Abstract
This study explored the biodiversity of Fucales along the islands of Lipari and Vulcano, almost ten years after the last studies on the phytobenthos of these islands. To carry out this research, a monitoring activity was conducted through both scuba dives and snorkelling activities. Through this study, a total of 13 species (four belonging to Cystoseira, three to Ericaria, two to Gongolaria, and four to Sargassum) was observed. When comparing the present data with past studies, nine species were previously reported, ten taxa were not found in the present study, and four taxa were reported here for the first time in the Aeolian Archipelago. The alpha (local) diversity was higher in Vulcano than in Lipari. Moreover, the beta (turnover) diversity showed a different assemblage in species grouping between the two islands. Since some of the observed species are currently considered rare or in regression, their finding in the Aeolian Archipelago is of particular interest. Therefore, they should receive special consideration, particularly in light of the possible future establishment of a marine protected area (MPA) in the Aeolian Archipelago.
1. Introduction
The Aeolian islands form an archipelago of seven large-sized emerged volcanos (Alicudi, Filicudi, Salina, Lipari, Vulcano, Panarea and Stromboli) and five islets (Basiluzzo, Dattilo, Lisca Nera, Bottaro and Lisca Bianca). They extend off the northern coast of Sicily, in the southeastern Tyrrhenian Sea [1]. These islands were formed about 1.3 million years ago, during the Quaternary Period, and the volcanic activity of the islands of Vulcano and Stromboli continues today [2]. As a result of their volcanic activity, there are many hydrothermal vents along this area, especially around Vulcano and Panarea [3]. These phenomena mainly consist of gaseous emissions of H2S, SO2, SO3 and CO2 at temperatures between 20° and 100 °C and beyond. In particular, the sulphurous emissions not only increase the seawater temperature, but also make it more turbid, due to the presence of the sulphur that is deposited all over the surrounding substrate and benthic organisms. In the areas close to the vents, the pH can vary from 4.5 to 7.4 [4].
The Aeolian Archipelago is characterised by a heterogeneous morphology, with hard bottoms and steep slopes, making the marine environment of these islands a zone of high biodiversity, hosting a wide array of habitats and communities that reach a large depth range [1].
In 1982, this archipelago was formally identified and proposed as a potential marine protected area (MPA), through the Italian law 979/1982 [1,5]. In 2016, the Italian Government committed to establishing an MPA in the Aeolian Archipelago and started the official process to create it. This process can be achieved through an in-depth knowledge of the marine life along these islands. Regarding the marine flora and vegetation of the Aeolian Archipelago, many studies were realised around the 1990s and the first decade of the 2000s.
In particular, historical data are available for the macroalgal flora. Indeed, previous studies were performed by Cavaliere [6,7,8], who mainly focused on the area of the Strait of Messina. Subsequently, other studies on the macroalgal flora were reported by Giaccone [9], Cormaci et al. [10,11], Alongi et al. [12], Acunto et al. [13] and Catra et al. [14] for the islands of Filicudi, Lipari, Panarea, Stromboli, Salina and Vulcano. Finally, Giaccone et al. [5] realised a comprehensive study regarding the macroalgal flora of the Aeolian Archipelago, reporting all the data collected from the previous studies.
Regarding the phytobenthic stands associated with hydrothermal vents, some studies were performed around the stretches of the coastline of Vulcano, in Panarea and on the small island of Bottaro [4,15,16,17,18]. The last study on the flora on the Aeolian Archipelago was performed by Bellissimo et al. [19], who described the community of Ericaria brachycarpa (J. Agardh) Molinari & Guiry (reported as Cystoseira brachycarpa J. Agardh) in an area affected by hydrothermal vents off the eastern coast of Panarea.
Although, in the past, several studies were carried out along the Aeolian Archipelago, there is still a lack of knowledge regarding the perennial macroalgae in these areas [13], particularly those belonging to the genera Cystoseira C. Agardh, Ericaria Stackhouse, Gongolaria Boehmer and Sargassum C. Agardh (order Fucales), which are generally very sensible to anthropogenic effects [20,21,22,23], making them useful indicators of water quality, according to the Water Framework Directive (2000/60/EC) and the Marine Strategy Framework Directive (MSFD, 2008/56/EC).
Although these species are considered among the most important habitat-forming species in the Mediterranean waters, providing very productive, complex and diverse habitats at different depths [23], they are currently considered quite rare due to the dramatic losses they have experienced in the last decades [24,25]. Thus, the current boundaries and distribution ranges for many taxa are, in most cases, still unknown [26].
Therefore, the aims of this work are: (1) to gain up-to-date knowledge of the Fucales on Lipari and Vulcano, which could be used to improve the management of the future MPA; (2) to compare the current data with those extracted by previous studies; (3) to evaluate alpha and beta diversity, comparing the study areas of these two islands; (4) to compare the data of these islands with those obtained from the previous study of the Pantelleria island [27].
2. Materials and Methods
To carry out this research, a monitoring activity was conducted during the first week of July 2023 along the coasts of Lipari and Vulcano (Figure 1). Eleven sites (seven in Lipari and four in Vulcano) were monitored through both scuba dives and snorkelling activities. The sites were selected with the staff from the diving center in accordance with the sea-weather conditions. In particular, a total of 10 scuba dives and 4 snorkelling activities were carried out, each lasting from 40 min to 1 h, covering an area of approximately 250–300 m2 (Table 1).
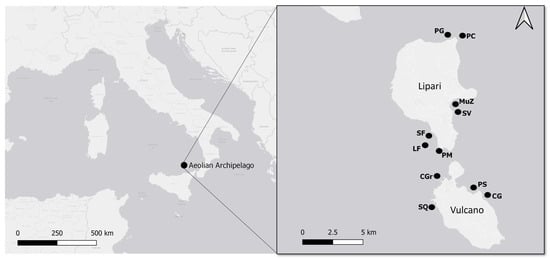
Figure 1.
Geographical locations of the study areas in the Mediterranean Sea. The black dots indicate the investigated sites (see Table 1) on the islands Lipari and Vulcano.

Table 1.
Details of study sites, including their abbreviations, GPS coordinates, dates and survey activities conducted during this study.
A visual census activity was performed through random courses [28]. This technique was selected because it makes it possible to explore different environments and depths [29,30]. The scuba dives were carried out perpendicular to the coastline, following the seabed geomorphology from the surface to about 40 m, depending on the site. The snorkelling activities were conducted along the coastline following the same range of depths (from 0.5 to 1.5 m of depth). The snorkelling sessions were performed in order to observe the environments closer to the coast and, thus, obtain a complete overview of the biodiversity of Fucales along the islands.
During the courses, Fucalean species were identified in situ and photographed using two underwater cameras, Olympus TG-6 and TG-4 (Olympus Corporation, Tokyo, Japan). This approach was useful to minimize the sampling effort on these threatened species.
For each species, photographs were taken to estimate the canopy density according to the 4 classes proposed by Thibaut et al. [31]: absence of individuals, scattered individuals, abundant patches, almost-continuous-to-continuous stands.
Data on the depth and seawater temperature were taken through a Suunto D6i dive computer. In order to be able to identify each taxon, photographs were taken of the frond, base, apex, receptacles and tophules, if present. When species examination was not possible in the field, a few fragments of thalli were taken and stored in exsiccata for subsequent examination with the stereomicroscope. When receptacles were collected, transverse sections of conceptacles were made through a razor blade in order to evaluate the presence of antheridia and oogonia and, thus, assess the maturity of receptacles. The stored material was deposited in the private collection of G.M. Some fragments were taken for future molecular analyses. Species identification was performed following Gómez-Garreta et al. [32], Cormaci et al. [33], Rodríguez-Prieto et al. [34] and Blanfuné et al. [35].
To compare the current presence of the Fucales along these islands with the historical data, the following reference research was consulted: Giaccone [4,5,9,15], Cormaci et al. [10,11], Alongi et al. [12], Acunto et al. [13,16], Catra et al. [14], Acunto and Rindi [17], Bellissimo et al. [18,19].
Alpha diversity was evaluated using species richness, defined as the total number of species identified at each site. However, since this measure is strongly dependent on sampling size and effort, the point diversity was also evaluated through Simpson index, Shannon index and Margalef index [36,37,38]. These indices were used because Shannon index is a statistic index, which depends on species richness, Simpson index is a dominance index, which gives more weight to common or dominant species [39] and, finally, Margalef index is a species-richness index, which compensates for sampling effects and can be used in conjunction with indices sensitive to dominance [40].
Diversity indices were calculated using the software Past 4.03.
Beta diversity among sites on the two islands was estimated using Jaccard similarity matrix constructed on presence/absence data, through the software Primer 7. The patterns were visualised using non-metric multidimensional scaling (non-metric MDS) inferred from Jaccard similarity matrix.
3. Results
3.1. Previous and Current Distributions of Fucales
During the monitoring activity, a total of 13 species were found: four belonging to Cystoseira (C. compressa, C. foeniculacea, C. pustulata and C. cf. micheleae), three to Ericaria (E. dubia, E. brachycarpa, E. funkii), two to Gongolaria (G. montagnei var. compressa and G. sauvageauana), and four to Sargassum (S. vulgare, S. hornschuchii, S. furcatum and S. trichocarpum).
