Abstract
Insect farming is gaining attention as a promising area for exploring probiotic bacteria, which can benefit both insect health and various industries. Silkworm farming is a key industry in Thailand; however, challenges such as disease susceptibility and optimising growth require innovative solutions for sustainable practices. Our study addresses this by assessing lactic acid bacteria (LAB) in native Thai silkworm faeces, which accumulate as natural by-products during the rearing process. We conducted biochemical tests, including those for catalase, haemolytic activity, bile salt tolerance, antimicrobial activity, antibiotic susceptibility, and cell surface hydrophobicity, along with taxonomic classification. Out of 102 isolates, eight potential probiotics were selected, with five showing strong probiotic traits like acid and bile salt tolerance and cell surface hydrophobicity, enhancing gut survivability. These isolates also displayed antagonistic activity against pathogens like Staphylococcus aureus, Salmonella typhimurium, Escherichia coli, and Pseudomonas aeruginosa. Safety assessments confirmed their safety, with no haemolytic activity and sensitivity to antibiotics like chloramphenicol and amoxicillin. These LAB isolates (SP04, SP06, SP44, SP64, and SP67), identified as Enterococcus faecalis strain NBRC 100481, show promise as in vitro probiotics for silkworm rearing, calling for further in vivo evaluation.
1. Introduction
As the global population continues to increase, the popularity of insects for both food and feed applications is steadily increasing. This trend is gaining momentum because of their remarkable nutritional profile and sustainable production practices, making insects a compelling solution to the challenges posed by the growing world population [1]. Insect farming may not only support protein self-sufficiency but can also offer novel means for organic waste management and the generation of value-added products, including fertilisers from rearing residues [2]. Bombyx mori (Lepidoptera: Bombycidae; Linnaeus, 1758) is one of the most important farmed insects in Thailand [3]. Throughout its life cycle, the silkworm undergoes various stages, starting as an egg, progressing through five larval instars, transforming into a pupa, and finally emerging as an adult silk moth (Figure 1). The polyvoltine silkworm can be reared year-round and is primarily found in tropical regions, such as Thailand. Beyond its traditional use in silk production, B. mori serves as a valuable insect model in life sciences and plays an important role in antiviral agent screening (e.g., silkworm–baculovirus infection model) [4] and in environmental monitoring [5], and also as food [6] and feed [7].
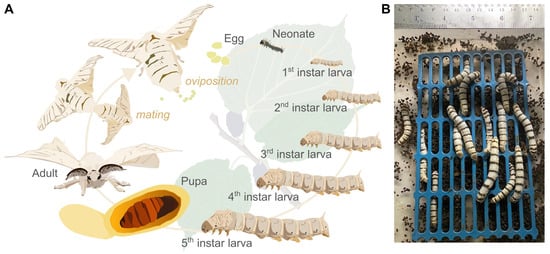
Figure 1.
(A) Silkworm (Bombyx mori) life cycle and (B) faeces harvested under aseptic conditions in a laminar flow cabinet.
Only the monophagous larvae of B. mori exclusively feed on mulberry (Morus spp.) leaves, generating faeces that are rich in nutrients and microorganisms [8]. This waste material represents a significant by-product of silkworm rearing and is produced in large quantities in Southeast Asian countries, including Thailand. Silkworm faeces, rich in (plant) nutrients and mixed with undigested substrate residues (frass), are used as organic fertilisers and in biogas production [9,10,11]. In Thailand, silkworm frass serves as a vital fertiliser for mulberry plantations, offering organic nutrients and showcasing sustainable agricultural practices. China has leveraged silkworm faeces in traditional medicine, emphasising their high fat, protein, and amino acid contents [12]. Reports indicate its use in pharmaceutical supplements and the food industry [13], such as oral iron supplementation, to ameliorate iron deficiency associated with anaemia [14]. Moreover, silkworm faeces have been shown to have antiviral and anti-inflammatory effects and can be used to treat symptoms related to rheumatoid arthritis [6]. Silkworm faeces primarily consist of organic matter and ash, constituting about 84–90% and 16–20% of their biomass, respectively [15]. In particular, faeces from silkworms reared on a natural mulberry leaf diet show an enriched content of amino acids, carbohydrates, and lipids [16].
Insects are generally considered a rich source of probiotic microorganisms that reside in their guts, where they provide metabolic support and defence mechanisms that support insect health [17]. These microbes are promising targets for biotechnological applications and have previously been isolated based on their strong cellulolytic [18], amylolytic [18], and xylolytic [19] properties. A metagenomic study by Yeruva et al. [20] revealed diverse patterns of microbial communities across various silkworm breeds, highlighting the need for further in-depth characterisation of silkworm-derived probiotics.
This study focused on the potential of lactic acid bacteria (LAB) isolated from faeces of a native Thai silkworm strain, considering the beneficial probiotic qualities of many LABs. Dong et al. [21] revealed that B. mori reared on fresh mulberry leaves had a higher bacterial diversity in their guts than those reared on an artificial diet. LAB, particularly those from the Lactobacillus genus, are widely acknowledged for their positive impact on host health, interacting with the gastrointestinal mucosa, stimulating the immune system, and decreasing pathogen infections [22]. Bermudez-Brito et al. [23] demonstrated and summarised six major mechanisms of action of probiotics: (a) enhancement of the epithelial barrier, (b) increased adhesion to the intestinal mucosa, (c) inhibition of pathogen adhesion, (d) competitive exclusion of pathogenic microorganisms, (e) production of antimicrobial substances, and (6) modulation of the immune system. Therefore, defining probiotic properties involves multiple screening criteria, such as tolerance to low pH and bile salts, inhibition of pathogens, cell surface hydrophobicity, safety assessments based on haemolytic activity, susceptibility to antibiotics, and the potential to induce host immune responses [24,25]. Bacteria belonging to the enterococci genus are widely recognised for their probiotic benefits and play a crucial role as food additives for human consumption [26]. Specifically, E. faecalis isolated from the meconium of newborns possesses notable probiotic properties suitable for applications that foster human health [27]. Enterococci have been extensively used to promote animal health and husbandry practices [28]. As feed additives, enterococci contribute to the induction of the immune system, thereby enhancing resistance against pathogens and supporting overall health.
Given the medicinal properties of silkworm faeces, this study aimed to isolate and screen LAB from the larval faeces of a Thai B. mori strain, with a focus on characterising their probiotic potential.
