Seasonal Soil Respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands

 ,
,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
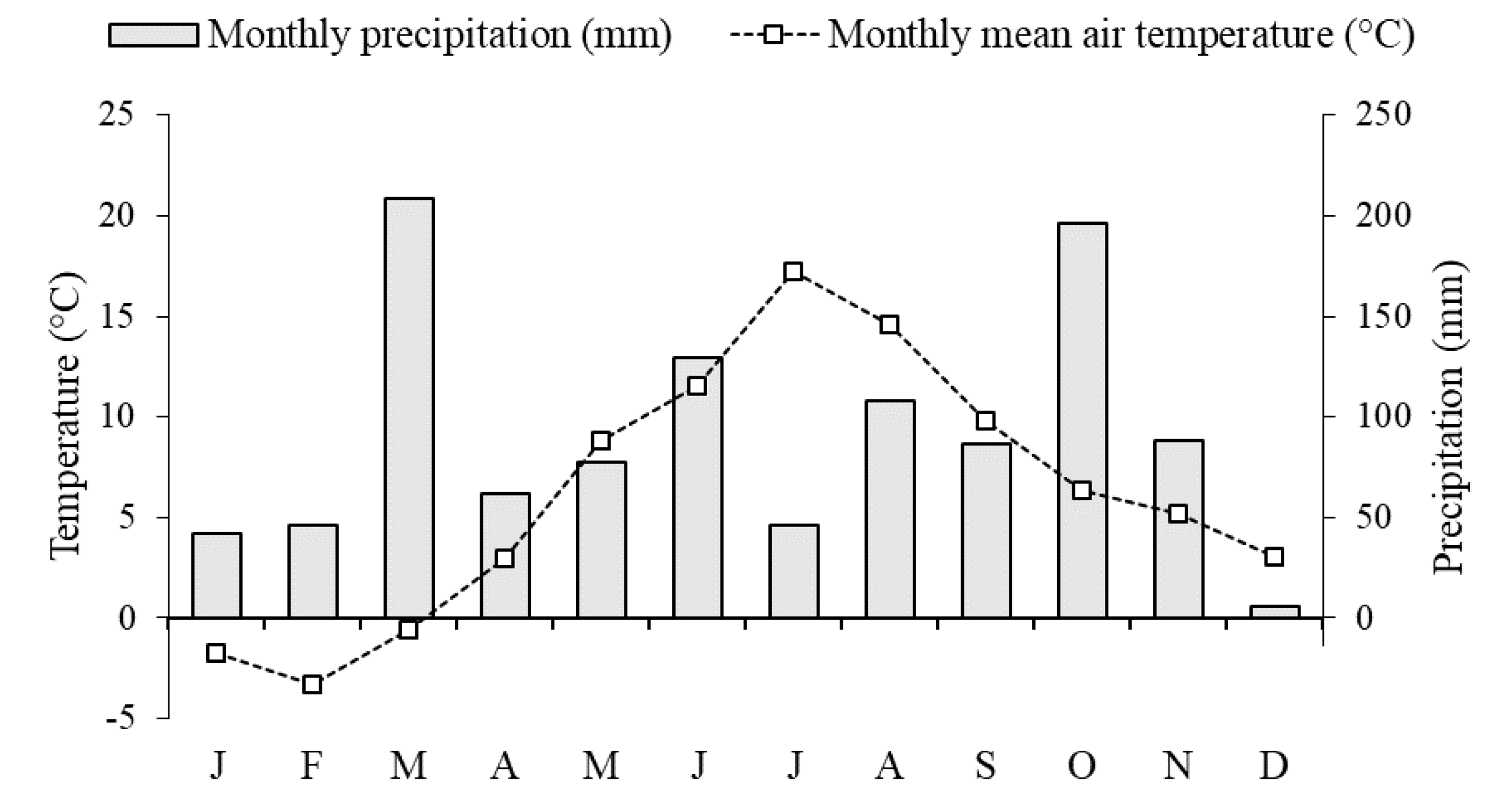
2.1. Study Area
2.2. Soil CO2 Efflux, Temperature and Water Content Analysis
2.3. Data Analysis
3. Results and Discussion
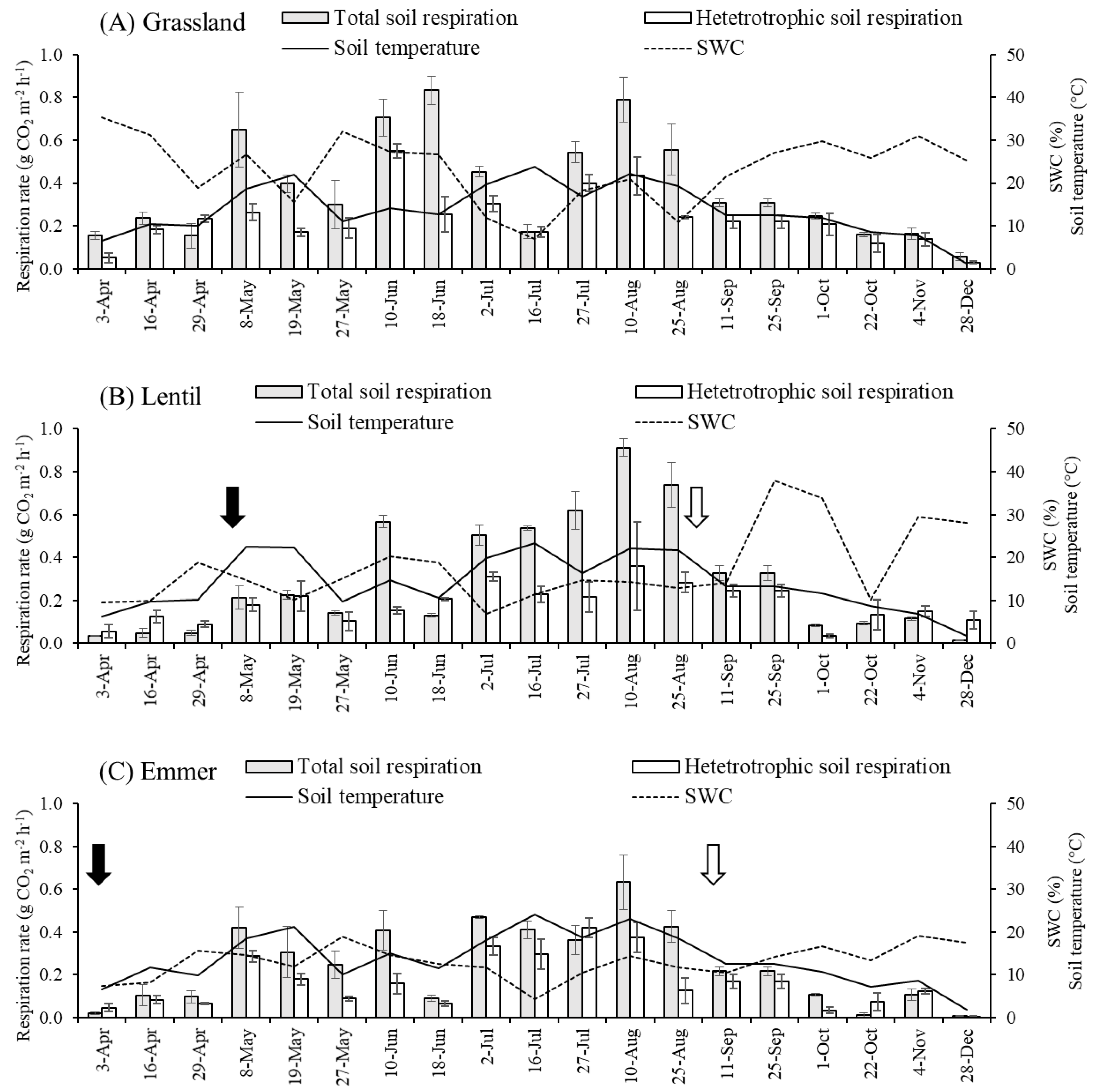
3.1. Soil CO2 Dynamics
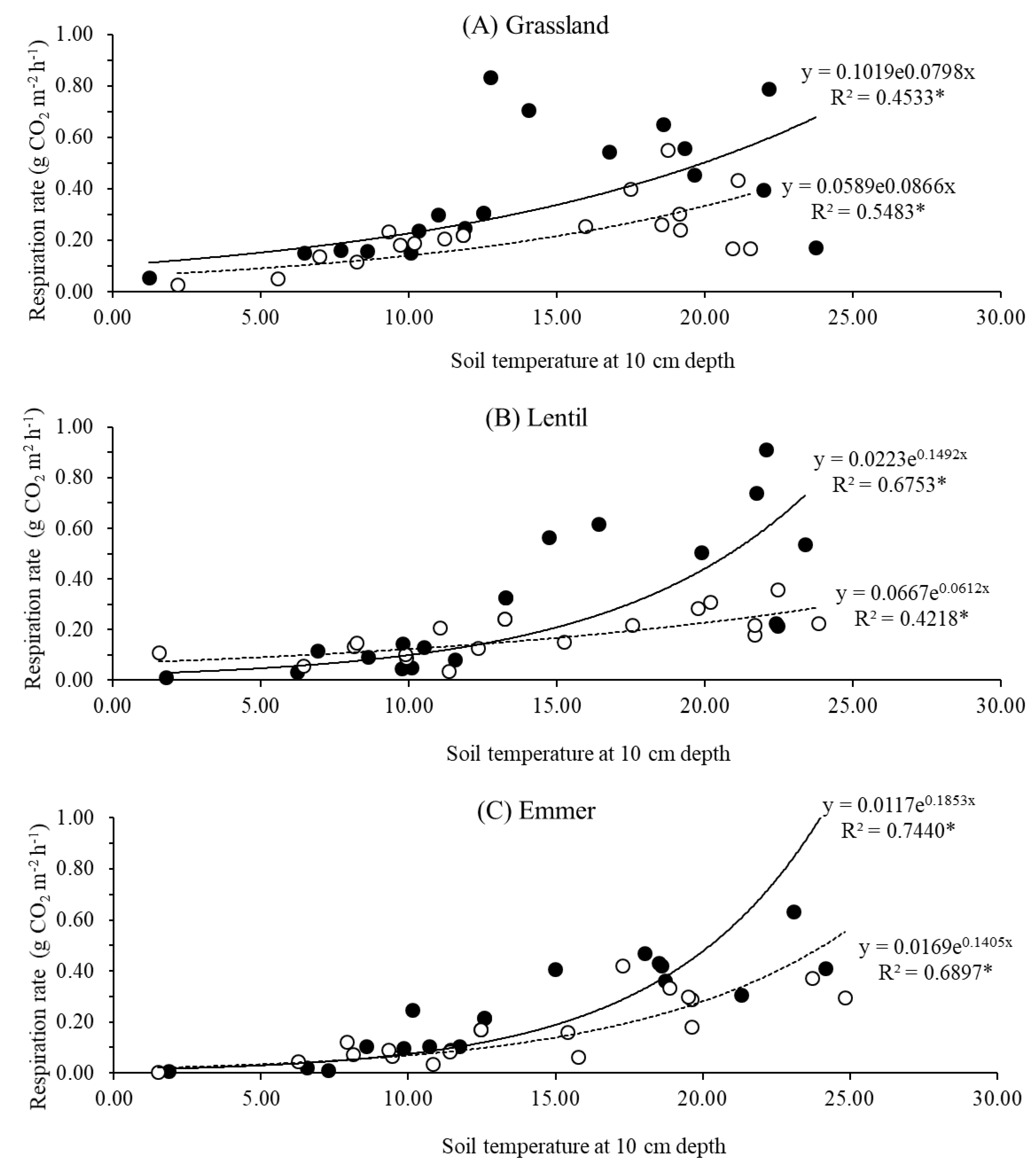
3.2. Relationships between Soil CO2 Efflux and Soil Temperature and Water Content
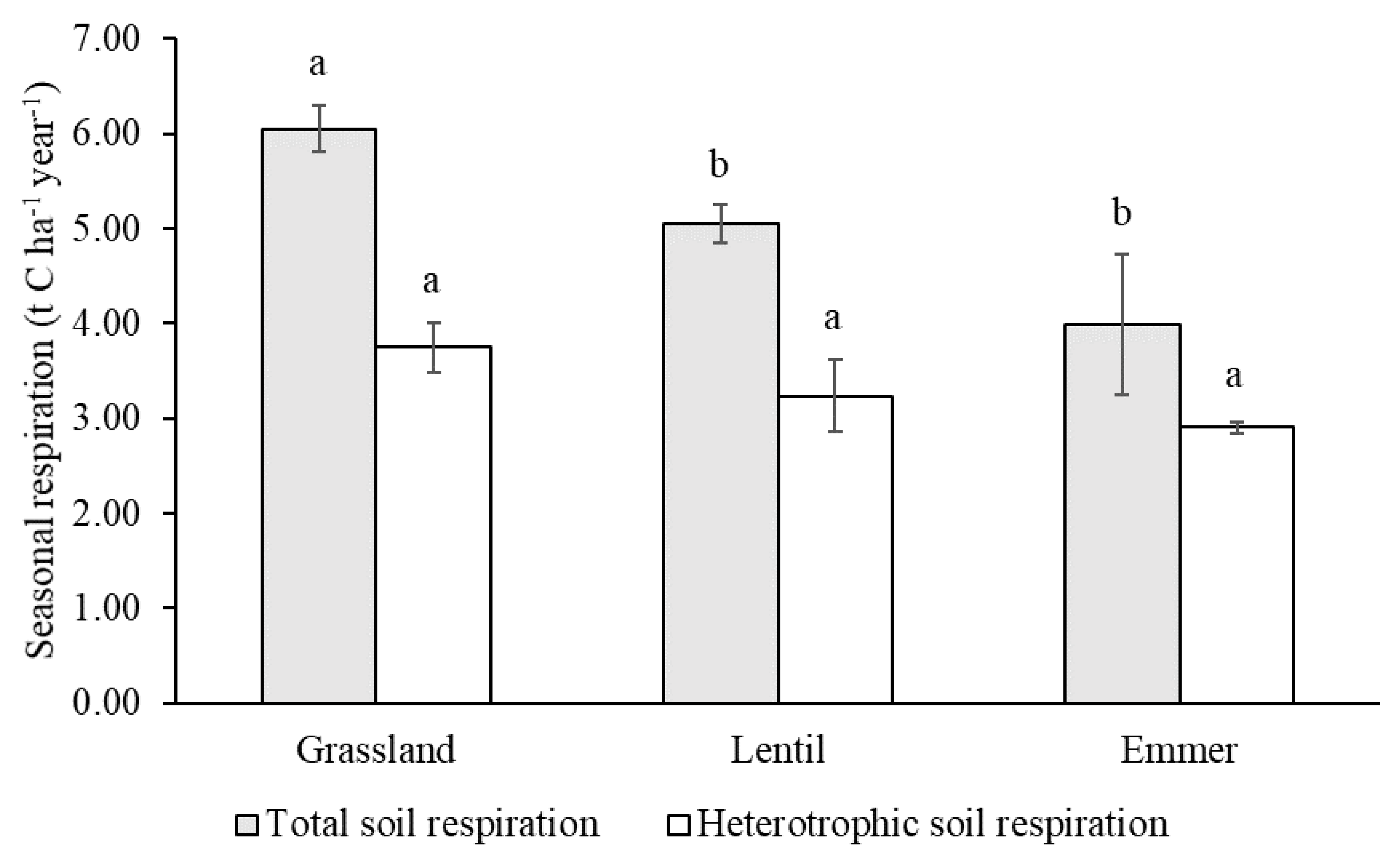
3.3. Seasonal Cumulative Total Respiration and Heterotrophic Respiration Rates
3.4. Soil Carbon Stock
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- D’Ottavio, P.; Francioni, M.; Trozzo, L.; Sedić, E.; Budimir, K.; Avanzolini, P.; Trombetta, M.F.; Porqueddu, C.; Santilocchi, R.; Toderi, M. Trends and approaches in the analysis of ecosystem services provided by grazing systems: A review. Grass Forage Sci. 2018, 73, 15–25. [Google Scholar] [CrossRef]
- Falkowski, P.; Scholes, R.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Högberg, P.; Linder, S.; et al. The Global Carbon Cycle A Test of Our Knowledge of Earth as a System. Science 2000, 290, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Mcgonigle, T.P.; Turner, W.G. Grasslands and Croplands Have Different Microbial Biomass Carbon Levels per Unit of Soil Organic Carbon. Agriculture 2017, 7, 57. [Google Scholar] [CrossRef]
- Selecky, T.; Bellingrath-Kimura, S.; Kobata, Y.; Yamada, M.; Guerrini, I.; Umemura, H.; Dos Santos, D. Changes in Carbon Cycling during Development of Successional Agroforestry. Agriculture 2017, 7, 25. [Google Scholar] [CrossRef]