According to the available research, 19 Fucalean species were previously observed along the coasts of these islands (Table 2). Of these, nine species were observed in the present study: E. brachycarpa, C. compressa, C. pustulata, E. funkii, G. sauvageauana, C. foeniculacea, G. montagnei var. compressa, S. hornschuchii and S. vulgare. In addition, ten species were not observed (C. humilis var. myriophylloides, E. amentacea, G. barbata, E. crinita, G. montagnei and G. montagnei var. tenuior, E. selaginoides, E. zosteroides, S. acinarium, S. flavifolium) and four species are here reported for the first time for the Aeolian Archipelago (E. dubia, C. micheleae, S. furcatum and S. trichocarpum).
Table 3 provides information regarding the explored sites (range of depths and seawater temperatures), the distribution of the fucalean taxa found at these sites, the ranges of the depths at which they were documented and the presence of mature receptacles.
Table 2.
Comparison of previous and current records of Fucales on the islands of Lipari and Vulcano.
Table 2.
Comparison of previous and current records of Fucales on the islands of Lipari and Vulcano.
| Species | This Study | Past | Reference |
|---|---|---|---|
| Cystoseira compressa (Esper) Gerloff & Nizamuddin | x | x | [5] |
| Cystoseira foeniculacea (Linnaeus) Greville | x | x | as Cystoseira discors [4] |
| Cystoseira humilis var. myriophylloides (Sauvageau) J. H. Price & D. M. John | x | [5] | |
| Cystoseira cf. micheleae Verlaque, Blanfuné, Boudouresque, Thibaut & Sellam | x | ||
| Cystoseira pustulata (Ercegovic) Neiva & Serrão | x | x | as Cystoseira fimbriata v. pustulata [9] |
| Ericaria amentacea (C. Agardh) Molinari & Guiry | x | as Cystoseira amentacea v. stricta [5] | |
| Ericaria crinita (Duby) Molinari & Guiry | x | as Cystoseira crinita [5] | |
| Ericaria dubia (Valiante) Neiva & Serrão | x | ||
| Ericaria selaginoides (Linnaeus) Molinari & Guiry | x | as Cystoseira tamariscifolia [5] | |
| Ericaria zosteroides (C. Agardh) Molinari & Guiry | x | as Cystoseira zosteroides [5] | |
| Ericaria brachycarpa (J. Agardh) Molinari & Guiry | x | x | as Cystoseira brachycarpa v. brachycarpa [5] |
| Ericaria funkii (Gerloff & Nizamuddin) Molinari & Guiry | x | x | as Cystoseira jabukae [5] |
| Gongolaria barbata (Stackhouse) Kuntze | x | as Cystoseira barbata v. barbata [5] | |
| Gongolaria montagnei (J. Agardh) Kuntze | x | as Cystoseira spinosa v. spinosa [5] | |
| Gongolaria montagnei var. tenuior (Ercegović) Molinari & Guiry | x | as Cystoseira spinosa v. tenuior [5] | |
| Gongolaria montagnei var. compressa (Ercegović) Verlaque, Blanfuné, Boudouresque & Thibaut | x | x | as Cystoseira spinosa v. compressa [5] |
| Gongolaria sauvageauana (Hamel) Molinari & Guiry | x | x | as Cystoseira sauvageauana [5] |
| Sargassum acinarium (Linnaeus) Setchell | x | [5] | |
| Sargassum flavifolium Kützing | x | [5] | |
| Sargassum vulgare C. Agardh | x | x | [5] |
| Sargassum hornschuchii C. Agardh | x | x | [5] |
| Sargassum furcatum Kützing | x | ||
| Sargassum trichocarpum J. Agardh | x |
Table 3.
Distribution of the reported taxa at the sites (see Table 1), with information on the finding-depth range (in meters), temperature range (°C) and, as noted with the asterisk, the presence of receptacles.
Table 3.
Distribution of the reported taxa at the sites (see Table 1), with information on the finding-depth range (in meters), temperature range (°C) and, as noted with the asterisk, the presence of receptacles.
| Lipari | Vulcano | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PC | SV | PG | PM | SF | LF | MuZ | CG | PS | SQ | CGr | |
| Depth range (m) | 0–33 | 0–26 | 0–34 | 0–28 | 0–22 | 0–31 | 0–0.5 | 0–29 | 0–22 | 0–37 | 0–33 |
| Temperature range (°C) | 25°–16° | 25°–19° | 25°–18° | 25°–17° | 25°–18° | 25°–19° | 25° | 26°–17° | 26°–16° | 25°–16° | 26°–17° |
| Taxa | |||||||||||
| Cystoseira compressa | 0.5 | 8 * | 0–0.5 * | 0–0.5 | 0.5–1 | 0.5–1 | 7–10 | 0–5 * | |||
| Cystoseira foeniculacea | 5–29 | 8–19 | 12 | 13 | 7–21 | 11–13 | |||||
| Cystoseira cf. micheleae | 6–8 | 8–14 | |||||||||
| Cystoseira pustulata | 3–5 * | 4–9 * | |||||||||
| Ericaria dubia | 31 | 27 | 30 | ||||||||
| Ericaria brachycarpa | 5–10 | 3–6 | 8–13 | 0.5–15 | 12–17 | 0.2–15 | 0.8–1 | 7 | 0.5–13 | ||
| Ericaria funkii | 27 * | 16–26 * | 15–27 | 18–21 * | 17–22 | 23 * | 16–31 * | 24–31 | |||
| Gongolaria montagnei var. compressa | 28 | 6–28 | 7–20 | ||||||||
| Gongolaria sauvageauana | 16–23 | 20–27 | 17–19 | 9–16 | 11 * | 7–20 * | |||||
| Sargassum vulgare | 0.5–1 | 12–15 * | 15 | 2–9 * | |||||||
| Sargassum hornschuchii | 7–17 * | ||||||||||
| Sargassum furcatum | 12–28 * | 18–26 | 21 | 21 | 10–18 * | 8–12 | 12–24 | ||||
| Sargassum trichocarpum | 15–26 * | 17 | 15–21 * | ||||||||
3.2. Alpha and Beta Diversity
The average number of species per site was 5.64 ± 0.83, with values of 4.28 ± 0.89 at Lipari and 8 ± 0.71 at Vulcano. A total number of 30 species was found at Lipari and 32 at Vulcano.
The Simpson index (Table 4) ranged from a value of 1 (representing the lowest diversity), found at the sites of Secca del Villaggio (SV) and Zona Municipio (MuZ) (Lipari), to a value of 0.1 (corresponding to the highest diversity) in the site of Capo Grillo (CG) (Vulcano). By contrast, the Shannon and Margalef indices showed the highest values (representing the highest levels of diversity) on the island of Vulcano, particularly at Capo Grillo (CG), Parete della Sirena (PS) and Capo Grosso (CGr). In addition, on the island of Lipari, the highest values were observed at Punta Castagna (PC), Le Formiche (LF) and Pietra Menalda (PM), respectively (Table 4).
Table 4.
Simpson, Shannon and Margalef diversity indices calculated for the different examined sites.
Regarding the beta diversity among the sites, a different pattern was observed in the species’ grouping between the two islands (Figure 2). The Jaccard similarity matrix constructed based on the presence/absence of species displayed that the most distant sites were Parete della Sirena (PS) at Vulcano and Zona Municipio (MuZ) and Secca del Villaggio (SV) at Lipari.
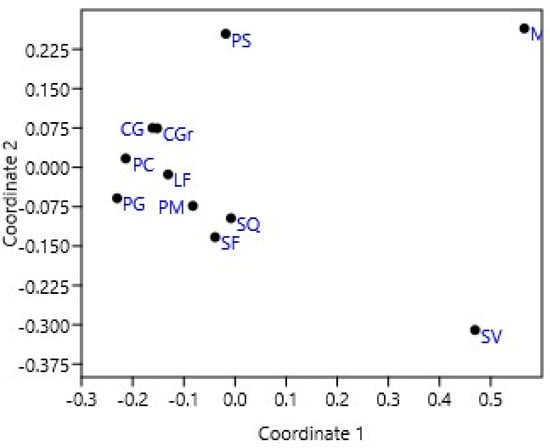
Figure 2.
Non-metric multidimensional scaling ordination based on Jaccard similarity calculated using presence/absence data.
The sites with the highest similarity (70%) were Punta Castagna (PC) and Capo Grillo (CG). A similarity of 66.7% was observed between the sites of Pietra Menalda (PM) and Secca delle Formiche (SF), Le Formiche (LF) and Capo Grosso (CGr) and Secca delle Formiche and Scoglio del Quaglietto (SQ). Finally, the sites of Capo Grillo (CG) and Capo Grosso (CGr) showed a similarity of 63.6%.
3.3. List of the Found Species
For each found species, details on the taxonomic information, morphological observations, habitat and distribution, as well as further remarks, are given. The validity of the species names was checked by consulting Algaebase [41].
- Cystoseira compressa (Esper) Gerloff & Nizamuddin (Figure 3A–E)
 Figure 3. Morphology of Cystoseira compressa specimens from Aeolian islands. (A) A specimen from deep waters; (B) a shallower specimen with receptacles; (C) specimens from exposed sites; (D,E) specimens from the infralittoral fringe, resembling Ericaria amentacea.
Figure 3. Morphology of Cystoseira compressa specimens from Aeolian islands. (A) A specimen from deep waters; (B) a shallower specimen with receptacles; (C) specimens from exposed sites; (D,E) specimens from the infralittoral fringe, resembling Ericaria amentacea.
