Abstract
In 2020, a plant displaying promising attributes as both an attractant and bioinsecticide for Oryctes rhinoceros (Coleoptera: Scarabaeidae), known locally as “Hujan Emas”, was discovered at PT Sampoerna Agro Tbk, Indonesia. Despite its potential, the identity of this plant remains unknown. This study aims to reveal the plant’s identity and ascertain its potential as an attractant and bioinsecticide against O. rhinoceros. Identification was performed using morphological and sequence analyses of the MaturaseK, chloroplast rps16, and chloroplast rpl16 genes. Field observations were conducted over 9 months, focusing on the number of adult O. rhinoceros visiting the plant and the number captured using synthetic pheromone (ethyl 4-methyloctanoate) traps. Bioinsecticidal activity assessments focused on extracts from the plant’s bark, stem, and leaves. Morphological characterization and sequence analysis conclusively affirmed the plant’s identity as Senna multijuga (Fabales: Fabaceae). The total number of O. rhinoceros specimens obtained from S. multijuga was 1.6 times higher (836 beetles) than that obtained using the pheromone trap (512 beetles), suggesting that S. multijuga has better attractant capability than ethyl 4-methyloctanoate. The leaf extract exhibited insecticidal activity against larvae, while the stem and bark extracts demonstrated insecticidal activity against both larvae and adults. Larval mortality ranged from 1.25% to 3.75%, while adult mortality was notably higher, ranging from 12.5% to 25%. This is the first report on the capability of S. multijuga as an attractant and bioinsecticide against O. rhinoceros.
1. Introduction
Oryctes rhinoceros (L., 1758) (Coleoptera: Scarabaeidae), commonly known as the coconut rhinoceros beetle (CRB), is a significant pest of coconut plants, including oil palm and other plants belonging to the palm family [1,2,3]. The O. rhinoceros beetles attack and penetrate the plant’s growing tip, spreading to attack other plant tissues and damaging the plant’s meristem, thereby inhibiting its growth. The pest’s infestation disrupts the plant’s photosynthesis process, leading to a decrease in production [1]. The yield loss caused by this pest in oil palm plants can reach up to 25% [3]. The holes created by O. rhinoceros can serve as entry points for other pests or plant pathogens, exacerbating the damage caused [1].
The rhinoceros beetle lives and reproduces in organic matter derived from plant residues or animal dung [4,5]. Its biological cycle consists of one pupal and four larval stages. O. rhinoceros requires a time span of 100 to 250 days to reach the adult stage. Instars 1 to 3 are the most vulnerable larval stages and are the most suitable for control measures. Instar 3 larvae are the most voracious and have a lifespan of 60–165 days. They subsequently transform into pupae after the pre-pupal stage (instar 4 larvae), lasting 8–13 days [4,6].
The emergence of O. rhinoceros as an important oil palm pest at PT Sampoerna Agro Tbk prompted the company to focus specifically on its control. In 2020, the infestation of O. rhinoceros on non-yielding oil palm at PT Sampoerna Agro Tbk covered 1764.4 hectares, with infestation levels fluctuating monthly between 0.1% and 38.1%. Within the affected area, 55% (966 ha) experienced severe infestation (>5%), while 35% (798 ha) had mild infestation (<5%). Various methods have been employed to control O. rhinoceros, such as planting legume cover crops, manually removing larvae, installing pheromone traps, and using carbosulfan insecticide. However, these efforts have not yet produced satisfactory outcomes. The considerable cost of controlling O. rhinoceros on non-yielding oil palm amounts to IDR 14,743,476 (USD 932.71) per hectare; this has led PT Sampoerna Agro Tbk to explore alternative control approaches that are more effective, cost-efficient, and eco-friendly.
Various plant substances have been reported to play a role as insecticides, and their use is becoming a safe, environmentally friendly, and sustainable control alternative. The use of plant substances as insecticides does not leave harmful residues on crop yields. The search for local plant substances available around the cultivation area, making them easy to find and apply, continues to be carried out, including insecticides to control O. rhinoceros.
In 2020, a plant that appeared to have potential as an attractant and bioinsecticide for O. rhinoceros was discovered in the PT Sampoerna Agro Tbk plantation in Indonesia, locally known as “Hujan Emas” (golden rain). Many O. rhinoceros beetles were observed to visit the branches of the plant and then die after eating part of the plant for 1–2 days. Until now, the identity of this plant was unknown, and no scientific studies have been carried out on its capability as a bioinsecticide for controlling O. rhinoceros. This research aims to determine the identity and confirm the ability of “Hujan Emas” as an attractant and a bioinsecticide against O. rhinoceros.
2. Materials and Methods
2.1. Identification of “Hujan Emas” (Golden Rain) Plants
2.1.1. Morphological Characteristics
Morphological characteristics include both vegetative and generative features. Vegetative characteristics encompass the canopy, branches, stems, and leaves, while generative characteristics comprise flowers and seeds. The leaf parts, including branches, flowers, and fruits, were collected from 2-year-old golden rain plants in January 2023 from the oil palm plantation of PT Sampoerna Agro Tbk. The plant samples were then cleaned of impurities and preserved as dried specimens (herbarium). The sample was placed in a transparent plastic bag and identified by Dr. Silva Abraham, S.Si, M.Si, at the Directorate of Scientific Collection Management, National Research and Innovation Agency (BRIN), Cibinong, Bogor, West Java, Indonesia (registration ID 77561).
2.1.2. Molecular Identification
Primer design: Three pairs of primers were created for the identification of the golden rain plant. These primer pairs were designed based on the sequences of the MaturaseK (MatK), chloroplast rps16 (rps16), and chloroplast rpl16 (rpl16) genes. These genes serve as standard plant barcodes, and they have been widely used to identify many plant species [7,8] including Senna spp. [9]. The reference strains used for constructing the dendrogram were retrieved from GenBank (https://www.ncbi.nlm.nih.gov/ (accessed on 26 December 2023).