- Houghton, R.A.; House, J.I.; Pongratz, J.; Van Der Werf, G.R.; Defries, R.S.; Hansen, M.C.; Le Quéré, C.; Ramankutty, N. Carbon emissions from land use and land-cover change. Biogeosciences 2012, 9, 5125–5142. [Google Scholar] [CrossRef]
- Caballero, R.; Molle, G.; Roggero, P.P.; Bagella, S.; D’Ottavio, P.; Papanastasis, V.P.; Fotiadis, G.; Sidiropoulou, A. Grazing systems and biodiversity in Mediterranean areas: Spain, Italy and Greece. Pastos 2009, 39, 9–154. [Google Scholar]
- Wang, W.; Fang, J. Soil respiration and human effects on global grasslands. Glob. Planet. Chang. 2009, 67, 20–28. [Google Scholar] [CrossRef]
- Xu, L.; Baldocchi, D.D. Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California. Agric. For. Meteorol. 2004, 123, 79–96. [Google Scholar] [CrossRef]
- Hoffmann, I.; From, T.; Boerma, D. Ecosystem services provided by livestock species and breeds, with special consideration to the contributions of small-scale livestock keepers and pastoralists. FAO Commission on genetic resources for food and agriculture. Background study paper 66(1). 2014. Available online: http://www.fao.org/3/a-at598e.pdf (accessed on 27 July 2019).
- Monaci, E.; Polverigiani, S.; Neri, D.; Bianchelli, M.; Santilocchi, R.; Toderi, M.; Ottavio, P.D.; Vischetti, C. Effect of contrasting crop rotation systems on soil chemical and biochemical properties and plant root growth in organic farming: First results. Ital. J. Agron. 2017, 12, 364–374. