Screening for Salt Tolerance in Four Local Varieties of Phaseolus lunatus from Spain

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growth and Stress Treatments
2.3. Soil Analysis
2.4. Plant Growth Parameters
2.5. Photosynthetic Pigments
- Chl a (µg mL−1) = 12.21 (A663) − 2.81 (A646),
- Chl b (µg mL−1) = 20.13 (A646) − 5.03 (A663),
- Caro (µg mL−1) = (1000A470 − 3.27[chl a] − 104[chl b])/227.
- Final values were expressed in mg g−1 DW.
2.6. Ion Concentration Measurements
2.7. Osmolyte Quantification
2.8. Statistical Analysis
3. Results
3.1. Substrate and Plant Growth Analyses
3.2. Photosynthetic Pigments
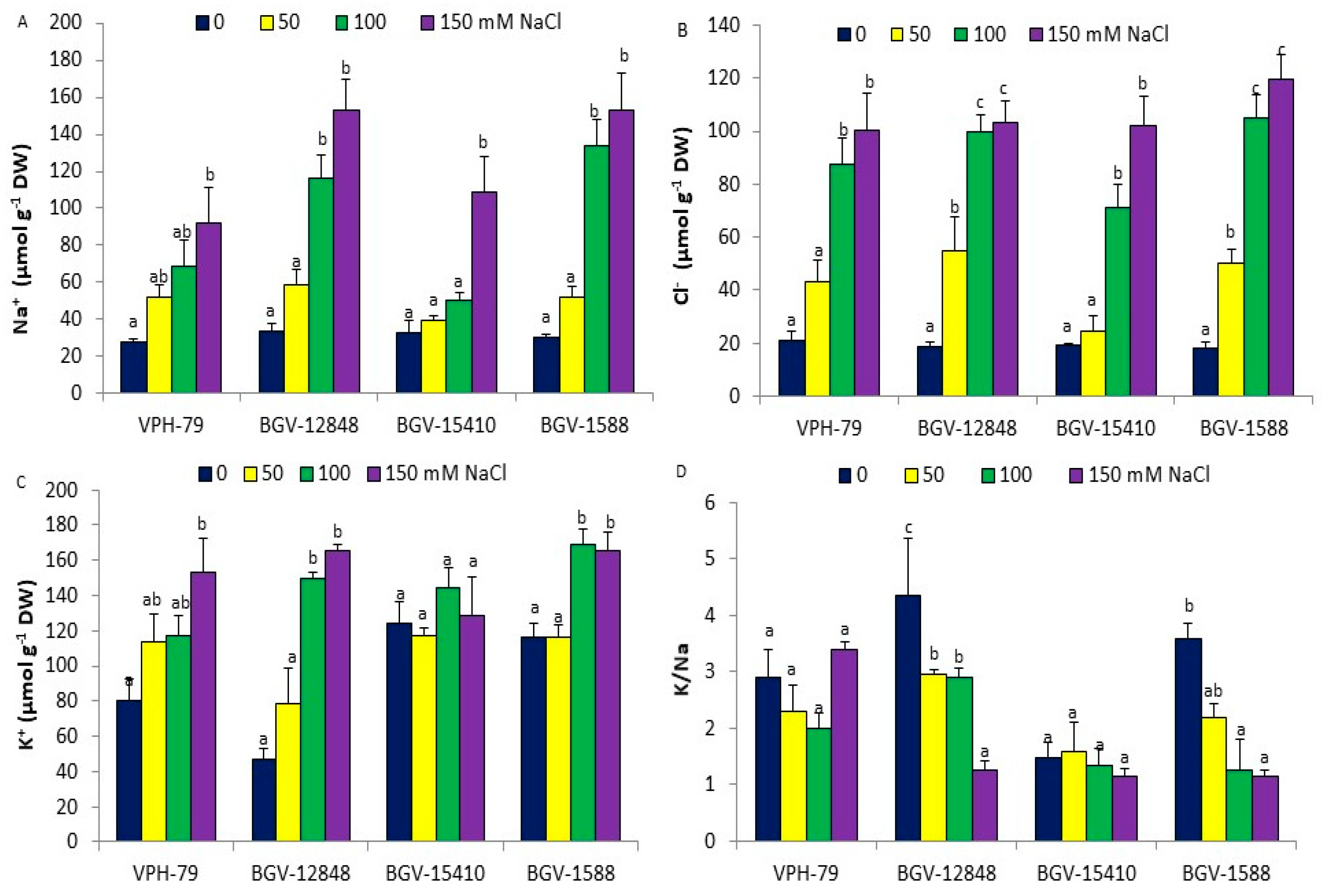
3.3. Ions Accumulation
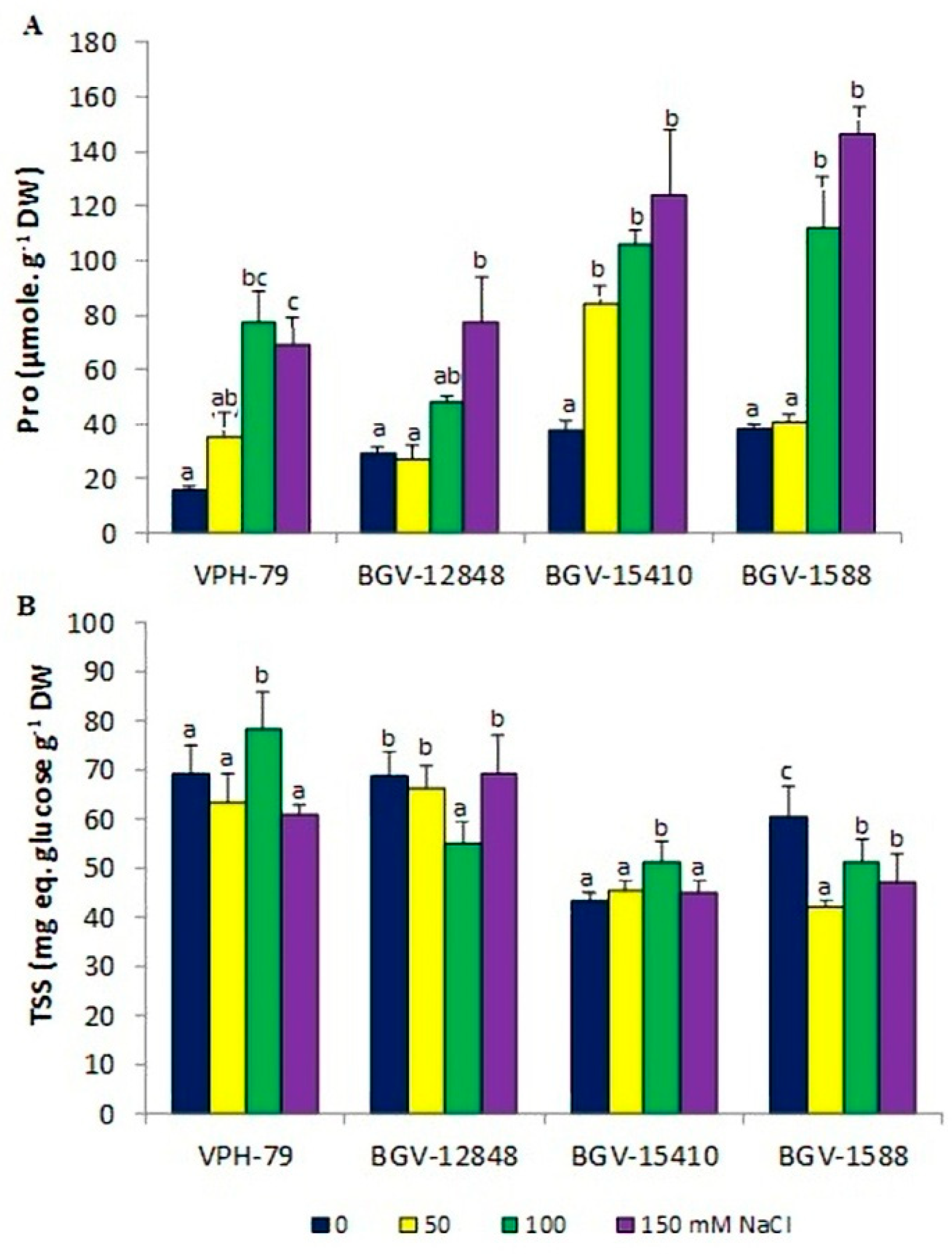
3.4. Osmolytes Accumulation
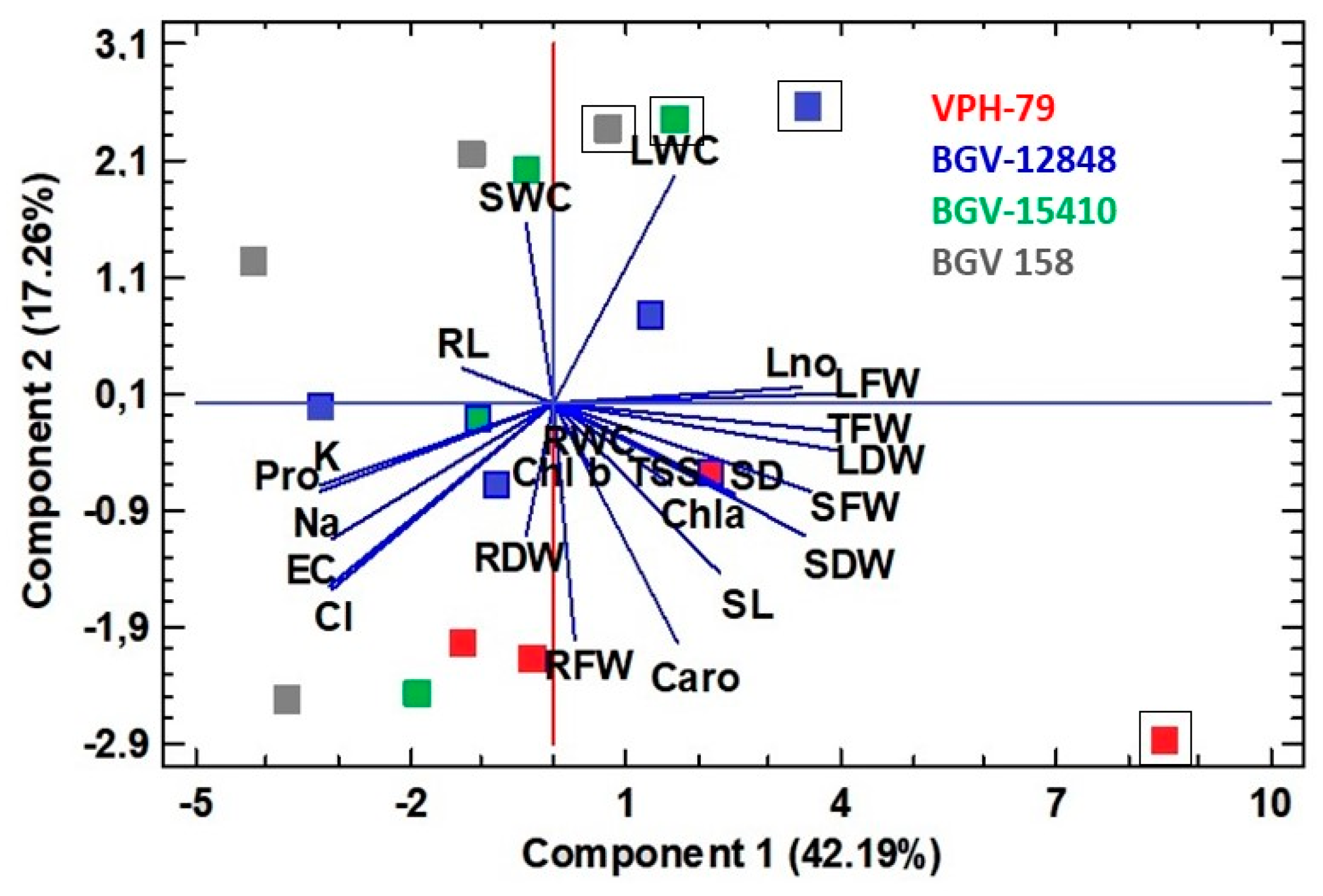
3.5. Principal Component Analysis (PCA)
4. Discussion
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- López-Alcocer, J.J.; Lépiz-Ildefonso, R.; González-Eguiarte, D.R.; Rodríguez-Macías, R.; López-Alcocer, E. Morphological variability of wild Phaseolus lunatus from the western region of Mexico. Rev. Fitotec. Mex. 2016, 39, 49–58. [Google Scholar]
- Martínez-Castillo, J.; Camacho-Pérez, L.; Villanueva-Viramontes, S.; Andueza-Noh, R.H.; Chacón-Sánchez, M.I. Genetic structure within the Mesoamerican gene pool of wild Phaseolus lunatus (Fabaceae) from Mexico as revealed by microsatellite markers: Implications for conservation and the domestication of the species. Am. J. Bot. 2014, 101, 851–864. [Google Scholar] [CrossRef]
- Baudoin, J.P. Phaseolus lunatus L. In Protabase; Brink, M., Belay, G., Eds.; PROTA (Plant Resources of Tropical Africa/Ressources végétales de l’Afrique tropicale): Wageningen, The Netherlands, 2006. [Google Scholar]