2. Materials and Methods
2.1. Silkworm Rearing and Harvesting of Fresh Faeces
Silkworm eggs of a Thai strain were purchased from a silkworm farm in the Kudrung District, Mahasarakham Province, Thailand. The eggs were incubated at 25–26 °C for 10 days. After hatching, the larvae were fed with mulberry leaves thrice a day (at 06:00, 12:00, and 18:00). Faecal samples were collected from 100 larvae on day 4 of the 5th instar (Figure 1A) under sterile conditions in a laminar flow cabinet to avoid cross-contamination (Figure 1B).
2.2. Isolation of Lactic Acid Bacteria (LAB) and Assessment of Acid Tolerance
The inoculum was prepared by mixing 10 g of silkworm faeces with 90 mL of sterile 0.9% saline solution. De Man–Rogosa–Sharpe (MRS) broth (pH 2.5) was inoculated with 100 µL of the silkworm faeces suspension and incubated at 37 °C for 2 h to determine the acid tolerance of lactic acid bacteria [29]. After the acid treatment, the samples were serially diluted from 10−1 to 10−4. Each dilution was plated in four replicates by spreading 100 µL on MRS agar (pH 6.2) supplemented with 1% CaCO3 and 0.004% bromocresol purple (BCP). The plates were incubated at 37 °C for 48 h under anaerobic conditions using an Anaeropack® system (Mitsubishi Gas Chemical Co. Inc., Tokyo, Japan). LAB colonies exhibiting characteristic morphological features and producing a yellow colour from BCP were selected for the screening of probiotic properties. Of the 102 colonies, 74 changed their colour to yellow and were surrounded by a clear zone.
2.3. Gram Staining and Catalase Test
Gram staining was conducted by transferring a bacterial culture onto a glass microscope slide using a sterilised inoculation loop and smearing it with water. The culture was stained with crystal violet for 1 min, followed by fixing with iodine dye for an additional minute. Discolouration was achieved by washing with 70% ethanol. Finally, the culture was counterstained with safranin for 30 s and then rinsed with water. Morphological features of the bacteria were observed under a light microscope at 40× magnification.
For the catalase test, the bacterial culture was smeared onto a dry and cleaned slide using a sterilised inoculating loop, and a drop of hydrogen peroxide was added to the culture. Immediate observation of bubbles indicated a positive catalase reaction, whereas the absence of bubbles indicated a negative catalase reaction. Only colonies showing gram-positive and catalase-negative reactions were selected. The selected colonies were then evaluated for probiotic properties and stored at −20 °C in MRS broth containing 20% glycerol for further studies.
2.4. Haemolytic Activity
The haemolytic activity of LAB was assessed following a modified protocol using human blood [30,31]. Overnight cultures of each LAB isolate in MRS broth were streaked onto Columbia agar plates (BD Difco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 5% human blood. The plates were incubated at 37 °C for 48 h. Subsequently, the bacterial plates were examined for the presence of haemolytic zones, categorised as alpha haemolysis (greenish zone), beta haemolysis (clear zone), or gamma haemolysis (no haemolysis) around the LAB colonies. Only the strains exhibiting gamma haemolysis were considered safe. The selected LAB isolates were then subjected to antibiotic assays, with Staphylococcus aureus serving as a positive control, showing a visible clear zone for beta haemolysis [32].
2.5. Bile Salt Tolerance Assay
The survival of LAB under acidic conditions with bile salts was also investigated. The bile salt tolerance of the isolates was evaluated by adding 0.3% (w/v) bile salt (Oxgall, Himedia, Maharashtra, India) to MRS broth (pH 2.5). For the test, a 5 mL culture of LAB isolates grown overnight in MRS broth at 37 °C under anaerobic conditions was used. Viable LAB colonies were determined using serial dilution and plate counting. MRS broth without LAB was used as a negative control. Survival rate was assessed by contrasting counts of viable colonies (mean log(cfu)/mL) before and after exposure to bile salts using the following Equation (1):
2.6. Cell Surface Hydrophobicity Assay
To assess the bacterial adhesion of LAB isolates to solvents, cell surface hydrophobicity was determined following established protocols [33,34]. LAB suspensions were prepared in phosphate-buffered saline (PBS) and centrifuged at 6500 rpm for 5 min. The PBS-resuspended LAB were mixed with an equal volume of physiological saline to obtain an optical density of 0.5 at a wavelength of 600 nm (OD600). Three mL of this suspension was transferred to an autoclaved tube and 1 mL of xylene was added. After shaking on a vortex shaker for 1 min, the tubes were left undisturbed for 15 min to facilitate phase separation. Subsequently, the absorbance of the aqueous phase was measured at a wavelength of 600 nm. Hydrophobicity was calculated from three replications, representing the percent decrease in the OD of the original bacterial suspension due to LAB partitioning into the hydrocarbon layer [35]. The hydrophobicity of the cells was measured at OD600, and the cell surface hydrophobicity ratio (%) was calculated using the following Equation (2) [29]:
2.7. Evaluation of LAB Isolates against Bacteria Pathogens
The antibacterial activities of the isolates against four human pathogens (Escherichia coli ATCC 25922, Salmonella typhimurium ATCC 14028, Pseudomonas aeruginosa, and Staphylococcus aureus ATCC 25923) were assessed using the agar spot method [36]. Sixteen LAB isolates were spotted onto the surface of MRS agar and incubated at 37 °C for 24 h under anaerobic conditions. After culturing, 50 µL of each bacterial pathogen was inoculated into a Brain Heart Infusion (BHI, 0.5% soft agar) and poured onto the MRS agar plates. The plates were incubated at 37 °C for another 24 h, and inhibition activity was observed. The diameter of the clear zone for each isolate against each indicator pathogen was measured, and inhibition levels were classified as (−) for no inhibition, (+) for 0.5–6 mm, (++) for 7–12 mm, and (+++) for more than 12 mm inhibition [37].
2.8. Sensitivity of LAB to Antibiotics
Antibiotic susceptibility was assessed using the Bauer–Kirby disk diffusion method [38] with commercial antibiotic discs (Himedia, Maharashtra, India). Five antibiotics, penicillin (10 units), chloramphenicol (30 µg), gentamicin (10 µg), tetracycline (30 µg), and amoxicillin (30 µg), were tested [39,40]. LAB isolates were swabbed on Mueller–Hinton agar, and antibiotic discs were placed on the plate before incubation at 37 °C for 24 h. The inhibition zone around each antibiotic disc was measured, and LAB sensitivity was categorised as sensitive (“S”), intermediate (“I”), or resistant (“R”) based on the enclosed zone size interpretative charts (Himedia): amoxicillin (S ≥ 18 mm; I = 14–17 mm; R ≤ 13 mm), chloramphenicol (S ≥ 18 mm; I = 13–17 mm; R ≤ 12 mm), gentamicin (S ≥ 15 mm; I = 13–14 mm; R ≤ 12 mm), penicillin (S ≥ 15 mm; R ≤ 14 mm), and tetracycline (S ≥ 15 mm; I = 12–14 mm; R ≤ 11 mm).