- Basionym: Fucus compressus Esper, 1799.
- Synonyms: Cystoseira flicina Bory, Cystoseira abrotanifolia f. fimbriata Sauvageau, Fucus fimbriatus Desfontaines, 1799, Cystoseira fimbriata Bory, 1832.
- Morphological observations of Aeolian specimens: The thalli of this species are caespitose, almost completely flattened, without spiniform appendages and with a small discoid holdfast. The thalli are 4–12 cm in length. The apices are small, smooth and non-prominent. The primary branches are flattened and have with an alternating and distichous disposal. The higher-order branches are flattened or cylindrical, always with an alternate–distich arrangement. They feature pedicellate receptacles, which are lanceolate–fusiform and can be simple or branched.
- Habitat: This species shows great adaptability to different habitats and environments. During the monitoring activities, it was observed from the surface to a depth of 10 m, in both sheltered and exposed waters. In deeper waters, only scattered individuals were observed, while in shallower waters, almost continuous stands were observed.
- Distribution: This species was found in eight of the eleven explored sites, on both Lipari and Vulcano (Table 3).
- Further remarks: It was observed that this species has colonised the infralittoral fringe, replacing E. amentacea and E. selaginoides, which were previously documented at these islands (see Table 2). As already demonstrated by Mangialajo et al. [42], the reasons for this substitution could be related to a high level of pollution of the surface waters, due to the continuous passage and discharge of ships, particularly during summer. This replacement probably led to an alteration or adaptation in morphological form, characterised by long fronds with a three-dimensional, pyramidal shape, similar to those of E. amentacea, capable of withstanding a high degree of wave motion and prolonged air exposure. This hypothesis was also suggested by Mangialajo et al. [42], who observed that the lengths of the axes of C. compressa increased towards the urbanised shores as a result of their release from competition with E. amentacea. However, Mangialajo et al. [42] did not exclude the potential effects of other environmental factors.
- Cystoseira foeniculacea (Linnaeus) Greville (Figure 4A–C)
 Figure 4. Morphology of Cystoseira foeniculacea specimens from Aeolian islands. (A) Habit; (B) spinose and prominent apex; (C) a specimen growing on a pebble.
Figure 4. Morphology of Cystoseira foeniculacea specimens from Aeolian islands. (A) Habit; (B) spinose and prominent apex; (C) a specimen growing on a pebble.
- Basionym: Fucus foeniculaceus Linnaeus
- Synonyms: Cystoseira abrotanifolia (Linnaeus) C. Agardh, C. concatenata (Linnaeus) C. Agardh, C. discors (Linnaeus) C. Agardh, C. ercegovicii Giaccone, Fucus abrotanifolius Linnaeus, F. barbatus Linnaeus, F. concatenatus Linnaeus, F. discors Linnaeus, Phyllacantha concatenata (Linnaeus) Kützing.
- Morphological observations of Aeolian specimens: This species is caespitose, with several axes entirely covered by small spiniform appendages, which provide a characteristic knotty habit. The thalli are 5–7 cm long. The holdfast is wide and discoid. The apices of this species are spinose and very prominent in relation to the insertions of the primary branches. The latter are cylindrical, covered with small spines and feature secondary branches with a distichous and alternating disposal. The last-order branches are filiform.
- Habitat: C. foeniculacea was observed in the infralittoral, at depths from 5 m to 21 m, as scattered individuals or forming small patches.
- Distribution: C. foeniculacea was found at six (three in Lipari and three in Vulcano) of the eleven studied sites (Table 3).
- Further remarks: This species was often observed growing on small pebbles (5–6 cm) on coarse gravel substrate.
 Figure 5. Morphology of Cystoseira cf. michaelae specimens from Aeolian islands. (A) Foliose habit; (B) detail of the apex surrounded by tophules; (C) detail of an ovoid smooth–tuberculate tophule; (D) a small-sized specimen growing on a pebble.
Figure 5. Morphology of Cystoseira cf. michaelae specimens from Aeolian islands. (A) Foliose habit; (B) detail of the apex surrounded by tophules; (C) detail of an ovoid smooth–tuberculate tophule; (D) a small-sized specimen growing on a pebble.
- Basionym: Cystoseira micheleae Verlaque, Blanfuné, Boudouresque, Thibaut & Sellam.
- Synonyms: Cystoseira granulata var. turneri Montagne.
- Morphological observations of Aeolian specimens: This species has a characteristic foliose habit. It is non-caespitose with an axis originating from a robust discoid base, which can be single or branched. The thalli are 4–8 cm in length. The apex is spinose and not prominent, often surrounded by tophules. The latter are ovoid, from spinose to smooth–tuberculate with ageing. The primary branches are cylindrical at the base and then tend to be flattened with an inconspicuous midrib. They can be alternately branched in one or several planes, with simple or bifid-spine-like appendages. Receptacles were not observed.
- Habitat: This species was observed in the infralittoral, at depths from 6 m to 14 m, forming dense patches. According to Blanfuné et al. [35] and as observed during the monitoring, it prefers sub-horizontal to sloping bottoms, with slopes of up to 45°. We also observed small-sized thalli growing on pebbles.
- Distribution: This species was observed on the island of Vulcano at Capo Grillo and Parete della Sirena (Table 3). The latter site is characterised by gaseous emissions of H2S, SO2, SO3.
- Further remarks: This species was described by Montagne [43], from Algeria, as C. granulata var. turneri Montagne. Between 2014 and 2015, Sellam et al. [44] collected some samples in the regions of Tipaza and Algiers that corresponded to Montagne’s taxon. They elevated the Montagne variety to species status, giving it a new name, Cystoseira micheleae Verlaque (as Cystoseira michaelae Verlaque et al.), due to the existence of C. turneri (Yendo) Roberts, and they designed the lectotype. Furthermore, the authors distinguished C. micheleae from C. montagnei (now Gongolaria montagnei) through the following characteristics: in the former, the tophules are spinose when young and become tuberculate when older, while in the latter, they remain spinose; and in the former, the receptacles are both basal and intercalary near the tophules, and terminal on the branchlets, while in the latter, they are only in the terminal position. To date, this species has only been reported along the coasts of Algeria and Tunisia [35]. However, some of the reports of G. montagnei up until 2017, the date of the publication of the work by Sellam et al. [44], could have been misidentifications referring to this taxon and, therefore, the distribution of C. michelae could be wider.
The specimens observed during this study showed a wide array of tophules, from spinose to tuberculate and rough. Thus, they match the description by Sellam et al. [44]. However, since receptacles were not observed, the identification at species level was not certain. Future molecular analyses could be useful to compare these specimens with the African specimens and to characterise this entity genetically.
- Cystoseira pustulata (Ercegovic) Neiva & Serrão (Figure 6A–E)
 Figure 6. Morphology of Cystoseira pustulata specimens from Aeolian islands. (A) Habit; (B) a fertile specimen; (C) detail of receptacles; (D) a fertile specimen with aerocysts; (E) detail of receptacles sustained by flattened aerocysts.
Figure 6. Morphology of Cystoseira pustulata specimens from Aeolian islands. (A) Habit; (B) a fertile specimen; (C) detail of receptacles; (D) a fertile specimen with aerocysts; (E) detail of receptacles sustained by flattened aerocysts.
- Basionym: Cystoseira abrotanifolia subsp. pustulata Ercegovic.
- Synonym: Cystoseira compressa subsp. pustulata (Ercegovic) Verlaque.
- Morphological observations of Aeolian specimens: This species has a very delicate and minute habit. It is characterised by caespitose thalli that are 7–12 cm in length, which are attached to the substrate by a small discoid holdfast. The apices are smooth, tiny and not prominent. The primary branches are cylindrical-to-compressed, with an alternating and distichous arrangement. They are sprinkled with prominent cryptostomata that provide a pustulate aspect, from which the name “pustulata” derives. The secondary and tertiary branches are cylindrical and slender. They feature tiny, lanceolate and fusiform receptacles, which can be simple or bifurcated and may be sustained by a flattened aerocyst.
- Habitat: Scattered individuals from this species were observed in the infralittoral, at depths from 3 m to 9 m.
- Distribution: C. pustulata was found only at the sites of Pietra Menalda and Parete della Sirena, in Lipari and Vulcano, respectively (Table 3).
- Further remarks: This species was previously reported on the island of Vulcano as C. fimbriata v. pustulata by Giaccone [9]. Subsequently, Giaccone et al. [5] reported this entity as a synonym of C. humilis v. humilis in the list of phytobenthic species of the Aeolian archipelago. Recently, Neiva et al. [26] pointed out that the previous records of C. humilis could be misidentifications and might actually refer to C. pustulata, which has been defined as a separate and independent species.
- Ericaria dubia (Valiante) Neiva & Serrão (Figure 7A–E)
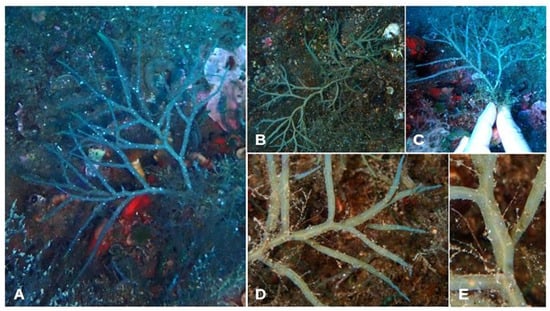 Figure 7. Morphology of Ericaria dubia specimens from Aeolian islands. (A) erect specimen; (B) creeping specimen; (C,D) branching disposal; (E) detail of the midrib.
