Abstract
Alternariol (AOH) and alternariol monomethyl ether (AME) are secondary metabolites produced by fungi belonging to the genus Alternaria, which generally contaminate fruits but also cereal crops and vegetables. The objective of this study was to investigate if the co-exposure of the swine epithelial intestinal cell line (IPEC-1) to a mixture of mycotoxins would cause an increase in toxicity as compared with exposure to a single toxin. The effects of individual toxins as well as those of their combination (1:1 ratio), in a range of 1–250 μM, were assessed in vitro for the cell viability of proliferating IPEC-1 cells and then on parameters related to the oxidative stress. Our results indicate that both AOH and AME significantly decreased the IPEC-1 cell viability, but the cytotoxicity induced by the AOH + AME combination was significantly higher than that induced by the exposure to the individual toxins. The main interaction type ranged from slight synergy for the AOH-AME combination affecting 25% of cell viability (CI = 0.88), which evolved into a synergistic effect for a higher level of cytotoxicity IL50 (CI = 0.41) and a strong synergistic effect at IL90 (CI = 0.10). In addition, we investigated the effects of two low concentrations (2.5 μM and 5 μM) of AOH and AME mycotoxins administered individually or in combination on oxidative stress in IPEC-1 cells. Both AOH and AME can induce an increase in reactive oxygen species—ROS (+) cells%—and oxidative damage in porcine IPEC-1 cells. At least an additive effect was observed when the cells were exposed to the combination of AOH-AME, consisting of an increase in the percentage of ROS (+) cells and the oxidation of lipids, proteins, and DNA as compared with the individual toxin effect. A breakdown of the antioxidant defense was observed in IPEC-1 cells after the exposure to individual toxins, related to the decrease in the activity of antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT), but no additive or synergic effect resulted after the exposure to the mixture of the toxins. In conclusion, our data indicate that both AOH and AME interfere with cell proliferation and oxidative stress. Moreover, the exposure of IPEC-1 cells to the combination of AOH and AME mycotoxins had a dose-dependent synergistic effect on IPEC-1 cell viability. Also, the oxidative damage induced in IPEC-1 cells by the combination of AOH and AME was stronger than the effects of individual toxins. However, the signaling pathways responsible for the toxicity of AOH, AME, and their combinations need further investigations in order to provide important data for risk assessments in swine in the case of the contamination of feed with Alternaria toxins.
1. Introduction
Alternaria fungi are plant pathogens that can damage field crops or can cause crop spoilage after harvesting, leading to significant economic losses [1]. They are also common allergens in humans and cause severe allergic reactions [2]. Additionally, the molds of the Alternaria genus are able to produce mycotoxins that represent a threat to food safety since some of them have been proven to be harmful to humans and animals [3]. Most of the toxicological studies published to date were focused on the analysis of toxic effects of a single mycotoxin without taking into consideration the exposure to combinations of Alternaria toxins as well as their possible interactions—additive, synergistic, or antagonistic [4]. However, the simultaneous contamination of food and feed commodities with mycotoxins is a rule rather than an exception. For example, a survey program monitoring the mycotoxin occurrence in feed and feed raw materials has shown a case of contamination with 139 fungal metabolites in 83 analyzed feed samples [5]. In this survey, alternariol (AOH) and alternariol monomethyl ether (AME) were the most frequently occurring Alternaria mycotoxins, with AOH being detected in 80% of samples in a range between 2.8 and 221 μg/kg and AME in 82% of samples in a range of 1.4–733 μg/kg [5]. Different surveys have indicated that AOH and AME occur in the same food commodities mostly in similar concentrations ranges (1:1 ratio) [6,7]. Exposure to Alternaria mycotoxins was responsible for cytotoxic, genotoxic, mutagenic, and carcinogenic effects, as demonstrated by in vitro toxicological experiments involving bacterial and mammalian cells [8,9]. Oxidative stress seems to be one of the principal mechanisms involved in Alternaria mycotoxin toxicity, the toxins responsible for the oxidation of proteins, lipids, and DNA and for an increase in the oxidative status, with a modulation of the redox cellular balance [10,11,12]. Few studies have investigated the effect of Alternaria mycotoxin mixtures, and all of them were realized in vitro using human cell lines. It can be expected that AOH and AME, mycotoxins with a common mode of action, may have additive or synergic effects [4,13]. Indeed, several studies have indicated that the AOH-AME combination has additive and synergic effects on cell viability as compared with the exposure to a single toxin [4,14,15]. However, another recent study indicated that the exposure of a human gastric epithelial cell line to a mixture of AME and AOH results in antagonistic cytotoxic effects at low concentrations that turn into additive and synergistic effects at higher toxin concentrations [16]. Among farm animals, swine are particularly exposed to the effects of mycotoxin through their alimentation based on cereals [17]. An analysis of 1141 pig feed samples from the years 2014 to 2019 has shown a contamination with a median concentration of 4 μg/kg and a prevalence of 50.7% for AOH and a median concentration of 3 μg/kg and a prevalence of 40.3% for AME [18]. Currently, Alternaria toxins are not regulated in food and feed in Europe, and the EFSA considers that the exposure to Alternaria toxins through the diet is higher than the toxicological threshold of concern (EFSA 2022). As also stated by the EFSA, there is a need for new research to assess the risks posed by Alternaria mycotoxins in farm animals, as the knowledge on their toxic effects as well as the occurrence data in feed are insufficient to establish the health risks [8]. Many studies in the literature have investigated the toxicity of individual toxins using mainly in vitro models. For example, our previous in vitro study has shown that AME triggers apoptosis and oxidative damage in swine intestinal epithelial cells [19]. To date, according to our knowledge, there are no risk assessment studies in the literature concerning the effects of the co-exposure to Alternaria mycotoxins in farm animals using either in vitro or in vivo studies. Therefore, the present study aims to provide important data concerning the effect of individual and binary combinations of the two most frequently occurring Alternaria mycotoxins, AOH and AME, in swine, using a pig epithelial intestinal cell line (IPEC-1), as the first cells exposed to the toxins that enter the body.
2. Materials and Methods
Reagents. The reagents and kits were purchased mainly from Sigma Aldrich (St. Louis, MO, USA), if no other company is specified.
Cell cultures. IPEC-1 (2 × 105 cells/mL), between the passages 125 and 133, were grown in 24- or 96-well plates in DMEM/F12 complete media, as already described [19]. After 24 h, the cells were treated for another 24 h with individual toxins (AOH or AME) or with their combination (1:1 ratio), in a 1–250 μM range for assessing cell viability. As shown by cytotoxicity studies, concentrations of toxins higher than 5 µM have a high cytotoxicity. For this reason, concentrations lower than 5 μM were used for the assessment of the parameters related to the oxidative stress: reactive oxygen species—ROS (+) cells%—antioxidant enzyme activity, and protein, lipid, and DNA oxidation.
Toxins. Pure AOH and AME (>98%) were dissolved in DMSO, aliquoted, and stored at −20 °C prior to dilution in cell culture medium. The final DMSO concentration was no greater than 1%.
Cell viability. The cytotoxic effect of both individual and a combination of mycotoxins was assessed on IPEC-1 subjected to 24 h treatment with 0 to 250 μM AOH, AME, or a combination thereof in a 1:1 ratio according to the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] test. The colorimetric reaction was based on the reduction of MTT to purple formazan crystals by metabolically active cells, as described in previous research [19]. The data are expressed as a percentage of the control.
Analysis of mycotoxin interactions with regard to cell viability. The types of interactions that affect IPEC-1 cell viability after exposure to a binary mixture of mycotoxins were assessed according to the Chou and Talalay method [20] and CompuSyn1.0 (ComboSyn, Inc., Washington, DC, USA, 2012), a program for the quantitation of synergism in drug combinations, as previously described [21]. The Chou–Talalay method is based on the median-effect equation; together with the isobologram method, it provides a visual assessment of the interactions between the toxins in the mixture. Said interactions were analyzed by plotting the isobolograms and calculating the combination index (CI) values for binary combinations of AOH and AME.
