of 0.08 mg/kg, confirming the safety of the harvested tobacco samples [63]. Seboctylamine acetate, known for its remarkable efficacy, broad-spectrum activity, and relatively low toxicity, is widely used in the protection of various crops. This compound effectively inhibits the growth of a variety of plant pathogens while being safe for the crops themselves, without causing phytotoxicity [66]. However, there are few reports on methods for detecting seboctylamine acetate residues [67]. Han et al. developed a method for determining pesticide residues of seboctylamine acetate in apples, cottonseed, peppers, tomatoes, grapes, and pears using ultra-high-performance liquid chromatography–tandem mass spectrometry (UPLC-MS/MS). This method employs a modified QuEChERS technique for sample purification and is analyzed using multiple reaction monitoring (MRM) mode methods. Acetonitrile and 1.0 mmol/L ammonium acetate containing 0.1% formic acid were used as the mobile phase. Quantitative analysis was conducted using the external standard method, yielding average recoveries ranging from 75.6% to 106.2%, with relative standard deviations (RSD) not exceeding 8.8%. The limits of quantification (LOQs) were determined to be 2.0 µg/kg for apples, peppers, tomatoes, grapes, and pears, and 10.0 µg/kg for cottonseed [68]. Although this agent exhibits significant inhibitory effects on yellow spot disease in oyster mushrooms, its residue levels on P. ostreatus remain unclear. The residue analysis method proposed by Han is specifically designed for plant-based foods and is, therefore, not directly applicable to oyster mushrooms, presenting certain limitations in its use. In response, this study developed a new analytical method for determining bactericide residues on edible mushrooms. We established a method to measure seboctylamine acetate residue levels, and our experimental results indicated that, with active ingredient doses of 1.8% seboctylamine acetate AS and 6% kasugamycin WP, set at 216 mg/L and 43.2 mg/L, respectively, the deposition reached 250 mL/m2 within 48 h. The final residue concentrations were 0.033 mg/kg and 0.045 mg/kg, respectively, both below the maximum residue limits established for P. ostreatus, demonstrating that their application on oyster mushrooms is safe.
5. Conclusions
In summary, this study has identified an effective and safe bactericide for the prevention and management of yellow spot disease in P. ostreatus, providing a foundation for the development of a residue detection method for seboctylamine acetate. This work also contributes to the potential future registration of the drug. The findings revealed that seboctylamine acetate exhibited superior antibacterial activity against P. tolaasii compared to the other compounds tested. Notably, the three concentrations of seboctylamine acetate did not significantly inhibit the mycelial growth of P. ostreatus. At a concentration of 3 mg/L, seboctylamine acetate demonstrated strong field control efficacy, achieving 83.00% and 82.11% in different trials, without adversely affecting the growth of P. ostreatus fruiting bodies. The residue analysis of seboctylamine acetate in P. ostreatus revealed relatively low levels, further supporting its suitability for use in controlling yellow spot disease in oyster mushrooms.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14122352/s1, Figure S1: The structural formula of Seboctylamine acetate. Figure S2: The safety test of Pleurotus ostreatus.
Author Contributions
Conceptualization, H.W.; Data curation, Y.Y., H.F. and Q.C.; Formal analysis, Y.Y. and Q.C.; Funding acquisition, Y.L. and H.W.; Investigation, Y.Y., H.F., Y.L., and X.L.; Methodology, Y.Y.; Project administration, H.W. and Y.L.; Resources, Y.L. and H.W.; Software, Y.Y. and H.F.; Supervision, H.W.; Visualization, H.W.; Writing—original draft, Y.Y. and H.W.; Writing—review and editing, Y.L. and H.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key R&D Program of China (2023YFD1401200) and the Modern Agricultural Technology Industry System of Shandong Province (SDAIT-07-04).
Institutional Review Board Statement
No applicable.
