Abstract
Improving seed germination and seedling development can potentially increase crop yield and improve quality in direct-seeded rice. This study aimed to detect loci or genes associated with rice seed germination. We reported the phenotypic analysis of seed germination in 103 rice accessions across two years, and a genome-wide association study (GWAS) was conducted to identify loci underlying the genetic regulation of seed germination. A total of seven genetic loci were found to be associated with seed germination, including five loci that overlapped with the previously reported loci/genes, and two novel loci. Of these, two loci (qGP2 and qGP4.1) were stable across different environments. GP4 (Germination percentage 4), encoding a 9-cis-epoxycarotenoid dioxygenase, was identified as the candidate gene of the major locus qGP4.1. A sequence analysis of GP4 revealed that four functional polymorphic sites in the coding region were significantly associated with germination percentage. The disruption of GP4 by gene editing resulted in faster seed germination and seedling establishment. Taken together, we have identified GP4 as a novel gene involved in rice seed germination, and we provide a potential target gene for improving rice seed vigor via gene editing or molecular breeding.
1. Introduction
Seed germination is a critical developmental phase transition that initiates the life cycle of flowering plants [1]. It is generally believed that seeds with low levels of vigor germinate slowly and have weak seedlings, while seeds with high vigor germinate more quickly and have the potential to build healthy seedlings successfully in the field [2]. Rice (Oryza sativa L.) is one of the major staple food crops globally. The direct seeding of rice has recently been widely applied in many Asian countries due to its technical simplicity and efficiency [3]. However, slow germination and poor seedling establishment can lead to yield loss in the direct-seeding cultivation technology for rice [4]. Hence, understanding the natural variations in seed germination can help to genetically improve rice growth and grain yield using molecular breeding strategies.
Seed germination is a complex trait and genetically controlled by numerous quantitative trait loci (QTLs) or genes in rice. To explore the genetic variation in rice seed germination, many QTLs have been dissected based on segregating populations derived from biparental crosses [5,6,7,8,9,10]. For example, the QTL qLTG3-1 regulated germination capacity under low-temperature stress by weakening the embryo surrounding tissues [6]. Seed dormancy 4 (Sdr4) was identified as a major QTL for seed dormancy and acted downstream of the seed maturation-related gene OsVP1 to inhibit rice pre-harvest sprouting [7]. The Seed Dormancy1-2 (qSD1-2) locus was found to be involved in regulating endosperm-imposed dormancy and plant height in rice [8]. Although the biparental linkage mapping approach can be effectively used for identification of chromosome regions for seed germination, very limited recombination events in the biparental mapping may lead to detecting QTLs in large intervals rather than candidate genes [11,12]. Over the past decade, GWAS has emerged as a popular approach to test millions of single-nucleotide polymorphisms (SNPs) associated with trait variation in crops due to its high-resolution mapping [13]. More recently, the GWAS approach has also been employed in an attempt to dissect the genetic architecture of seed germination in rice, and dozens of key candidate genes that contribute to seed germination have been identified and functionally verified, such as OsOMT, OsGH3-2, OsCLSY1, OsPK5, GF14h, qGR1-1, OsGA2ox5, Rc, OsTTL, OsSAPK1, and OsTMF [14,15,16,17,18,19,20,21,22,23,24]. However, more and new genes related to seed germination need to be discovered from rice germplasm.
Seed germination is a complex physiological process controlled by several hormones [25]. Abscisic acid (ABA) is one of the key hormones that can promote seed dormancy and suppress seed germination and early seedling growth [26]. The regulation of cellular ABA level is determined by the dynamic balance of biosynthetic and catabolic pathways, allowing plants to cope with changes in complex physiological and environmental conditions [27,28]. In higher plants, de novo ABA biosynthesis starts from the cleavage of carotenoid precursors [29]. To date, most ABA biosynthesis genes have been well characterized, including zeaxanthin epoxidase, 9-cis-epoxycarotenoid dioxygenase (NCED), and ABA aldehyde oxidase [30]. Among them, NCED catalyzes the oxidative cleavage of 9-cis-epoxycarotenoid to release xanthoxin, which is considered to be the rate-limiting step in ABA biosynthesis [31]. The first NCED gene was identified from the ABA-deficient maize mutant vp14 [32,33]. Since then, the function of NCED genes in diverse plant species have been widely identified and studied [34,35,36]. However, allelic variations in NCED genes associated with seed germination have rarely been reported in plants.
In this study, we conducted a GWAS using 700K SNPs in a genetic panel of 103 rice accessions to identify loci linked to seed germination. A total of seven loci associated with early seed germination were identified, of which qGP2 on chromosome 2 and qGP4.1 on chromosome 4 were stably expressed over the two years. Through a combination of gene sequencing and knockout verification, GP4, encoding 9-cis-epoxycarotenoid dioxygenase, was considered as the most likely candidate gene for the qGP4.1 locus associated with seed germination. These findings will help to facilitate molecular design in rice breeding for the genetic improvement of seed vigor.
2. Materials and Methods
2.1. Plant Materials
The INDICA varietal group used in this study was obtained from Rice Diversity Pane 1 (RDP1), including 64 indica and 39 aus accessions (Supplementary Table S1) [37]. The knockout mutant of the GP4 gene was generated in the wild-type (WT) background (cv Nipponbare) using the clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9 technology. A natural population of 103 rice accessions was planted in the spring of 2020 and 2021, respectively. The WT and gp4 mutant were planted in the summer of 2022. All rice plants were cultivated in an experimental field of the Zhongkai University of Agriculture and Engineering (Guangdong Province, China). Each INDICA accession, WT and GP4 mutant was planted in eight rows with eight plants per row (33 × 17 cm spacing). Field management followed normal local agricultural practices. The seeds were harvested at 35 days after flowering stored dry at room temperature (25 °C) for two months, and then stored in a refrigeration house (4 °C) before performing germination tests. Accessions with low seed-setting rate were excluded, leaving 101 accessions in 2020 and 91 accessions in 2021 kept for the phenotype analysis (Supplementary Table S1).
2.2. Evaluation of Seed Germination
Fifty healthy seeds of each replicate were placed on double sheets of filter paper moistened with 10 mL distilled water in 9 cm Petri dishes, and then incubated in a 25 °C greenhouse with a 12 h day/12 h night photoperiod. Seed germination was defined as an observed radicle emergence of 2 mm, and the seedling establishment was subject to the condition that the root length was equal to the seed length and the shoot length was half the seed length. The number of germinated seeds for 103 rice accessions was recorded two days after imbibition. The number of germinated seeds and seedlings for WT and mutant were recorded daily for one week. The traits of germination percentage, seedlings percentage and germination index were calculated according to the method that was described previously [38]. Three replicates of each accession or mutant line were included in this study.