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating Root and Soil Microbial Contributions to Soil Respiration: A Review of Methods and Observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Francioni, M.; Lai, R.; D’Ottavio, P.; Trozzo, L.; Kishimoto-Mo, A.W.; Budimir, K.; Baldoni, N.; Toderi, M. Soil respiration dynamics in forage-based and cereal-based cropping systems in central Italy. Sci. Agric. 2020, 77, 1–15. [Google Scholar] [CrossRef]
- Rong, Y.; Ma, L.; Johnson, D.A.; Yuan, F. Soil respiration patterns for four major land-use types of the agro-pastoral region of northern China. Agric. Ecosyst. Environ. 2015, 213, 142–150. [Google Scholar] [CrossRef]
- Mukumbuta, I.; Shimizu, M.; Hatano, R. Short-term land-use change from grassland to cornfield increases soil organic carbon and reduces total soil respiration. Soil Tillage Res. 2019, 186, 1–10. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. Updated world map of the Köppen-Geiger climate classification. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Budimir, K.; Trombetta, M.F.; Francioni, M.; Toderi, M.; D’Ottavio, P. Slaughter performance and carcass and meat quality of Bergamasca light lambs according to slaughter age. Small Rumin. Res. 2018, 164, 1–7. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to soil taxonomy. Soil Conserv. Serv. 2014, 12, 410. [Google Scholar]
- Paetz, A.; Wilke, B.M. Soil Sampling and Storage. Monit. Assess. Soil Bioremediat. 2005, 5, 1–45. [Google Scholar]
- Alberti, G.; Vedove, G.D.; Zuliani, M.; Peressotti, A.; Castaldi, S.; Zerbi, G. Changes in CO2 emissions after crop conversion from continuous maize to alfalfa. Agric. Ecosyst. Environ. 2010, 136, 139–147. [Google Scholar] [CrossRef]