Informed Consent Statement
No applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hassan, E.; Neama, A.; Khandsuren, B.; Xhensila, L.; Gréta, T.; Peter, H.; Yahya, E.; József, P. Edible mushrooms for sustainable and healthy human food: Nutritional and medicinal attributes. Sustainability 2022, 14, 4941. [Google Scholar] [CrossRef]
- Sganzerla, W.G.; Todorov, S.D.; Da, S.A.P.G. Research Trends in the study of edible mushroom: Nutritional properties and health benefits. Int. J. Med. Mushrooms 2022, 24, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Fu, Y.; Shi, M.; Wang, H.; Guo, J.Z. Pollution level and risk assessment of lead, cadmium, mercury, and arsenic in edible mushrooms from Jilin Province, China. J. Food Sci. 2021, 86, 3374–3383. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Yu, L.L.; Zhao, J.X.; Zhang, H.; Chen, W.; Zhai, Q.X. Role of dietary edible mushrooms in the modulation of gut microbiota. J. Funct. Foods 2021, 83, 104538. [Google Scholar] [CrossRef]
- Muszynska, B.; Grzywacz-Kisielewska, A.; Kala, K.; Gdula-Argasinska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Hu, D.D.; Gu, J.G.; Zuo, X.M.; Hu, Q.X.; Zhang, J.X. Evaluation of oyster mushroom strains for resistance to Pseudomonas tolaasii by inoculation in spawned substrates. Eur. J. Plant Pathol. 2013, 137, 119–126. [Google Scholar] [CrossRef]
- Piska, K.; Sulkowska-Ziaja, K.; Muszynska, B. Edible mushroom Pleurotus ostreatus (oyster mushroom)—ITS dietary significance and biological activity. Acta. Sci. Pol-Hortoru. 2017, 16, 151–161. [Google Scholar]
- Sang, Y.M.; Han, X.Z.; Gao, Y.C.; Hao, Z.T.; Zeng, Y.; Dai, S.S.; Xue, H.K. Research progress on biological activities of polysaccharides from edible fungus. Starch-Starke 2023, 75, 2200219. [Google Scholar] [CrossRef]
- Zhang, Y.R.; Wang, D.W.; Chen, Y.T.; Liu, T.T.; Zhang, S.S.; Fan, H.X.; Liu, H.C.; Li, Y. Healthy function and high valued utilization of edible fungi. Food Sci. Hum. Well 2021, 10, 408–420. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macro fungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Sánchez, C. Cultivation of Pleurotus ostreatus and other edible mushrooms. Appl. Microbiol. Biot. 2010, 85, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.X.; Du, H.J.; Hu, Q.H.; Yang, W.J.; Pei, F.; Xiao, H. Health benefits of edible mushroom polysaccharides and associated gut microbiota regulation. Crit. Rev. Food Sci. 2021, 62, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.T.; Pan, C.H.; Chien, Y.W.; Huang, H.Y. Edible mushrooms: Novel medicinal agents to combat metabolic syndrome and associated diseases. Curr. Pharm. Des. 2020, 26, 4970–4981. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Joshi, P.B.; Ahmad, Z.; Hemeg, H.A.; Olatunde, A.; Naz, S.; Hafeez, N.; Simal-Gandara, J. Edible mushrooms as potential functional foods in amelioration of hypertension. Phytother. Res. 2023, 37, 2644–2660. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B.J. Anti-obesity effects of medicinal and edible mushrooms. Molecules 2018, 23, 2880. [Google Scholar] [CrossRef]
- Hu, H.J.; Wang, J.R.; Cheng, X.H.; Liu, Y.; Zhang, X.Y. Preliminary studies on the effects of oyster mushroom spherical virus China strain on the mycelial growth and fruiting body yield of the edible mushroom Pleurotus ostreatus. Biology 2022, 11, 574. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Bellettini, S.; Fiorda, F.A.; Pedro, A.C.; Bach, F.; Fabela-Morón, M.F.; Hoffmann-Ribani, R. Diseases and pests noxious to Pleurotus spp. mushroom crops. Rev. Argent. Microbiol. 2018, 50, 216–226. [Google Scholar] [CrossRef]