2.3. Genome-Wide Association Study
A high-SNP-density dataset (700K SNPs) of 103 rice accessions was downloaded from the Rice Diversity database (http://www.ricediversity.org/data/index.cfm, accessed on 11 July 2020), which has been published previously [39]. SNPs with a low minor allele frequency (<5%) and high missing ratio (>25%) were excluded from the SNP dataset, leaving 343,464 SNPs. A GWAS was performed with a mixed linear model implemented in the R package GAPIT 3.0 [40]. We used a significance threshold of p ≤ 10−4 (i.e., −log10 p ≥ 4) to identify significant marker-trait associations, according to a previous GWAS on this population [37,39]. The GWAS results were presented as Manhattan plots using the qqman package in R. Any two lead SNPs within a 200 kb interval were grouped into one association locus [17].
2.4. Generation and Identification of gp4 Mutant
The plasmid vector of the CRISPR/Cas9 system targeting GP4 locus was constructed as described previously [41]. CRISPR targeting sites were designed using CRISPR-PLANT (http://omap.org/crispr/CRISPRsearch.html, accessed on 22 September 2021). The two targeted site spacers in the exon of GP4 were 5′-CCGTCGTCATCGTCTGCTCCTGT-3′ and 5′-CCGGCACGGTGAACCGGTCGCTG-3′. Transgenic rice plants were generated via the Agrobacterium-mediated transformation method, as previously reported [42]. The DNA fragments covering the two target sites were PCR-amplified from T2 plants with specific primers (5′ primer: CGCCGTCTCTAGAATGAGGT, 3′ primer: CCACCTCCATCTTCCCGG). The PCR products were then sequenced and analyzed to identify mutations.
2.5. Data Analysis
Descriptive statistics, including mean, standard deviation (SD), coefficient of variation (CV%), range, and heritability were calculated. Data were analyzed by analysis of variance (ANOVA) using the Excel 2017 software program. The significant differences between WT and gp4 mutant, between C type and A type of lead SNP, and between Hap1 and Hap2 of GP4 were identified using Student’s t-test. Broad-sense heritability (h2) was calculated using the equation h2 = σ2g/(σ2e/rt + σ2ge/t + σ2g) × 100%, where σ2g is the genotypic variance, σ2ge is the genotype-by-environment interaction variance, σ2e is the experimental error variance, and t and r are the number of environments (years) and replicates, respectively. Correlation coefficient was calculated using Pearson correlation analysis. The phenotypic data were transformed into normal distributions using the square root transformation. The normality of trait distributions was evaluated by the Kolmogorov–Smirnov test (cut-off at p = 0.01). Pearson correlation and normality testing were performed using SPSS v21.0 (IBM SPSS Statistics, Chicago, IL, USA).
3. Results
3.1. Characterization of Seed Germination in INDICA Varietal Group
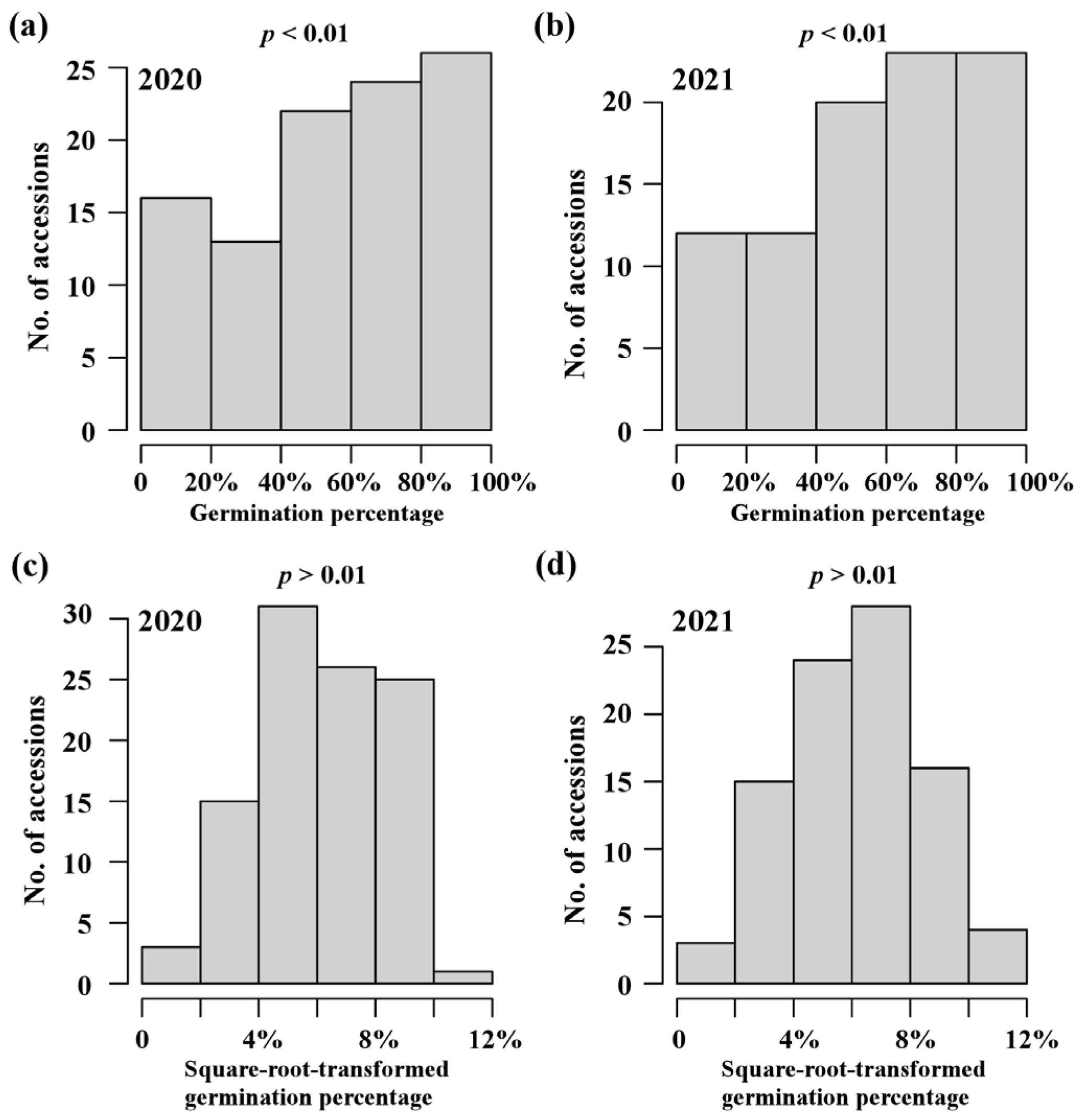
To investigate the variation in rice seed germination, a population of 103 rice accessions belonged to INDICA varietal group in RDP1 was used to estimate germination trait (germination percentage at two days after imbibition) in 2020 and 2021 (Supplementary Table S1). Wide phenotypic variations were observed in both years (Table 1). As shown in Figure 1a,b, the frequency distributions of phenotypic data displayed a similar pattern in both years, with more in the high germination percentage side. In particular, the four accessions, NSFTV30 (origin Vietnam), NSFTV43 (origin China), NSFTV208 (origin India), and NSFTV349 (origin China), had a high germination percentage of more than 90% across two years, indicating that they could be used as a potential high-vigor donor in breeding programs. A highly significant correlation (Pearson’s correlation coefficient > 0.8; p value < 0.01) was observed between the years. Additionally, the phenotypic data for this population were quite repeatable across years, with broad-sense heritability estimated at 94.19% (Table 1). These findings indicated that wide variations in germination percentage in the INDICA varietal group were mostly determined by the genetic factors and were thus suitable for further genetic mapping. Due to non-normality distribution for germination percentages (Figure 1a,b; p value < 0.01), the raw data were square-root-transformed to achieve approximate normal distributions (Figure 1c,d; p value > 0.01) prior to GWAS.