- De Ron, A.M.; Santalla, M.; Rodiño, A.P.; González, A.M.; Godoy, L.; Mansilla, J.P.; Blair, M. Judia. In Las Variedades Locales en la Mejora Genética de Plantas; Ruiz de Galarreta, J.I., Prohens, J., Tierno, R., Eds.; Servicio Central de Publicaciones del Gobierno Vasco: Donostia-San Sebastián, Spain, 2016; pp. 155–170. ISBN 978-84-457-3395-0. [Google Scholar]
- Casado, S.; Gonzáles, J.M.; Varela, F.; Rosselló, J.; Carrascosa, M.; Soriano, J.J.; Camarillo, J.M. Estudio Diagnóstico Sobre la Biodiversidad Cultivada y la Agricultura Ecológica; Sociedad Española de Agricultura Ecológica—Red de Semillas “Resembrando e Intercambiando”: Seville, Spain, 2009. [Google Scholar]
- Jackson, M.; Ford-Lloyd, B.; Parry, M. Plant Genetic Resources and Climate Change; CABI Climate Change: Wallingford, UK, 2013; Volume 4. [Google Scholar]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Intergovernmental panel on climate change. In Proceeding of the 5th Assessment Report, WGII, Climate Change 2014: Impacts, Adaptation, and Vulnerability; Cambridge University Press: Cambridge, UK; Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 15 July 2018).
- Owens, S. Salt of the earth. Genetic engineering may help to reclaim agricultural land lost due to salinisation. EMBO Rep. 2001, 2, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Grattan, S.; Grieve, C.M. Salinity–mineral nutrient relations in horticultural crops. Sci. Hortic. 1999, 78, 127–157. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genomics 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef]
- Bayuelo-Jiménez, J.S.; Debouck, D.G.; Lynch, J.P. Salinity Tolerance in Phaseolus species during early vegetative growth. Crop Sci. 2002, 42, 2184–2192. [Google Scholar] [CrossRef]
- Rodrigues Do Nascimento, M.G.; Ursulino Alves, E.; Mauricio da Silva, M.L.; Marques Rodrigues, C. Lima bean (Phaseolus lunatus L.) seeds exposed to different salt concentrations and temperatures. Rev. Caatinga 2017, 30, 738–747. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; Circular 347, 2nd ed.; University of California Agricultural Experiment Station: Berkeley, CA, USA, 1950. [Google Scholar]