Figure 7. Morphology of Ericaria dubia specimens from Aeolian islands. (A) erect specimen; (B) creeping specimen; (C,D) branching disposal; (E) detail of the midrib.
- Basionym: Cystoseira dubia Valiante.
- Synonym: Cystoseira fucoides Ercegovic.
- Morphological observations of Aeolian specimens: This species is pseudocaespitose, with an axis that prematurely divides. It can have an erect or more creeping habit. The thalli are 4–7 cm long. The apex is smooth and not prominent. In situ, it shows slight iridescence. The primary branches have a cylindrical basal part and a flattened distal part. The branching disposal is distichous and alternate, in one plane. All the branches have an entire margin and are run throughout by an evident midrib. Receptacles were not observed.
- Habitat: E. dubia was found as scattered individuals or as forming small patches in the lower infralittoral–circalittoral, at depths from 27 m to 31 m.
- Distribution: This species was recorded at three sites: one in Lipari (Punta Castagna) and two in Vulcano (Capo Grillo and Capo Grosso) (Table 3).
- Further remarks: The annual cycle of the development of a E. dubia population was well studied by Tita [45] in Cannizzaro (on the eastern coast of Sicily). Currently, in this area, this species is completely absent (according to our personal observation). As a deep species, E. dubia is particularly threatened by pollution, turbidity, sedimentation from watersheds, trawling and competition with non-indigenous species [35]. Despite its rarity, the status of this species in many Mediterranean areas is still unknown. Therefore, the finding of this species in the Aeolian islands is valuable and needs to be addressed in the context of the MPA-creation process.
- Ericaria brachycarpa (J. Agardh) Molinari & Guiry (Figure 8A–C)
 Figure 8. Morphology of Ericaria brachycarpa specimens from Aeolian islands. (A) Habit; (B) holdfast; (C) apices.
Figure 8. Morphology of Ericaria brachycarpa specimens from Aeolian islands. (A) Habit; (B) holdfast; (C) apices.
- Basionym: Cystoseira brachycarpa J. Agardh
- Synonyms: Cystoseira brachycarpa J. Agardh, Cystoseira caespitosa Sauvageau, Carpodesmia brachycarpa (J. Agardh) Orellana & Sansón
- Morphological observations of Aeolian specimens: This species is caespitose, with several thalli that originate from a wide and irregular holdfast. The thalli are 10–14 cm in length. The axes are rough and knotty due to the fronds sinking to the sea floor during the resting period. The apices are not prominent and they are covered by spinose appendages. The primary branches are cylindrical and can feature spines. The higher-order branches are cylindrical, without spines. Receptacles were not observed.
- Habitat: E. brachycarpa was observed in the infralittoral, at depths from 0.8 m to 17 m, forming well-structured and continuous stands.
- Distribution: This species is widely distributed in Lipari and Vulcano, indeed, it was observed at nine of the eleven examined sites (Table 3).
- Further remarks: Recently, Neiva et al. [26] highlighted the existence of the two cryptic species, E. brachycarpa and E. balearica (Sauvageau) Neiva, Ballesteros & Serrão, which are well genetically differentiated and geographically separated. The E. balearica is distributed in the Balearic Sea and on the Sicilian Island of Pantelleria, while E. brachycarpa is present in Greece and along the northern coast of Sicily. Since one of these Sicilian localities (Capo Milazzo) is near the Aeolian archipelago, it is likely that the specimens found in Lipari and Vulcano belong to the entity E. brachycarpa. Future molecular analyses could confirm this hypothesis.
- Ericaria funkii (Gerloff & Nizamuddin) Molinari & Guiry (Figure 9A–F)
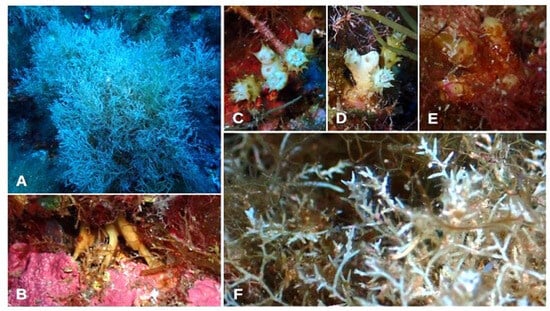 Figure 9. Morphology of Ericaria funkii specimens from Aeolian islands. (A) Habit; (B) digitiform holdfast; (C,D) apices; (E) oblong tophules; (F) receptacles.
Figure 9. Morphology of Ericaria funkii specimens from Aeolian islands. (A) Habit; (B) digitiform holdfast; (C,D) apices; (E) oblong tophules; (F) receptacles.
- Basionym: Cystoseira funkii Schiffner ex Gerloff & Nizamuddin
- Synonym: Carpodesmia funkii (Schiffner ex Gerloff & Nizamuddin) Orellana & Sansón
- Morphological observations of Aeolian specimens: This species is non-caespitose, with an axis that has a coralloid aspect. The thalli are 13–15 cm in length. In situ, this species shows strong iridescence. The thalli are attached to the substrate through haptera or a digitiform holdfast. The apex is smooth and not prominent. The primary branches are cylindrical and spiniform. They feature higher-order branches with an alternate disposal. The tophules are oblong and spinose or rough. They are distributed along the axis, near the base, further contributing to providing the thallus with this coralloid aspect. The receptacles are positioned on the terminal branchlets and are diffuse. Conceptacles are located at the base of the spine.
- Habitat: This species was found in the lower infralittoral–circalittoral, at depths from 15 m to 31 m, forming dense and continuous stands.
- Distribution: This species was found at eight of the eleven explored sites, in both Lipari and Vulcano (Table 3).
- Further remarks: In the past, in the Aeolian archipelago, the presence of Cystoseira jabukae Ercegovic was documented [5]. Some authors considered E. funkii to be a synonym of C. jabukae. However, Verlaque et al. [46], by studying specimens of C. jabukae collected from Corsica and comparing them with the description by Ercegovic [47], concluded that C. jabukae and C. funkii are two distinct species, and considered the previous reports from the western Mediterranean and the Ionian Sea to refer to C. funkii [33]. This C. funkii is distinguished by the following characters: a haptera-like holdfast; a short primary axis that branches out into divaricated and radially arranged secondary axes; spiny tophules spaced along the axis; the lower portion of the primary branches bearing spines; abundant spinose appendages; and receptacles with spinose appendages [46]. Given the aforementioned features, we believe that the specimens observed during this study belong to E. funkii.
Recently, this species was also found along the Cilento coast and was genetically analysed by Rendina et al. [48]. This confirms the presence of this entity in the Tyrrhenian Sea since a long time, demonstrating a stable trend.
- Gongolaria montagnei var. compressa (Ercegović) Verlaque, Blanfuné, Boudouresque & Thibaut (Figure 10A–C)
 Figure 10. Morphology of Gongolaria montagnei var. compressa specimens from Aeolian islands. (A) Habit; (B) arrangement of tophules along the length of the thallus; (C) detail of tophules.
Figure 10. Morphology of Gongolaria montagnei var. compressa specimens from Aeolian islands. (A) Habit; (B) arrangement of tophules along the length of the thallus; (C) detail of tophules.
- Basionym: Cystoseira adriatica subsp. compressa Ercegovic.
- Synonyms: Cystoseira adriatica subsp. compressa Ercegovic, Cystoseira platyramosa Ercegovic, Cystoseira adriatica subsp. intermedia Ercegovic, Cystoseira adriatica var. intermedia (Ercegovic) Giaccone, Cystoseira spinosa var. compressa (Ercegovic) Cormaci, G. Furnari, Giaccone, Scammacca & D. Serio, Cystoseira montagnei var. compressa (Ercegovic) M. Verlaque, Blanfuné, Boudouresque, Thibaut & Sellam.
- Morphological observations of Aeolian specimens: This species is non-caespitose, with an axis that rarely divides, attached to the substrate by a compact discoid holdfast. The thalli are 12–15 cm long. The apex is spinose and not prominent. The primary branches originate from the tophules; they are initially cylindrical and then become flattened with serrated margins and a central midrib. All the branches have an alternating disposal and can feature spiniform appendages. The tophules are distributed along the length of the thallus, especially in its upper part. They are ovoid and covered by spines. Receptacles were not observed.
- Habitat: This species was found in the infralittoral, at depths from 6 m to 28 m, as both scattered individuals and sparse patches.
- Distribution: G. montagnei var. compressa was reported at one site (Punta Castagna) in Lipari and at two sites (Capo Grillo and Parete della Sirena) in Vulcano (Table 3).
- Further remarks: In the literature consulted, it was observed that this species is still present in Lipari and Vulcano (Table 2), demonstrating a quite stable trend.
- Gongolaria sauvageauana (Hamel) Molinari & Guiry (Figure 11A–C)
 Figure 11. Morphology of Gongolaria sauvageauana specimens from Aeolian islands. (A) Habit; (B) branched axis; (C) detail of the spinose and prominent apex.
Figure 11. Morphology of Gongolaria sauvageauana specimens from Aeolian islands. (A) Habit; (B) branched axis; (C) detail of the spinose and prominent apex.
- Basionym: Cystoseira sauvageauana Hamel.