Table 1.
Descriptive statistics of germination percentage in the mapping population.
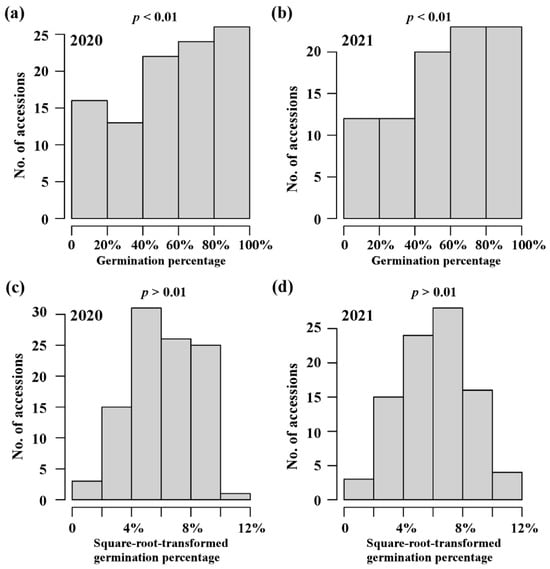
Figure 1.
Phenotypic variation in seed germination in INDICA varietal group. The distribution of germination percentage at two days after imbibition in 2020 (a) and 2021 (b). The distribution of square-root-transformed germination percentage in 2020 (c) and 2021 (d). Normality of the distributions was assessed by the Kolmogorov–Smirnov test (cut-off at p = 0.01).
3.2. GWAS of Seed Germination in INDICA Varietal Group
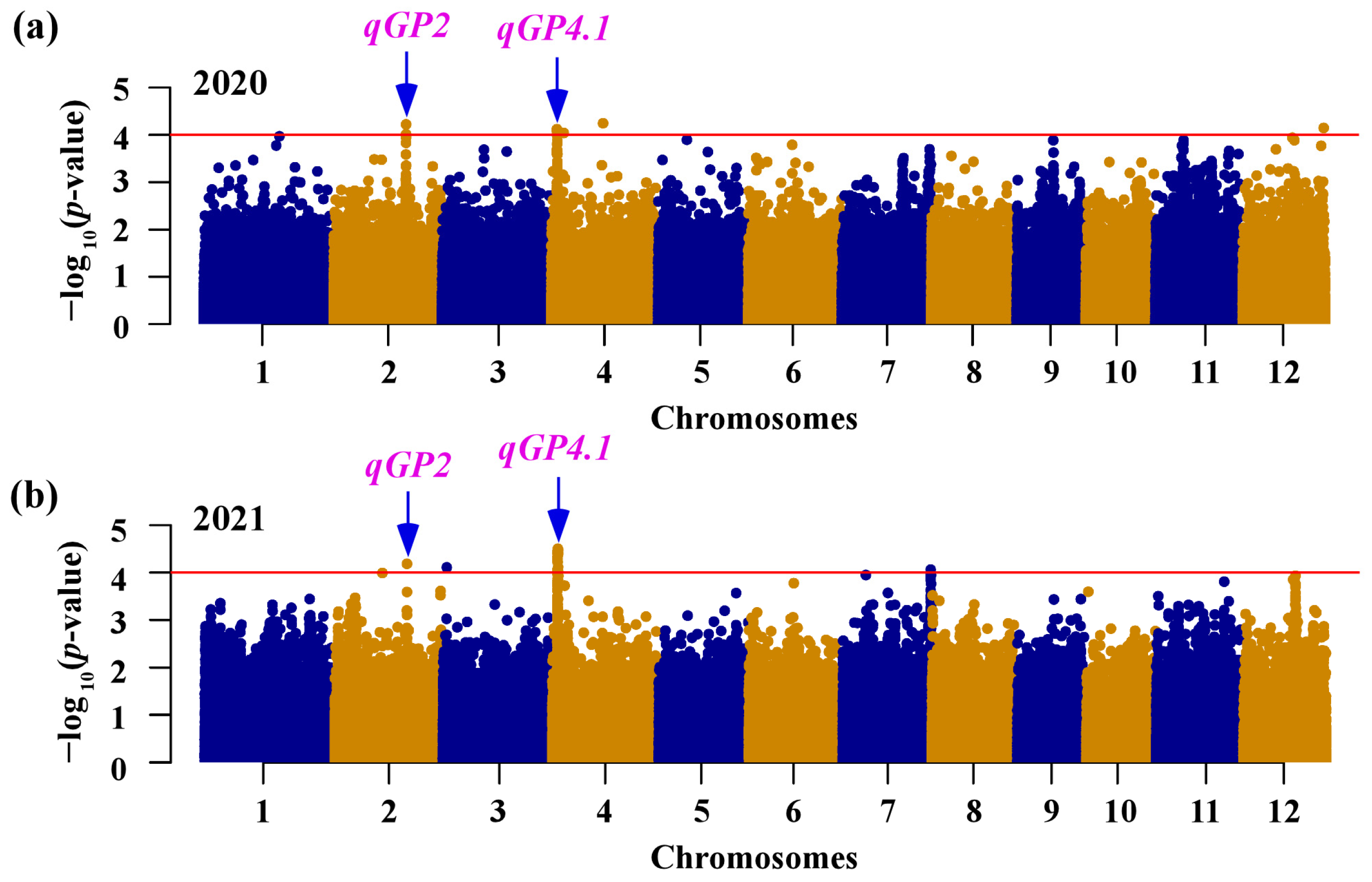
GWAS was performed in INDICA varietal group to reveal the genetic basis of the natural variation in seed germination using MLM method for 343,464 SNPs. A total of seven loci exhibited significant associations with the germination percentage, which was distributed on chromosome 2, 3, 4, 7, and 12 (Table 2). Two of these loci (qGP2 and qGP4.1) were repeatedly detected in two consecutive years (Figure 2; Table 2). We identified five loci (qGP2, qGP3, qGP4.1, qGP7, and qGP12) co-located with previously identified QTLs or genes related to seed germination [16,43,44,45,46,47], and two loci (qGP4.2 and qGP4.3) were newly discovered in the present study (Table 2). Interestingly, qGP4.1 was the locus with the multiple s significant SNPs in both years, suggesting that it is a major genetic locus for germination percentage in the INDICA varietal group. Therefore, we subsequently selected this locus for further investigation.
Table 2.
Loci associated with germination percentage.