2.9. Colony PCR, Sanger Sequencing, and Taxonomic Identification
For a single colony PCR, a mixture of 0.5 µL 27F forward primer (5′-AGAGTTTGATCATGGCTCA-3′), 0.5 µL 1492R reverse primer (5′-TACGGTTACCTTGTTACGACTT-3′), 0.5 µL 2% BSA, and 12.5 µL RedTaq Polymerase 2× master mix (containing 1.5 mM MgCl2, VWR, Radnor, PA, USA) was prepared and combined with 11 µL PCR-grade H2O. Bacterial isolates selected from the enrichment plates were carefully transferred into reaction tubes containing 25 µL of the PCR mix using sterile pipette tips. Colony PCR was performed according to the thermocycler program described in Table S1. Subsequently, the PCR products were purified using the GeneElute PCR Clean-up Kit (Sigma-Aldrich, St. Louise, MO, USA). Prior to Sanger sequencing of the entire 16S rRNA gene, 2 µL of the 27F forward primer was added to each purified PCR product. The resulting full-length 16S reads were subjected to nucleotide BLAST search for taxonomic assignment. All the samples were amplified and sequenced in duplicate for more reliable verification.
2.10. Statistical Analysis
The experiments were carried out in triplicate (n = 3), and the results are presented as the mean ± standard deviation. Data analysis was performed in R (v. 4.3.1) using one-way analysis of variance, and statistical significance was defined as p < 0.05. Prior to analysis, normality of the data was checked using the Shapiro–Wilk test. Tukey’s Honest Significant Difference post hoc test was used for pairwise comparisons of groups.
3. Results
3.1. Morphological and Physiological Study of Lactic Acid Bacteria
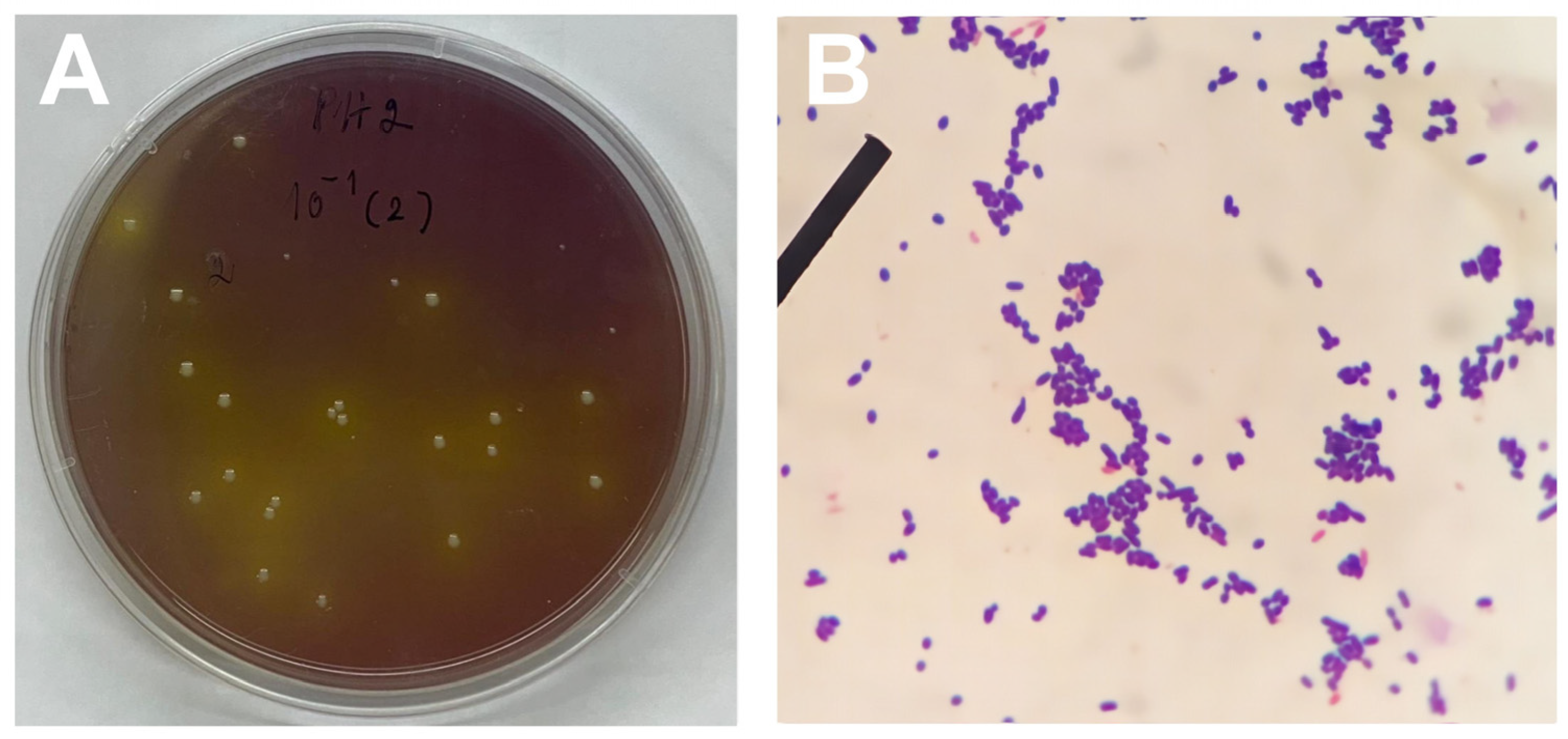
Through morphological analysis of the colonies, 102 LAB colonies derived from the faeces of a local Thai silkworm strain were identified. Initial characterisation of these isolated LAB involved criteria such as gram positivity, exhibiting either cocci or rod shapes (Figure 2A,B) and demonstrating survival in an acidic environment (pH 2.5) through the hydrolysis of CaCO3 combined with bromocresol purple on MRS agar. Notably, 74 isolates exhibited morphological features indicative of their potential as probiotic candidates, specifically for gram-positive bacteria. Additionally, all isolated strains demonstrated defence mechanisms against oxidative stress, as evidenced by a catalase-positive reaction.