- DNA Extraction
DNA extraction was performed using the cetyl trimethylammonium bromide (CTAB) method as described by [10] with slight modifications. A total of 0.5 g of the golden rain plant sample was placed into a mortar, and 600 µL TE buffer pH (8.0) (1st Base, Malaysia) was added. Then, the sample was ground until smooth. After grinding, the mortar was sealed with aluminum foil and incubated at −40 °C overnight. Following this, the lid was opened, and the sample was ground again for 15 min. A total of 0.5 mL of the ground material was transferred to a 1.5 mL tube, and then 400 µL of 2% CTAB was added and homogenized before being incubated at 65 °C for 1 h. After incubation, 500 µL of phenol chloroform isoamyl alcohol (P:C:I) (25:4:1) was added and homogenized, and this mixture was centrifuged at 14,000 rpm for 10 min. A total of 600 µL of the supernatant was collected and transferred to a new 1.5 mL tube, and chloroform isoamyl alcohol (C:I) (24:1) was added in an equal volume (600 µL, the same as the obtained supernatant volume), homogenized, and then centrifuged at 14,000 rpm for 10 min. A total of 400 µL of the supernatant was collected and transferred to a new tube. Then, 400 µL of isopropanol was added and incubated at −20 °C for 20 min. After incubation, the tube was centrifuged for 10 min, and the supernatant was discarded. A total of 500 µL of 70% ethanol was added, and the mixture was centrifuged at 14,000 rpm for 10 min. The entire supernatant was then discarded, and the obtained pellet was incubated for 1 day. Following this, 20 µL of TE buffer pH (8.0) (1st Base, Malaysia) was added to the pellet, and the extracted DNA was ready for PCR use. Centrifugation was performed using a Microspin12 centrifuge (Biosan, Latvia).
- DNA Amplification
Amplification was performed using the Sensoquest Thermal Cycler Machine (Germany). In this study, three pairs of primers for the identification of the golden rain plant were successfully designed based on the sequences of the MaturaseK (MatK) (matKsenF and matKsenR), chloroplast rps16 (rps16) (rps16senF and rps16senR), and chloroplast rpl16 (rpl16) (rpl16senF and rpl16senR) genes (Table 1). PCR was carried out using a total volume of 25 µL, consisting of 12.5 µL Master Mix (Red Mix) (Bioline, Memphis, TN, USA), 1 µL forward primer, 1 µL reverse primer, 1 µL DNA template (approximately 1 µg/µL), and 9.5 µL sterile water. DNA amplification involved an initiation cycle at 95 °C for 5 min, followed by 30 cycles consisting of denaturation at 95 °C for 1 min, primer annealing (all primer sets) at 53 °C for 1 min, primer extension at 72 °C for 1 min, and a final elongation cycle at 72 °C for 5 min. The PCR results were electrophoresed using a 0.5% agarose gel suspended in 20 mL 1× Tris-Boric Acid-EDTA (TBE) buffer (1st Base, Malaysia) containing 1 µL ethidium bromide (EtBr 10 mg/mL). Electrophoresis was conducted using 1× TBE buffer at a voltage of 50 volts for 60 min. The results were visualized using a DigiDoc UV transilluminator (UVP, Upland, CA, USA).

Table 1.
Primers developed in this study.
- Sequencing and Analysis of Results
The PCR results were subsequently sent to 1st Base Malaysia for sequencing. The sequencing results were then processed using BioEdit ver. 7.2.6 for Windows [11] and submitted to the basic local alignment search tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 26 December 2023) to determine potential identities. A dendrogram was created using Mega 11 for Windows [12] employing the maximum likelihood method. The reference strains used were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/ (accessed on 26 December 2023) (Supplementary Table S1).
2.2. The Potential of the “Hujan Emas” (Golden Rain) Plant as an Attractant for O. rhinoceros Beetles
The golden rain plant was introduced and planted in PT Sampoerna Agro Tbk in 2015 as an ornamental plant. The potential of the golden plant as a bioinsecticide and attractant was first recognized in 2020. Further observations were performed to confirm its potential.
Additional field observations were performed in the oil palm plantation area of PT Sampoerna Agro Tbk, located in the Belida estate, divisions I and II, Ogan Komering Ilir (OKI), South Sumatra (3°48′28.613′′ S, 105°0′53.874′′ E), Indonesia (Figure 1). The experiment was performed using an inter-site (oversite) design [13] with two factors, namely, golden rain plants and commercial pheromone traps.
Figure 1.
Observation site (PT Sampoerna Agro, Tbk). (A) Map of Indonesia showing the location of South Sumatra province filled in red. (B) A map of South Sumatra (filled in yellow), with a red-colored box showing the location of PT Sampoerna Agro Tbk Belida estate, divisions I and II, in Ogan Komering Ilir (OKI).
The field observations were conducted once per month over 9 months (January to September) in 2020 on 25 golden rain plants, covering 625 hectares with a density of 1 plant per 25 hectares. These plants were 18 months old. The golden rain plants were planted since 2018 at a spacing of 8–10 m in proximity to the installation area of commercial pheromone traps. The data collected included the number of O. rhinoceros beetles successfully captured when visiting the golden rain plants, both perched and engaging in activities on the trunk or found dead beneath the plants. The capture results for the pheromone traps represent the population of beetles trapped in the pherotrap. The pheromone trap used was ethyl 4-methyloctanoate, a synthetic attractant (a sex pheromone) for O. rhinoceros. Observation of the beetles trapped in pheromone traps was performed once per month over 9 months (January to September) in 2020. A total of 299 pheromone traps were installed, covering an area of 897 hectares with a density of 1 trap per 3 hectares. Installation was carried out by placing the pheromone on the trap and suspending it on a wooden structure at a height of 3 m (Figure 2).
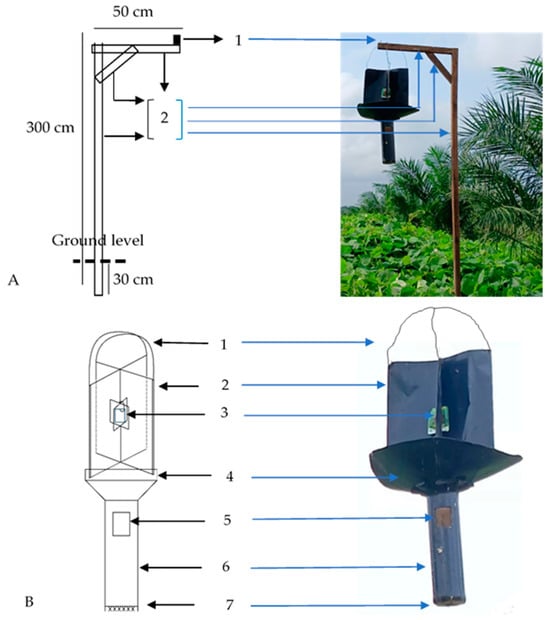
Figure 2.