Reactive oxygen species. IPEC-1 cells were exposed to 0–5 μM AOH, AME, and a combination of the two (in a 1:1 ratio) for 24 h. Quantification of ROS (+) cells% was performed on treated IPEC-1 cells via flow-cytometry analysis, as previously described [19], using a Muse Cell Analyzer system and a specific kit (Muse Oxidative Stress Kit) (Merck KGaA, Darmstadt, Germany). The data are expressed as the cells’ ROS (+) percentage.
Antioxidant enzyme activity. IPEC-1 cells were exposed to AOH, AME, and a combination of the two (in a 1:1 ratio) at a concentration of 0 to 5 μM for a 24 h period. The activities of catalase (CAT), glutathione peroxidase (GPx), and superoxide dismutase (SOD) were analyzed based on IPEC-1 cell lysates using specific kits (Cayman Chemical Company—Ann Arbor, MI, USA) according to the manufacturer’s instructions [19].
Assessment of lipid peroxidation. Lipid oxidation was determined in IPEC-1 cells that underwent 24 h of exposure to AOH, AME, and a combination of the two (ratio 1:1) at a concentration of 0 to 5 μM from cell sample lysates in phosphate-buffered saline. After the protein concentrations in the cell lysates had been identified and normalized to 1000 ng/mL protein, thiobarbituric acid-reactive substances (TBARSs) were measured according to the established protocol [19]. The results were expressed as nmol malondialdehyde/mL.
Assessment of protein oxidation. IPEC-1 cells were exposed to AOH, AME, and a combination of the two (ratio 1:1) at a 0 to 5 μM concentration for 24 h. Protein oxidation was determined in cell lysates, and the reactions between 2,4-dinitrophenyl hydrazine (Merck KGaA, Darmstadt, Germany) and protein carbonyls were monitored spectrophotometrically according to a previously described method [19]. The results are expressed in nmol/mg of carbonyl content.
Assessment of DNA oxidation. DNA oxidation was measured in IPEC-1 samples after the cells underwent 24 h of exposure to AOH, AME, and a combination of the two (in a 1:1 ratio) in concentrations ranging from 0 to 5 μM. The immune–enzymatic method was implemented according to the established protocol [19] using a DNA Oxidative Damage ELISA Kit (Cayman Chemical Company, Ann Arbor, MI, USA). The results are expressed in pg/mL of 8-hydroxy-2′-deoxyguanosine.
Statistical analysis. The cell viability, ROS (+) cells %, activity of SOD, CAT, and GPX, lipid peroxidation, protein oxidation, and DNA oxidation results represent the mean ± standard error of the mean (SEM) of three independent experiments, with two replicates for each experiment (n = 6). Differences among groups were tested using one-way ANOVA of the GraphPad Prism 9 software (GraphPad Software, Boston, MA, USA, 2020), followed by Fisher’s PSLD test. Statistical significance between the experimental groups was considered at a p-value < 0.05.
3. Results
3.1. Effects of AOH, AME, and a Combination of the Two on IPEC-1 Cell Viability
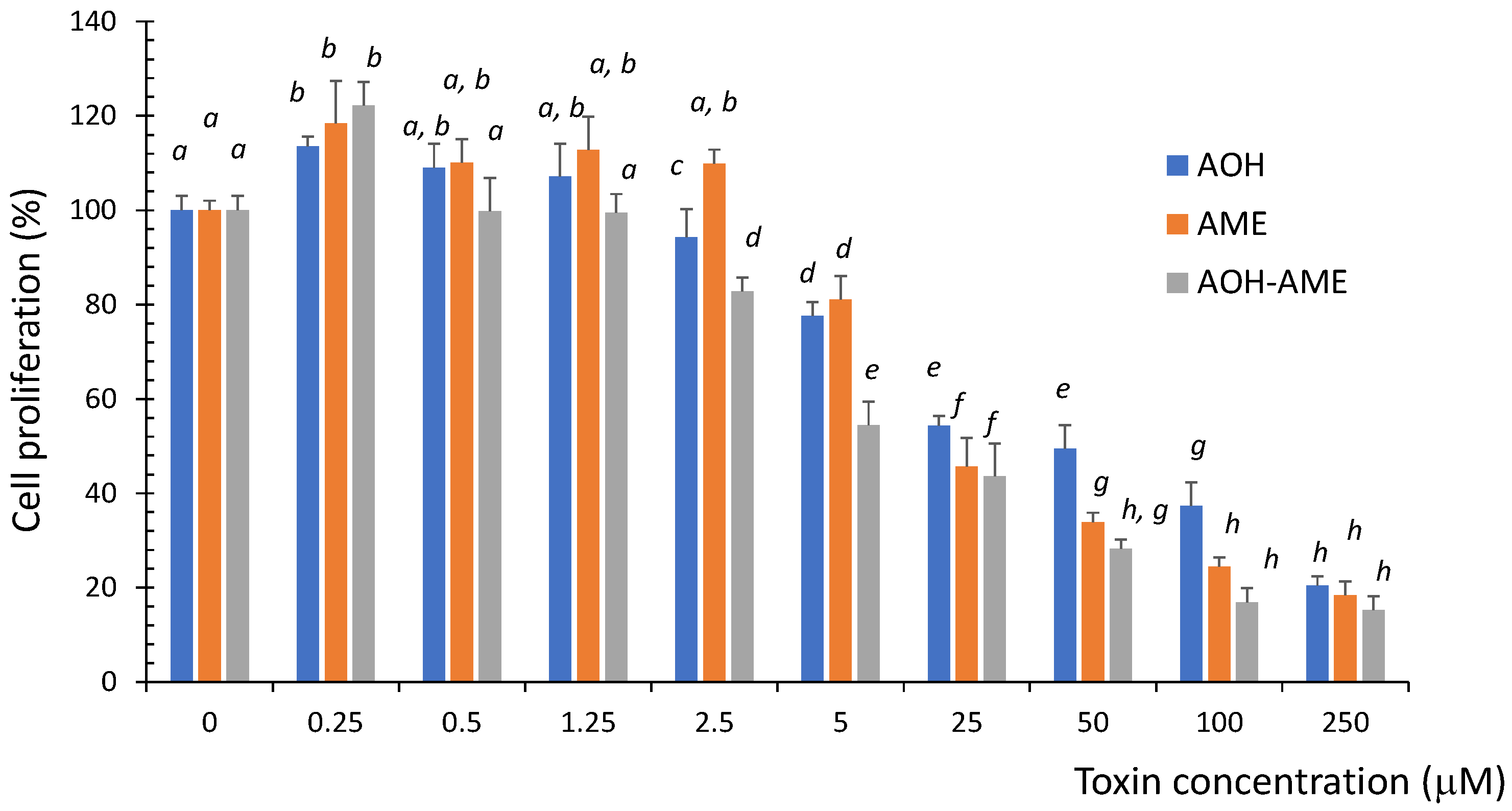
Proliferating IPEC-1 cells were exposed to different concentrations of AOH, AME, or a combination of the two in a ratio of 1:1, and the cytotoxicity was evaluated using an MTT test. As shown in Figure 1, exposure to both the individual toxins as well as a combination of the two significantly decreased the cell viability in a dose-dependent manner. The cytotoxicity induced by the AOH + AME combination was significantly higher than that induced by the exposure to AOH or AME alone, especially for concentrations above 2.5 μM.
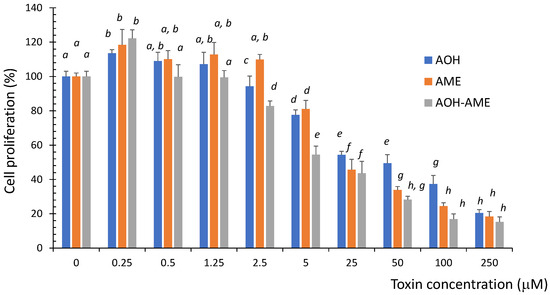
Figure 1.