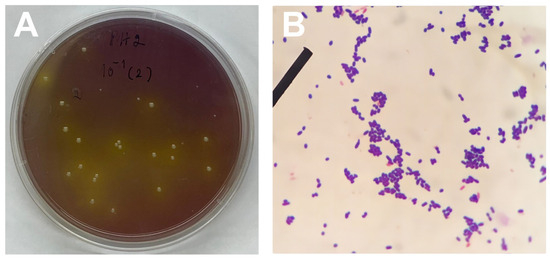
Figure 2.
Preliminary characterisation of lactic acid bacteria (LAB) isolated from faeces of silkworms: (A) single colonies of LAB isolated on selective MRS agar, (B) microscopic (40× magnification) observation of gram-positive LAB.
3.2. Haemolytic Activity, Bile Salt Tolerance, and Cell Surface Hydrophobicity
To assess the safety of the selected LAB, the haemolytic reaction was determined by streaking LAB on Columbia blood agar containing 5% (v/v) human blood. Out of the 74 isolates, 49 isolates did not exhibit a halo zone, confirming that they were gamma haemolytic. In contrast, a clear haemolytic zone (beta haemolysis) was observed around S. aureus, which served as positive control.
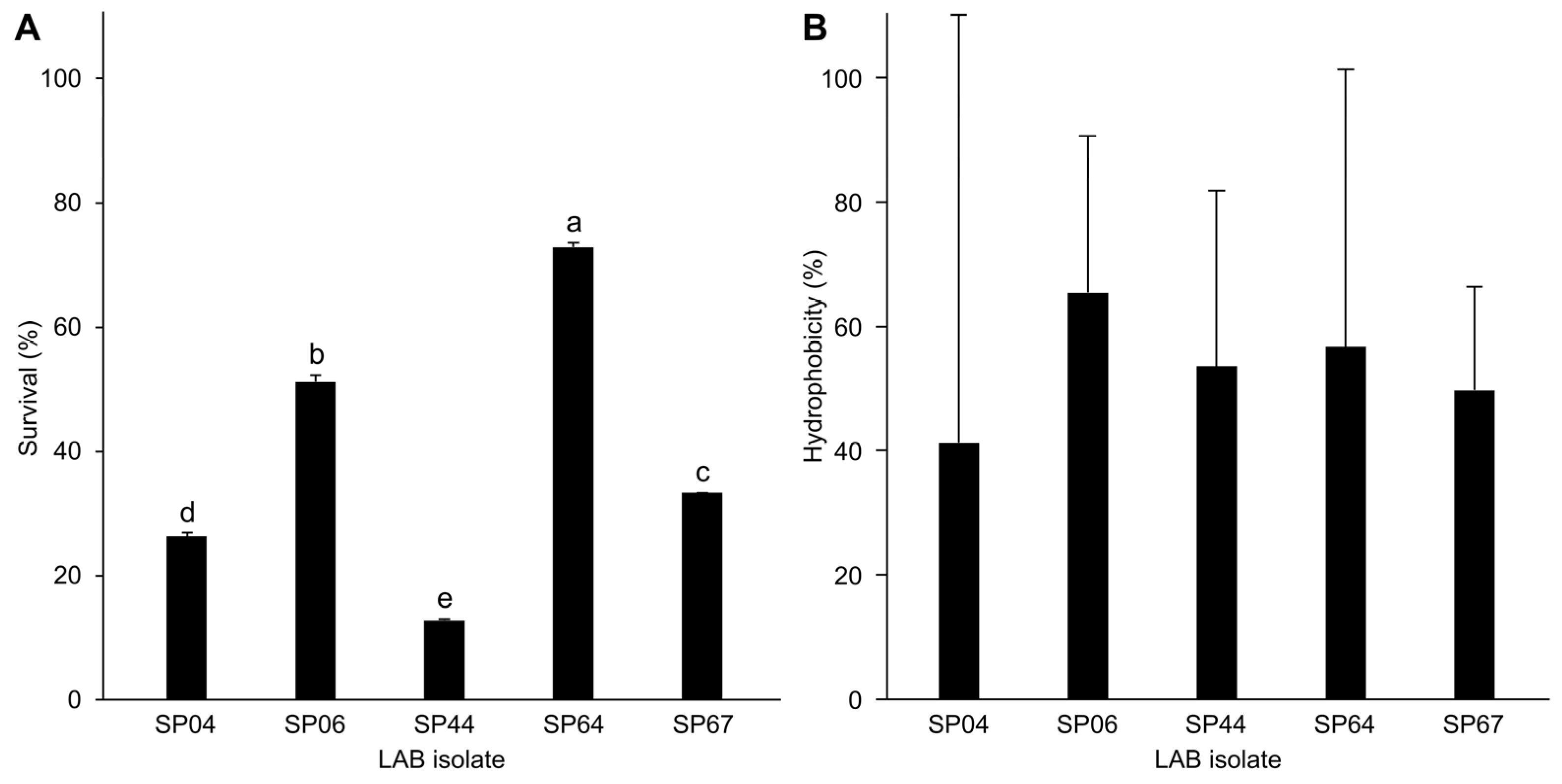
Among the LAB isolates, SP04, SP06, SP44, SP64, and SP67 displayed notable acid and bile salt tolerance (Figure 3A). After 24 h in bile salt conditions, their survival rates were 26.3 ± 0.6%, 51.2 ± 1.0%, 12.8 ± 0.3%, 72.9 ± 0.8%, and 33.3 ± 0.0%, resulting in highly significant differences across the isolates (F(4,10) = 3938, p < 0.001 ***).
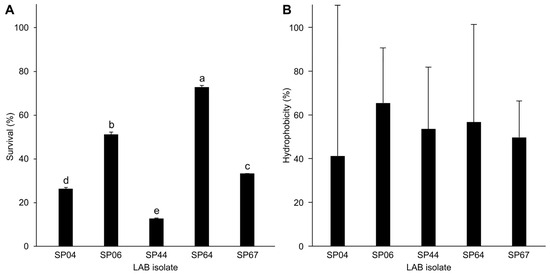
Figure 3.
(A) Selected lactic acid bacteria (LAB) isolates’ tolerance to bile salt (n = 3) (ANOVA: F(4,10) = 3938, p < 0.001). (B) Cell surface hydrophobicity of selected LAB (n = 3). No significant differences between LAB were observed in terms of hydrophobicity (ANOVA: F(4,10) = 0.141, p = 0.94). Values are presented as mean ± standard deviation. Lower-case letters indicate statistically significant differences between groups.
Our study revealed that LAB isolates exhibited hydrophobicity ranging from 41% to 65% on average (Figure 3B). Notably, we observed significant variation among replicates within each isolate yet detected no significant differences across distinct isolates (F(4,10) = 0.141, p = 0.94).