Structure and installation of the Oryctes rhinoceros trap used: (A) Support pole. 1. Retaining nail; 2. wood (diameter 5 cm). (B) O. rhinoceros trap. 1. Wire (diameter 0.2 cm); 2. iron plate (35 × 40 cm), edges folded 2 cm for strength and tied with a wire to hold them together; 3. a hole (5 × 8 cm2) for placing the pheromone trap; 4. a funnel (diameter 3.5 cm), attached to the iron plate; 5. a hole (5 × 7 cm2) to collect the caught O. rhinoceros; 6. a PVC pipe (diameter 10.16 cm; length 40–50 cm); 7. mosquito net as a cover for the bottom pipe.
2.3. Laboratory Testing of the Plant Extract’s Ability as a Bioinsecticide against O. rhinoceros
2.3.1. Preparation of the Golden Rain Plant Extract
The aerial parts of the bark, stem, and leaves of the golden rain plant were cut into thin pieces and dried at 50 ± 1 °C for 24 h using a 30–1060 drying oven (Memmert, Büchenbach, Germany). The dried samples weighed a total of 5 kg and were ground and passed through a No. 80 mesh testing sieve GB/T6003.1-2012 (Xinxiang Xiyang Screening Machinery Manufacturing Co., Ltd., Xinxiang, China). Subsequently, they were macerated separately with 20 L of a 96% ethanol solution for 4 weeks. This extended maceration period aimed to ensure that all compounds in the sample were completely extracted. The resulting filtrate was then filtered using No. 42 Whatman filter paper (Whatman International Ltd., Cardiff, England) and evaporated with a rotary evaporator (IKA RV 10 Rotary Evaporator, Wertheim, Germany) until a residue was obtained. The residue was then dissolved in 500 mL of distilled water and poured into a 1000 mL separatory funnel (Schott Duran, Mainz, Germany), followed by the addition of 500 mL of ethyl acetate. This resulted in the formation of 2 phases. The separatory funnel was then closed and vigorously shaken to mix the 2 phases of the solution. Subsequently, the funnel was inverted, and the stopcock was opened to release excess vapor pressure. After this, it was left to stand to allow the separation of the 2 phases (ethyl acetate phase and water phase). The ethyl acetate phase and water phase were separated, and each was transferred to storage bottles. The water phase underwent re-extraction by mixing 500 mL of ethyl acetate into the separatory funnel. The extraction process in the separatory funnel was repeated 3 times to obtain 1500 mL of ethyl acetate phase and 500 mL of water phase. Each water and ethyl acetate fraction was evaporated with a rotary evaporator to obtain a residue. Subsequently, insecticidal activity testing against O. rhinoceros larvae and adults was conducted.
2.3.2. Experimental Design
This research was conducted using factorial experiment 4 × 2 in randomized complete block design (RCBD). The first factor was S. multijuga plant part extracts (bark, stem, leaves, and control) and second factor was the solvent used (ethyl acetate and water). The observation was performed at 24, 48, 72, and 96 h after application of the plant extracts. All treatments were repeated 4 times, and each treatment unit consisted of 5 test insects.
2.3.3. Preparation of Test Insects
One hundred forty larvae (instar 3) and 140 1-month-old adult O. rhinoceros were collected from the oil palm plantation of PT Sampoerna Agro Tbk. The larvae and adults were placed in a plastic box (64 × 43 × 14.5 cm3) containing organic material derived from decomposed oil palm plant residues. The larvae and adults were reared inside the box (using a rearing method developed by PT Sampoerna Agro Tbk) until the test was conducted.
2.3.4. Application of Plant Extracts
The concentration of each golden rain plant part extract (stem, bark, and leaves) was 10,000 ppm [14]. The application of the golden rain plant extracts was performed by taking 20 µL of the sample solution using a micropipette and dropping it onto the mouthparts of O. rhinoceros larvae or adults. This method was carried out because it was suspected that the extract from parts of the S. multijuga plant is a stomach poison to these pests. This suspicion arose based on field observations showing that O. rhinoceros dies after consuming parts of the S. multijuga plant. Subsequently, the larval and adult O. rhinoceros were placed in a plastic jar (18 cm in diameter and 20 cm in height) containing organic material derived from decomposed oil palm plant residues.
2.3.5. Observations and Data Collection
Observations of test insect mortality were conducted 24, 48, 72, and 96 h after application. Test insect mortality (percentage mortality) (M) was calculated using the following formula: the number of dead insects (n) divided by the total number of observed insects (N) multiplied by 100% [(n/N) × 100%] [15].
2.3.6. Data Analysis
The mortality data obtained were tested for homogeneity using the Barlett test and then analyzed using factorial ANOVA, followed by Duncan’s multiple range test at a significance level of 5%. Statistical analysis was performed using R Statistical Software (version 4.1.1, R Foundation for Statistical Computing, Vienna, Austria) [16].
3. Results
3.1. Senna multijuga Has the Identity of the “Hujan Emas” (Golden Rain) Plant
3.1.1. Morphological Observations Indicate That the Golden Rain Plant Is Senna multijuga
The morphological characteristics obtained revealed that the golden rain plant sample belonged to the Fabaceae family, specifically the species Senna multijuga (Rich.), H.S. Irwin and Barneby, 1982 [17]. The appearance of the golden rain plant is shown in Figure 3.

Figure 3.
Senna multijuga golden rain plant. (A) Tree appearance. (B) Branching. (C) Leaves. (D) Flowers. (E) Fruit (pod). (F) Seeds.
3.1.2. Molecular Identification Confirmed That the Golden Rain Plant Is Senna multijuga
Based on BLAST analysis, the sequencing results using the matKsenF/matKsenR primer pair showed 99.86% sequence similarity to S. multijuga (Acc. No. KX538469). The sequencing results using the rps16senF/rps16senR primer pair indicated 99.76% sequence similarity to S. multijuga var. multijuga (Acc. No. AM086962). The sequencing results using the rpl16senF/rpl16senR primer pair demonstrated 99.77% sequence similarity to S. multijuga var. multijuga (Acc. No. AM086762) (Table 2).
Table 2.
Results of the analysis using the BLAST program.