The effect of exposure to 0 to 250 μM of AOH, AME, and a combination of the two on the viability of IPEC-1 cells; the results represent the mean ± standard error of the mean (SEM) of three independent experiments, n = 6. Differences among groups were tested using one-way ANOVA followed by Fisher’s PSLD test. a–h indicate significant differences between treatments (p < 0.05).
3.2. Types of Interactions between Mycotoxins Related to IPEC-1 Cell Viability
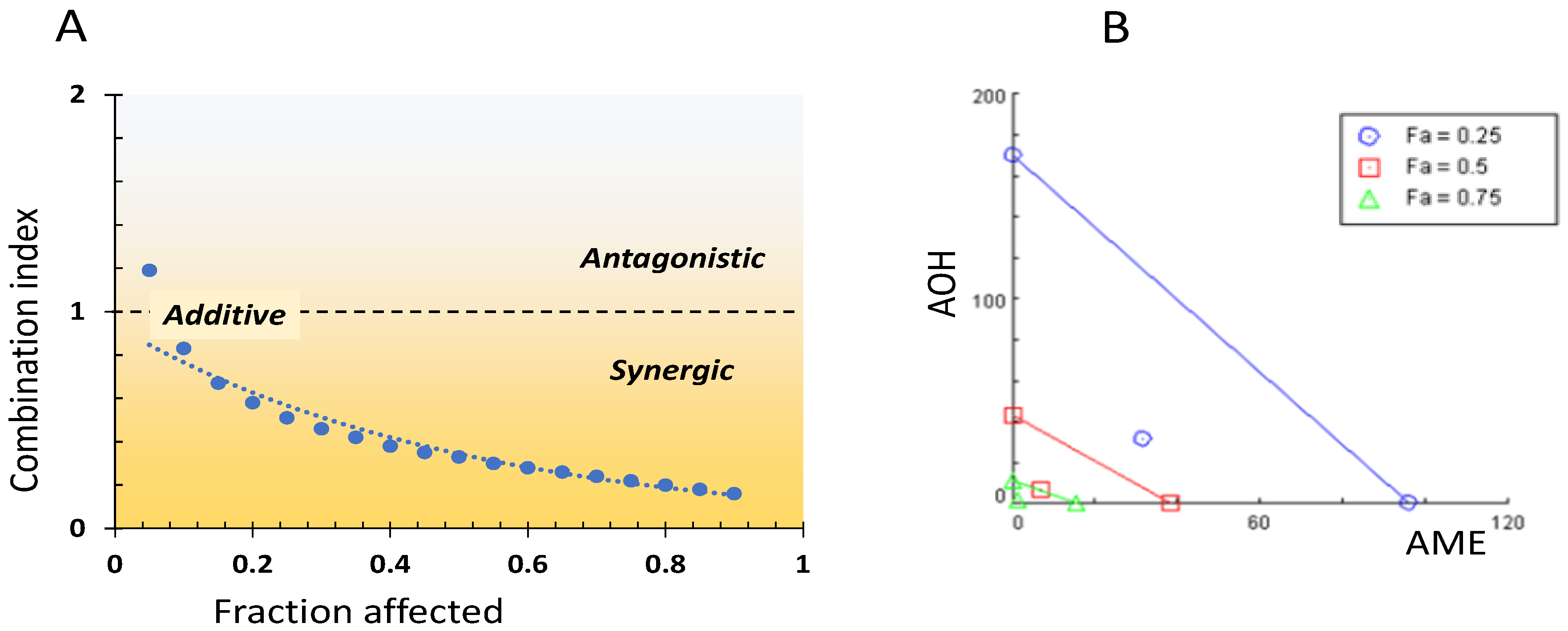
Isobologram and a combination index method were used to assess the types of interactions between AOH, AME, and AOH + AME. These interactions were categorized as additive, synergic, or antagonistic. The values for the combination index for the binary mycotoxin combination that induces an inhibition level (IL) of 25, 50, and 75%, as well as the dose reduction index necessary for the corresponding reduction in cell viability, are presented in Table 1. With the exception of the lowest fa (0.05), for which there is an antagonistic effect, in the whole range of fa (fractions affected) the AOH + AME combination has CI values below one, which indicate an additive effect evolving into a synergistic effect. This changes with the toxin concentration, with CI values decreasing from 0.83 for fa = 0.10 to 0.16 for fa = 0.9.

Table 1.
Combination and dose reduction index values for the combination of AOH and AME in relation to cell viability.
As shown in Table 1, the DRI (dose reduction index) ranges from 5.3- to 8.96-fold for AOH and 3.03- to 19.8-fold for AME indicating the reduction factor for the concentration of each mycotoxin in the combination as compared with the concentration of the individual toxin due to the synergistic action of the toxin mixture. The increase in the DRI values for inhibition levels of 25, 50, 75, and 90% indicates that the synergic effect increases with the concentration of the toxins.
The dose–effect curve parameters (Dm and m) for the cytotoxic effects of the mycotoxins are presented in Table 2. The highest median-effect dose required to induce a 50% inhibition of the cell was calculated for AOH, while the calculated Dm was 1.12 times lower for AME and 3.17 lower for the combination of toxins.
Table 2.
Dose–effect relationship parameters for AOH, AME, and their binary combinations in relation to cell viability.
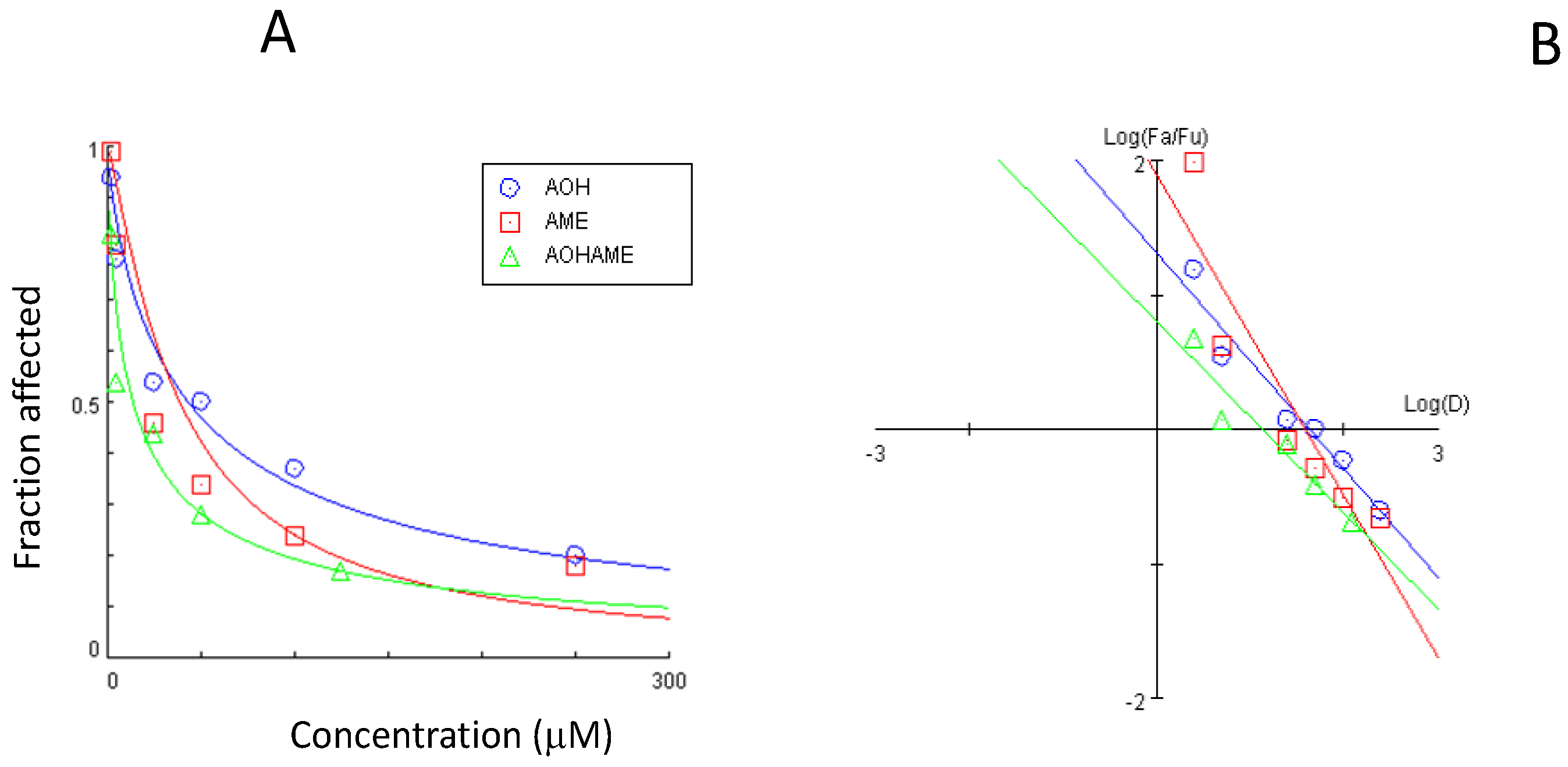
The dose–effect curve and the median-effect plot for the AOH and AME combination are presented in Figure 2.