- Synonyms: Cystoseira selaginoides var. polyoedematis Sauvageau, Cystoseira sauvageauana Hamel, Cystoseira sauvageauana var. polyoedematis (Sauvageau) Hamel, Cystoseira sicula Schiffner ex Gerloff & Nizamuddin, Treptacantha sauvageauana (Hamel) Orellana & Sansón.
- Morphological observations of Aeolian specimens: This species is non-caespitose, with an axis that can be simple or branched, attached to the substrate by a discoid holdfast. The thalli are 12–16 cm in length. The apex is spinose and very prominent in relation to the insertion of the primary branches. The primary branches are long and cylindrical, and they are slender in relation to the axis. They can feature spiniform appendages in their lower portions. The higher-order branches are cylindrical and widely spaced, and they decrease in length towards the top. The receptacles are on the terminal branchlets and they are compact and cylindrical, with a few spiniform appendages. Conceptacles can be observed at the base of the spine, when present.
- Habitat: This species was found forming patches or almost continuous stands in the infralittoral, at depths from 9 m to 27 m.
- Distribution: This species was found at six sites: four in Lipari (Punta Castagna, Pietra Menalda, Secca delle Formiche and Le Formiche) and two in Vulcano (Capo Grillo and Scoglio del Quaglietto) (Table 3).
- Further remarks: The decline of G. sauvageauana in several Mediterranean areas has been widely documented [25] and, thus, it can now be considered a rare species. In the Aeolian archipelago, this taxon was reported in the past, and it is still present today. Therefore, the presence of this species on these islands deserves special consideration, particularly in light of the establishment of the MPA.
- Sargassum vulgare C. Agardh (Figure 12A–C)
 Figure 12. Morphology of Sargassum vulgare specimens from Aeolian islands. (A) Habit; (B) lanceolate foliose branches; (C) detail of receptacles.
Figure 12. Morphology of Sargassum vulgare specimens from Aeolian islands. (A) Habit; (B) lanceolate foliose branches; (C) detail of receptacles.
- Basionym: Sargassum vulgare C. Agardh, nom. illeg.
- Synonyms: Fucus salicifolius S. G. Gmelin, Sargassum megalophyllum Montagne, Sargassum coarctatum Kützing, Sargassum vulgare var. megalophyllum (Montagne) Vickers.
- Morphological observations of Aeolian specimens: This species has a single knotty axis, simple or branched, which is attached to the substrate through a discoid holdfast. The thalli are 5–8 cm long. The primary branches are cylindrical and feature secondary branches, which have a distichous–alternating arrangement. The foliose branches are lanceolate with an evident midrib, with toothed or wavy margins. Aerocysts were not observed. The receptacles are composed (with a sterile pedicel) and are fusiform, warty, simple or branched.
- Habitat: Scattered individuals of this species were found from the upper infralittoral (0.5–1 m) to a depth of 15 m.
- Distribution: The S. vulgare was found at four sites: one in Lipari (Le Formiche) and three in Vulcano (Parete della Sirena, Scoglio del Quaglietto and Capo Grosso) (Table 3).
- Further remarks: This species has a stable trend on these islands.
- Sargassum hornschuchii C. Agardh (Figure 13A–D)
 Figure 13. Morphology of Sargassum hornschuchii specimens from Aeolian islands. (A) Habit; (B) foliose branches; (C) receptacles; (D) arrangement of receptacles.
Figure 13. Morphology of Sargassum hornschuchii specimens from Aeolian islands. (A) Habit; (B) foliose branches; (C) receptacles; (D) arrangement of receptacles.
- Basionym: Sargassum hornschuchii C. Agardh.
- Synonym: Stichophora hornschuchii (C. Agardh) Kützing.
- Morphological observations of Aeolian specimens: This species is erect, with a single smooth axis, attached to the substrate by a discoid holdfast. The thalli are 5–8 cm in length. The primary branches are flattened at the base, and sometimes slightly toothed at the margins. The foliose branchlets are narrow, lanceolate with a distichous–alternating disposal. They have a central midrib and entire, smooth or slightly toothed margins. Aerocysts were not observed. The receptacles are warty, flattened or triangular. They are composed, brought by a sterile pedicel, which is well developed and branched.
- Habitat: This species was found in the infralittoral at depths from 7 to 17 m, as scattered individuals or small patches.
- Distribution: The S. hornschuchii was only recorded at the site of Scoglio del Quaglietto, on the island of Vulcano (Table 3).
- Further remarks: Thibaut et al. [24,49] documented the decline of this species in many Mediterranean areas. In the Aeolian Archipelago, this species has been reported in the past [5], and it is still stable there. Thus, it should receive special attention in the process of MPA creation.
- Sargassum furcatum Kützing (Figure 14A–C)
 Figure 14. Morphology of Sargassum furcatum specimens from Aeolian islands. (A) Habit; (B) detail of the foliose branches branched up to 4 times; (C) detail of the receptacles.
Figure 14. Morphology of Sargassum furcatum specimens from Aeolian islands. (A) Habit; (B) detail of the foliose branches branched up to 4 times; (C) detail of the receptacles.
- Basionym: Sargassum furcatum Kützing
- Synonyms: Sargassum vulgare f. furcatum (Kützing) J. Agardh
- Morphological observations of Aeolian specimens: This species has a creeping habit. The thalli, which are 5–8 cm long, are composed of a single axis, attached to the substrate by a discoid holdfast. The primary and secondary branches are knotty and feature foliose branches, which can be lanceolate or lobate, with pointed apexes and smooth or toothed margins, with an evident midrib that follows the bifurcation of the branches. Indeed, this species takes the name “furcatum” because its foliose branches can be divided up to four times. Aerocysts were not observed. The receptacles are cylindrical, lanceolate, warty and branched, forming a bunch. They are usually situated at the axilla of the foliose branches.
- Habitat: Scattered individuals were found in the infralittoral, at depths from 8 m to 28 m.
- Distribution: The S. furcatum was recorded at seven of the eleven examined sites: four in Lipari (Punta Castagna, Parete dei Gabbiani, Punta Menalda and Le Formiche) and three in Vulcano (Capo Grillo, Parete della Sirena and Capo Grosso).
- Further remarks: This species, with an Atlantic and Pacific distribution [41], was first reported in the Mediterranean by Flores-Moya and Conde [50], in Spain. Subsequently, in 2021, it was found in several locations along the eastern coast of Sicily [51]. Through this monitoring activity, S. furcatum was also reported in the Aeolian archipelago, representing a further record of this non-indigenous species in Sicily.
- Sargassum trichocarpum J. Agardh (Figure 15A–C)
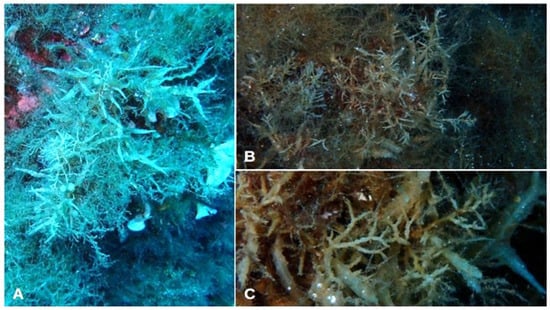 Figure 15. Morphology of Sargassum trichocarpum specimens from Aeolian islands. (A) Habit; (B) arrangement of receptacles; (C) receptacles.
Figure 15. Morphology of Sargassum trichocarpum specimens from Aeolian islands. (A) Habit; (B) arrangement of receptacles; (C) receptacles.
- Basionym: Sargassum trichocarpum J. Agardh.
- Synonyms: Sargassum boryanum Montagne, Sargassum vulgare var. trichocarpum J. Agardh.
- Morphological observations of Aeolian specimens: This species is erect, with a single smooth axis, which is attached to the substrate by a discoid holdfast. The thalli are 7–12 cm in length. The primary branches are cylindrical and smooth. The foliose branches are elongated and narrow, with wavy or serrated margins and a central midrib. The aerocysts are spherical and brought by a slender pedicel. The receptacles are cylindrical, slender and branched several times in one plane.
- Habitat: This species was found in the infralittoral, at depths from 15 m to 26 m, forming small patches on the seabed.
- Distribution: This species was recorded at three sites (Capo Grillo, Parete della Sirena and Capo Grosso) of Vulcano (Table 3).
- Further remarks: In the literature consulted, it was observed that this species was not previously documented in the Aeolian archipelago (Table 2). Thus, the record presented here is the first for these islands. Since, as reported by Thibaut et al. [49] and Ballesteros and Weitzmann [52], S. trichocarpum is a regressing and rare species in the Mediterranean Sea, this record should receive special consideration in the context of the creation of MPAs.
4. Discussion
This study explored the current diversity of Fucales along Lipari and Vulcano, almost ten years after the last studies on the phytobenthos on these islands. Comparing the present data with those obtained in the past, it was observed that 10 taxa were missed: C. humilis var. myriophylloides, E. amentacea, E. crinita, E. selaginoides, E. zosteroides, G. barbata, G. montagnei and G. montagnei var. tenuior, S. acinarium, S. flavifolium. Nevertheless, nine species were still observed on these islands (C. compressa, C. foeniculacea, C. pustulata, E. brachycarpa, E. funkii, G. montagnei var. compressa, G. sauvageauana, S. hornschuchii and S. vulgare) and four taxa (C. cf. micheleae, E. dubia, S. furcatum and S. trichocarpum) are here reported for the first time for the Aeolian Archipelago.