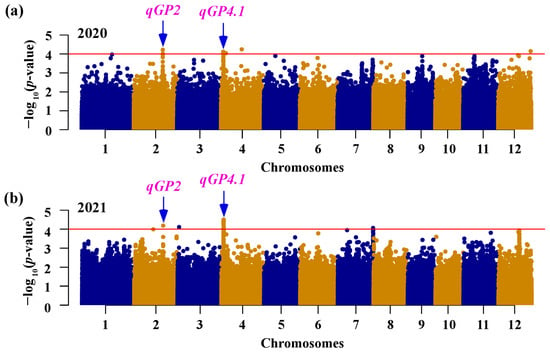
Figure 2.
GWAS of seed germination. Manhattan plots for germination percentage at two days after imbibition in 2020 (a) and 2021 (b). The horizontal lines represent the genome-wide significance threshold of −log10 p = 4. The arrows indicate the loci that are stable across two years.
3.3. Identification of GP4 as a Candidate Gene for Seed Germination
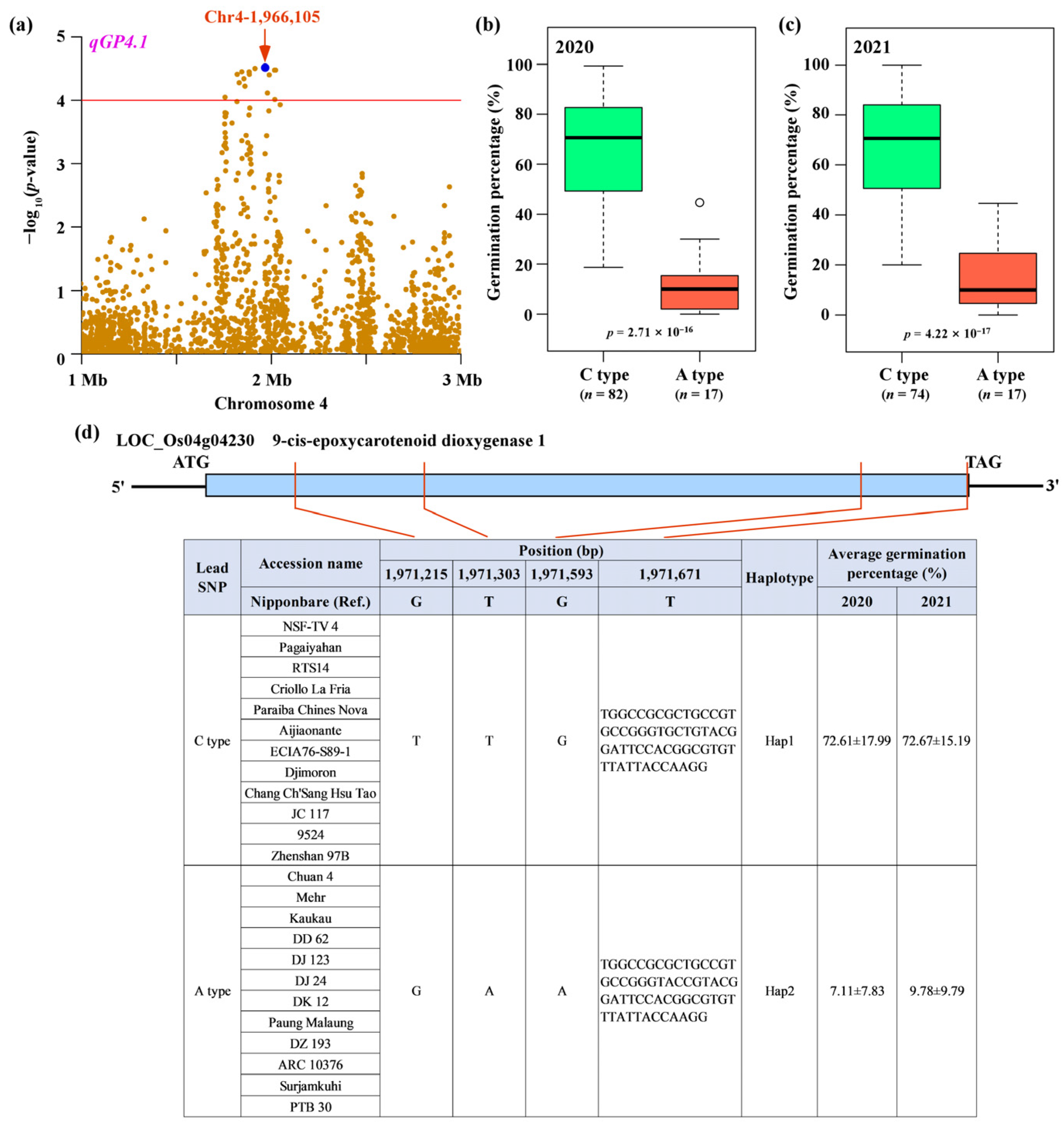
The most significant SNP (lead SNP) for qGP4.1 was located at 1,966,105 bp on chromosome 4 (Figure 3a). To estimate the effect of the lead SNP on the phenotypes, we performed a statistical analysis of germination percentage between two allelic groups stratified by the lead SNP (Supplementary Table S2), and the results showed that the rice accessions carrying C-type had significantly higher germination percentage than those carrying A-type across two years (Figure 3b,c). Within the qGP4.1 locus, we found that the gene LOC_Os04g04230 encoding a putative 9-cis-epoxycarotenoid dioxygenase 1, was immediately adjacent to the lead SNP within a 5 kb distance (Supplementary Figure S1). In previous studies, 9-cis-epoxycarotenoid dioxygenases have been found to be involved in seed dormancy and germination in plants [48,49]. Therefore, we deduced that LOC_Os04g04230 was a strong candidate gene in this associated locus. To investigate the contribution of the natural variation in LOC_Os04g04230 to the variation in germination percentage in rice, we sequenced the coding region of LOC_Os04g04230 from 12 accessions with C-type and 12 accessions with A-type, and compared it to the reference sequence. A total of seven polymorphic sites were identified within the coding region of LOC_Os04g04230 (Supplementary Figure S2). Three SNPs (1,971,215, 1,971,303, and 1,971,593 bp) were nonsynonymous substitution SNPs and resulted in amino acid changes, one INDEL (1,971,671 bp) caused frameshift mutation, and the remaining three SNPs (1,971,565, 1,971,595, and 1,971,664 bp) were synonymous substitution SNPs and caused no amino acid change (Supplementary Figure S2). Therefore, we predicted that nonsynonymous SNP and INDEL variations (1,971,215 bp, 1,971,303 bp, 1,971,593, and 1,971,671 bp) in LOC_Os04g04230 may have a genetic effect on seed germination by altering protein function. Of these key variants, LOC_Os04g04230 could be classified into two haplotypes (Figure 3d). Accessions carrying Hap1 showed significantly higher germination percentage than those carrying Hap2 across two years (p = 2.71 × 10−16 in 2020, p = 4.22 × 10−17 in 2021; Figure 3d). Therefore, LOC_Os04g04230 was the most likely candidate for the qGP4.1 locus, and designated tentatively as Germination percentage 4 (GP4).