3.3. Selected LAB Isolates against Bacterial Pathogens
The selected LAB isolates were subjected to an agar spot test on MRS agar against indicator bacteria, demonstrating their inhibitory effects on pathogenic strains. The degree of inhibition, measured by the clear zone, varied between 4.2 mm and 19.8 mm, as detailed in Table 1.

Table 1.
Antimicrobial testing of selected lactic acid bacteria (LAB) against pathogenic bacteria in MRS broth (pH 6.5) (n = 3). Evaluation based on Vélez et al. (2007) [37]: 0.5–6 mm = (+), 7–12 mm = (++), >12 mm = (+++).
3.4. Antibiotic Susceptibility Test
To confirm the suitability of probiotic candidates for use as feed supplements, it is crucial to evaluate the sensitivity of the isolates to a relevant range of antibiotics of both human and animal importance. The five selected LAB isolates were subjected to antibiotic susceptibility testing using various antibiotics. In line with the guidelines by the European Food Safety Authority, two antibiotic groups were considered: (I) agents inhibiting cell wall synthesis (amoxicillin and penicillin) and (II) agents inhibiting protein synthesis (chloramphenicol, gentamicin, and tetracycline). Notably, the tested isolates demonstrated sensitivity to amoxicillin, chloramphenicol, penicillin, and tetracycline as denoted by the letter ‘S’ (sensitive). Four out of five isolates, however, showed resistance (‘R’) against gentamicin, and for isolate SP44, this antibiotic showed intermediate inhibitory (‘I’) effects, as detailed in Table 2.
Table 2.
Antibiotic susceptibility of selected lactic acid bacteria (LAB) isolates. Data on the measured inhibition zones are expressed as mean diameter ± standard deviation (n = 3). Susceptibility was determined based on the size interpretative charts enclosed by the antibiotic disc manufacturer (Himedia, Maharashtra, India). (R) = resistant, (S) = sensitive, (I) = intermediate inhibition.
3.5. Genetic Identification
The sequences derived from Sanger sequencing of the five isolates were all identified as Enterococcus faecalis strain NBRC 100481 (Accession no.: NR_113902.1). Comprehensive information, including raw sequences and detailed nucleotide BLAST output, is provided in Table S2.
4. Discussion
In the context of sericulture, silk production has been promoted and developed to support a circular economy. Recently, the silk-producing industry has found applications beyond traditional clothing, extending to uses in human food and animal feed [41]. As a by-product of insect mass-rearing, insect faeces have gained popularity, particularly as a novel agricultural fertiliser, due to their vast microbial and nutritional diversity and soil-enhancing properties [42,43]. In this study, we aimed to harness the potential of waste from silkworm farming by isolating LAB from the faeces of a Thai strain of B. mori and investigate its potential use as probiotics. The results indicated that these LAB isolates met the probiotic criteria established by Bermudez-Brito et al. (2012) [23]. Preliminary isolation tests were conducted using MRS nutrient media (MRS agar and MRS broth) at an acidified pH of 2.5. These tolerant isolates were further assessed for their probiotic properties. Five selected isolates demonstrated bile salt hydrolase activity and the ability to adhere to the epithelial cells. They were sensitive to amoxicillin, chloramphenicol, penicillin, and tetracycline, but showed resistance to gentamicin, indicating that their growth could be inhibited by a large selection of antibiotics if necessary. Based on their properties, the isolated LAB can be considered potential candidates for probiotics in animal feed [44]. However, caution should be paid to resistance to, e.g., aminoglycoside antibiotics such as gentamicin, which is frequently observed in LAB isolated from farmed animals [45]. The exposure of these animals to antibiotics increases the selective pressure on microbial communities in the host guts, favouring the proliferation of resistant microbes and driving the acquisition of resistances via horizontal gene transfer [45]. Artificial environments such as insect mass-rearing facilities may have similar effects on microbial populations. Although LAB, such as enterococci isolated from wild animals, also demonstrate resistances to antibiotics; farmed animals, pets, and humans constitute their primary reservoir [46]. Therefore, special attention should be directed towards monitoring the spread of resistances and associated virulence factors.
Sanger sequencing identified the isolates as Enterococcus faecalis strain NBRC 100481, a microaerophile mesophilic representative of the family Enterococcaceae. Recently, E. faecalis was found to be an effective antagonist against infections in B. mori caused by Microsporidia, including Nosema bombycis, the pathogen responsible for Pébrine disease in their larvae [47,48]. Enterococci are also capable of causing opportunistic infections [49] but are typically known for their probiotic effects when present as commensal gut colonisers. In healthy silkworms, enterococci make up a considerable amount of the gut microbiota, indicating their relevance to host health [50]. However, other representatives of enterococci such as Enterococcus mundtii, can have adverse effects on B. mori larvae and cause the flacherie disease [51], while the same species has been proven to protect another insect species, Tribolium castaneum, against Bacillus thuringiensis [52].
A critical aspect of probiotic bacteria is their ability to survive in acidic conditions in the gut. In our investigation involving 102 LAB isolates, all met the primary criteria for probiotic properties, displaying gram-positivity and growth in MRS broth with an acidic pH of 2.5 to a neutral pH of 6.7. These findings align with previous reports emphasising the importance of LAB viability in low-pH environments [53,54,55].
In this study, 102 different isolates were identified, of which 74 were found to be gram-positive and tested negative for catalase, as indicated by the absence of bubbles [56]. Out of these 74 isolates, 49 were cocci-shaped and exhibited gamma haemolysis. Additionally, bacterial cell surface hydrophobicity, assessed through auto-aggregation, demonstrated strong hydrophobicity to the hydrocarbon xylene, further supporting their potential as indigenous probiotics in the host gut [57,58].
Considering the importance of antibiotic resistance in probiotics, our results showed that the five LAB isolates identified as probiotic candidates exhibited susceptibility to common antibiotics [6]. This suggests that probiotics isolated from the faeces of silkworm larvae are safe for use in silkworm farming, benefiting from their origin in the native host’s gut environment and potentially offering advantages over probiotics isolated from other sources [59]. Moreover, these five isolates exhibited strong inhibitory effects on common pathogens such as E. coli, S. aureus, S. typhimurium, and P. aeruginosa. Enterococci are ubiquitous and can act as both probiotics or opportunistic pathogens depending on the strain and environmental conditions [60], though the specific mechanisms of their pathogen inhibition were not covered in this study. However, previous research has shown that E. faecalis can effectively colonise the intestine and exhibits immunoregulatory activities, including the activation of immune system cells [60]. Effective probiotics typically adhere to the gut lining, providing a barrier and excreting antimicrobial peptides against pathogens [61].