- González-Ubierna, S.; Lai, R. Modelling the effects of climate factors on soil respiration across Mediterranean ecosystems. J. Arid Environ. 2019, 165, 46–54. [Google Scholar] [CrossRef]
- Fan, L.C.; Yang, M.Z.; Han, W.Y. Soil Respiration under Different Land Uses in Eastern China. PLoS ONE 2015, 10, e0124198. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.; Pegoraro, E.; Tedeschi, V.; De Parri, I.; Jarvis, P.G.; Valentini, R. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob. Chang Biol. 2002, 8, 851–866. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A.; Lou, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Chang. Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Yuste, J.C.; Janssens, I.A.; Carrara, A.; Meiresonne, L.; Ceulemans, R. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest. Tree Physiol. 2003, 23, 1263–1270. [Google Scholar] [CrossRef]
- Almagro, M.; López, J.; Querejeta, J.I.; Martínez-Mena, M. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biol. Biochem. 2009, 41, 594–605. [Google Scholar] [CrossRef]
- Bahn, M.; Rodeghiero, M.; Anderson-Dunn, M.; Dore, S.; Gimeno, C.; Drösler, M.; Williams, M.; Ammann, C.; Berninger, F.; Flechard, C.; et al. Soil respiration in European grasslands in relation to climate and assimilate supply. Ecosystems 2008, 11, 1352–1367. [Google Scholar] [CrossRef]
- Mancinelli, R.; Campiglia, E.; Di Tizio, A.; Marinari, S. Soil carbon dioxide emission and carbon content as affected by conventional and organic cropping systems in Mediterranean environment. Appl. Soil Ecol. 2010, 46, 64–72. [Google Scholar] [CrossRef]