- Gil, R.; Bautista, I.; Boscaiu, M.; Lidón, A.; Wankhade, S.; Sánchez, H.; Lilinares, J.; Vicente, O. Responses of five Mediterranean halophytes to seasonal changes in environmental conditions. AoB Plants 2014, 6, plu049. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Weimberg, R. Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plant. 1987, 70, 381–388. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Reberd, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Boscaiu, M. Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 2016, 11, e0160236. [Google Scholar] [CrossRef] [PubMed]
- Taffouo, V.D.; Wamba, O.F.; Youmbi, E.; Nono, G.V.; Akoa, A. Growth, yield, water status and ionic distribution response of three bambara groundnut (Vigna subterranea (L.) Verdc.) landraces grown under saline conditions. Int. J. Bot. 2010, 6, 53–58. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defense systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Soussi, M.; Ocana, A.; Lluch, C. Effect of salt stress on growth, photosynthesis and nitrogen fixation in chickpea (Cicer arietinum L.). J. Exp. Bot. 1998, 49, 1329–1337. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hort. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Jacobi, B. Function of bean roots and stems in sodium retention. Plant Physiol. 1964, 39, 445–449. [Google Scholar] [CrossRef]
- Kramer, D.; Läuchli, A.; Yeo, A.R.; Gullasch, J. Transfer cells in roots of Phaseolus coccineus: Ultrastructure and possible function in exclusion of sodium from the shoot. Ann. Bot. 1977, 41, 1031–1040. [Google Scholar] [CrossRef]
- Seemann, J.R.; Critchley, C. Effects of salt stress on the growth, ion content, stomatal behaviour and photosynthetic capacity of a salt-sensitive species, Phaseolus vulgaris L. Planta 1985, 164, 151–162. [Google Scholar] [CrossRef]