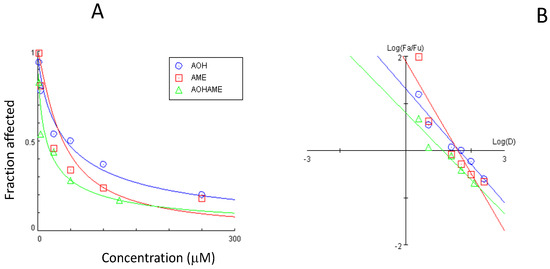
Figure 2.
Dose–effect curve (A) and median-effect plot (B) for the combined AOH and AME in relation to IPEC-1 cell viability.
The interaction between AOH and AME concerning the cytotoxic effect on IPEC-1 cells was assessed via the CI. Figure 3A shows the CI values for cytotoxicity levels between 5 and 95%. The main interaction type ranged from slight synergy for the AOH-AME combinations affecting 25% of cell viability (CI = 0.88) to a synergistic effect for higher levels of cytotoxic IL50 (CI = 0.41), and at IL90, a strong synergistic effect was observed (CI = 0.10).
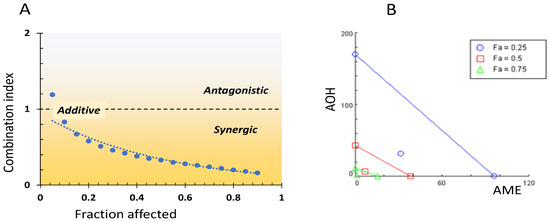
Figure 3.
Interrelation between combination index and fraction affected (A) and isobologram for the combined AOH-AME (B).
The isobologram for the AOH + AME combination corresponding to 25, 50, and 75% inhibition of cell viability is presented in Figure 3B. The additive effect is represented by the diagonal lines between AOH and AME concentrations, while the points situated below the lines illustrate the synergy between the two mycotoxins for different fraction percentages (25, 50, and 75%).
3.3. Effects of AOH, AME, and Combinations Thereof on Reactive Oxygen Species
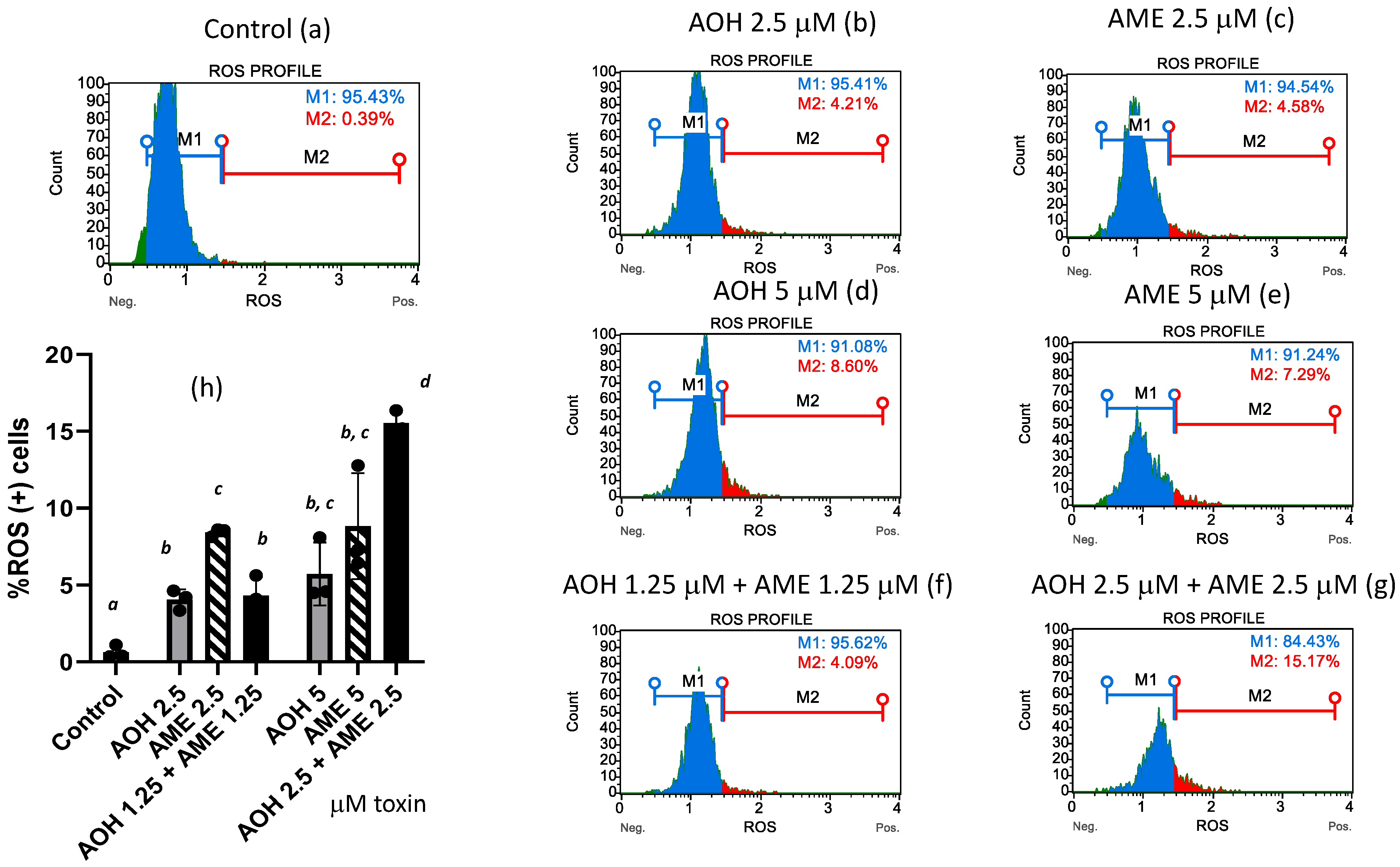
Oxidative stress is triggered by an increase in the intracellular levels of reactive oxygen species (ROS), which damage lipids, proteins, and DNA. In our study, flow-cytometry analysis revealed that both individual toxins and the toxin mixture caused the ROS (+) cells% to increase (Figure 4). While the low-toxin mixture failed to increase the ROS (+) cells to a higher percentage than was achieved by the individual toxins, the highest concentration of the toxin mixture triggered an increase in the percentage of ROS (+) cells that was 2.7 and 1.75 times greater than the effects of AOH and AME administered on an individual basis.
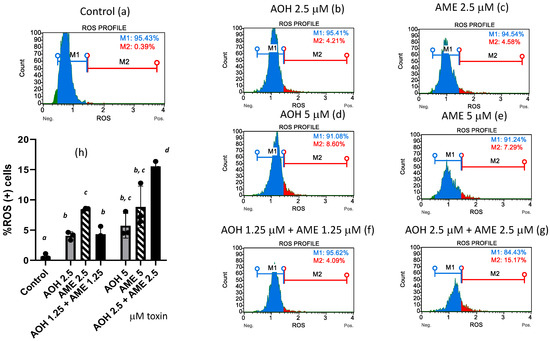
Figure 4.