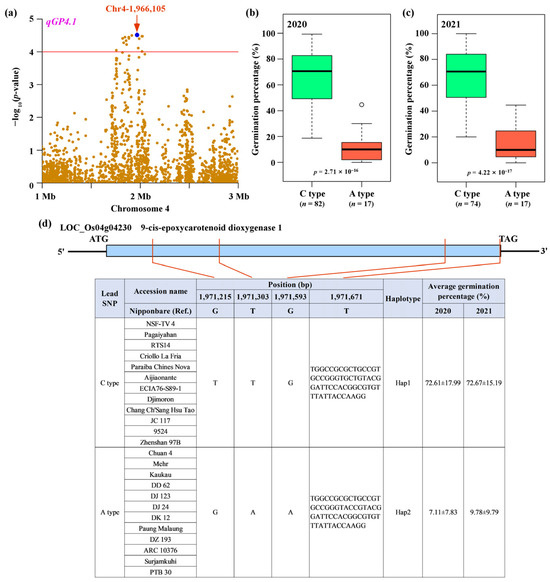
Figure 3.
GP4 is the causal gene for qGP4.1. (a) Local Manhattan plot for qGP4.1 on chromosome 4. Significance test of germination percentage for the allelic groups of rice accessions divided by the lead SNP (1,966,105 bp, C/A) for 2020 (b) and 2021 (c). (d) Haplotype analysis of GP4 showing an association of the four functional polymorphic sites with germination percentage. The p-values were obtained from Student’s t-test.
3.4. Functional Validation of GP4 through CRISPR-Cas9 System
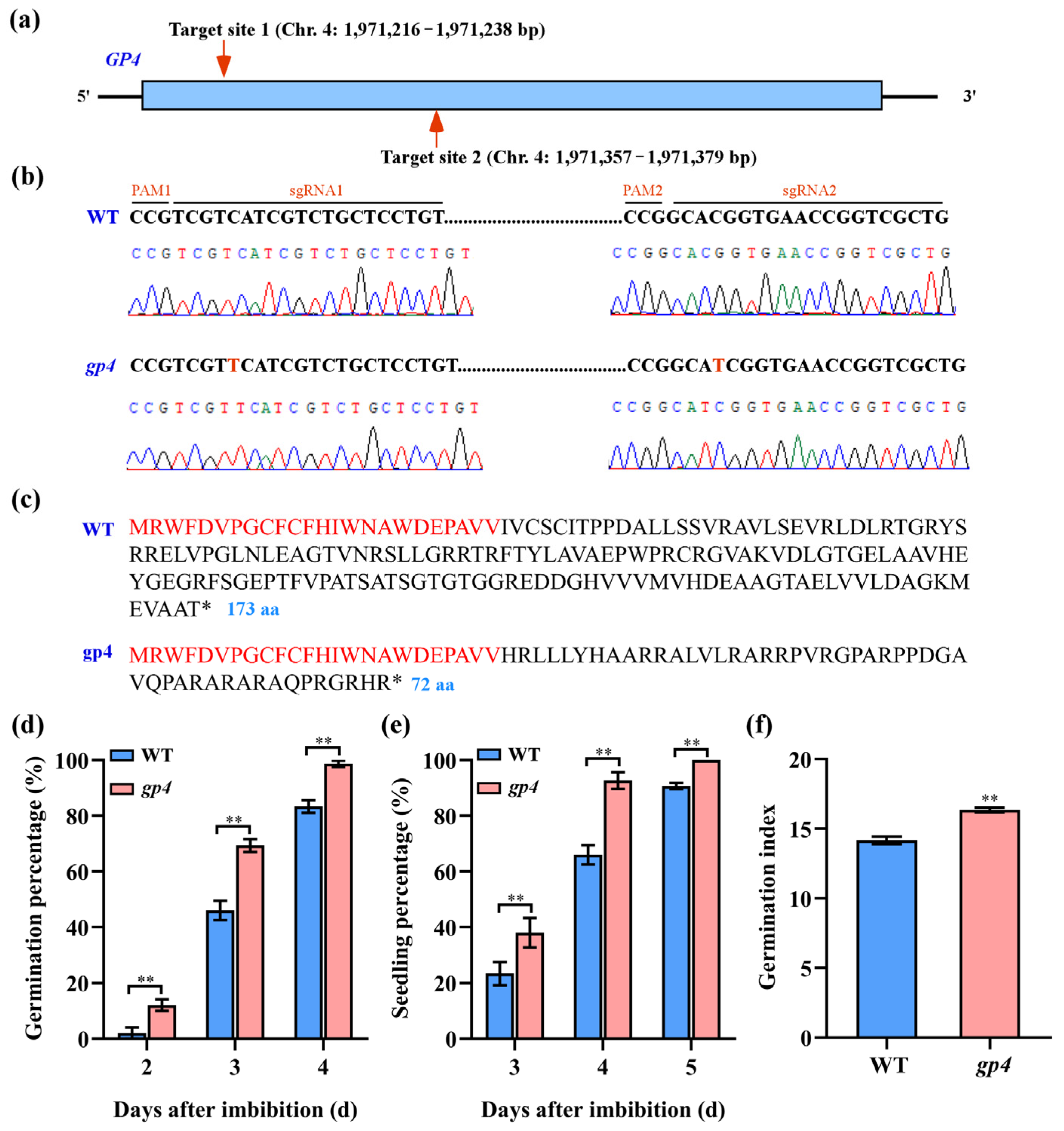
To investigate the potential involvement of the GP4 gene in regulating seed germination, we employed sgRNA-mediated CRISPR/Cas9 technology to create gp4 mutant in the Nipponbare background using two sgRNAs (Figure 4a). Through this approach, we identified one homozygous gp4 mutant, which contained a 1 bp insertion at the two target sites, respectively (Figure 4b). The insertions were predicted to induce a shift in the reading frame and a premature termination of protein translation (Figure 4c). The phenotype analysis showed that the germination percentages at day 2 to day 4 and the seedling percentages at day 3 to day 5 of gp4 mutant were significantly higher than those of WT (Figure 4d,e). We also noted that the gp4 mutant had a significantly higher germination index than WT after 7 days of germination (Figure 4f). These data suggested that the GP4 gene functioned as a negative regulator to participate in rice seed germination.
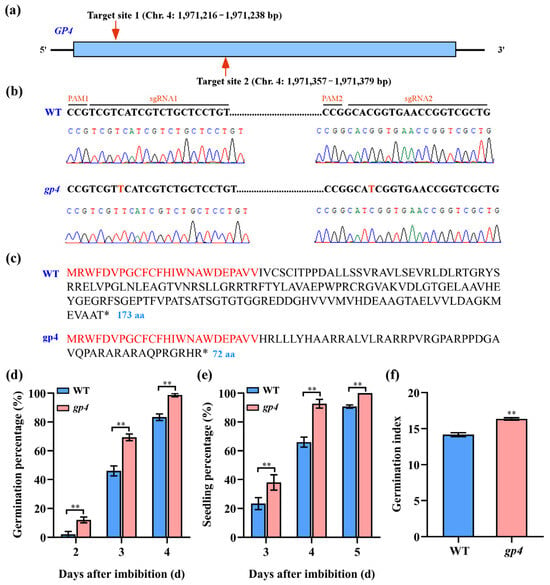
Figure 4.