- Bayuelo-Jiménez, J.S.; Jasso-Plata, N.; Ochoa, I. Growth and physiological responses of Phaseolus species to salinity stress. Int. J. Agron. 2012, 80, 207–222. [Google Scholar] [CrossRef]
- Gama, P.B.S.; Inanaga, S.; Tanaka, K.; Nakazawa, R. Physiological response of common bean. (Phaseolus vulgaris L.) seedlings to salinity stress. Afr. J. Biotechnol. 2007, 6, 79–88. [Google Scholar]
- Al Hassan, M.; Morosan, M.; López-Gresa, M.P.; Prohens, J.; Vicente, O.; Boscaiu, M. Salinity-induced variation in biochemical markers provides insight into the mechanisms of salt tolerance in common (Phaseolus vulgaris) and runner (P. coccineus) beans. Int. J. Mol. Sci. 2016, 17, 1582. [Google Scholar] [CrossRef] [PubMed]
- Gierth, M.; Mäser, P. Potassium transporters in plants- Involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 2007, 581, 2348–2356. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Navarro, A.; Rubio, F. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.; Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkov, V.; Wang, B.; Dominy, P.J.; Fricke, W.; Amtmann, A. Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ. 2003, 27, 1–14. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Saradhi, P.; Alia, P.; Arora, S.; Prasad, K.V. Proline accumulates in plants exposed to UV radiation and protects them against UV induced peroxidation. Biochem. Biophys. Res. Commun. 1995, 209, 1–5. [Google Scholar] [CrossRef]
- Siripornadulsil, S.; Train, S.; Verma, D.P.S.; Sayre, R.T. Molecular mechanisms of proline mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Cárdenas-Avila, M.L.; Verde-Star, J.; Maiti, R.K.; Foroughbakhch-P, R.; Gámez-González, H.; Martínez-Lozano, S.; Núñez-González, M.A.; García Díaz, G.; Hernández-Piñero, J.L.; Morales-Vallarta, M.R. Variability in accumulation of free proline on in vitro calli of four bean (Phaseolus vulgaris L.) cultivars exposed to salinity and induced moisture stress. Phyton 2006, 75, 103–108. [Google Scholar]