In this study, LAB were isolated from the faeces of a polyvoltine Thai silkworm strain. Previous research has shown a higher prevalence of Enterococcus sp. in polyvoltine than in bivoltine silkworms [20]. Probiotic criteria include the ability to reduce midgut pH from alkaline to acidic, which is a key factor in preventing infectious diseases caused by microorganisms, such as viruses, bacteria, and microsporidia.
Lately, lactic acid bacteria have been increasingly used as supplements and food additives in animals, including insects [19,62]. However, the mechanisms and interactions between silkworms and LAB remain unclear, and further research of LAB survival in the gut is necessary. Our results suggest that LAB supplementation may offer opportunities to enhance health while potentially reducing pathogens, particularly in preventing viral and bacterial infections in silkworms, an area with limited reports to date.
5. Conclusions
Our study was designed to capitalise on the utilisation of waste from silkworm farming. Lactic acid bacteria isolates were specifically selected from the faeces of a local strain of B. mori in Thailand for their promising probiotic potential. The five selected isolates demonstrated bile salt hydrolase activity and the ability to adhere to epithelial cells. Notably, all five isolates were sensitive to amoxicillin, chloramphenicol, penicillin, and tetracycline and exhibited resistance only to gentamicin. Considering their favourable probiotic properties, the selected LAB isolates were deemed potential candidates for use as probiotics in animal feed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14060924/s1, Table S1: Cycler program used for colony PCR with 27F and 1492R primers to amplify the 16S rRNA gene of bacterial isolates; Table S2: Representative sequences of the five isolates generated by Sanger sequencing using 27F and 1492R primers and their corresponding nucleotide BLAST results for taxonomic identification.
Author Contributions
Conceptualization, S.S.; methodology, S.S.; validation, S.S.; formal analysis, S.S., S.C., S.H. and T.K.; investigation, S.S., S.C., S.H. and T.K.; resources, S.S. and T.K.; writing—original draft preparation, S.S.; writing—review and editing, S.H. and T.K.; visualization, S.S. and T.K.; supervision, S.S.; project administration, S.S.; funding acquisition, S.S. and T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was financially supported by ‘Thailand Science Research and Innovation (TSRI)’. Moreover, the research reported in this publication was jointly supported by the ASEAN-European Academic University Network (ASEA-UNINET), the Austrian Federal Ministry of Education, Science and Research (BMBWF), and the OeAD—Austria’s Agency for Education and Internationalisation.
Institutional Review Board Statement
The research conducted in this study involved the use of B. mori, a widely cultivated model insect globally. As such, the study did not raise concerns related to experimental animal welfare, and no additional ethical approval was required for the procedures followed in this research.
Data Availability Statement
The 16S rRNA gene nucleotide sequences of the identified isolates have been publicly deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 20 May 2024)) under submission SUB14404462, covering accession numbers PP727389-PP727398.
Acknowledgments
The authors express their gratitude to the two anonymous reviewers for their valuable feedback, which significantly enhanced the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Alexandratos, N.; Bruinsma, J.; Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; Agricultural Development Economics Division, Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar] [CrossRef]
- Zhou, Y.; Xiao, R.; Klammsteiner, T.; Kong, X.; Yan, B.; Mihai, F.-C.; Liu, T.; Zhang, Z.; Kumar Awasthi, M. Recent Trends and Advances in Composting and Vermicomposting Technologies: A Review. Bioresour. Technol. 2022, 360, 127591. [Google Scholar] [CrossRef] [PubMed]
- Hanboonsong, Y.; Jamjanya, T.; Durst, P.B. Six-Legged Livestock: Edible Insect Farming, Collection and Marketing in Thailand; RAP Publication; Food and Agriculture Organization of the United Nations, Regional Office for Asia and the Pacific: Bangkok, Thailand, 2013; ISBN 978-92-5-107578-4. [Google Scholar]
- Orihara, Y.; Hamamoto, H.; Kasuga, H.; Shimada, T.; Kawaguchi, Y.; Sekimizu, K. A Silkworm–Baculovirus Model for Assessing the Therapeutic Effects of Antiviral Compounds: Characterization and Application to the Isolation of Antivirals from Traditional Medicines. J. Gen. Virol. 2008, 89, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, H.; Kurokawa, K.; Kaito, C.; Kamura, K.; Manitra Razanajatovo, I.; Kusuhara, H.; Santa, T.; Sekimizu, K. Quantitative Evaluation of the Therapeutic Effects of Antibiotics Using Silkworms Infected with Human Pathogenic Microorganisms. Antimicrob. Agents Chemother. 2004, 48, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, S.; Duan, H.; Wang, J.; Yan, W. Silkworm Pupae: A Functional Food with Health Benefits for Humans. Foods 2022, 11, 1594. [Google Scholar] [CrossRef] [PubMed]
- Banday, M.T.; Adil, S.; Sheikh, I.U.; Hamadani, H.; Qadri, F.I.; Sahfi, M.E.; Sait, H.S.A.W.; Abd El-Mageed, T.A.; Salem, H.M.; Taha, A.E.; et al. The Use of Silkworm Pupae (Bombyx mori) Meal as an Alternative Protein Source for Poultry. Worlds Poult. Sci. J. 2023, 79, 119–134. [Google Scholar] [CrossRef]
- Park, J.-H.; Lee, D.-G.; Yeon, S.-W.; Kwon, H.-S.; Ko, J.-H.; Shin, D.-J.; Park, H.-S.; Kim, Y.-S.; Bang, M.-H.; Baek, N.-I. Isolation of Megastigmane Sesquiterpenes from the Silkworm (Bombyx mori L.) Droppings and Their Promotion Activity on HO-1 and SIRT1. Arch. Pharm. Res. 2011, 34, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Łochyńska, M.; Frankowski, J. The Biogas Production Potential from Silkworm Waste. Waste Manag. 2018, 79, 564–570. [Google Scholar] [CrossRef]