Functional validation of GP4 via gene editing. (a) Generation of gp4 mutant using CRISPR/Cas9 system. (b) The target sequence region of GP4 is PCR amplified and sequenced to identify mutations. (c) Comparison of deduced amino acid sequences of homozygous mutant gp4 and the WT; identical amino acid sequence is highlighted in red. (d) Comparison of germination percentage between the WT and gp4 mutant on the 2nd, 3rd, and 4th days. (e) Comparison of seedling percentage between the WT and gp4 mutant on the 3rd, 4th, and 5th days. (f) Comparison of germination index between the WT and gp4 mutant. Identical amino acid sequence was highlighted in red. The asterisk indicated a stop codon. Statistical differences between the WT and gp4 mutant were analyzed using Student’s t-test. ** Significant difference between the WT and gp4 mutant (p < 0.01).
4. Discussion
Seed germination is an important criterion of seed vigor, and greater seed germination and vigor is closely correlated with better a seedling performance and yield potential in the direct seeding of rice [50]. Therefore, breeding elite rice varieties with a high seed vigor suitable for direct seeding is a sustainable method to protect rice yields. In this study, we conducted genetic mapping on an INDICA varietal group and tried to identify the functional loci that could potentially be selected for use in rice breeding. Previous findings reported that the INDICA group had a significantly higher level of seed vigor than JAPONICA during germination [14,19,21]. However, the difference in seed germination within the INDICA varietal group remains unclear. Our results showed that the INDICA varietal group had high phenotypic variations in germination percentage in both years. This finding suggests that this population had a diverse genetic background and that it would be beneficial to explore the genetic basis of variations in seed germination.
In the past decade, GWAS has become increasingly popular to uncover the genetic underpinnings of complex traits in crops [51]. In this study, a total of seven loci were found to be associated with germination percentage through GWAS in two years. Three loci were consistent across two years (qGP2 and qGP4.1), while five loci (qGP3, qGP4.2, qGP4.3, qGP7, and qGP12) were identified in each single year, suggesting a presence of a significant genotype-environment interaction. The positions of the lead SNP for all identified loci were used to search for known QTL/genes reported in previous studies. In addition to two novel loci, we identified five loci that overlapped with previously identified QTLs/genes. For example, qGP2 co-localized with the regions of qDOM2.3 for seed dormancy and qPP2-1 for seed protrusion [43,44]; qGP4.1 coincided with the regions of qRI4a for anaerobic germination tolerance and qPP4 for seed protrusion [44,46]; qGP12 overlapped with the intervals of qSD12-1 for seed dormancy [47]; qGP3 was found near TE, a gene for seed germination and seedling growth [45]; and qGP7 was adjacent to OsCLSY1, a gene for anaerobic germination [16]. These observations verified the reliability of identifying genetic loci related to seed germination by GWAS. Multiple significant associations were found to be located at qGP4.1 locus in both years, and we therefore selected it for further investigation. Interestingly, the lead SNP for qGP4.1 was located in close proximity to the GP4 gene, which encoded NCED, a key enzyme for ABA biosynthesis. A sequencing and haplotype analysis demonstrated that four functional polymorphic sites (1,971,215 bp, 1,971,303 bp, 1,971,593 and 1,971,671 bp) within the coding region of GP4 were strongly associated with the germination percentage. Accessions with Hap1 of GP4 generally have a higher seed vigor than accessions with Hap2 of GP4, demonstrating that Hap1 could be used as the donor parent for breeding elite rice varieties with high seed vigor.
NCED expression is a main determinant of ABA accumulation, and increased transcript levels of NCED lead to increased ABA levels in plants [36,48]. In previous studies, several NCED mutants have been identified, and their functions in seed dormancy, seed germination, and post-germination growth have been studied. For example, Arabidopsis AtNCED6 and AtNCED9 were required for ABA biosynthesis in developing seeds, and seeds of the double mutant, Atnced6 Atnced9, showed reduced dormancy [52]. The CRISPR/Cas9-induced mutations of rice OsNCED3 resulted in decreased seed dormancy, enhanced preharvest sprouting, and increased post-germination seedling growth [53,54]. In this study, we found that the mutation of GP4 led to increased seed germination and enhanced early seedling growth. One possible explanation is that it is likely that GP4 also regulated seed germination through controlling ABA biosynthesis in seeds. However, the exact molecular mechanisms by which GP4 regulates seed germination need to be elucidated in further studies. In this study, a functional analysis in the Nipponbare background preliminarily proved that the knockout of the GP4 gene can improve seed germination in JAPONICA rice. However, a large difference in allele frequency at 1,971,215 bp and 1,971,671 bp was found between INDICA and JAPONICA (Supplementary Figure S3), suggesting a possible functional differentiation. Thus, further functional studies in INDICA rice are required to verify GP4 function.
5. Conclusions
In summary, we identified seven genetic loci for germination percentage by GWAS, and three of these loci were stable across two years, including qGP2 and qGP4.1. GP4, a NCED encoding gene, was identified as the best candidate for qGP4.1 locus. GP4 knockout using CRISPR/Cas9 displayed faster germination and seedling establishment, and suggest that GP4 is a promising genetic resource for the improvement of rice seed vigor in molecular breeding.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14091613/s1, Figure S1: LOC_Os04g04230 (GP4) lies adjacent to the lead SNP (Chr4-1,966,105) of qGP4.1 locus; Figure S2: Sequence analysis of the candidate gene GP4 from 12 accessions with C-type and 12 accessions with A-type; Figure S3: Allele frequency of four functional polymorphic sites in INDICA and JAPONICA; Table S1: List of 103 rice accessions used for GWAS. Table S2: The lead SNP (Chr4-1,966,105) genotypes of all accessions.