- Jiménez-Bremont, J.F.; Becerra-Flora, A.; Hernández-Lucero, E.; Rodríguez-Kessler, M.; Acosta-Gallegos, J.A.; Ramírez Pimentel, J.G. Proline accumulation in two bean cultivars under salt stress and the effect of polyamines and ornithine. Biol. Plant. 2006, 50, 763–766. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Treatment (mM NaCl) | VPH-79 | BGV-12848 | BGV-15410 | BGV 1588 |
|---|---|---|---|---|---|
| Soil EC1:5 | 0 | 0.40 ± 0.03 a | 0.33 ± 0.05 a | 0.28 ± 0.02 a | 0.47 ± 0.14 a |
| 50 | 0.81 ± 0.12 a | 1.02 ± 0.11 ab | 1.02 ± 0.29 b | 1.09 ± 0.21 ab | |
| 100 | 1.57 ± 0.11 b | 1.39 ± 0.15 b | 1.68 ± 0.07 c | 1.31 ± 0.19 b | |
| 150 | 1.82 ± 0.15 b | 1.81 ± 0.44 b | 1.81 ± 0.05 c | 1.77 ± 0.07 b | |
| p | 0.000 | 0.000 | 0.000 | 0.000 | |
| Root length (cm) | 0 | 27.20 ± 3.33 a | 26.60 ± 1.80 a | 39.75 ± 3.60 | 45.20 ± 3.82 |
| 50 | 31.50 ± 1.37 a | 32.50 ± 1.5 b | 33.00 ± 2.58 | 35.67 ± 2.40 | |
| 100 | 40.00 ± 1.15 b | 25.20 ± 1.95 a | 37.50 ± 4.33 | 34.00 ± 4.97 | |
| 150 | 27.00 ± 3.05 a | 30.00 ± 1.00 b | 39.00 ± 3.90 | 41.00 ± 6.54 | |
| p | 0.000 | 0.026 | 0.497 | 0.297 | |
| Root FW (g) | 0 | 4.65 ± 0.83 | 3.02 ± 0.59 | 2.78 ± 0.18 a | 2.48 ± 0.17 ab |
| 50 | 3.54 ± 0.41 | 2.81 ± 0.41 | 2.59 ± 0.36 a | 1.99 ± 0.47 a | |
| 100 | 3.67 ± 0.35 | 2.73 ± 0.41 | 4.17 ± 0.59 ab | 3.37 ± 0.80 ab | |
| 150 | 3.81 ± 1.14 | 2.52 ± 0.37 | 6.24 ± 1.07 b | 4.03 ± 0.56 b | |
| p | 0.240 | 0.882 | 0.001 | 0.046 | |
| Root WC (%) | 0 | 88.60 ± 0.46 | 91.18 ± 1.02 | 83.71 ± 1.25 | 84.08 ± 6.67 |
| 50 | 89.09 ± 0.84 | 98.87 ± 0.51 | 86.26 ± 2.07 | 84.03 ± 0.98 | |
| 100 | 87.64 ± 0.24 | 91.01 ± 0.70 | 81.83 ± 4.61 | 87.74 ± 0.75 | |
| 150 | 88.23 ± 5.50 | 89.17 ± 0.52 | 85.33 ± 0.73 | 87.14 ± 0.59 | |
| p | 0.445 | 0.462 | 0.868 | 0.109 | |