The phylogenetic analysis of the matK gene sequence indicated that the analyzed plant belonged to the same group as the reference strain S. multijuga (Acc. No KX538469) (Supplementary Figure S1A). Furthermore, the phylogenetic analysis of the rps16 and rpl16 gene sequences showed that the analyzed plant fell within the group of the reference strain S. multijuga var. multijuga ((Acc. No. AM086962) (Supplementary Figure S1B) and (Acc. No. 086762) (Supplementary Figure S1C), respectively). The molecular identification findings align with and reinforce the morphological assessment, conclusively identifying the golden rain plant as S. multijuga. The sequence data of the golden rain plant have been deposited in GenBank with the following accession numbers: PP270335 (TanBmat; MatK), PP270334 (TanBrps; rps16), and PP270333 (TanBrpl; rpl16).
3.2. Senna multijuga Has the Ability to Attract Adult O. rhinoceros
The number of beetle captures (when visiting S. multijuga) in the first three months remained below that of the pheromone trap captures, but then started to increase from the fourth month. The number of beetles collected on S. multijuga plants was higher and differed markedly in the ninth month compared to the captures in the pheromone traps (Figure 4). The observations over the 9-month period indicate that S. multijuga plants had control potential in effectively attracting O. rhinoceros beetle pests compared to the pheromone trap. The number of O. rhinoceros beetles captured on S. multijuga plants was equivalent to 1.6 times the number caught by the pheromone trap.
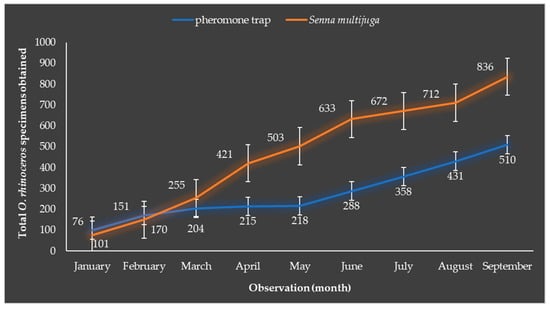
Figure 4.
The number of Oryctes rhinoceros specimens obtained from the pheromone trap and Senna multijuga plants during the 9-month observation period. The total number of O. rhinoceros beetles obtained was cumulative, representing the sum of beetles captured each month. Error bars represent the standard errors of the means of total O. rhinoceros specimens captured.
3.3. Senna multijuga Has Potential as a Bioinsecticide for O. rhinoceros
In addition to its role as an attractant for O. rhinoceros, the plant S. multijuga var. multijuga also had the ability to act as an insecticide against O. rhinoceros. The O. rhinoceros beetles that landed on it died 1–2 days after feeding on parts of the S. multijuga subsp. multijuga plant (Figure 5).

Figure 5.
Oryctes rhinoceros visiting Senna multijuga: (A) Oryctes rhinoceros perched on the stem of Senna multijuga. (B) Feeding activity on the bark of S. multijuga. (C) Dead Oryctes rhinoceros (fallen to the ground) after feeding on the bark of S. multijuga.
- Mortality of Oryctes rhinoceros larvae
Treatment with the water extract of S. multijuga bark 48 h after application showed a mortality rate of 1.25%, the water extract of S. multijuga stem showed 1.25%, the water extract of S. multijuga leaf showed 3.75%, and the ethyl acetate extract of S. multijuga leaf showed a mortality rate of 3.75%. The mortality of O. rhinoceros larvae is caused by the presence of toxic active components in the plant. In the subsequent observations, no additional insect deaths were found in any treatment (Figure 6).
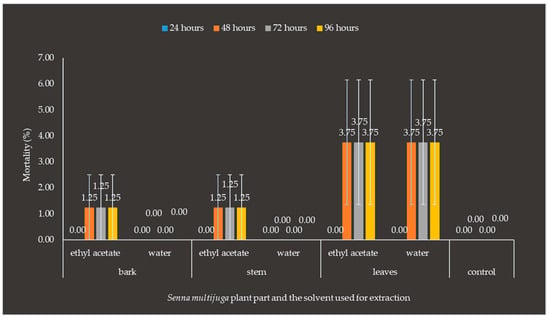
Figure 6.
Mortality of Oryctes rhinoceros larvae after the application of Senna multijuga plant extracts. Deaths were observed after the application of stem, bark, and leaf extracts using ethyl acetate solvent and after the application of leaf extracts using ethyl acetate and water solvents. Error bars represent the standard errors of the means of mortality of O. rhinoceros larvae after the application of S. multijuga plant part extract.
The statistical analysis revealed that different plant parts have a significant impact on the mortality rate of O. rhinoceros larvae. On the other hand, the type of solvent used in the experiments did not affect the mortality rate of the larvae. Furthermore, the analysis showed no significant interaction between the plant parts and the solvents used (Table 3).
Table 3.
Statistical analysis of the influence of plant part, solvent, observation time, and their interactions on the mortality of Oryctes rhinoceros larvae.
The application of S. multijuga leaf extract resulted in a significantly higher mortality rate for O. rhinoceros larvae compared to the control group, as well as the groups treated with bark and stem extracts. This finding indicates that the leaf extract possesses potent larvicidal properties that are more effective than those found in other parts of the plant. The mortality rate for larvae exposed to the leaf extract was markedly greater, highlighting the potential of S. multijuga leaves as a powerful natural insecticide (Figure 7).
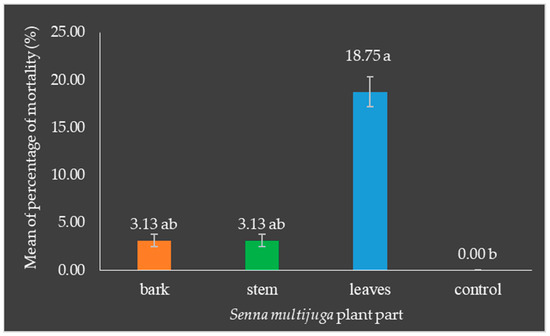
Figure 7.
Mean mortality of Oryctes rhinoceros larvae: Application of leaf extract resulted in higher mortality, which was significantly different from the control, bark, and stem extracts. The percentage mortality was analyzed using factorial ANOVA followed by Duncan’s multiple range test (DMRT) at a 5% significance level using R Statistical Software. Error bars represent the standard errors of the means of mortality of O. rhinoceros larvae. Numbers followed by the same letter are not significantly different. Different colors represent different treatments.
- Mortality of Oryctes rhinoceros beetle
The observation results 24 h after application of S. multijuga plant extract showed that the stem water extract and stem ethyl acetate extract exhibited a mortality rate of 12.5% for O. rhinoceros beetles. At 48 h after application, the S. multijuga bark ethyl acetate extract showed a mortality rate of 12.5% for O. rhinoceros beetles, and the stem water extract showed a mortality rate of 25%. The mortality of O. rhinoceros beetles was caused by the presence of toxic active components in the S. multijuga plant. In the subsequent observations, no more dead test insects were found (Figure 8).