Effect of AOH, AME, and a combination thereof on the % of ROS (+) cells. Quantification of ROS (+) cells% was performed by flow-cytometry analysis in IPEC-1 cells that underwent 24 h of exposure to AOH, AME, and a combination of the two (ratio 1:1) in concentrations from 0 to 5 μM. M1 = %ROS (−) cells; M2 = %ROS (+) cells. Figures (a–g) represent flow-cytometry histograms for different treatments; figure (h) represents the effect of AOH, AME, and a combination of the two on the % of ROS (+) cells; the results represent the mean ± standard error of the mean (SEM) of three independent experiments, n = 6. Differences among groups were tested using one-way ANOVA followed by Fisher’s PSLD test. a,b,c,d indicate significant differences between different treatments (p < 0.05).
3.4. Effects of AOH, AME, and Their Combinations on Protein, Lipid, and DNA Oxidation
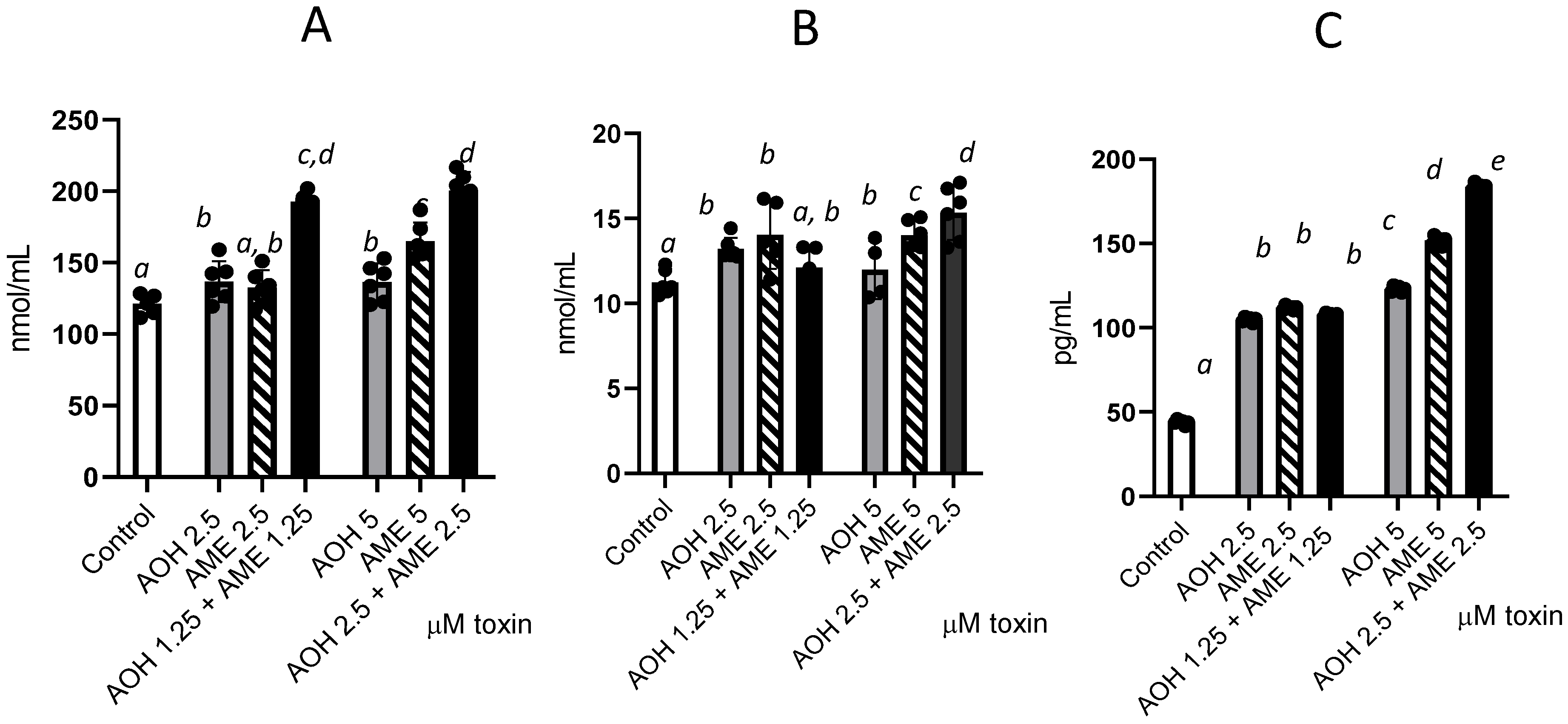
We further investigated the individual and combined toxins’ capacity to trigger oxidative stress by measuring the oxidation of lipids, protein, and DNA (Figure 5) and observed significant increases for the tested concentrations of 2.5 and 5 μM compared to the control. Compared with individual toxins, the combined mixture induced an increase in protein oxidation, as demonstrated by the significant rise in protein carbonyl in cells for both concentrations (2.5 and 5 μM), which was indicative of an additive effect. For lipid oxidation, this effect was observed only for the highest dose of the toxin combination, which led to a 21% increase in TBARSs compared with that observed for AOH alone and an 8.5% increase compared with AME alone. Similarly, for DNA oxidation, the effect of the toxin mixture was observed only for the highest dose, which induced 33% and 17.3% increases in 8 oxo-2′-deoxyguanosine compared with the individual toxins.
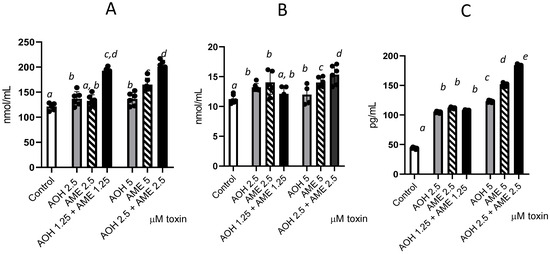
Figure 5.
Effects of AOH, AME, and a combination of the two in concentrations from 0 to 5 μM on different parameters associated with oxidative stress—lipid oxidation (TBARSs), protein oxidation (protein carbonyl), and DNA oxidation (8 oxo dG); the results represent the mean ± standard error of the mean (SEM) of three independent experiments, n = 6. (A) Effect of AOH, AME, and a combination of the two on protein carbonyl concentrations; (B) effect of AOH, AME, and a combination of the two on TBARSs; (C) effect of AOH, AME, and a combination of the two on 8-Oxo-2′-deoxyguanosine. Differences among groups were tested using one-way ANOVA followed by Fisher’s PSLD test. a–e indicate significant differences between treatments (p < 0.05).
3.5. Effects of AOH, AME, and a Combination of the Two on Antioxidant Enzyme Activity in IPEC-1 Cells
As literature data have demonstrated that Alternaria toxins affect oxidative stress, we wanted to investigate the effect of AOH, AME, and a mixture of the two on the activities of catalase, glutathione peroxidase, and superoxide dismutase enzymes in IPEC-1 cells.