| Stem length (cm) | 0 | 188.56 ± 0.85 | 115.00 ± 12.7 | 131.40 ± 11.1 | 162.20 ± 11.5 b |
| 50 | 162.67 ± 15.9 | 150.25 ± 8.51 | 148.90 ± 25.9 | 121.20 ± 11.5 ab | |
| 100 | 172.83 ± 11.7 | 147.20 ± 9.98 | 129.50 ± 6.85 | 88.20 ± 5.04 a | |
| 150 | 155.33 ± 21.4 | 115.33 ± 11.6 | 159.00 ± 7.37 | 132.75 ± 21.5 ab | |
| p | 0.073 | 0.068 | 0.636 | 0.002 | |
| Stem diameter (cm) | 0 | 5.88 ± 0.16 b | 6.01 ± 0.09 | 5.00 ± 0.36 | 4.49 ± 0.33 ab |
| 50 | 4.75 ± 0.41 a | 5.09 ± 0.36 | 4.49 ± 0.21 | 5.03 ± 0.24 b | |
| 100 | 4.91 ± 0.14 ab | 5.36 ± 0.34 | 5.16 ± 0.25 | 3.99 ± 0.11 a | |
| 150 | 5.34 ± 0.29 ab | 4.13 ± 0.60 | 4.49 ± 0.21 | 4.92 ± 0.20 b | |
| p | 0.037 | 0.079 | 0.306 | 0.014 | |
| Stem FW (g) | 0 | 30.79 ± 2.73 b | 14.99 ± 5.12 | 17.54 ± 2.08 | 16.78 ± 2.85 b |
| 50 | 16.77 ± 4.12 a | 12.81 ± 2.78 | 10.62 ± 1.87 | 11.79 ± 1.17 ab | |
| 100 | 13.61 ± 1.24 a | 11.65 ± 1.48 | 11.20 ± 2.35 | 7.77 ± 1.12 a | |
| 150 | 12.78 ± 1.33 a | 6.96 ± 0.93 | 13.02 ± 1.67 | 13.89 ± 0.79 ab | |
| p | 0.004 | 0.257 | 0.098 | 0.011 | |
| Stem WC (%) | 0 | 81.22 ± 0.88 a | 85.53 ± 2.12 a | 84.29 ± 2.57 a | 83.25 ± 10.55 a |
| 50 | 77.90 ± 0.76 a | 80.93 ± 2.03 a | 83.07 ± 2.45 a | 81.50 ± 1.36 a | |
| 100 | 79.00 ± 1.47 a | 81.01 ± 0.55 a | 83.07 ± 1.25 a | 84.67 ± 1.03 a | |
| 150 | 78.28 ± 1.51 a | 81.90 ± 0.55 a | 81.76 ± 0.57 a | 83.37 ± 0.49 a | |
| p | 0.162 | 0.107 | 0.897 | 0.153 | |
| Leaf number | 0 | 44.80 ± 2.80 b | 19.20 ± 4.02 | 33.20 ± 4.04 b | 27.20 ± 3.70 b |
| 50 | 22.83 ± 5.61 a | 16.25 ± 4.44 | 18.60 ± 3.64 ab | 19.00 ± 1.89 ab | |
| 100 | 17.83 ± 0.98 a | 15.20 ± 1.15 | 24.67 ± 3.65 ab | 10.60 ± 1.16 a | |
| 150 | 12.67 ± 1.45 a | 9.33 ± 1.52 | 13.75 ± 3.38 a | 10.80 ± 1.73 a | |
| p | 0.001 | 0.115 | 0.020 | 0.000 | |
| Leaf FW (g) | 0 | 61.18 ± 5.53 b | 42.49 ± 8.25 b | 25.05 ± 3.64 b | 20.26 ± 3.47 b |
| 50 | 26.80 ± 9.83 a | 25.05 ± 3.64 a | 20.95 ± 3.91 b | 13.05 ± 2.68 ab | |