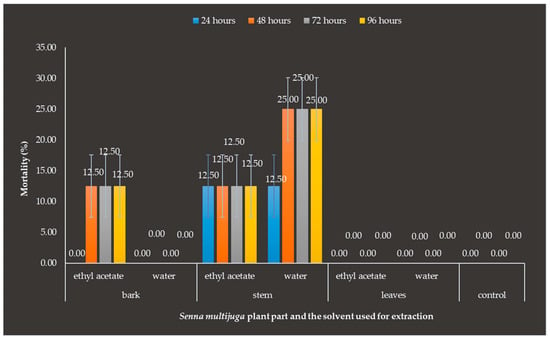
Figure 8.
Mortality of Oryctes rhinoceros beetles after the application of Senna multijuga plant extracts. Deaths were observed in the application of bark with ethyl acetate as the solvent, as well as in the stem when both ethyl acetate and water were used as the solvents. Error bars represent the standard errors of the means of mortality of O. rhinoceros beetles after the application of S. multijuga plant part extract.
Unlike the data on the mortality of O. rhinoceros larvae, the statistical analysis revealed that both the plant parts and the solvent used significantly influence the mortality of O. rhinoceros beetles. Precisely, different plant parts exhibited varying degrees of effectiveness in causing beetle mortality, suggesting that some parts of the plant contain higher concentrations of bioactive compounds that are lethal to the beetles. Additionally, the choice of solvent used for extracting these compounds also played a crucial role in determining the mortality rates. The interaction between plant parts and solvents was found to be significant, indicating that the effectiveness of the plant extracts in killing the beetles depends on the combination of both the plant part and the solvent used (Table 4).
Table 4.
Statistical analysis of the influence of plant part, solvent, observation time, and their interactions on the mortality of Oryctes rhinoceros beetles.
The extract of S. multijuga plant parts, along with the type of solvent used for extraction, had a significant impact on the mortality rate of O. rhinoceros beetles. The results indicate that different parts of the S. multijuga plant, when extracted with various solvents, vary in their effectiveness at inducing beetle mortality. The application of the S. multijuga stem extract resulted in the highest mortality rate for O. rhinoceros beetles compared to the bark and leaf extracts treatments. This suggests that the stem of S. multijuga contains particularly potent bioactive compounds or higher concentrations of these compounds that are more effective at killing the beetles (Figure 9).
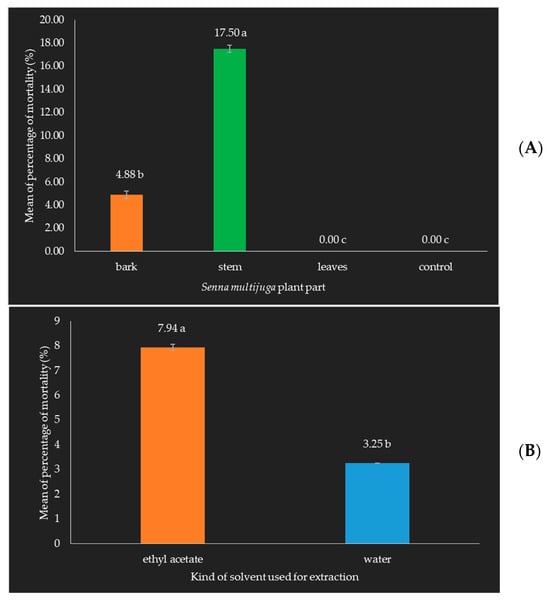
Figure 9.
Mean mortality of O. rhinoceros beetles: (A) Extract of S. multijuga plant parts. (B) Solvent used for extraction. Application of the stem extract resulted in a higher mortality rate, which was significantly different from that of the control, leaves, and bark extracts. The solvent used for extraction significantly influenced the mortality of O. rhinoceros beetles. The percentage mortality was analyzed using factorial ANOVA followed by Duncan’s multiple range test (DMRT) at a 5% significance level using R Statistical Software. Error bars represent the standard errors of the means of mortality of O. rhinoceros beetles. Numbers followed by the same letter are not significantly different. Different colors represent different treatments.
4. Discussion
Senna multijuga (false sicklepod; golden shower) (syn. Cassia multijuga) is a plant native to northern South America. It is also native to Mexico and several regions in Malaysia and Indonesia [18]. Initially known for its use as an ornamental plant along roadsides due to its attractive yellow flowers and evergreen leaves [17,19], S. multijuga is currently classified as a weed [18,20]. The plant produces seeds that are easily dispersed by wind and human activities [18]. Senna multijuga has spread and can be found in various regions worldwide due to its adaptability to various soil types. This plant is reported to be distributed in several regions in Indonesia, including Java, Sumatra, and Nusa Tenggara [18].
Various beneficial chemical compounds have been reported to be present in parts of Senna spp. plants, including S. multijuga [21,22,23,24,25]. Alkaloids are among the compounds reported to be present in S. multijuga. Alkaloids are phytoconstituents known for their biological and pharmacological activities, which can be utilized in the field of human health [20,21,22]. Pyridine alkaloids derived from the leaves of S. multijuga have been reported to act as acetylcholinesterase (AChE) inhibitors, including 7′-multijuguinone, 12′-hydroxy-7′-multijuguinone [21], 12′-hydroxy-7′-multijuguinol, 12′-hydroxy-8′-multijuguinol, methyl multijuguinate, 7′-multijuguinol, and 8′-multijuguinol [22].
In the field of plant protection, the inhibition of AChE is one of the targets of several types of active ingredients in insecticides [26,27]. AChE inhibition has been proven to be fatal for insects [28]. Organophosphates and carbamates are well-known AChE inhibitors and are widely used as insecticidal agents [27,29].
In this study, it is evident that the S. multijuga plant can attract O. rhinoceros, making it easy to capture and eliminate. The number of O. rhinoceros beetles attracted and captured by the S. multijuga plants was significantly higher than the number of insects captured by the pheromone traps. The average weekly capture of O. rhinoceros by the pheromone traps was 12 O. rhinoceros beetles, while the S. multijuga plants were capable of capturing 22, despite the fact that 299 pheromone traps were installed compared to only 25 S. multijuga plants. This indicates that the S. multijuga plant has the potential to be used as an attractant and trap for O. rhinoceros beetles.