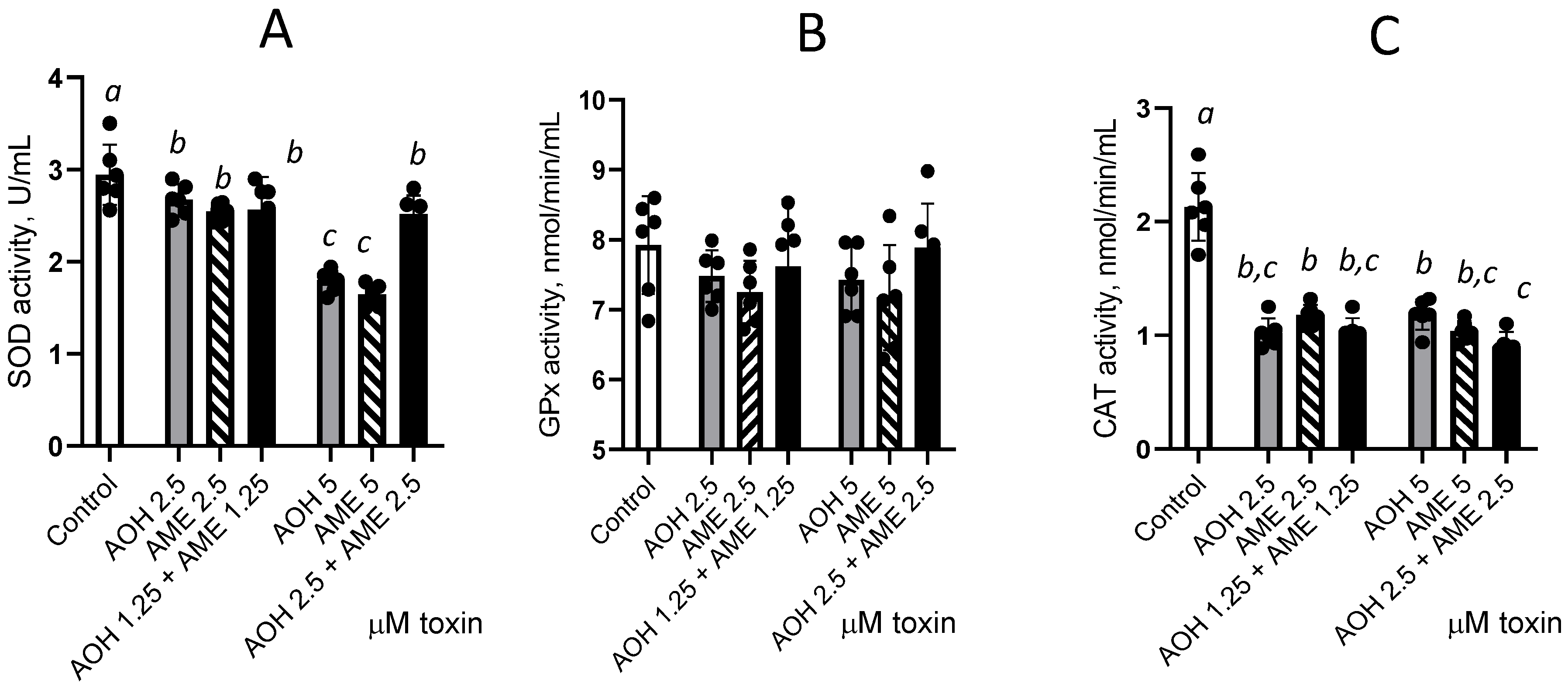
As shown in Figure 6, exposure to individual toxins or their combination significantly decreases SOD and CAT enzyme activity compared with the control, while we observed no such effect on GPx activity. Individual toxins induced a dose-dependent decrease in SOD activity by 1.1 and 1.64 times for 2.5 and 5 μM AOH and by 1.16 and 1.79 times for 2.5 and 5 μM AME, respectively. No additive or synergic effect was induced by the combination of AOH and AME compared with the individual toxin effect for 2.5 μM, while an antagonistic effect was observed for the 5 μM concentration for SOD. A significant decrease in CAT activity was induced by the toxins when they were administered both individually and in combination, but this effect was not dose-dependent, and there were no significant effects due to the interactions between the individual toxins.
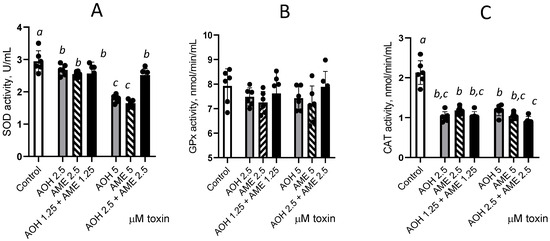
Figure 6.
Effect of AOH, AME, and their combination in concentrations from 0 to 5 μM on the activity of the enzymes involved in the oxidative stress. The activities of superoxide dismutase (SOD), figure (A), glutathione peroxidase (GPx), Figure (B), and catalase (CAT), Figure (C), were analyzed from IPEC-1 cell lysates; the results represent the mean ± standard error of the mean (SEM) of three independent experiments, n = 6. Differences among groups were tested using one-way ANOVA followed by Fisher’s PSLD test. a,b,c indicate significant differences between different treatments (p < 0.05).
4. Discussion
Recent surveys have indicated that cereals are frequently contaminated with Alternaria toxins [3] and with multiple mycotoxins from the same genera. One study conducted between 2014 and 2016 that investigated the contamination of Slovenian cereal samples with Alternaria and other mycotoxins revealed that more than 43% of positive samples were co-contaminated with Alternaria mycotoxins [22]. In particular, studies concerning the presence of AOH and AME identified both toxins in more than 90% of wheat samples in a ratio between 0 and 2.28 [23] or in a ratio between 1 and 3.7 in wheat flour samples [6]. Exposure to co-occurring Alternaria mycotoxins present in the same sample can affect the health of humans and animals alike to a greater extent than exposure to an individual toxin due to the possible cumulative effect of the combined mycotoxins [24]. This effect can be assumed as both AOH and AME have a common production pathway and AME can occur as a result of AOH methylation [25]. Exposure to mycotoxins can occur through the consumption of contaminated products such as cereals [26].
The currently available data concerning Alternaria toxicity are insufficient for an adequate risk assessment and, consequentially, for the establishment of maximum limits in food and feed [27].
Pigs eat a diet rich in cereals, and health issues may occur in animals that consume contaminated feed [28]. Moreover, risk assessment studies in livestock, and especially in swine, are scarce and cannot provide sufficient data to determine the maximum level of toxins that can be present in feed without negative health repercussions [8].
The intestinal epithelium represents the largest barrier between the organism and the external environment. It is responsible for the absorption of nutrients, water, and electrolytes but also for maintaining an effective defense against xenobiotics, including mycotoxins [29]. Several mycotoxins alter the normal intestinal functions of the gastrointestinal tract, such as nutrient absorption and the barrier function [30]. After ingestion, the mycotoxins come into contact with the intestinal epithelial cells, which are the first cells exposed to their toxic effects.
Literature data have proven that Alternaria can trigger toxic effects, exhibiting cytotoxic, genotoxic, and estrogenic properties, as shown by both in vitro and in vivo studies [31]. However, the toxic effect of Alternaria on the intestinal epithelium is not fully understood [32].
For this reason, we assessed the effect of individual and binary combinations of the two most frequently occurring Alternaria mycotoxins, AOH and AME, in swine, using an intestinal cell line—IPEC-1—as these cells are the first to be exposed to the toxins upon their entry to the body. Our data have shown that 24 h of exposure of proliferating IPEC-1 cells to Alternaria toxins induced a dose-dependent decrease in cell viability (Figure 1). Previous studies have indicated that AOH is cytotoxic, inducing cell cycle arrest in the G2/M phase and apoptosis through the mitochondrial pathway [33,34]. AME also induces DNA strand breaks by interacting with DNA topoisomerase and can cause cell cycle arrest [4,19]. In our study, AME triggered a stronger cytotoxic effect in IPEC-1 cells compared with AOH after 24 h, with statistically significant toxin concentrations of 10, 50, and 100 μM. Similar results were obtained in HepG2 cells, where AME was found to be more cytotoxic than AOH and induced significant decreases in cell viability after 24 h of exposure to concentrations of 50 and 100 μM [35]. In Caco2 cells, a similar exposure time (24 h) to AOH and AME at concentrations between 3.125 and 100 μM had no effects on cell viability; a decrease in the number of viable Caco2 was observed only after 48 h of exposure. This effect is more important for AME than for AOH. Other studies have shown that the cytotoxicity of AME in two different cell lines, HepG2 and Caco2, was comparable to that of AOH [4].