| 100 | 12.12 ± 2.44 a | 18.85 ± 2.51 a | 19.23 ± 4.20 b | 5.08 ± 1.90 a | |
| 150 | 8.89 ± 1.40 a | 7.04 ± 1.29 a | 9.82 ± 4.16 a | 5.08 ± 1.90 a | |
| p | 0.000 | 0.015 | 0.020 | 0.000 | |
| Leaf WC (%) | 0 | 85.27 ± 0.18 a | 89.88 ± 2.33 b | 86.53 ± 0.91 | 84.78 ± 0.60 |
| 50 | 82.63 ± 2.29 a | 85.14 ± 0.65 ab | 87.98 ± 2.12 | 84.78 ± 0.60 | |
| 100 | 81.64 ± 2.26 a | 84.66 ± 0.28 ab | 79.24 ± 10.64 | 85.09 ± 0.41 | |
| 150 | 78.66 ± 3.57 a | 83.07 ± 1.52 a | 80.99 ± 4.18 | 78.89 ± 6.49 | |
| p | 0.000 | 0.035 | 0.610 | 0.104 | |
| Total FW Reduction (%) | 0 | 0 a | 0 a | 0 a | 0 a |
| 50 | 51.25 b | 32.77 b | 38.21 b | 32.09 b | |
| 100 | 69.57 bc | 45.08 b | 24.67 b | 55.73 b | |
| 150 | 73.63 c | 59.39 b | 36.68 b | 41.78 b | |
| p | 0.000 | 0.000 | 0.043 | 0.000 |
| Parameter | Treatment (VAR 1) | Species (VAR 2) | VAR1 × VAR2 |
|---|---|---|---|
| EC | 0.000 | 0.932 | 0.785 |
| Root length | 0.871 | 0.000 | 0.004 |
| Root FW | 0.002 | 0.001 | 0.012 |
| Root WC | 0.493 | 0.557 | 0.309 |
| Stem diameter | 0.082 | 0.057 | 0.027 |
| Stem length | 0.404 | 0.000 | 0.017 |
| Stem FW | 0.000 | 0.000 | 0.033 |
| Stem WC | 0.063 | 0.000 | 0.763 |
| Leaves no. | 0.000 | 0.000 | 0.763 |
| Leaf FW | 0.000 | 0.000 | 0.000 |
| Leaf WC | 0.044 | 0.232 | 0.632 |
| Total FW | 0.000 | 0.000 | 0.000 |
| Chl a | 0.647 | 0.000 | 0.715 |
| Chl b | 0.785 | 0.989 | 0.008 |
| Caro | 0.014 | 0.000 | 0.000 |
| Pro | 0.000 | 0.000 | 0.002 |
| Na+ | 0.000 | 0.000 | 0.010 |
| K+ | 0.000 | 0.002 | 0.010 |
| Cl− | 0.000 | 0.003 | 0.606 |
| K+/Na+ | 0.000 | 0.000 | 0.009 |
| TSS | 0.539 | 0.000 | 0.041 |
| Pigment (mg g−1 DW) | Treatment (mM NaCl) | VPH-79 | BGV-12848 | BGV-15410 | BGV 1588 |
|---|---|---|---|---|---|
| Chl a | 0 | 6.33 ± 0.07 | 6.24 ± 0.77 | 5.14 ± 0.44 | 4.17 ± 0.66 |
| 50 | 5.60 ± 0.44 | 5.71 ± 0.85 | 6.05 ± 0.37 | 3.81 ± 0.27 | |
| 100 | 5.41 ± 0.97 | 6.50 ± 0.98 | 5.84 ± 1.02 | 3.03 ± 0.85 | |