The use of pheromone traps to control O. rhinoceros is one method that has been reported to be quite effective [5,30]. The direct capture of adult O. rhinoceros can reduce the population and, at the same time, suppress the overall population by reducing the number of reproductive insects. No negative impacts are caused by this control method [30].
Since its discovery in the 1990s, the male aggregation pheromone of O. rhinoceros, ethyl 4-methyloctanoate (E4-MO), has been widely used for the detection, monitoring, and trapping of this pest insect [31]. In addition to detecting the presence of and monitoring O. rhinoceros, the use of attractants has proven effective in controlling this pest [32]. The search for plant extracts that can be used as attractants for O. rhinoceros is currently underway. The potential use of pineapple skin extract has been reported by [33]. The use of mature coconut trees has also been reported by [34]. The results of this current study indicate that S. multijuga has the potential to be used as an O. rhinoceros attractant. This has not been investigated or reported until now. We are conducting an ongoing study to determine the chemical compounds contained in different parts of the S. multijuga plant that may have the ability to attract O. rhinoceros.
Field observations also indicate that some O. rhinoceros beetles were found dead after visiting and feeding on parts of the S. multijuga plants. The deceased O. rhinoceros beetles fell to the ground around the plant. This suggests the possibility that the plant also acts as an insecticide. Laboratory testing was conducted to confirm this assumption.
Testing of the S. multijuga plant extract’s ability as a bioinsecticide was conducted on O. rhinoceros beetles and larvae. Three parts of the plant (leaves, bark, and stem) were further investigated for their potential as bioinsecticides. Extraction was performed using two solvents, namely, water and ethyl acetate, to obtain compounds from different parts of the S. multijuga plant that are polar and non-polar in nature.
The test results indicate that each tested part of the plant has insecticidal capability against O. rhinoceros. The presence of pyridine alkaloids in S. multijuga [21,22] is considered one of the reasons for its insecticidal properties. The existence of pyridine alkaloids has been reported in the leaves of S. multijuga [21,22]. However, the presence of these alkaloids in the stem and bark is not yet known. Further studies will be conducted to determine the chemical compounds present in the stem and bark of S. multijuga, including the presence and types of pyridine alkaloids.
Some S. multijuga plant part extracts appeared to not cause death in O. rhinoceros. This is suspected to be due to O. rhinoceros being able to degrade (neutralize) the toxic effect of the plant extract [35]. Some insects are reported to have genes capable of expressing resistance to insecticides [36,37]. Three major enzyme families, namely, cytochrome P450s (P450s), carboxylesterases (COEs), and glutathione-S transferases (GSTs), have been reported to be responsible for insecticide resistance [38,39]. Insecticide resistance is also reported to be influenced by symbionts present in the insect’s body, although not all toxins that enter can be neutralized [40,41]. Symbionts that have been reported to play a role in triggering insect resistance to insecticides include Arsenophonus [42], Burkholderia sp. [43], Citrobacter sp. [44], Enterococcus sp. [45], Pseudomonas melophtora [46], Symbiotaphrina kochi [47], and Wolbachia [48]. In addition to the type of toxin, the population of symbionts present in the insect and the level of toxin concentration also affect the level of insect resistance [40,49]. The presence of resistance genes and symbionts capable of neutralizing insecticides, especially in O. rhinoceros, would be an interesting area of further study.
Although extracts from different parts of the S. multijuga plant can cause mortality in both larvae and adult O. rhinoceros, the resulting mortality percentages are low, ranging from 1.25% to 3.75% for larvae and 12.5% to 25% for O. rhinoceros beetles. This condition raises the suspicion that S. multijuga plays a more dominant role as an attractant rather than as an insecticide. Further investigation regarding the potential of S. multijuga as an attractant will be conducted, including the search for active ingredients capable of attracting adult O. rhinoceros. Additionally, the identification of compounds with insecticidal properties against O. rhinoceros larvae and adults will also be pursued.
5. Conclusions
In this study, we observed that S. multijuga demonstrates attractant and bioinsecticidal capabilities against O. rhinoceros. In particular, S. multijuga exhibited a greater ability to attract O. rhinoceros compared to the widely used synthetic pheromone ethyl 4-methyloctanoate. Its bioinsecticidal activities against both larvae and adults were also confirmed in a laboratory investigation. The leaf extract demonstrated insecticidal activity against larvae, while the stem and bark extracts showed insecticidal effects against both larvae and adults. The chemical compound responsible for the attractant and insecticidal activities remains unclear at this time. Further investigations will be conducted to characterize and identify the chemical components of the leaf, stem, and bark extracts and evaluate their effects on the mortality of and ability to attract O. rhinoceros.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14091477/s1, Table S1. Reference used in this study; Figure S1. Analysis of the matK gene sequence of the golden rain plant (TanBmat; Acc. No. PP270335) using the maximum likelihood method and the Kimura 2-parameter model [50] with 1000x bootstrap. The dendrogram was created using the MEGA 11 program for Windows [13]. A. Dendrogram based on the matK gene sequence analysis. The golden rain plant appears to be in the same group as Senna multijuga (KX538469). B. Dendrogram based on the rps16 gene. The golden rain plant (TanBrps; Acc. No. PP270334) appears to be in the same group as Senna multijuga var. multijuga (Acc. No. AM086962). Coffea arabica (Acc. No. GW447360) is used as an outgroup. C. Dendrogram based on the rpl16 gene. The golden rain plant (TanBrpl; Acc. No. PP270333) appears to be in the same group as Senna multijuga var. multijuga (Acc. No. AM086962). Libidibia coriaria is used as an outgroup in all three dendrograms with Acc. No KM219789.1 (matK), NC_026677.1 (rpl16) and KF522107.1 (rps16).
Author Contributions
Conceptualization, R.W., S.A., S.D.A., D.A., S.S., Y.F. and R.S.; methodology, R.W., S.A., S.D.A., S.S., Y.F. and R.S.; software, R.S.; validation, R.W., S.A., D.A., R.H., S.S. and R.S.; formal analysis, S.S., R.S., Y.F., R.H., S.A. and S.D.A.; investigation, S.A., S.D.A. and R.S.; resources, R.W., S.A., S.D.A., A.A., S.S. and D.A.; data curation, S.A., S.D.A., A.A., Y.F. and R.S.; writing—original draft preparation, R.S. and Y.F.; writing—review and editing, R.W., S.A., S.D.A., R.H. and S.S.; visualization, R.S. and Y.F.; supervision, D.A. and R.H.; project administration, S.D.A.; funding acquisition, R.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by PT Sampoerna Agro Tbk, Jakarta, Indonesia, grant number 133/BSM/III/23/RO/CA.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets used or analyzed in this study are available from the corresponding author on reasonable request.