- Raich, J.W.; Tufekcioglu, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Zhu, Y.; Pei, J.; Xie, C.; Yu, Y.; Ye, J.; Fang, C.; Yuan, Z.; Gong, Y. Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa-pasture of the Loess Plateau, China. Sci. Total Environ. 2017, 590, 729–738. [Google Scholar]
- Bao, X.; Zhu, X.; Chang, X.; Wang, S.; Xu, B.; Luo, C.; Zhang, Z.; Wang, Q.; Rui, Y.; Cui, X. Effects of soil temperature and moisture on soil respiration on the Tibetan plateau. PLoS ONE 2016, 11, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Zechmeister-Boltenstern, S.; Jandl, R. Carbon losses due to soil warming: Do autotrophic and heterotrophic soil respiration respond equally? Glob. Chang. Biol. 2009, 15, 901–913. [Google Scholar] [CrossRef]
- Smith, P. Land use change and soil organic carbon dynamics. Nutr. Cycl. Agroecosyst. 2008, 81, 169–178. [Google Scholar] [CrossRef]
- Agnoletti, M. Italian Historical Rural Landscapes. Cultural Values for the Environment and Rural Development; Agnoletti, M., Ed.; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Boix-Fayos, C.; De Vente, J.; Albaladejo, J.; Martínez-Mena, M. Soil carbon erosion and stock as affected by land use changes at the catchment scale in Mediterranean ecosystems. Agric. Ecosyst. Environ. 2009, 133, 75–85. [Google Scholar] [CrossRef]
- Nadeu, E.; Van Oost, K.; Boix-Fayos, C.; De Vente, J. Importance of land use patterns for erosion-induced carbon fluxes in a Mediterranean catchment. Agric. Ecosyst. Environ. 2014, 189, 181–189. [Google Scholar] [CrossRef]