| 150 | 5.25 ± 0.28 | 6.02 ± 0.65 | 5.25 ± 0.28 | 3.89 ± 0.88 | |
| p | 0.560 | 0.669 | 0.917 | 0.315 | |
| Chl b | 0 | 3.17 ± 0.49 | 2.37 ± 0.26 | 2.80 ± 0.22 | 2.22 ± 0.07 |
| 50 | 2.35 ± 0.26 | 2.14 ± 0.39 | 2.43 ± 0.41 | 2.38 ± 0.11 | |
| 100 | 1.89 ± 0.39 | 2.32 ± 0.18 | 2.44 ± 0.78 | 2.77 ± 0.80 | |
| 150 | 1.58 ± 0.13 | 2.13 ± 0.14 | 1.93 ± 0.69 | 2.02 ± 0.38 | |
| p | 0.054 | 0.985 | 0.521 | 0.77 | |
| Caro | 0 | 2.69 ± 1.04 b | 0.67 ± 0.09 | 0.49 ± 0.04 | 0.54 ± 0.15 a |
| 50 | 0.97 ± 0.25 a | 0.91 ± 0.16 | 0.73 ± 0.14 | 0.63 ± 0.21 ab | |
| 100 | 1.26 ± 0.13 a | 0.90 ± 0.14 | 0.62 ± 0.22 | 0.84 ± 0.17 ab | |
| 150 | 1.48 ± 0.16 ab | 1.29 ± 0.14 | 1.08 ± 0.24 | 1.32 ± 0.11 b | |
| p | 0.045 | 0.263 | 0.087 | 0.030 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arteaga, S.; Al Hassan, M.; Chaminda Bandara, W.M.; Yabor, L.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Screening for Salt Tolerance in Four Local Varieties of Phaseolus lunatus from Spain. Agriculture 2018, 8, 201. https://doi.org/10.3390/agriculture8120201
Arteaga S, Al Hassan M, Chaminda Bandara WM, Yabor L, Llinares JV, Boscaiu M, Vicente O. Screening for Salt Tolerance in Four Local Varieties of Phaseolus lunatus from Spain. Agriculture. 2018; 8(12):201. https://doi.org/10.3390/agriculture8120201
Chicago/Turabian StyleArteaga, Sugenith, Mohamad Al Hassan, Wijesinghe M. Chaminda Bandara, Lourdes Yabor, Josep V. Llinares, Monica Boscaiu, and Oscar Vicente. 2018. "Screening for Salt Tolerance in Four Local Varieties of Phaseolus lunatus from Spain" Agriculture 8, no. 12: 201. https://doi.org/10.3390/agriculture8120201
APA StyleArteaga, S., Al Hassan, M., Chaminda Bandara, W. M., Yabor, L., Llinares, J. V., Boscaiu, M., & Vicente, O. (2018). Screening for Salt Tolerance in Four Local Varieties of Phaseolus lunatus from Spain. Agriculture, 8(12), 201. https://doi.org/10.3390/agriculture8120201