- Radványi, D.; Gere, A.; Jókai, Z.; Fodor, P. Rapid evaluation technique to differentiate mushroom disease-related moulds by detecting microbial volatile organic compounds using HS-SPME-GC-MS. Anal. Bioanal. Chem. 2015, 407, 537–545. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, Y.; Xie, J.B.; Zhao, S.; Qin, W.T.; Song, Q.G.; Wang, S.X.; Rong, C.B. Bacterial infection induces ultrastructural and transcriptional changes in the king oyster mushroom (Pleurotus eryngii). Microbiol. Spectr. 2022, 10, e0144522. [Google Scholar] [CrossRef]
- Lee, C.J.; Yun, H.S.; Jhune, C.S.; Cheong, J.C.; Yoo, Y.B. Occurrence of bacterial soft rot of Pleurotus ostreatus caused by burkholderia gladiolipv. agaricicolain korea. J. Plant Pathol. 2010, 92, 235–240. [Google Scholar]
- Sajben, E.; Manczinger, L.; Nagy, A.; Kredics, L.; Vágvölgyi, C. Characterization of pseudomonads isolated from decaying sporocarps of oyster mushroom. Microbiol. Res. 2010, 166, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.P.; Iacobellis, N.S. Characterization of fluorescent pseudomonads responsible for the yellowing of oyster mushroom (Pleurotus ostreatus). Phytopathol. Mediterr. 2014, 53, 54–65. [Google Scholar]
- Xiao, Y.; Chen, K.; Liu, K.F.; Wei, Y.C.; Ma, X.L.; Bian, Y.B. Pathogens and symptom diversity of yellow blotch disease in Pleurotus ostreatus. Acta Edulis Fungi 2021, 28, 159–166. [Google Scholar]
- Gea, F.J.; Navarro, M.J.; Suz, L.M. Cobweb disease on oyster culinary-medicinal mushroom (Pleurotus ostreatus) caused by the mycoparasite Cladobotryum mycophilum. J. Plant Pathol. 2019, 101, 349–354. [Google Scholar] [CrossRef]
- Chun, S.C.; Ahn, Y.N.; Khan, S.M.; Chung, I.M.; Won, H.Y.; Jhune, C.S.; Park, Y.J. The microbial population in the air of cultivation facility of oyster mushrooms. J. Microbiol. 2012, 50, 1053–1057. [Google Scholar] [CrossRef]
- Tajalipour, S.; Hassanzadeh, N.; Khabbaz Jolfaee, H.; Heydari, A.; Ghasemi, A. Biological control of mushroom brown blotch disease using antagonistic bacteria. Biocontrol Sci. Technol. 2014, 24, 473–484. [Google Scholar] [CrossRef]
- Huang, P.J. Varieties Resistance and Bactericide Screening to Pseudomonas tolaasii in Pleurostus ostrestus. Master’s Thesis, Huazhong Agricuture University, Wuhan, China, 2012. [Google Scholar]
- Liu, K.J. Research progress on edible fungus diseases. World Agric. 1991, 5, 32–34. [Google Scholar]
- Zhang, R.D.; Hu, D.D.; Zuo, X.M.; Wang, H.X.; Jiang, R.B. Research progress on bacterial brown spot disease of oyster mushroom and button mushroom. J. Plant Prot. 2007, 5, 549–554. [Google Scholar]
- Li, Q.; Chen, S.S.; Zhang, S.R.; Li, C.J.; Zhou, J.Z.; Ma, X.D.; Li, X.F. Bioconcentration study of Xinjunan in zebrafish. Environ. Monit. Assess. 2011, 183, 113–120. [Google Scholar] [CrossRef]
- Li, L.K.; Yuan, S.K.; Yan, Q.P.; Cui, X.L.; Jiang, H. Study on the anti-microbial activity of Xinjunan to 15 plant pathogens. Pestic. Sci. Manag. 2010, 31, 32–34. [Google Scholar]
- Zhao, H.; Liu, J.Y.; Chang, M.C.; Meng, J.L.; Zhang, M.L. Identification of a new pathogen of the brown spot disease of rough-skin oyster mushroom and its biological control. J. Edible Fungi 2019, 26, 113–118+8. [Google Scholar]