- Drewnik, M.; Musielok, Ł.; Stolarczyk, M.; Mitka, J.; Gus, M. Effects of exposure and vegetation type on organic matter stock in the soils of subalpine meadows in the Eastern Carpathians. Catena 2016, 147, 167–176. [Google Scholar] [CrossRef]
- Novelli, L.E.; Caviglia, O.P.; Piñeiro, G. Increased cropping intensity improves crop residue inputs to the soil and aggregate-associated soil organic carbon stocks. Soil Tillage Res. 2017, 165, 128–136. [Google Scholar] [CrossRef]
- Toderi, M.; Francioni, M.; Seddaiu, G.; Roggero, P.P.; Trozzo, L.; D’Ottavio, P. Bottom-up design process of agri-environmental measures at a landscape scale: Evidence from case studies on biodiversity conservation and water protection. Land Use Policy 2017, 68, 295–305. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Cultivation | pH | Sand (%) | Silt (%) | Clay ( %) | Gravel (%) | Soil Bulk Density (g cm−3) | Wilting Point (%) | Field Capacity (%) | Total Organic C (%) |
|---|---|---|---|---|---|---|---|---|---|
| Grassland | 7.6 ± 0.13 | 60.83 ± 1.36 | 35.70 ± 2.65 | 3.47 ± 1.31 | 65.33 ± 5.03 | 0.92 ± 0.06 | 45.30 ± 4.23 | 56.16 ± 6.53 | 14.68 ± 0.77 |
| Lentil | 8.0 ± 0.01 | 43.37 ± 2.08 | 45.53 ± 1.42 | 11.10 ± 2.72 | 72.33 ± 4.04 | 1.10 ± 0.11 | 24.64 ± 1.96 | 37.21 ± 3.61 | 6.80 ± 1.27 |
| Emmer | 7.9 ± 0.04 | 48.03 ± 1.00 | 44.87 ± 0.46 | 7.10 ± 0.62 | 67.67 ± 2.52 | 0.80 ± 0.10 | 33.39 ± 1.78 | 44.64 ± 12.9 | 10.40 ± 1.38 |
| Experimental Field | Soil Respiration | a | b | R10 | Q10 |