Most of these species are listed as “endangered or threatened” macroalgae in Annex II of the Barcelona Convention [53]. Nevertheless, unfortunately, Cystoseira s.l. species have not yet been included on the IUCN Red List, even though the codes LC (low-concern species), RA (rare species) and CR (critically endangered species) were proposed, according to the species, by Verlaque et al. [53]. On the other hand, S. hornschuchii was considered EN (endangered species), while S. trichocarpum was considered VU (vulnerable species) [53]. Therefore, the finding of these species in the Aeolian Archipelago is of particular interest and should receive attention in the future, especially regarding the possible future creation of an MPA zone.
Regarding C. cf. micheleae, the finding of this species in the Aeolian Archipelago is remarkable, since, to date, the known distribution of this species includes only Algeria and Tunisia [35]. However, this needs further investigation due to the lack of all the features necessary for species identification and the need to characterise this taxon through genetic analyses.
The record of S. furcatum along the coasts of the Aeolian Archipelago is also noteworthy. Considering the provenance of this species, as well as the fact that it was recently found for the first time in widely studied areas in the past, it might be assumed that the species is non-indigenous [51]. The present report on the Aeolian Archipelago could indicate the additional spread of this species in the Tyrrhenian Sea. The finding of fertile individuals both along the Ionian coasts of Sicily and in the Aeolian Archipelago could demonstrate that S. furcatum is establishing reproductive and, possibly, self-sustaining populations in the Mediterranean Sea.
Overall, when comparing the past and current fucalean diversity, it can be noted that in the last ten years, a regression occurred in the taxa of the infralittoral fringe, such as E. amentacea and E. selaginoides (see Table 2), and that they were replaced by C. compressa. Even the upper infralittoral, where, previously, E. crinita and G. barbata (Table 2) were reported, is now mostly colonised by E. brachycarpa stands. This process might be due to biotic homogenisation related to anthropogenic effects [54]. In particular, this replacement could be attributed to an increment in the pollution rate due to the discharge and passage of ships, particularly during summer, as also observed in the city of Genova by Mangialajo et al. [42]. Therefore, it can be assumed that this substitution might have led C. compressa to adopt a pyramidal and three-dimensional form, in order to better adapt to this environment, characterised by high hydrodynamism and air exposure. Mangialajo et al. [42] observed an increase in the thallus length of C. compressa in proximity to the most anthropised areas, as a result of the competition with E. amentacea. Nevertheless, the potential effects of other factors cannot be excluded and, thus, further studies are necessary to better investigate this process.
The evaluation of the alpha diversity in each study area highlighted a generally higher level of diversity at the sites of Vulcano than in Lipari, despite the lower number of areas (four) explored on the former island. This difference could be due to the fact that Lipari is subjected to more anthropogenic effects than Vulcano. One of anthropic activities that profoundly altered the sea bottom was the cumbersome extraction of pumice, whose sandy wastes currently cover much of the seabed of the northwestern sector of Lipari [55]. The estimation of the beta diversity across the study areas showed a difference in the pattern of the species’ grouping between these two islands. Moreover, the same analysis indicates that the most different assemblage was observed at the Parete della Sirena site, on the island of Vulcano. This different pattern might be related to the gaseous emissions of H2S, SO2 and SO present at this site [55].
By comparing the biodiversity of the fucalean species of Lipari and Vulcano with that of Pantelleria [27], it can be noted that there is a lower number of taxa in the Aeolian islands (13) than in Pantelleria (19). This is most likely because the island of Pantelleria is located in the Sicily Channel, where the modified Atlantic water (MAW) from the Atlantic Sea encounters the Levantine intermediate water (LIW) from the eastern Mediterranean [56], favouring the dispersion of species from one side of the Mediterranean to the other [57]. Nevertheless, both in Pantelleria and on the Aeolian Islands, deep-sea species, such as G. montagnei var. compressa and E. dubia, have remained stable over time. This could be due to the action of the cool currents of deep bottoms, which might offer a spatial refuge [58], maintaining well-structured deep fucalean populations [59].
In this study, a large difference in temperature between deeper and shallower waters was observed: at depths from ca. 20 m to 40 m, the temperature range was 16–19 °C, while at depths from 0 m to 10 m, the temperature range was 25–26 °C (Table 3). By comparing the historical data reported by the website of the Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA) [60] for the station of Ginostra (located in the Aeolian Archipelago), an increase in surface-seawater temperature of ca. 2 °C from 2013 to 2023 was observed. This temperature increase might have contributed to the regression of shallow species along the Aeolian Archipelago, as already documented on other surface species, such as E. crinita [61], E. selaginoides [62] and G. barbata [63]. Therefore, considering the foreseen scenario of temperature and thermal anomaly increments [64], it will be necessary to keep monitoring these species and to identify the possible existence of climatic refugia that could guarantee the existence and survival of these Fucales in the future [61].
5. Conclusions
Cystoseira s.l. (comprising the three genera of Cystoseira, Ericaria and Gongolaria) and Sargassum species are ecosystem engineers in Mediterranean rocky coastal environments, forming dense forests and representing one of the most productive Mediterranean marine ecosystems [34,65]. Nevertheless, in the last decades, these species have undergone a severe decline and loss in many coastal Mediterranean areas, due to several anthropogenic stressors, such as coastal urbanisation, marine pollution, eutrophication, sedimentation and competition with invasive species [23,42,66,67,68,69], which are ultimately intensified by the effects of climate change and thermal anomalies [61,63,64,70,71]. Moreover, the knowledge on the distribution and health status of many fucalean taxa and their populations is still scarce or limited [72]. Indeed, in several Mediterranean areas, the studies on these species are quite outdated [27,73] and, thus, it is important to update the current information on these habitat-forming species, in order to protect them and favour their conservation, future survival or ecological restoration. In this study, we reported the current distribution of fucalean stands in the Aeolian Archipelago about ten years after the last studies on these species.
Cystoseira s.l. and Sargassum species are considered “species of community interest” in the European Habitat Directive (92/43/EEC, 1992). Nevertheless, more effective protection regimes, as MPAs, are currently required to halt and limit the decline of these marine Mediterranean forests.
At the international level, the Aeolian archipelago was designated in 2001 as a UNESCO World Heritage site. At the national level, the Italian government identified the Aeolians as a potential MPA, and in 2016, the government publicly committed to the designation of this archipelago as an MPA [1]. However, the official process is still ongoing. Therefore, during the process of the MPA’s establishment, it will be important to consider the current distribution of the fucalean taxa reported here, paying special attention to the species that are currently considered rare or critically endangered.
Author Contributions
Conceptualization, G.M. and A.L.; methodology, G.M. and A.L.; validation, D.S. and S.B.; formal analysis, G.M. and S.B.; writing—original draft preparation, G.M. and A.L.; writing—review and editing, D.S. and S.B.; supervision, D.S. and S.B.; project administration, S.B.; funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was also supported by the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4—Call for tender no. 3138 of 16 December 2021, rectified by Decree no. 3175 of 18 December 2021 by the Italian Ministry of University and Research, funded by the European Union—NextGenerationEU, Award Number: project code CN_00000033, Concession Decree No. 1034, of 17 June 2022, adopted by the Italian Ministry of University and Research, Project title “National Biodiversity Future Center—NBFC”, and the Biodiversa+ FORESCUE (Biodiversa2021-134) project, BiodivProtect call on “Supporting the protection of biodiversity and ecosystems across land and sea”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated during this study are included in this published article.