- Zhao, H. Study on Isolation, Identification, Infection Mechanism, and Biological Control of the Pathogen of Brown Spot Disease in Oyster Mushroom. Master’s Thesis, Shanxi Agricuture University, Jinzhong, China, 2019. [Google Scholar]
- Xue, T.Z.; Kai, L.M.; Ming, Y.S.; Jin, M.Z.; Li, J.L.; Shi, Q.N. Isolation and identification of pathogens of Morchella sextelata bacterial disease. Front. Microbiol. 2023, 14, 1231353. [Google Scholar]
- Alamer, A.S.I.; Tomah, A.A.; Li, B.; Zhang, J.Z. Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China. Agriculture 2020, 10, 37. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.L.; Ma, L.G.; Wang, P.; Li, C.S.; Qi, J.S. Identification of Pythium species as pathogens of garlic root rot. J. Plant Pathol. 2021, 103, 259–267. [Google Scholar] [CrossRef]
- Liu, Z.L.; Wu, S.J.; Chen, X.F. Biological properties of pathogen yellow spot disease in Pleurotus pulmonarius and toxicity test of different fungicides in laboratory. Edible Fungi China 2024, 43, 67–72. [Google Scholar]
- Chen, Q.Q.; Qiu, Y.; Yuan, Y.Z.; Wang, K.Y.; Wang, H.Y. Biocontrol activity and action mechanism of Bacillus velezensis strain SDTB038 against Fusarium crown and root rot of tomato. Front. Microbiol. 2022, 13, 994716. [Google Scholar] [CrossRef]
- Niu, J.J. Study on Pesticides Control of the Bacterial Disease of Mushroom. Master’s Thesis, Hebei University of Science Technology, Shijiazhuang, China, 2014. [Google Scholar]
- Qiu, Y.; Yan, H.H.; Sun, S.M.; Wang, Y.Q.; Zhao, X.R.; Wang, H.Y. Use of Bacillus velezensis SDTB022 against tobacco black shank (TBS) and the biochemical mechanism involved. Biol. Control 2022, 165, 104785. [Google Scholar] [CrossRef]
- Du, Y.X.; Zhu, C.Y.; Ruan, H.C. Control efficacy of seven fungicides against Mycogone perniciosa and their safety evaluation to Agaricus bisporus. Chin. J. Pestic. Sci. 2013, 15, 635–642. [Google Scholar]
- Zhang, D.; Sun, X.L.; Ma, X.Y. Inhibition of fungicide on sclerotinias clerotiorum of cucumber at different growth stages and field control effects. Pak. J. Bot. 2023, 55, 1175–1179. [Google Scholar] [CrossRef]
- Fang, Q.K.; Yao, G.Y.; Shi, Y.H.; Ding, C.C.; Wang, Y.; Wu, X.W.; Hua, R.M.; Cao, H.Q. Residue dynamics and risk assessment of prochloraz and its metabolite 2,4,6-trichlorophenol in apple. Molecules 2017, 22, 1780. [Google Scholar] [CrossRef]
- NY 5099-2002; Guideline for the Efficacy Evaluation of Fungicides. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2002.
- NY/T 1965.1-2010; Pesticides—Guidelines for the Field Efficacy Trials: Part 1. Fungicides. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2010.
- Qin, G.F.; Chen, Y.; He, F.R.; Yang, B.X.; Zou, K.T.; Shen, N.M.; Zuo, B.; Liu, R.X.; Zhang, W.; Li, Y.B. Risk assessment of fungicide pesticide residues in vegetables and fruits in the mid-western3 region of China. J. Food Compos. Anal. 2021, 95, 103663. [Google Scholar] [CrossRef]
- Marquette, C.A.; BlumL, L.J. Applications of the luminol chemiluminescent reaction in analytical chemistry. Anal. Bioanal. Chem. 2006, 385, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Santana-Mayor, A.; Rodríguez-Ramos, R.; Herrera-Herrera, A.V.; Socas-Rodríguez, B.; Rodríguez-Delgado, M.A. Updated overview of QuEChERS applications in food, environmental and biological analysis (2020–2023). Trac-Trend. Anal. Chem. 2023, 169, 117375. [Google Scholar] [CrossRef]
- GB 2763-2021; National Food Safety Standard—Maximum Residue Limits for Pesticides in Food. National Health Commission of the People’s Republic of China: Beijing, China, 2021.