Acknowledgments
We express our gratitude to the Faculty of Agriculture, University of Lampung, for granting us permission to use their research facilities during this study. We also extend our thanks to PT Sampoerna Agro Tbk, Jakarta, Indonesia, for providing funding, materials, and sequencing support for this study. Additionally, we thank Silva Abraham, at the Directorate of Scientific Collection Management, National Research and Innovation Agency (BRIN), Cibinong, Bogor, West Java, Indonesia, for conducting the morphological identification of the S. multijuga plants.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bedford, G.O. Biology, ecology, and control of palm rhinoceros beetles. Annu. Rev. Entomol. 1980, 25, 309–339. [Google Scholar] [CrossRef]
- Hochberg, M.E.; Waage, J.K. A model for the biological control of Oryctes rhinoceros (Coleopteran: Scarabaeidae) by means of pathogens. J. Appl. Ecol. 1991, 28, 514–531. [Google Scholar] [CrossRef]
- Kalidas, P. Pest problems of oil palm and management strategies for sustainability. Agrotechnology 2012, SS11, 1. [Google Scholar] [CrossRef]
- Lever, R. Pests of the Coconut Palm; Food and Agriculture Organization: Rome, Italy, 1969; pp. 125–133. [Google Scholar]
- Manjeri, G.; Muhamad, R.; Tan, S.G. Oryctes rhinoceros beetles, an oil palm pest in Malaysia. Annu. Res. Rev. 2014, 4, 3429–3439. [Google Scholar] [CrossRef]
- Kalshoven, L.G.E. The Pests of Crops in Indonesia; Ichtiar Baru-Van Hoeve: Jakarta, Indonesia, 1981; pp. 463–468. [Google Scholar]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef]
- Kelchner, S.A. Group II introns as phylogenetic tools: Structure, function, and evolutionary constraints. Am. J. Bot. 2002, 89, 1651–1669. [Google Scholar] [CrossRef] [PubMed]
- Marazzi, B.; Endress, P.K.; Queiroz, L.P.; Conti, E. Phylogenetic relationships within Senna (Leguminosae, Cassiinae) based on three chloroplast DNA regions: Patterns in the evolution of floral symmetry and extrafloral nectaries. Am. J. Bot. 2006, 93, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Suharjo, R.; Swibawa, G.; Prasetyo, J.; Fitriana, Y.; Danaatmadja, Y.; Budiawan, A.; Roberts, S.; Noorhajati, N.; Amad, M.; Thines, M. Peronosclerospora australiensis is a synonym of P. maydis, which is widespread on Sumatra, and distinct from the most prevalent Java maize downy mildew pathogen. Mycol. Prog. 2020, 19, 1309–1315. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 4, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Maimunah, D.; Irwan, S.N.R.; Indradewa, D. Pertumbuhan widelia (Wedelia trilobata (L) Hitchc) pada tingkat naungan berbeda di jalur hijau Kota Yogyakarta [Widelia (Wedelia trilobata (L) Hitchc) growth at different shading levels in the roadside greenery of Yogyakarta City]. J. Ilmu Pertan. Indones. 2020, 25, 547–555. [Google Scholar] [CrossRef]
- Salinas-Sánchez, D.O.; Flores-Franco, G.; Avilés-Montes, D.; Valladares-Cisneros, M.G.; Arias-Ataide, D.M.; Mendoza-Catalán, M.A.; Sotelo-Leyva, C. Bioactivity of a linoleic acid–rich fraction of Ricinus communis L. (Euphorbiaceae) leaves against the yellow sugarcane aphid, Sipha flava (Hemiptera: Aphididae). J. Food Prot. 2021, 84, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Fitriana, Y.; Purnomo; Hariri, A.M. Uji efikasi ekstrak gulma siam terhadap mortalitas hama pencucuk buah kakao (Helopeltis spp.) di laboratorium) [Efficacy test of siam weed extract on the mortality of cocoa mirid bugs (Helopeltis spp.) in the laboratory]. J. HPT. Tropika. 2012, 12, 85–91. [Google Scholar] [CrossRef]
- R Core Team. R: A Languane and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 14 January 2024).
- Irwin, H.S.; Barneby, R.C. The American Cassiinae: A Synoptical Revision of Leguminosae Tribe Cassieae Subtribe Cassiinae in the New World; New York Botanical Garden: New York, NY, USA, 1982; pp. 1–918. [Google Scholar]
- Datiles, M.J.; Acevedo-Rodrígue, P. Senna multijuga (November Shower); CABI Compendium: Oxford, UK, 2014. [Google Scholar] [CrossRef]
- Ribeiro-costa, C.S.; Reynaud, D.T. Bruchids from Senna multijuga (Rich) I. & B (Caesalpinaceae) in Brazil with description of two new species. Coleopt. Bull. 1998, 52, 245–252. [Google Scholar]
- Local Land Services. North Coast Regional Strategic Weed Management Plan 2023–2027; North Coast Local Land Services: New South Wales, Australia, 2022; pp. 1–98. [Google Scholar]
- Serrano, M.A.; Pivatto, M.; Francisco, W.; Danuello, A.; Regasini, L.O.; Lopes, E.M.; Lopes, M.N.; Young, M.C.; Bolzani, V.S. Acetylcholinesterase inhibitory pyridine alkaloids of the leaves of Senna multijuga. J. Nat. Prod. 2010, 73, 482–484. [Google Scholar] [CrossRef] [PubMed]
- Francisco, W.; Pivatto, M.; Danuello, A.; Regasini, L.O.; Baccini, L.R.; Young, M.C.; Lopes, N.P.; Lopes, J.L.; Bolzani, V.S. Pyridine alkaloids from Senna multijuga as acetylcholinesterase inhibitors. J. Nat. Prod. 2012, 75, 408–413. [Google Scholar] [CrossRef]
- Oladeji, O.S.; Adelowoc, F.E.; Oluyoria, A.P. The genus Senna (Fabaceae): A review on its traditional uses, botany, phytochemistry, pharmacology and toxicology. S. Afr. J. Bot. 2021, 138, 1–32. [Google Scholar] [CrossRef]
- Alshehri, M.M.; Quispe, C.; Herrera-Bravo, J.; Sharifi-Rad, J.; Tutuncu, S.; Aydar, E.F.; Topkaya, C.; Mertdinc, Z.; Ozcelik, B.; Aital, M.; et al. A review of recent studies on the antioxidant and antiinfectious properties of senna plants. Oxidative Med. Cell. Longev. 2022, 6025900. [Google Scholar] [CrossRef]