Author Contributions
Conceptualization, B.Y.; Investigation, B.Y., S.C., G.C., Z.P. and J.L.; Writing—original draft preparation, B.Y. and S.C.; Writing—review and editing, Y.Z. and B.Y.; Visualization, B.Y. and S.C.; Supervision, B.Y.; Project administration, B.Y. and Y.Z.; Funding acquisition, B.Y. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Open Competition Program of Top Ten Critical Priorities of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province (Grant No. 2022SDZG05, 2023SDZG05 and 2024SDZG05), the National Natural Science Foundation of China (Grant No. 32101802), the Guangzhou Basic and Applied Basic Research Foundation (Grant No. SL2022A04J01557), the Improvement of Scientific Research Capacity for Key Construction Discipline of Guangdong Province (Grant No. 2022ZDJS023), and the Science and Technology Innovation Fund for Graduate Students of Zhongkai University of Agriculture and Engineering (Grant No. KJCX2024028).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data from the experiments presented in this study are available on request to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lee, K.; Lee, H.G.; Yoon, S.; Kim, H.U.; Seo, P.J. The Arabidopsis MYB96 transcription factor is a positive regulator of ABSCISIC ACID-INSENSITIVE4 in the control of seed germination. Plant Physiol. 2015, 168, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Hou, D.; Li, Y.; Chao, H.; Zhang, K.; Wang, H.; Xiang, J.; Raboanatahiry, N.; Wang, B.; Li, M. Integration of proteomic and genomic approaches to dissect seed germination vigor in Brassica napus seeds differing in oil content. BMC Plant Biol. 2019, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- Mahender, A.; Anandan, A.; Pradhan, S.K. Early seedling vigour, an imperative trait for direct-seeded rice: An overview on physio-morphological parameters and molecular markers. Planta 2015, 241, 1027–1050. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Yang, R.; Wang, F.; Fu, J.; Yang, W.; Bai, T.; Wang, S.; Yin, H. Effects of gibberellin priming on seedling emergence and transcripts involved in mesocotyl elongation in rice under deep direct-seeding conditions. J. Zhejiang Univ. Sci. B 2021, 22, 1002–1021. [Google Scholar] [CrossRef]
- Miura, K.; Lin, S.; Yano, M.; Nagamine, T. Mapping quantitative trait loci controlling seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet. 2002, 104, 981–986. [Google Scholar] [CrossRef]
- Fujino, K.; Sekiguchi, H.; Matsuda, Y.; Sugimoto, K.; Ono, K.; Yano, M. Molecular identification of a major quantitative trait locus, qLTG3–1, controlling low-temperature germinability in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 12623–12628. [Google Scholar] [CrossRef]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice. Proc. Natl. Acad. Sci. USA 2010, 107, 5792–5797. [Google Scholar] [CrossRef]
- Ye, H.; Feng, J.; Zhang, L.; Zhang, J.; Mispan, M.S.; Cao, Z.; Beighley, D.H.; Yang, J.; Gu, X.Y. Map-Based Cloning of Seed Dormancy1-2 identified a gibberellin synthesis gene regulating the development of endosperm-imposed dormancy in rice. Plant Physiol. 2015, 169, 2152–2165. [Google Scholar]
- Lai, Y.; Cheng, J.; He, Y.; Yang, B.; Wang, Z.; Zhang, H. Identification of QTLs with additive, epistatic, and QTL × seed maturity interaction effects for seed vigor in rice. Plant Mol. Biol. Rep. 2015, 34, 160–171. [Google Scholar] [CrossRef]
- Yang, J.; Yang, G.; Yang, M.; Su, L.; Xia, A.; Li, D.; Huang, C.; Zhou, D.; Liu, Y.; Wang, H.; et al. Quantitative trait locus analysis of seed germination and early seedling growth in rice. Front. Plant Sci. 2019, 10, 1582. [Google Scholar] [CrossRef]
- Riedelsheimer, C.; Lisec, J.; Czedik-Eysenberg, A.; Sulpice, R.; Flis, A.; Grieder, C.; Altmann, T.; Stitt, M.; Willmitzer, L.; Melchinger, A.E. Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize. Proc. Natl. Acad. Sci. USA 2012, 109, 8872–8877. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R.; Hay, F.R. Variation in seed longevity among diverse Indica rice varieties. Ann. Bot. 2019, 124, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Han, B. Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 2014, 65, 531–551. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, B.; Xu, J.; Peng, L.; Sun, S.; Huang, Z.; Jiang, X.; He, Y.; Wang, Z. A genome-wide association study reveals that the 2-oxoglutarate/malate translocator mediates seed vigor in rice. Plant J. 2021, 108, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Fan, K.; Wang, Y.; Tian, L.; Zhang, C.; Sun, W.; He, H.; Yu, S. OsGRETCHENHAGEN3-2 modulates rice seed storability via accumulation of abscisic acid and protective substances. Plant Physiol. 2021, 186, 469–482. [Google Scholar] [CrossRef]
- Castano-Duque, L.; Ghosal, S.; Quilloy, F.A.; Mitchell-Olds, T.; Dixit, S. An epigenetic pathway in rice connects genetic variation to anaerobic germination and seedling establishment. Plant Physiol. 2021, 186, 1042–1059. [Google Scholar] [CrossRef]
- Yang, B.; Chen, M.; Zhan, C.; Liu, K.; Cheng, Y.; Xie, T.; Zhu, P.; He, Y.; Zeng, P.; Tang, H.; et al. Identification of OsPK5 involved in rice glycolytic metabolism and GA/ABA balance for improving seed germination via genome-wide association study. J. Exp. Bot. 2022, 73, 3446–3461. [Google Scholar] [CrossRef]
- Yoshida, H.; Hirano, K.; Yano, K.; Wang, F.; Mori, M.; Kawamura, M.; Koketsu, E.; Hattori, M.; Ordonio, R.L.; Huang, P.; et al. Genome-wide association study identifies a gene responsible for temperature-dependent rice germination. Nat. Commun. 2022, 13, 5665. [Google Scholar] [CrossRef]
- Dai, L.; Lu, X.; Shen, L.; Guo, L.; Zhang, G.; Gao, Z.; Zhu, L.; Hu, J.; Dong, G.; Ren, D.; et al. Genome-wide association study reveals novel QTLs and candidate genes for seed vigor in rice. Front. Plant Sci. 2022, 13, 1005203. [Google Scholar] [CrossRef]
- Yang, B.; Zeng, J.; Chen, S.; Li, S.; Wu, L.; Wan, X. Genome-wide association study reveals the genetic basis of seed germination in Japonica rice. Agriculture 2022, 13, 118. [Google Scholar] [CrossRef]
- Yang, B.; Chen, S.; Zheng, Z.; Zeng, J.; Liu, J.; Zhao, H.; Zheng, Y. Genome-wide association studies for rice seed germination under drought stress using 3VmrMLM. Food Energy Secur. 2024, 13, e529. [Google Scholar] [CrossRef]
- Prasad, C.T.M.; Kodde, J.; Angenent, G.C.; Hay, F.R.; McNally, K.L.; Groot, S.P.C. Identification of the rice Rc Gene as a main regulator of seed survival under dry storage conditions. Plant Cell Environ. 2023, 46, 1962–1980. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Zhu, P.; Chen, Y.; Chen, X.; Liu, K.; Chen, S.; Hu, J.; He, Y.; Xie, T.; Luo, S.; et al. Identification of a key locus, qNL3.1, associated with seed germination under salt stress via a genome-wide association study in rice. Theor. Appl. Genet. 2023, 136, 58. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ma, Y.; Zhao, H.; Guo, L.; Guo, Y.; Liu, C. Genome-wide association studies identified OsTMF as a gene regulating rice seed germination under salt stress. Front. Plant Sci. 2024, 15, 1384246. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Tuấn, P.A.; Izydorczyk, M.S.; Ayele, B.T. Ethylene regulates post-germination seedling growth in wheat through spatial and temporal modulation of ABA/GA balance. J. Exp. Bot. 2019, 71, 1985–2004. [Google Scholar] [CrossRef]