- Schusterova, D.; Mraz, P.; Uttl, L.; Drabova, L.; Kocourek, V.; Hajslova, J. Pesticide residues in fresh and processed edible mushrooms from Czech markets. Food Addit. Contam. B 2023, 16, 384–392. [Google Scholar] [CrossRef]
- Aditya; Neeraj; Jarial, R.S.; Jarial, K.; Bhatia, J.N. Comprehensive review on oyster mushroom species (Agaricomycetes): Morphology, nutrition, cultivation and future aspects. Heliyon 2024, 10, e26539. [Google Scholar] [CrossRef]
- Suada, I.K.; Sudarma, I.; Kim, B.S.; Cha, J.Y.; Ohga, S. Fungal contaminant threaten oyster mushroom (Pleurotus ostreatus (Jacq. ex Fr.) Kummer) Cultivation in Bali. J. Fac. Agr. Kyushu. Univ. 2015, 60, 309–313. [Google Scholar] [CrossRef]
- Kim, M.K.; Seuk, S.W.; Lee, Y.H.; Kim, H.R.; Cho, K.M. Fungicide sensitivity and characterization of cobweb disease on a Pleurotus eryngii mushroom crop caused by Cladobotryum mycophilum. Plant Pathol. J. 2014, 30, 82–89. [Google Scholar] [CrossRef]
- Nurziya, Y.; Hao, J.Z.; Wei, P.; Jia, W.J.; Guan, J.H.; Wang, Y.H.; Gai, Q.W. Occurrence and identification of bacterial brown patch of processing mushrooms in Northern Xinjiang. Xinjiang Agric. Sci. 2012, 49, 2234–2238. [Google Scholar]
- Zhang, Q.; Zhang, C.H.; Song, H.Q.; Song, L.L.; Ma, H.J.; Wang, Z.H. Control effect of three compound fungicides on rhizopus and Pleurotus ostreatus diseases. J. Henan Agric. Sci. 2016, 45, 67–71. [Google Scholar]
- Zhu, T.T.; Ling, C.X. Effect of the different fungicides on the Toxicity of Pleurotus ostreatus and Trichoderma spp. J. Anhui Agric. Sci. 2011, 39(1410–1411+1414), 1410–1414. [Google Scholar]
- Ye, H.L.; Xia, X.Q.; Hu, R.P. Inhibitory effects of several biological fungicides on Neurospora sp. in edible fungus. J. Anhui Agric. Sci. 2015, 43, 121–122. [Google Scholar]
- Li, X.S.; Jiang, Y.P.; Shan, W.L.; Pan, C.P. Dissipation and residues detection of dioctyl diethylene triamine acetate in rice plant and environment by QuEChERS method and liquid chromatography/electrospray tandem mass spectrometry. B Environ. Contam. Tox. 2010, 84, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lü, K. Field efficacy of 1.8% seboctylamine acetate solution in controlling pepper virus disease. Hubei Plant Prot. 2012, 5, 10–11. [Google Scholar]
- Li, Q.Z. Analysis of the efficacy of several agents in controlling passion fruit mosaic virus disease. South Agric. 2017, 11, 9–10. [Google Scholar]
- Zang, H.Y.; Yang, X.; Gu, C.Y.; Sun, J.Z.; Pan, R.; Wang, Y.X.; Gao, T.C.; Yuan, S.K.; Chen, Y. A specific high toxicity of xinjunan (Dioctyldiethylenetriamine) to xanthomonas by affecting the iron metabolism. Microbiol. Spectr. 2023, 11, e0438222. [Google Scholar] [CrossRef]
- Jin, L.; Yang, J.; Wang, K. Mechanistic Study of the Antifungal Action of Alkyl Polyamine Fungicide Octylamine through Inhibition of Fungal ATP Synthesis. In Proceedings of the Annual Meeting of the Chinese Society of Plant Pathology; Anhui Agricultural University, Plant Protection Institute: Hefei, China, 2023; p. 627. [Google Scholar]
- Jin, M.J. Analysis of the 8% Xinjunan Acetate AS Residue on Tobacco and Dynamic Digestion Study. Master’s Thesis, Guizhou University, Guiyang, China, 2016. [Google Scholar]
- Le, L.H.T.; Tran-Lam, T.T.; Cam, T.Q.; Nguyen, T.N.; Dao, Y.H. Pesticides in edible mushrooms in Vietnam. Food Addit. Contam. B 2021, 14, 139–148. [Google Scholar] [CrossRef]
- Wen, Y.J.; Xiao, Z.Y.; Sun, Z.W. Safety risk assessment of pesticide residues in edible fungi and dietary exposure risk evaluation. J. Agric. Sci. 2024, 14, 77–82. [Google Scholar]
- Du, P.Q.; Wu, X.H.; Xu, J.; Dong, F.S.; Shi, Y.C.; Li, Y.B.; Liu, X.G.; Zheng, Y.Q. Different residue behaviors of four pesticides in mushroom using two different application methods. Environ. Sci. Pollut. Res. Int. 2018, 25, 8377–8387. [Google Scholar] [CrossRef]
- Jiao, J.; Shi, Y.C.; Wu, D. Residual dynamic of imidacloprid in Agrocybe aegerita and its substrates. Agrochemicals 2019, 58, 115–117. [Google Scholar]
- Han, C.; Hu, B.Z.; Chen, S.B.; Wang, N.; Hou, J.B.; Jin, N.; Shen, Y. Determination of Xinjunan pesticide residue in foodstuffs of plant origin by a modified QuEChERS method and ultra performance liquid chromatography-tandem mass spectrometry. LWT 2021, 151, 112101. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).