- Zibaee, E.; Javadi, B.; Sobhani, Z.; Akaberi, M.; Farhadi, F.; Amiri, M.S.; Baharara, H.; Sahebkarf, A.; Emami, S.A. Cassia species: A review of traditional uses, phytochemistry and pharmacology. Pharmacol. Res. Mod. Chin. Med. 2023, 9, 100325. [Google Scholar] [CrossRef]
- Hazuki, I.N.; Shukor, M.Y. Acetylcholinesterase as an in vitro assay for insecticides: A mini review. J. Environ. Microbiol. Toxicol. 2018, 6, 7–12. [Google Scholar] [CrossRef]
- Araújo, M.F.; Castanheira, E.M.S.; Sousa, S.F. The buzz on insecticides: A review of uses, molecular structures, targets, adverse effects, and alternatives. Molecules 2023, 28, 3641. [Google Scholar] [CrossRef] [PubMed]
- Rajashekar, Y.; Raghavendra, A.; Bakthavatsalam, N. Acetylcholinesterase inhibition by biofumigant (Coumaran) from leaves of Lantana camara in stored grain and household insect pests. BioMed Res. Int. 2014, 187019. [Google Scholar] [CrossRef]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Persp. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Bedford, G.O. Advances in the control of rhinoceros beetle, Oryctes rhinoceros in oil palm. J. Oil Palm Res. 2014, 26, 183–194. [Google Scholar]
- Bedford, G.O. Biology and management of palm dynastid beetles: Recent advances. Annu. Rev. Entomol. 2013, 58, 353–372. [Google Scholar] [CrossRef]
- Paudel, S.; Jackson, T.A.; Mansfield, S.; Ero, M.; Moore, A.; Marshall, S.D.G. Use of pheromones for monitoring and control strategies of coconut rhinoceros beetle (Oryctes rhinoceros): A review. Crop Prot. 2023, 17, 106400. [Google Scholar] [CrossRef]
- Munawaroh, L.; Wirman, S.P.; Fitrya, N.; Syahputra, R.F.; Gesriantuti, N. Pendekatan baru pegendalian kumbang tanduk secara otomatis pada tanaman kelapa sawit yang ditanam ulang: Perangkap feromon kulit nanas [A novel approach for automated rhinoceros beetle control in oil palm replanting: Pineapple peel-derived trap]. J. Ilm. Pertan. 2013, 20, 200–208. [Google Scholar] [CrossRef]
- Alfiler, A.R.R. Increased attraction of Oryctes rhinoceros aggregation pheromone, ethyl 4-methyloctanoate, with coconut wood. CORD 1999, 15, 34. [Google Scholar] [CrossRef]
- Huber, M.; Roder, T.; Irmisch, S.; Riedel, A.; Gablenz, S.; Fricke, J.; Rahfeld, P.; Reichelt, M.; Paetz, C.; Liechti, N.; et al. A beta-glucosidase of an insect herbivore determines both toxicity and deterrence of a dandelion defense metabolite. Elife 2021, 10, e68642. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Horne, I.; Sutherland, T.D.; Russell, R.J. The genomics of insecticide resistance. Genome Biol. 2003, 4, 202. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H. The molecular genetics of insecticide resistance. Genetics 2013, 194, 807–815. [Google Scholar] [CrossRef]
- Ranson, H.; Claudianos, C.; Ortelli, F.; Abgrall, C.; Hemingway, J.; Sharakhova, M.V.; Unger, M.F.; Collins, F.H.; Feyereisen, R. Evolution of supergene families associated with insecticide resistance. Science 2002, 298, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-C.; Chien, T.-Y.; Hu, C.-C.; Chen, M.-J.M.; Wu, W.-J.; Feng, H.-T.; Haymer, D.S.; Chen, C.-Y. Discovery of genes related to insecticide resistance in bactrocera dorsalis by functional genomic analysis of a de novo assembled transcriptome. PLoS ONE 2012, 7, e40950. [Google Scholar] [CrossRef] [PubMed]
- Blanton, A.G.; Peterson, B.F. Symbiont-mediated insecticide detoxification as an emerging problem in insect pests. Front. Microbiol. 2020, 11, 547108. [Google Scholar] [CrossRef] [PubMed]
- Rupawate, P.S.; Roylawar, P.; Khandagale, K.; Gawande, S.; Ade, A.B.; Jaiswal, D.K.; Borgave, S. Role of gut symbionts of insect pests: A novel target for insect-pest control. Front. Microbiol. 2023, 14, 1146390. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Chen, M.; Yue, L.; Xing, K.; Li, T.; Kang, K.; Liang, Z.; Yuan, L.; Zhang, W. A distinct strain of Arsenophonus symbiont decreases insecticide resistance in its insect host. PLoS Genet. 2018, 14, e1007725. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. An ancient but promiscuous host-symbiont association between Burkholderia gut symbionts and their heteropteran hosts. ISME J. 2011, 5, 446–460. [Google Scholar] [CrossRef]
- Cheng, D.; Guo, Z.; Riegler, M.; Xi, Z.; Liang, G.; Xu, Y. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 13. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut microbiota mediate insecticide resistance in the diamondback moth, Plutella xylostella (L.). Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef]
- Boush, G.M.; Matsumura, F. Insecticidal degradation by Pseudomonas melophthora, the bacterial symbiote of the apple maggot. J. Econ. Entomol. 1967, 60, 918–920. [Google Scholar] [CrossRef]
- Shen, S.K.; Dowd, P.F. Detoxification spectrum of the cigarette beetle symbiont Symbiotaphrina kochii in culture. Entomol. Exp. Appl. 1991, 60, 51–59. [Google Scholar] [CrossRef]
- Berticat, C.; Rousset, F.; Raymond, M.; Berthomieu, A.; Weill, M. High Wolbachia density in insecticide-resistant mosquitoes. Proc. R. Soc. London Ser. B 2002, 269, 1413–1416. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Somani, J.; Roy, S.; Babu, A.; Pandey, A.K. Insect microbial symbionts: Ecology, interactions, and biological significance. Microorganisms 2023, 11, 2665. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 11–20. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).