Acknowledgments
The authors wish to thank all the staff at the diving centre, La Gorgonia, for their willingness, kindness and expertise. Moreover, the authors would like to express their gratitude to the three anonymous reviewers for their valuable comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Álvarez, H.; Perry, A.L.; Blanco, J.; García, S.; Aguilar, R. Towards the Creation of a Marine Protected Area in the Aeolian Islands. Results of the 2018 Aeolian Expediton, 1st ed.; Oceana: Madrid, Spain, 2019; pp. 1–136. [Google Scholar]
- Francalanci, L.; Avanzinelli, R.; Tommasini, S.; Heuman, A. A west east geochemical and isotopic traverse along the volcanism of the Aeolian Island arc, southern Tyrrhenian Sea, Italy: Inferences on mantle source processes. Geol. Soc. Am. Spec. Pap. 2007, 418, 235. [Google Scholar] [CrossRef]
- Esposito, V.; Andaloro, F.; Canese, S.; Bortoluzzi, G.; Bo, M.; Di Bella, M.; Giordano, P.; Spagnoli, F.; La Cono, V.; Yakimov, M.M.; et al. Exceptional discovery of a shallow-water hydrothermal site in the SW area of Basiluzzo islet (Aeolian archipelago, South Tyrrhenian Sea): An environment to preserve. PLoS ONE 2018, 13, e0190710. [Google Scholar] [CrossRef]
- Giaccone, G. Associazioni algali e fenomeni secondari di vulcanismo nelle acque marine di Vulcano (Mar Tirreno). Plant Biosyst. 1969, 103, 353–366. [Google Scholar] [CrossRef]
- Giaccone, G.; Cormaci, M.; Furnari, G.; Scammacca, B.; Alongi, G.; Catra, M.; Di Martino, V.; Marino, G.; Serio, D. Biodiversità vegetale marina dell’arcipelago “Isole Eolie”. Boll. Accad. Gioenia Nat. Sci. Catania 1999, 32, 191–242. [Google Scholar]
- Cavaliere, A. Ricerche sulla flora algologica dello Stretto di Messina. Primo contributo. Boll. Ist. Bot. Univ. Catania 1957, 1, 155–179. [Google Scholar]
- Cavaliere, A. Quelques notes sur les algues recueillies aux Isoles Eoliennes pendant una breve campagne thalassographique réalisée de Novembre 1957 a Fevrier 1958. Rapp. Commis. Int. Explor. Sci. Mer Médit. 1958, 15, 185–186. [Google Scholar]
- Cavaliere, A. Ricerche sulla flora algologica dello Stretto di Messina. Secondo contributo. Boll. Ist. Bot. Univ. Catania 1959, 3, 79–88. [Google Scholar]
- Giaccone, G. Raccolte di fitobenthos sulla banchina continentale Italiana. Plant Biosyst. 1969, 103, 485514. [Google Scholar] [CrossRef]
- Cormaci, M.; Furnari, G.; Scammacca, B.; Serio, D.; Pizzuto, F.; Alongi, G.; Dinaro, R. La vegetazione marina di substrato duro dell’isola di Salina (Isole Eolie). Boll. Accad. Gioenia Sci. Nat. Catania 1992, 25, 115–144. [Google Scholar]
- Cormaci, M.; Furnari, G.; Serio, D.; Pizzuto, F. Osservazioni sulle fitocenosi bentoniche dell’isola di Salina (Isole Eolie). In Parchi Naturali e Aree Protette; Guerrini, A., Ed.; Atti del Secondo Workshop del Progetto Strategico “Clima Ambiente e Territorio del Mezzogiorno” del C.N.R. Salina 20–30 Maggio 1990; Litografia Idonea G.: Catania, Italy, 1994; pp. 339–365. [Google Scholar]
- Alongi, G.; Pizzuto, F.; Scammacca, B. La flora sommersa dell’isola di Vulcano (Isole Eolie). Boll. Accad. Gioenia Sci. Nat. Catania 1993, 26, 273–291. [Google Scholar]
- Acunto, S.; Matagliati, F.; Rindi, F.; Rossi, F. Osservazioni floristiche sui popolamenti bentonici della baia di levante dell’Isola di Vulcano (Isole Eolie). Biol. Mar. Medit. 1997, 4, 348–350. [Google Scholar]
- Catra, M.; Alongi, G.; Giaccone, G. La flora sommersa dell’Isola di Filicudi (Isole Eolie). Boll. Accad. Gioenia Sci. Nat. Catania 1999, 32, 99–114. [Google Scholar]
- Giaccone, G. Contributo allo studio dei popolamenti algali del Basso Tirreno. Ann. Dell’Univ. Di Ferrara Nuova Ser. Bot. 1971, 4, 17–43. [Google Scholar]
- Acunto, S.; Matagliati, F.; Cinelli, F. Osservazioni sui popolamenti bentonici di un’area interessata da attività idrotermale nei pressi dell’Isola di Panarea (Isole Eolie). Biol. Mar. Medit. 1996, 3, 434–436. [Google Scholar]
- Acunto, S.; Rindi, F. Variabilità spaziale di popolamenti fitobentonici in relazione ad attività idrotermali nella Baia di Levante dell’isola di Vulcano (Isole Eolie): Studio preliminare. Biol. Mar. Medit. 1997, 4, 351–352. [Google Scholar]
- Bellissimo, G.; Rull Lluch, J.; Di Maida, G.; Pirrotta, M.; Tomasello, A.; Calvo, S. Influence of hydrothermal vents on phytobenthic communities in the Aeolian Islands (Tyrrhenian Sea): Preliminary results. In Proceedings of the 4th Mediterranean Symposium on Marine Vegetation, Yasmine-Hammamet, Tunisia, 2–4 December 2010. [Google Scholar]
- Bellissimo, G.; Rull Lluch, J.; Tomasello, A.; Calvo, S. The community of Cystoseira brachycarpa J. Agardh emend. Giaccone (Fucales, Phaeophyceae) in a shallow hydrothermal vent area of the Aeolian Islands (Tyrrhenian Sea, Italy). Plant Biosyst. 2014, 148, 21–26. [Google Scholar] [CrossRef]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef]
- Mangialajo, L.; Ruggieri, N.; Asnaghi, V.; Chiantore, M.; Povero, P.; Cattaneo-Vietti, R. Ecological status in the Ligurian Sea: The effect of coastline urbanisation and the importance of proper reference sites. Mar. Pollut. Bull. 2007, 55, 30–41. [Google Scholar] [CrossRef]
- Asnaghi, V.; Chiantore, M.; Bertolotto, R.M.; Parravicini, V.; Cattaneo-Vietti, R.; Gaino, F.; Moretto, P.; Privitera, D.; Mangialajo, L. Implementation of the European water framework directive: Natural variability associated with the CARLIT method on the rocky shores of the Ligurian Sea (Italy). Mar. Ecol. 2009, 30, 505–513. [Google Scholar] [CrossRef]
- Sales, M.; Cebrian, E.; Tomas, F.; Ballesteros, E. Pollution impacts and recovery potential in three species of the genus Cystoseira (Fucales, Heterokontophyta). Estuar. Coast. Shelf Sci. 2011, 92, 347–357. [Google Scholar] [CrossRef]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfune, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in the French Riviera: The harbinger of future extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Neiva, J.; Bermejo, R.; Medrano, A.; Capdevila, P.; MillaFigueras, D.; Afonso, P.; Ballesteros, E.; Sabour, B.; Serio, D.; Nóbrega, E.; et al. DNA barcoding reveals cryptic diversity, taxonomic conflicts and novel biogeographical insights in Cystoseira s.l. (Phaeophyceae). Eur. J. Phycol. 2022, 58, 351–375. [Google Scholar] [CrossRef]
- Marletta, G.; Lombardo, A. The Fucales (Ochrophyta, Phaeophyceae) of the Island of Pantelleria (Sicily Channel, Mediterranean Sea): A new contribution. Ital. Bot. 2023, 15, 137–163. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Chauvet, C.; Duval, C.; Galzin, R.; Lejeune, P.; Barnabé, G.; Blanc, F.; Chevalier, R.; Duclerc, J.; et al. The underwater observation of fish communities and fish populations: Methods and problems. Rev. Ecol. Terre Vie 1985, 40, 467–540. [Google Scholar]
- La Mesa, G.; Salvati, E.; Agnesi, S.; Tunesi, L. Assessment of coastal fish assemblages before the establishment of a new marine protected area in central Mediterranean: Its role in formulating zoning proposal. Mediterr. Mar. Sci. 2017, 18, 11–21. [Google Scholar] [CrossRef]
- Marletta, G.; Lombardo, A. Assessment of grazing impact on deep canopy-forming species in the western Ionian Sea, Central Mediterranean. Int. J. Aquat. Biol. 2020, 8, 365–376. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Cottalorda, J.M.; Hereu, B.; Susini, M.L.; Verlaque, M. Unexpected temporal stability of Cystoseira and Sargassum forests in Port-Cros, one of the oldest Mediterranean marine National Parks. Cryptogam. Algol. 2016, 37, 61–90. [Google Scholar] [CrossRef]
- Gómez-Garreta, A.; Barceló-Martí, M.C.; Ribera-Siguan, M.A.; Rull-Lluch, J. Cystoseira C. Agardh. In Flora Phycologica Iberica, 1st ed.; Gómez-Garreta, A., Ed.; Universidade de Murcia: Múrcia, Spania, 2001; Volume 1, Fucales; pp. 99–166. [Google Scholar]
- Cormaci, M.; Furnari, G.; Catra, M.; Alongi, G.; Giaccone, G. Flora marina bentonica del Mediterraneo: Phaeophyceae. Boll. Accad. Gioenia Sci. Nat. Catania 2012, 45, 1–508. [Google Scholar]
- Rodríguez-Prieto, C.; Ballesteros, E.; Boisset, F.; Afonso Carrilo, J. Guía de Las Macroalgas y Fanerógamas Marinas del Mediterráneo Occidental; Ediciones Omega: Barcelona, Spain, 2013; pp. 1–656. [Google Scholar]
- Blanfuné, A.; Verlaque, M.; Boudouresque, C.F.; Rozis, E.; Thibaut, T. Les Forêts Marines de France et de Méditerranée. Guide de Détermination des Espèces-Ingénieurs. Sargassaceae, Fucales, Phaeophyceae; Presses Universitaries de Provence: Marseille, France, 2022; pp. 1–207. [Google Scholar]
- Gray, J.S. The measurement of marine species diversity, with an application to the benthic fauna of the Norwegian continental shelf. J. Exp. Mar. Biol. Ecol. 2000, 250, 23–49. [Google Scholar] [CrossRef]
- Danovaro, R.; Bianchelli, S.; Gambi, C.; Mea, M.; Zeppilli, D. α-, β-, γ-, δ-and ε-diversity of deep-sea nematodes in canyons and open slopes of Northeast Atlantic and Mediterranean margins. Mar. Ecol. Prog. Ser. 2009, 396, 197–209. [Google Scholar] [CrossRef]
- Bianchelli, S.; Buschi, E.; Danovaro, R.; Pusceddu, A. Biodiversity loss and turnover in alternative states in the Mediterranean Sea: A case study on meiofauna. Sci. Rep. 2016, 6, 34544. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed]
- Gamito, S. Caution is needed when applying Margalef diversity index. Ecol. Indic. 2010, 10, 550–551. [Google Scholar] [CrossRef]
- AlgaeBase. Available online: https://www.algaebase.org (accessed on 30 July 2023).