- Cheng, C.; Wang, Z.; Ren, Z.; Zhi, L.; Yao, B.; Su, C.; Liu, L.; Li, X. SCFAtPP2-B11 Modulates ABA signaling by facilitating SnRK2.3 degradation in Arabidopsis thaliana. PLoS Genet. 2017, 13, e1006947. [Google Scholar] [CrossRef]
- Cutler, A.J.; Krochko, J.E. Formation and breakdown of ABA. Trends Plant Sci. 1999, 4, 472–478. [Google Scholar] [CrossRef]
- Yan, J.; Zhao, C.; Zhou, J.; Ye, Y.; Wang, P.; Zhu, X.; Tang, G.; Bressan, R.A.; Zhu, J. The miR165/166 mediated regulatory module plays critical roles in ABA homeostasis and response in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006416. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Kim, D.H.; Hwang, I. ABA homeostasis and signaling involving multiple subcellular compartments and multiple receptors. Plant Cell Rep. 2013, 32, 807–813. [Google Scholar] [CrossRef]
- Hauser, F.; Waadt, R.; Schroeder, J.I. Evolution of abscisic acid synthesis and signaling mechanisms. Curr. Biol. 2011, 21, R346–R355. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A.D. Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.H.; Tan, B.C.; Gage, D.A.; Zeevaart, J.A.D.; McCarty, D.R. Specific oxidative cleavage of carotenoids by VP14 of maize. Science 1997, 276, 1872–1874. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.C.; Schwartz, S.H.; Zeevaart, J.A.D.; McCarty, D.R. Genetic control of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 1997, 94, 12235–12240. [Google Scholar] [CrossRef] [PubMed]
- Burbidge, A.; Grieve, T.M.; Jackson, A.; Thompson, A.; McCarty, D.R.; Taylor, I.B. Characterization of the ABA-deficient tomato mutant notabilis and its relationship with maize Vp14. Plant J. 1999, 17, 427–431. [Google Scholar] [CrossRef]
- Chernys, J.T.; Zeevaart, J.A.D. Characterization of the 9-cis-epoxycarotenoid dioxygenase gene family and the regulation of abscisic acid biosynthesis in avocado. Plant Physiol. 2000, 124, 343–354. [Google Scholar] [CrossRef]
- Qin, X.; Zeevaart, J.A.D. Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance. Plant Physiol. 2002, 128, 544–551. [Google Scholar] [CrossRef]
- Zhao, K.; Tung, C.-W.; Eizenga, G.C.; Wright, M.H.; Ali, M.L.; Price, A.H.; Norton, G.J.; Islam, M.R.; Reynolds, A.; Mezey, J.; et al. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun. 2011, 2, 467. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Bao, Y.; Wang, F.; Zhang, H. Quantitative trait loci analysis for rice seed vigor during the germination stage. J. Zhejiang Univ. Sci. B 2010, 11, 958–964. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.A.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- Xing, H.L.; Dong, L.; Wang, Z.P.; Zhang, H.Y.; Han, C.Y.; Liu, B.; Wang, X.C.; Chen, Q.J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Toki, S.; Hara, N.; Ono, K.; Onodera, H.; Tagiri, A.; Oka, S.; Tanaka, H. Early infection of scutellum tissue with Agrobacteriu allows high-speed transformation of rice. Plant J. 2006, 47, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Wang, Y.; Zhang, C.; He, H.; Yu, S. Genetic dissection of seed dormancy using chromosome segment substitution lines in rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1344. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Shi, J.; Gui, J.; Zhou, H.; Yan, Y.; Zhu, X.; Xie, B.; Liu, X.; He, J. Rice seed protrusion quantitative trait loci mapping through genome-wide association study. Plants 2024, 13, 134. [Google Scholar] [CrossRef]
- Lin, Q.; Wu, F.; Sheng, P.; Zhang, Z.; Zhang, X.; Guo, X.; Wang, J.; Cheng, Z.; Wang, J.; Wang, H.; et al. The SnRK2-APC/CTE regulatory module mediates the antagonistic action of gibberellic acid and abscisic acid pathways. Nat. Commun. 2015, 2, 7981. [Google Scholar] [CrossRef]
- Islam, M.R.; Naveed, S.A.; Zhang, Y.; Li, Z.; Zhao, X.; Fiaz, S.; Zhang, F.; Wu, Z.; Hu, Z.; Fu, B.; et al. Identification of candidate genes for salinity and anaerobic tolerance at the germination stage in rice by genome-wide association analyses. Front. Genet. 2022, 13, 822516. [Google Scholar] [CrossRef]
- Zhou, Y.; Xie, Y.; Cai, J.; Liu, C.; Zhu, H.; Jiang, R.; Zhong, Y.; Zhang, G.; Tan, B.; Liu, G.; et al. Substitution mapping of qtls controlling seed dormancy using single segment substitution lines derived from multiple cultivated rice donors in seven cropping seasons. Theor. Appl. Genet. 2017, 130, 1191–1205. [Google Scholar] [CrossRef]
- Martínez-Andújar, C.; Ordiz, M.I.; Huang, Z.; Nonogaki, M.; Beachy, R.N.; Nonogaki, H. Induction of 9-cis-epoxycarotenoid dioxygenase in Arabidopsis Thaliana seeds enhances seed dormancy. Proc. Natl. Acad. Sci. USA 2011, 108, 17225–17229. [Google Scholar] [CrossRef]
- Huo, H.; Dahal, P.; Kunusoth, K.; McCallum, C.M.; Bradford, K.J. Expression of 9-cis-EPOXYCAROTENOID DIOXYGENASE4 is essential for thermoinhibition of lettuce seed germination but not for seed development or stress tolerance. Plant Cell 2013, 25, 884–900. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, J.; Yang, T.; Dong, J.; Yang, W.; Chen, L.; Zhou, L.; Chen, J.; Liu, B.; Zhang, S.; et al. Genome-wide association mapping and gene expression analysis identify OsCPS1 as a new candidate gene controlling early seedling length in rice. Front. Plant Sci. 2022, 13, 976669. [Google Scholar] [CrossRef]
- Kadam, N.N.; Jagadish, K.; Struik, P.C.; van der Linden, C.G.; Yin, X. Incorporating genome-wide association into eco-physiological simulation to identify markers for improving rice yields. J. Exp. Bot. 2019, 70, 2575–2586. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V.; North, H.; Frey, A.; Sotta, B.; Seo, M.; Okamoto, M.; Nambara, E.; Marion-Poll, A. Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J. 2006, 45, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Guo, Y.; Liu, Y.; Zhang, F.; Wang, Z.; Wang, H.; Wang, F.; Li, D.; Mao, D.; Luan, S.; et al. 9-cis-epoxycarotenoid dioxygenase 3 regulates plant growth and enhances multi-abiotic stress tolerance in rice. Front. Plant Sci. 2018, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiang, Z.; Liu, M.; Wang, S.; Zhang, L.; Cai, D.; Huang, Y.; Mao, D.; Fu, J.; Chen, L. ABA biosynthesis gene OsNCED3 contributes to preharvest sprouting resistance and grain development in rice. Plant Cell Environ. 2022, 46, 1384–1401. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).