- Łochyńska, M.; Frankowski, J. Impact of Silkworm Excrement Organic Fertilizer on Hemp Biomass Yield and Composition. J. Ecol. Eng. 2019, 20, 63–71. [Google Scholar] [CrossRef]
- Van De Zande, E.M.; Wantulla, M.; Van Loon, J.J.A.; Dicke, M. Soil Amendment with Insect Frass and Exuviae Affects Rhizosphere Bacterial Community, Shoot Growth and Carbon/Nitrogen Ratio of a Brassicaceous Plant. Plant Soil 2023, 495, 631–648. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Somkhanngoen, C.; Sukrong, S. Potential Pharmaceutical Uses of the Isolated Compounds from Silkworm Excreta. Chiang Mai J. Sci. 2013, 41, 97. [Google Scholar]
- Tulp, M.; Bohlin, L. Unconventional Natural Sources for Future Drug Discovery. Drug Discov. Today 2004, 9, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wu, J.; Li, Q.; Gu, S.; Chen, Z.; Wang, K. Silkworm Feces Extract Improves Iron Deficiency Anemia via Suppressing Hepcidin Expression and Promoting Iron-Regulatory Proteins Expression. RSC Adv. 2017, 7, 50378–50388. [Google Scholar] [CrossRef]
- Neelagund, S.; Ingalhalli, S.; Savanurmath, C.; Hinchigeri, S.; Hiremath, M. Purification and Characterization of Antiviral Protein from Silkworm Fecal Matter. Casp. J. Environ. Sci. 2007, 5, 77–85. [Google Scholar]
- Qin, D.; Wang, G.; Dong, Z.; Xia, Q.; Zhao, P. Comparative Fecal Metabolomes of Silkworms Being Fed Mulberry Leaf and Artificial Diet. Insects 2020, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Huerta-García, A.; Álvarez-Cervantes, J. The Gut Microbiota of Insects: A Potential Source of Bacteria and Metabolites. Int. J. Trop. Insect Sci. 2024, 44, 13–30. [Google Scholar] [CrossRef]
- Liang, X.; Fu, Y.; Liu, H. Isolation and Characterization of Enzyme-Producing Bacteria of the Silkworm Larval Gut in Bioregenerative Life Support System. Acta Astronaut. 2015, 116, 247–253. [Google Scholar] [CrossRef]
- Unban, K.; Klongklaew, A.; Kodchasee, P.; Pamueangmun, P.; Shetty, K.; Khanongnuch, C. Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture. Insects 2022, 13, 136. [Google Scholar] [CrossRef]
- Yeruva, T.; Vankadara, S.; Ramasamy, S.; Lingaiah, K. Identification of Potential Probiotics in the Midgut of Mulberry Silkworm, Bombyx mori through Metagenomic Approach. Probiotics Antimicrob. Proteins 2020, 12, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.-L.; Zhang, S.-X.; Chen, Z.-H.; Tao, H.; Li, X.; Qiu, J.-F.; Cui, W.-Z.; Sima, Y.-H.; Cui, W.-Z.; Xu, S.-Q. Differences in Gut Microbiota between Silkworms (Bombyx mori) Reared on Fresh Mulberry (Morus alba Var. Multicaulis) Leaves or an Artificial Diet. RSC Adv. 2018, 8, 26188–26200. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Safety Evaluation of Certain Contaminants in Food; Food and Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Guarner, F. Probiotics. Int. J. Food Microbiol. 1998, 39, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Lähteinen, T.; Malinen, E.; Koort, J.M.K.; Mertaniemi-Hannus, U.; Hankimo, T.; Karikoski, N.; Pakkanen, S.; Laine, H.; Sillanpää, H.; Söderholm, H.; et al. Probiotic Properties of Lactobacillus Isolates Originating from Porcine Intestine and Feces. Anaerobe 2010, 16, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Yang, H.; Shigwedha, N.; Zhang, S.; Liu, F.; Zhang, L. Probiotic Effects and Metabolic Products of Enterococcus faecalis LD33 with Respiration Capacity. Foods 2022, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Al Atya, A.K.; Drider-Hadiouche, K.; Ravallec, R.; Silvain, A.; Vachee, A.; Drider, D. Probiotic Potential of Enterococcus faecalis Strains Isolated from Meconium. Front. Microbiol. 2015, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, Y.; Meng, Q.; Zhang, X.; Wang, X. Progress in the Application of Enterococcus faecium in Animal Husbandry. Front. Cell. Infect. Microbiol. 2023, 13, 1168189. [Google Scholar] [CrossRef] [PubMed]
- Jena, P.K.; Trivedi, D.; Thakore, K.; Chaudhary, H.; Giri, S.S.; Seshadri, S. Isolation and Characterization of Probiotic Properties of Lactobacilli Isolated from Rat Fecal Microbiota. Microbiol. Immunol. 2013, 57, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A.; Clogston, J.D.; Neun, B.W.; Hall, J.B.; Patri, A.K.; McNeil, S.E. Method for Analysis of Nanoparticle Hemolytic Properties in Vitro. Nano Lett. 2008, 8, 2180–2187. [Google Scholar] [CrossRef]
- Mangia, N.P.; Saliba, L.; Deiana, P. Functional and Safety Characterization of Autochthonous Lactobacillus paracasei FS103 Isolated from Sheep Cheese and Its Survival in Sheep and Cow Fermented Milks during Cold Storage. Ann. Microbiol. 2019, 69, 161–170. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Liu, Y.; Zhong, J.; Zhang, D. Assessing the Safety and Probiotic Characteristics of Lacticaseibacillus rhamnosus X253 via Complete Genome and Phenotype Analysis. Microorganisms 2023, 11, 140. [Google Scholar] [CrossRef]
- Boiocchi, F.; Porcellato, D.; Limonta, L.; Picozzi, C.; Vigentini, I.; Locatelli, D.P.; Foschino, R. Insect Frass in Stored Cereal Products as a Potential Source of Lactobacillus sanfranciscensis for Sourdough Ecosystem. J. Appl. Microbiol. 2017, 123, 944–955. [Google Scholar] [CrossRef]
- Hamidreza, T.; Fatemeh, T.; Hossein, M.; Mojtaba, Z.; Mahmood, S.; Parvin, S. Potential Probiotic of Lactobacillus johnsonii LT171 for Chicken Nutrition. Afr. J. Biotechnol. 2009, 8, 5833–5837. [Google Scholar] [CrossRef]
- Krausova, G.; Hyrslova, I.; Hynstova, I. In Vitro Evaluation of Adhesion Capacity, Hydrophobicity, and Auto-Aggregation of Newly Isolated Potential Probiotic Strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef]
- Schillinger, U.; Lücke, F.K. Antibacterial Activity of Lactobacillus sake Isolated from Meat. Appl. Environ. Microbiol. 1989, 55, 1901–1906. [Google Scholar] [CrossRef]