In our study, the exposure of IPEC-1 cells to AOH and AME both individually and in combination (in a 1:1 ratio) induced a dose-dependent decrease in cell viability that was more important for the combination of Alternaria toxins than for the individual toxins (Figure 1). Similarly, the exposure of two different cell lines (HepG2 and Caco2) to a binary mixture of AOH and AME in the same ratio as in our study and in concentrations of 4 μg/mL (equivalent to 15.5 μM) induced a more notable increase in cell death compared to the individual AME toxin [4]. Similarly, an assessment of the effect of the type of interactions between AOH (in concentrations from 1.875 to 30 µM) and Fusarium toxin enniatin B (in concentrations from 0.3125 to 5 µM) on the viability of Caco-2 cells has shown that the combination of enniatin B+AOH (1:6 ratio) had greater cytotoxicity than the individual toxins [15].
The interaction between AOH and AME and its cytotoxic effects were analyzed in our study according to the Chou–Talalay method [20] combined with an isobologram, which allowed us to discern whether the interactions were synergistic, additive, or antagonistic. Analyses of the cell viability data have shown that the interaction between the two mycotoxins is mainly synergistic and that the synergic effect is positively correlated with the rise in mycotoxin concentration resulting from the decrease in the CI. In the human gastric epithelial cell line (GES-1), combined AOH and AME had antagonistic effects on cell viability at low concentrations and additive and synergistic effects at high concentrations [16].
In our study, the median-effect dose required to induce 50% inhibition of the cell, calculated according to the Chou–Talalay method, was 43.2 μM for AOH, 38.5 μM for AME, and only 16.89 μM for combined AOH + AME. Bensassi et al. reported that a concentration of 120 μM AME was necessary to induce a 50% decrease in the viability of human colon carcinoma cells (HCT116 cells) according to flow-cytometry analyses [14]. In HT-29 cells exposed to AOH, the EC50 value for AOH was 72.5 μM, while in CaCo2, AOH concentrations up to 104.6 μM failed to induce a 50% decrease in cell viability regardless of the type of assay used [36]. In vitro assessment of the combinatory effects of AOH and another Alternaria mycotoxin—altertoxin II—administered in a 1:10 or 1:1 ratio (ATX II: AOH) mainly revealed additive effects in a hepatic cell line (HepG2) and in two intestinal cell lines (HT29 and HCEC-1CT cells) at lower doses, while antagonism was more commonly observed at the higher doses [37].
Oxidative stress results from disequilibrium between the reactive oxygen species (ROS) produced by and accumulating in the organism and said organism’s ability to mitigate the damaging effect of these products. ROS induction represents an additional mechanism that contributes to the toxicity of Alternaria toxins and plays an important role in their inhibitory effects on cell viability observed in various cellular models [10].
Our results have shown that both AOH and AME can induce an increase in ROS (+) cells% in porcine IPEC-1 cells. Other studies have indicated that Alternaria toxins trigger oxidative stress and are associated with ROS generation, the glutathione protective mechanism, and a change in antioxidant enzymatic defenses; these effects were primarily observed in in vitro studies [10,38]. In our study, AME had more important effects than AOH. Other studies have also found that AOH and AME increased the formation of dichloro-fluorescein in HT29 cells, which was indicative of ROS formation, but AOH induced more potent oxidative effects [39]. Concerning the different toxicological potencies of AOH and AME, these differences could be related to the use of different cell types and species, i.e., swine intestinal epithelial cell line (IPEC-1) vs. human colorectal adenocarcinoma cell line (HT29).
An additive effect was observed when the cells were exposed to the combination of AOH-AME, consisting of an increase in the percentage of ROS (+) cells compared with that observed in response to individual AOH and AME toxins. Such an increase in ROS production can lead to oxidative damage in DNA, lipids, and proteins [40]. The present data confirm our previous findings that AME significantly increases protein and DNA oxidation in IPEC-1 cells [19]. AOH also successfully increased the concentration of protein carbonyl as a marker of protein oxidation, of TBARSs as an indicator of lipid oxidation, and of 8-Oxo-2′-deoxyguanosine as a product of DNA oxidation, compared with the control. As in the case of ROS formation, the oxidative damage induced by the combination of AOH and AME was stronger than the effects of individual toxins, especially for the highest combined dose (2.5 μM AOH + 2.5 μM AME). Other studies indicated that the disruption of redox homeostasis following exposure to Alternaria toxins as a consequence of the increase in ROS levels was related to an alteration in the entire cell death pathway [34]. In addition, data have shown that Alternaria toxins’ capacity to cause oxidative stress and exert genotoxic effects is related to their capacity to act as a topoisomerase poison [41].
In order to neutralize free radicals, cells have developed an antioxidant system consisting of antioxidant enzymes as well as non-enzymatic antioxidants, which collectively reduce the oxidative state [42]. A breakdown of the antioxidant defense was observed in IPEC-1 cells after exposure to individual toxins, related to a decrease in the activity of SOD and CAT antioxidant enzymes, but no additive or synergic effect resulted from the exposure to the toxin mixture.
5. Conclusions
In this study, we aimed to investigate the effects of individual and binary combinations of the two most frequently occurring Alternaria mycotoxins, AOH and AME, in swine; we achieved this using a porcine epithelial intestinal cell line.
Our results concur with previous findings recounting additive or even synergistic effects on cell viability after co-exposure to AOH AME [4,15]. In our study, the exposure of IPEC-1 cells to AOH and AME, both individually and in combination, in a 1:1 ratio, induced a dose-dependent decrease in cell viability that was more significant for the combination of Alternaria toxins than for the individual toxins. This effect was mainly synergistic and was positively correlated with the increase in the mycotoxin concentration. As in previous studies [38,39], we found that exposure to the individual toxins triggered an increase in ROS production and subsequent oxidative damage to DNA, lipids, and proteins, as well as a decrease in the activity of the antioxidant enzymes SOD and CAT. The oxidative damage induced by the combined AOH and AME was more pronounced than the effects of the individual toxins, especially at the highest dose used in this study.
Our study was limited by its use of an in vitro as opposed to an in vivo model. However, the IPEC-1 cell line was not immortalized and was established from normal intestinal epithelium cells isolated from the jejunum and ileum of a neonatal pig [43] so the results obtained in vitro can be considered more reliable than those found using immortalized cell models (e.g., CaCo2). Further in vivo studies in swine are needed in order to confirm our results. The signaling pathways responsible for the toxicity of AOH, AME, and their combinations should also undergo further investigation; this will provide important risk assessment data for contaminated swine feed.
As many existing studies provide toxicological data concerning exposure to individual toxins, and the co-contamination of food and feed with mycotoxins is a frequent occurrence, our results provide important risk assessment data for swine feed co-contaminated with Alternaria toxins.