|---|---|---|---|---|---|
| Grassland | Total | 0.10 | 0.08 | 0.22 | 2.23 |
| Heterotrophic | 0.06 | 0.09 | 0.15 | 2.46 | |
| Lentil | Total | 0.02 | 0.15 | 0.09 | 4.48 |
| Heterotrophic | 0.07 | 0.06 | 0.13 | 1.82 | |
| Emmer | Total | 0.01 | 0.18 | 0.06 | 6.05 |
| Heterotrophic | 0.02 | 0.14 | 0.08 | 4.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francioni, M.; D’Ottavio, P.; Lai, R.; Trozzo, L.; Budimir, K.; Foresi, L.; Kishimoto-Mo, A.W.; Baldoni, N.; Allegrezza, M.; Tesei, G.; et al. Seasonal Soil Respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands. Agriculture 2019, 9, 165. https://doi.org/10.3390/agriculture9080165
Francioni M, D’Ottavio P, Lai R, Trozzo L, Budimir K, Foresi L, Kishimoto-Mo AW, Baldoni N, Allegrezza M, Tesei G, et al. Seasonal Soil Respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands. Agriculture. 2019; 9(8):165. https://doi.org/10.3390/agriculture9080165
Chicago/Turabian StyleFrancioni, Matteo, Paride D’Ottavio, Roberto Lai, Laura Trozzo, Katarina Budimir, Lucia Foresi, Ayaka Wenhong Kishimoto-Mo, Nora Baldoni, Marina Allegrezza, Giulio Tesei, and et al. 2019. "Seasonal Soil Respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands" Agriculture 9, no. 8: 165. https://doi.org/10.3390/agriculture9080165
APA StyleFrancioni, M., D’Ottavio, P., Lai, R., Trozzo, L., Budimir, K., Foresi, L., Kishimoto-Mo, A. W., Baldoni, N., Allegrezza, M., Tesei, G., & Toderi, M. (2019). Seasonal Soil Respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands. Agriculture, 9(8), 165. https://doi.org/10.3390/agriculture9080165