- Vélez, M.P.; Hermans, K.; Verhoeven, T.L.A.; Lebeer, S.E.; Vanderleyden, J.; De Keersmaecker, S.C.J. Identification and Characterization of Starter Lactic Acid Bacteria and Probiotics from Columbian Dairy Products. J. Appl. Microbiol. 2007, 103, 666–674. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Marchwińska, K.; Gwiazdowska, D. Isolation and Probiotic Potential of Lactic Acid Bacteria from Swine Feces for Feed Additive Composition. Arch. Microbiol. 2022, 204, 61. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Balcázar, J.L.; García, Y.; Halaihel, N.; Vendrell, D.; De Blas, I.; Merrifield, D.L.; Ruiz-Zarzuela, I. Identification and Characterization of Lactic Acid Bacteria Isolated from Rainbow Trout, Oncorhynchus mykiss (Walbaum), with Inhibitory Activity against Lactococcus garvieae: Trout Endogenous LAB Antagonise L. garvieae. J. Fish Dis. 2011, 34, 499–507. [Google Scholar] [CrossRef]
- Dhakal, R.; Sahu, R.K.; Das Gupta, D.; Saikia, M.; Borthakur, S.; Majumder, M.; Mech, S.; Thapa, M.; Dutta, P.; Kalita, J. Recycling of Protein Rich Silk Industry Waste for Potential Food and Therapeutic Application. Food Biosci. 2024, 60, 104461. [Google Scholar] [CrossRef]
- Praeg, N.; Klammsteiner, T. Primary study on frass fertilizers from mass-reared insects: Species variation, heat treatment effects, and implications for soil application at laboratory scale. J. Environ. Manag. 2024, 356, 120622. [Google Scholar] [CrossRef]
- Insam, H.; Klammsteiner, T.; Gómez-Brandòn, M. Biology of Compost. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2023; pp. 522–532. ISBN 978-0-12-409548-9. [Google Scholar]
- Menconi, A.; Kallapura, G.; Latorre, J.D.; Morgan, M.J.; Pumford, N.R.; Hargis, B.M.; Tellez, G. Identification and Characterization of Lactic Acid Bacteria in a Commercial Probiotic Culture. Biosci. Microbiota Food Health 2014, 33, 25–30. [Google Scholar] [CrossRef]
- Jaimee, G.; Halami, P.M. Emerging Resistance to Aminoglycosides in Lactic Acid Bacteria of Food Origin—An Impending Menace. Appl. Microbiol. Biotechnol. 2016, 100, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Klibi, N.; Aouini, R.; Borgo, F.; Ben Said, L.; Ferrario, C.; Dziri, R.; Boudabous, A.; Torres, C.; Ben Slama, K. Antibiotic Resistance and Virulence of Faecal Enterococci Isolated from Food-Producing Animals in Tunisia. Ann. Microbiol. 2015, 65, 695–702. [Google Scholar] [CrossRef]
- Zhang, X.; Feng, H.; He, J.; Muhammad, A.; Zhang, F.; Lu, X. Features and Colonization Strategies of Enterococcus faecalis in the Gut of Bombyx mori. Front. Microbiol. 2022, 13, 921330. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, H.; He, J.; Liang, X.; Zhang, N.; Shao, Y.; Zhang, F.; Lu, X. The Gut Commensal Bacterium Enterococcus faecalis LX10 Contributes to Defending against Nosema bombycis Infection in Bombyx mori. Pest Manag. Sci. 2022, 78, 2215–2227. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of Animal Origin. Microbiol. Spectr. 2018, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Lu, Y.; Zhang, H.; Kumar, D.; Liu, B.; Gong, Y.; Zhu, M.; Zhu, L.; Liang, Z.; Kuang, S.; et al. Effects of BmCPV Infection on Silkworm Bombyx mori Intestinal Bacteria. PLoS ONE 2016, 11, e0146313. [Google Scholar] [CrossRef]
- Cappellozza, S.; Saviane, A.; Tettamanti, G.; Squadrin, M.; Vendramin, E.; Paolucci, P.; Franzetti, E.; Squartini, A. Identification of Enterococcus mundtii as a Pathogenic Agent Involved in the “Flacherie” Disease in Bombyx mori L. Larvae Reared on Artificial Diet. J. Invertebr. Pathol. 2011, 106, 386–393. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Probiotic Enterococcus mundtii Isolate Protects the Model Insect Tribolium castaneum against Bacillus thuringiensis. Front. Microbiol. 2017, 8, 1261. [Google Scholar] [CrossRef]
- Cebeci, A.; Gürakan, C. Properties of Potential Probiotic Lactobacillus plantarum Strains. Food Microbiol. 2003, 20, 511–518. [Google Scholar] [CrossRef]
- Ripamonti, B.; Agazzi, A.; Bersani, C.; De Dea, P.; Pecorini, C.; Pirani, S.; Rebucci, R.; Savoini, G.; Stella, S.; Stenico, A.; et al. Screening of Species-Specific Lactic Acid Bacteria for Veal Calves Multi-Strain Probiotic Adjuncts. Anaerobe 2011, 17, 97–105. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile Salt and Acid Tolerance of Lactobacillus rhamnosus Strains Isolated from Parmigiano Reggiano Cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Bazireh, H.; Shariati, P.; Azimzadeh Jamalkandi, S.; Ahmadi, A.; Boroumand, M.A. Isolation of Novel Probiotic Lactobacillus and Enterococcus Strains from Human Salivary and Fecal Sources. Front. Microbiol. 2020, 11, 597946. [Google Scholar] [CrossRef] [PubMed]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, Autoaggregation and Hydrophobicity of 13 Strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Patel, A.K.; Ahire, J.J.; Pawar, S.P.; Chaudhari, B.L.; Chincholkar, S.B. Comparative Accounts of Probiotic Characteristics of Bacillus spp. Isolated from Food Wastes. Food Res. Int. 2009, 42, 505–510. [Google Scholar] [CrossRef]
- Asadi, A.; Lohrasbi, V.; Abdi, M.; Mirkalantari, S.; Esghaei, M.; Kashanian, M.; Oshaghi, M.; Talebi, M. The Probiotic Properties and Potential of Vaginal Lactobacillus spp. Isolated from Healthy Women against Some Vaginal Pathogens. Lett. Appl. Microbiol. 2022, 74, 752–764. [Google Scholar] [CrossRef]
- Krawczyk, B.; Wityk, P.; Gałęcka, M.; Michalik, M. The Many Faces of Enterococcus spp.—Commensal, Probiotic and Opportunistic Pathogen. Microorganisms 2021, 9, 1900. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.L. Antagonistic Activities of Lactobacilli and Bifidobacteria against Microbial Pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef]
- Savio, C.; Mugo-Kamiri, L.; Upfold, J.K. Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects. Insects 2022, 13, 376. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).