Author Contributions
Conceptualization, D.E.M.; methodology, G.C.P.; formal analysis, C.V.B.; investigation, C.V.B., A.M.P. and I.A.G.; writing—original draft preparation, D.E.M.; writing—review and editing, I.T.; project administration, D.E.M.; funding acquisition, D.E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Romanian Ministry of Research, Innovation and Digitalization, grant number PCE 42/2022.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data can be made available on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Miranda-Apodaca, J.; Artetxe, U.; Aguado, I.; Martin-Souto, L.; Ramirez-Garcia, A.; Lacuesta, M.; Becerril, J.M.; Estonba, A.; Ortiz-Barredo, A.; Hernández, A.; et al. Stress response to climate change and postharvest handling in two differently pigmented lettuce genotypes: Impact on alternaria alternata invasion and mycotoxin production. Plants 2023, 12, 1304. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhu, R. Alternaria allergy and immunotherapy. Int. Arch. Allergy Immunol. 2024, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nagda, A.; Meena, M. Alternaria mycotoxins in food and feed: Occurrence, biosynthesis, toxicity, analytical methods, control and detoxification strategies. Food Control 2024, 158, 110211. [Google Scholar] [CrossRef]
- den Hollander, D.; Holvoet, C.; Demeyere, K.; De Zutter, N.; Audenaert, K.; Meyer, E.; Croubels, S. Cytotoxic effects of alternariol, alternariol monomethyl-ether, and tenuazonic acid and their relevant combined mixtures on human enterocytes and hepatocytes. Front. Microbiol. 2022, 13, 849243. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef]
- Zhao, K.; Shao, B.; Yang, D.; Li, F.; Zhu, J. Natural occurrence of alternaria toxins in wheat-based products and their dietary exposure in china. PLoS ONE 2015, 10, e0132019. [Google Scholar] [CrossRef] [PubMed]
- Puvača, N.; Avantaggiato, G.; Merkuri, J.; Vuković, G.; Bursić, V.; Cara, M. Occurrence and determination of alternaria mycotoxins alternariol, alternariol monomethyl ether, and tentoxin in wheat grains by quechers method. Toxins 2022, 14, 791. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on the risks for animal and public health related to the presence of alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Crudo, F.; Hong, C.; Varga, E.; Del Favero, G.; Marko, D. Genotoxic and mutagenic effects of the alternaria mycotoxin alternariol in combination with the process contaminant acrylamide. Toxins 2023, 15, 670. [Google Scholar] [CrossRef]
- Tiessen, C.; Ellmer, D.; Mikula, H.; Pahlke, G.; Warth, B.; Gehrke, H.; Zimmermann, K.; Heiss, E.; Fröhlich, J.; Marko, D. Impact of phase i metabolism on uptake, oxidative stress and genotoxicity of the emerging mycotoxin alternariol and its monomethyl ether in esophageal cells. Arch. Toxicol. 2017, 91, 1213–1226. [Google Scholar] [CrossRef]
- Kowalska, K.; Kozieł, M.J.; Urbanek, K.A.; Habrowska-Górczyńska, D.E.; Domińska, K.; Piastowska-Ciesielska, A.W. Estrogen receptor β participates in alternariol-induced oxidative stress in normal prostate epithelial cells. Toxins 2021, 13, 766. [Google Scholar] [CrossRef] [PubMed]
- Puvača, N.; Tanasković, S.; Bursić, V.; Petrović, A.; Merkuri, J.; Shtylla Kika, T.; Marinković, D.; Vuković, G.; Cara, M. Optical characterization of Alternaria spp. Contaminated wheat grain and its influence in early broilers nutrition on oxidative stress. Sustainability 2021, 13, 4005. [Google Scholar] [CrossRef]
- Speijers, G.J.; Speijers, M.H. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; Sharaf el dein, O.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Combined effects of alternariols mixture on human colon carcinoma cells. Toxicol. Mech. Methods 2015, 25, 56–62. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Role of quercetin on caco-2 cells against cytotoxic effects of alternariol and alternariol monomethyl ether. Food Chem. Toxicol. 2016, 89, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Jia, B.; Wu, A. Cytotoxicities of co-occurring alternariol, alternariol monomethyl ether and tenuazonic acid on human gastric epithelial cells. Food Chem. Toxicol. 2023, 171, 113524. [Google Scholar] [CrossRef] [PubMed]
- Holanda, D.M.; Kim, S.W. Mycotoxin occurrence, toxicity, and detoxifying agents in pig production with an emphasis on deoxynivalenol. Toxins 2021, 13, 171. [Google Scholar] [CrossRef] [PubMed]
- Novak, B.; Rainer, V.; Sulyok, M.; Haltrich, D.; Schatzmayr, G.; Mayer, E. Twenty-eight fungal secondary metabolites detected in pig feed samples: Their occurrence, relevance and cytotoxic effects in vitro. Toxins 2019, 11, 537. [Google Scholar] [CrossRef]
- Marin, D.E.; Bulgaru, V.C.; Pertea, A.; Grosu, I.A.; Pistol, G.C.; Taranu, I. Alternariol monomethyl-ether induces toxicity via cell death and oxidative stress in swine intestinal epithelial cells. Toxins 2024, 16, 223. [Google Scholar] [CrossRef]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of hepg2 cells to zearalenone and its metabolites. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Babič, J.; Tavčar-Kalcher, G.; Celar, F.A.; Kos, K.; Knific, T.; Jakovac-Strajn, B. Occurrence of alternaria and other toxins in cereal grains intended for animal feeding collected in slovenia: A three-year study. Toxins 2021, 13, 304. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yoshizawa, T. Alternaria mycotoxins in weathered wheat from china. J. Agric. Food Chem. 2000, 48, 2920–2924. [Google Scholar] [CrossRef]
- Ruiz, M.J.; Macáková, P.; Juan-García, A.; Font, G. Cytotoxic effects of mycotoxin combinations in mammalian kidney cells. Food Chem. Toxicol. 2011, 49, 2718–2724. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, M.; Söderhäll, K. Alternatiol-o-methyltransferase fromalternaria alternata: Partial purification and relation to polyketide synthesis. Exp. Mycol. 1992, 16, 44–51. [Google Scholar] [CrossRef]
- Saleh, I.; Zeidan, R.; Abu-Dieyeh, M. The characteristics, occurrence, and toxicological effects of alternariol: A mycotoxin. Arch. Toxicol. 2024, 98, 1659–1683. [Google Scholar] [CrossRef]
- Crudo, F.; Varga, E.; Aichinger, G.; Galaverna, G.; Marko, D.; Dall’Asta, C.; Dellafiora, L. Co-occurrence and combinatory effects of alternaria mycotoxins and other xenobiotics of food origin: Current scenario and future perspectives. Toxins 2019, 11, 640. [Google Scholar] [CrossRef]
- Marin, D.E.; Bulgaru, C.V.; Anghel, C.A.; Pistol, G.C.; Dore, M.I.; Palade, M.L.; Taranu, I. Grape seed waste counteracts aflatoxin b1 toxicity in piglet mesenteric lymph nodes. Toxins 2020, 12, 800. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20; quiz 21–22. [Google Scholar] [CrossRef]
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- Schmutz, C.; Cenk, E.; Marko, D. The alternaria mycotoxin alternariol triggers the immune response of il-1β-stimulated, differentiated caco-2 cells. Mol. Nutr. Food Res. 2019, 63, e1900341. [Google Scholar] [CrossRef]
- Islam, M.T.; Martorell, M.; González-Contreras, C.; Villagran, M.; Mardones, L.; Tynybekov, B.; Docea, A.O.; Abdull Razis, A.F.; Modu, B.; Calina, D.; et al. An updated overview of anticancer effects of alternariol and its derivatives: Underlying molecular mechanisms. Front. Pharmacol. 2023, 14, 1099380. [Google Scholar] [CrossRef]
- Solhaug, A.; Vines, L.L.; Ivanova, L.; Spilsberg, B.; Holme, J.A.; Pestka, J.; Collins, A.; Eriksen, G.S. Mechanisms involved in alternariol-induced cell cycle arrest. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2012, 738–739, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hessel-Pras, S.; Kieshauer, J.; Roenn, G.; Luckert, C.; Braeuning, A.; Lampen, A. In vitro characterization of hepatic toxicity of alternaria toxins. Mycotoxin Res. 2019, 35, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Oxidative stress of alternariol in caco-2 cells. Toxicol. Lett. 2014, 229, 458–464. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Sack, M.; Jarolim, K.; Aichinger, G.; Somoza, M.M.; Marko, D. In vitro combinatory effects of the alternaria mycotoxins alternariol and altertoxin ii and potentially involved mirnas. Toxicol. Lett. 2017, 267, 45–52. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Oxidative DNA damage and disturbance of antioxidant capacity by alternariol in caco-2 cells. Toxicol. Lett. 2015, 235, 61–66. [Google Scholar] [CrossRef]
- Tiessen, C.; Fehr, M.; Schwarz, C.; Baechler, S.; Domnanich, K.; Böttler, U.; Pahlke, G.; Marko, D. Modulation of the cellular redox status by the alternaria toxins alternariol and alternariol monomethyl ether. Toxicol. Lett. 2013, 216, 23–30. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ros) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemöller, M.; Podlech, J.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the iiα isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Nossol, C.; Barta-Böszörményi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.K.; Rothkötter, H.J. Comparing Two Intestinal Porcine Epithelial Cell Lines (IPECs): Morphological Differentiation, Function and Metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).