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 2008, 358, 63–74. [Google Scholar] [CrossRef]
- Montagne, C. Cryptogames algériennes, ou plantes cellulaires recueillies par M. Roussel aux environs d’Alger, et publiées par le Docteur Camille Montagne. Ann. Sci. Nat. Bot. 1838, 2, 268–279 (+ Suppl. 337–345, plates 8–9). [Google Scholar]
- Sellam, L.N.; Blanfuné, A.; Boudouresque, C.F.; Thibaut, T.; Rebzani Zahaf, C.; Verlaque, M. Cystoseira montagnei J. Agardh and C. spinosa Sauvageau (Phaeophyceae, Sargassaceae): A taxonomic reappraisal of misused names, with the proposal of Cystoseira michaelae Verlaque et al. nom. et stat. nov. Cryptogam. Algol. 2017, 38, 133–157. [Google Scholar] [CrossRef]
- Tita, G. Aspects écologiques d’un peuplement à Cystoseira dubia Valiante. Mar. Life 1994, 4, 9–17. [Google Scholar]
- Verlaque, M.; Ballesteros, E.; Sala, E.; Garrabou, J. Cystoseira jabukae (Cystoseiraceae, Fucophyceae) from Corsica (Mediterranean) with notes on the previously misunderstood species C. funkii. Phycologia 1999, 38, 77–86. [Google Scholar] [CrossRef]
- Ercegovic, A. Sur les Cystoseira Adriatiques, leur Morphologie, Ecologie et Evolution. Fauna i Flora Jadrana, 1st ed.; Institut Oceanographie et Peche: Split, Croatia, 1952; Volume 2, pp. 1–212. [Google Scholar]
- Rendina, F.; Falace, A.; Alongi, G.; Buia, M.C.; Neiva, J.; Appolloni, L.; Marletta, G.; Russo, G.F. The Lush Fucales Underwater Forests off the Cilento Coast: An Overlooked Mediterranean Biodiversity Hotspot. Plants 2023, 12, 1497. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Verlaque, M.; Boudouresque, C.F.; Ruitton, S. The Sargassum conundrum: Very rare, threatened or locally extinct in the NW Mediterranean and still lacking protection. Hydrobiologia 2016, 781, 3–23. [Google Scholar] [CrossRef]
- Flores-Moya, A.; Conde, F. Fragmentos taxonómicos, corológicos, nomenclaturales y fitosociológicos (67–74). Acta Bot. Malacit. 1998, 23, 197–228. [Google Scholar] [CrossRef][Green Version]
- Tiralongo, F.; Akyol, O.; Al Mabruk, S.A.; Battaglia, P.; Beton, D.; Bitlis, B.; Borg, J.A.; Bouchoucha, M.; Çinar, M.E.; Crocetta, F.; et al. New Alien Mediterranean Biodiversity Records (August 2022). Mediterr. Mar. Sci. 2022, 23, 725–747. [Google Scholar] [CrossRef]
- Ballesteros, E.; Weitzmann, B. On the presence of a small population of Sargassun trichocarpum J. Agardh (Phaeophyceae: Fucales) in Catalonia (Northwestern Mediterranean). Butll. Inst. Catalana Hist. Nat. 2021, 85, 83–85. [Google Scholar]
- Verlaque, M.; Boudouresque, C.F.; Perret-Boudouresque, M. Mediterranean seaweeds listed as threatened under the Barcelona Convention: A critical analysis. Sci. Rep. Port Cros Natl. Park 2019, 33, 179–214. [Google Scholar]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Lombardo, A.; Marletta, G. The marine Heterobranchia (Mollusca: Gastropoda) fauna of the Aeolian archipelago (Tyrrhenian Sea). First contribution: Lipari and Vulcano. Int. J. Aquat. Biol. 2023, 11, 288–300. [Google Scholar] [CrossRef]
- Civitarese, G.; Gačić, M.; Lipizer, M.; Eusebi Borzelli, G.L. On the impact of the Bimodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian Seas (eastern Mediterranean). Biogeosciences 2010, 7, 3987–3997. [Google Scholar] [CrossRef]
- Ben Rais Lasram, F.; Tomasini, J.A.; Romdhane, M.S.; Do Chi, T.; Mouillot, D. Historical colonization of the Mediterranean Sea by Atlantic fishes: Do biological traits matter? Hydrobiologia 2008, 607, 51–62. [Google Scholar] [CrossRef]
- Darling, E.S.; Côté, I.M. Seeking resilience in marine ecosystems. Science 2018, 359, 986–987. [Google Scholar] [CrossRef] [PubMed]
- Serio, D.; Alongi, G.; Catra, M.; Cormaci, M.; Furnari, G. Changes in the benthic algal flora of Linosa Island (Straits of Sicily, Mediterranean Sea). Bot. Mar. 2006, 49, 135–144. [Google Scholar] [CrossRef]
- ISPRA, Istituto Superiore per la Protezione e la Ricerca Ambientale. Available online: https://www.mareografico.it/it/stazioni.html (accessed on 11 November 2023).
- Verdura, J.; Santamaría, J.; Ballesteros, E.; Smale, D.; Cefalì, M.E.; Golo, R.; de Caralt, S.; Vergés, A.; Cebrian, E. Local-scale climatic refugia offer sanctuary for a habitat-forming species during a marine heatwave. J. Ecol. 2021, 109, 1758–1773. [Google Scholar] [CrossRef]
- Celis-Plá, P.S.M.; Martinez, B.; Korbee, N.; Hall-Spencer, J.M.; Figueroa, F.L. Ecophysiological responses to elevated CO2 and temperature in Cystoseira tamariscifolia (Phaeophyceae). Clim. Chang. 2017, 142, 67–81. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Savonitto, G.; Lipizer, M.; Mancuso, P.; Ciriaco, S.; Srijemsi, M.; Falace, A. Climatic anomalies may create a long-lasting ecological phase shift by altering the reproduction of a foundation species. Ecology 2019, 100, e02838. [Google Scholar] [CrossRef] [PubMed]
- Garrabou, J.; Gómez-Gras, D.; Medrano, A.; Cerrano, C.; Ponti, M.; Schlegel, R.; Bensoussan, N.; Turicchia, E.; Sini, M.; Gerovasileiou, V.; et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Chang. Biol. 2022, 28, 5708–5725. [Google Scholar] [CrossRef] [PubMed]
- Orfanidis, S.; Rindi, F.; Cebrian, E.; Fraschetti, S.; Nasto, I.; Taskin, E.; Bianchelli, S.; Papathanasiou, V.; Kosmidou, M.; Caragnano, A.; et al. Effects of Natural and Anthropogenic Stressors on Fucalean Brown Seaweeds Across Different Spatial Scales in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 658417. [Google Scholar] [CrossRef]
- Arévalo, R.; Pinedo, S.; Ballesteros, E. Changes in the composition and structure of Mediterranean rocky-shore communities following a gradient of nutrient enrichment: Descriptive study and test of proposed methods to assess water quality regarding macroalgae. Mar. Pollut. Bull. 2007, 55, 104–113. [Google Scholar] [CrossRef]
- Sales, M.; Ballesteros, E. Shallow Cystoseira (Fucales: Ochrophyta) assemblages thriving in sheltered areas from Menorca (NW Mediterranean): Relationships with environmental factors and anthropogenic pressures. Estuar. Coast. Shelf Sci. 2009, 84, 476–482. [Google Scholar] [CrossRef]
- Tsiamis, K.; Panayotidis, P.; Salomidi, M.; Pavlidou, A.; Kleinteich, J.; Balanika, K.; Küpper, F.C. Macroalgal community response to re-oligotrophication in Saronikos Gulf. Mar. Ecol. Prog. Ser. 2013, 472, 73–85. [Google Scholar] [CrossRef]
- Vergés, A.; Tomas, F.; Cebrian, E.; Ballesteros, E.; Kizilkaya, Z.; Dendrinos, P.; Karamanlidis, A.A.; Spiegel, D.; Sala, E. Tropical rabbitfish and the deforestation of a warming temperate sea. J. Ecol. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Capdevila, P.; Hereu, B.; Salguero-Gómez, R.; Rovira, G.; Medrano, A.; Cebrian, E.; Garrabou, J.; Kersting, D.K.; Linares, C. Warming impacts on early life stages increase the vulnerability and delay the population recovery of a long-lived habitat forming macroalga. J. Ecol. 2019, 107, 1129–1140. [Google Scholar] [CrossRef]
- Falace, A.; Marletta, G.; Savonitto, G.; Carniel, F.C.; Srijemsi, M.; Bevilacqua, S.; Tretiach, M.; Alongi, G. Is the South-Mediterranean Canopy-Forming Ericaria giacconei (=Cystoseira hyblaea) a loser from Ocean Warming? Front. Mar. Sci. 2021, 8, 760637. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of vulnerable Cystoseira populations along the Italian infralittoral fringe, and relationships with environmental and anthropogenic variables. Mar. Pollut. Bull. 2018, 129, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Marletta, G. Status of shallow Fucales (Ochrophyta) assemblages in the bay of Brucoli, eastern coast of Sicily (Ionian Sea). Biodivers. J. 2021, 12, 1011–1026. [Google Scholar] [CrossRef]















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).