


Ecology and Phenology of the Subtidal Brown Alga Sargassum furcatum (Ochrophyta, Fucales), a Likely Non-Indigenous Species from the Mediterranean Sea


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Depth (m) | Substrate | Reference |
|---|---|---|---|
| Atlantic Ocean | |||
| Virgin Islands and Antilles | - | - | [20] |
| Venezuela | - | - | [21] |
| Salvage Islands | - | - | [22] |
| Cuba | 2–4 | Rocky | [9,23] |
| Brazil | 1–5 | Sandy | [19,24,25] |
| Trinidad and Tobago | - | - | [26] |
| Mexico | - | - | [27,28] |
| Canary Islands | - | - | [29] |
| Madeira | - | - | [30] |
| Costa Rica | 0–1.5 | Rocky and sandy | [31] |
| Azores | - | - | [32] |
| Puerto Rico | Up to 17 | - | [33] |
| Pacific Ocean | |||
| Philippines | - | - | [34] |
| Mediterranean Sea | |||
| Spain | 12 | - | [16] |
| Italy | 4–28 | Rocky and sandy | [14,18] |
| France and Corsica | Upper and medium infralittoral | - | [17] |
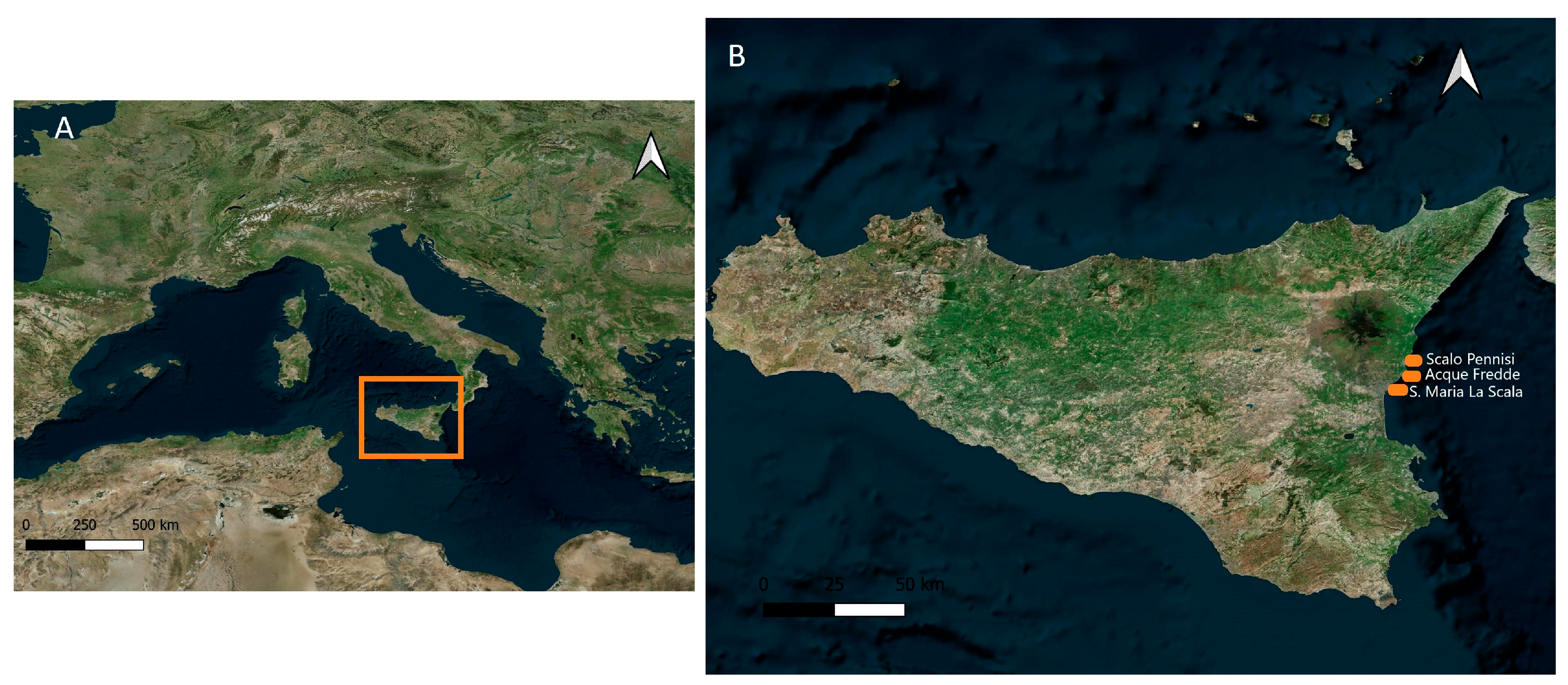
2. Materials and Methods
3. Results
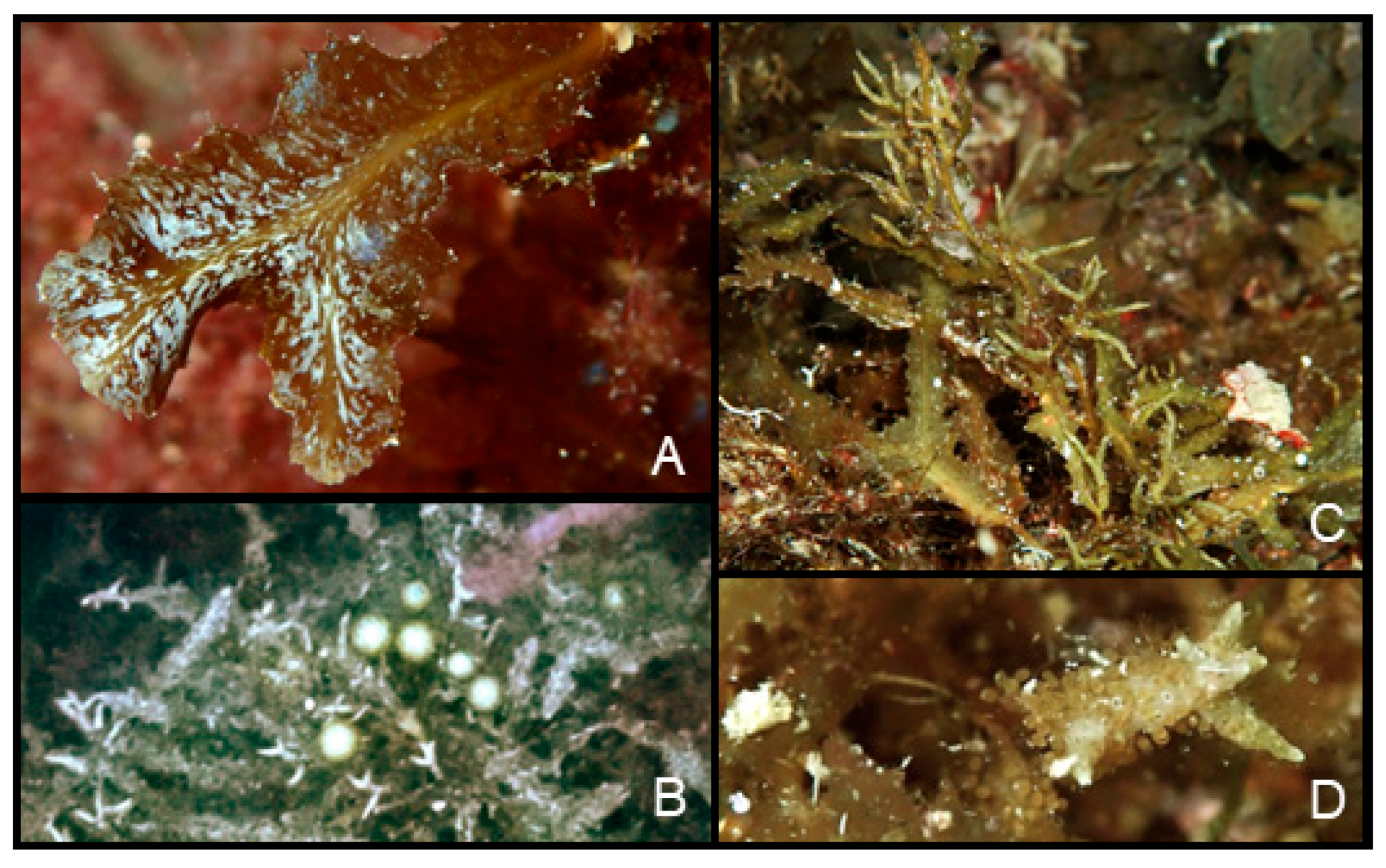
3.1. Description of the Studied Specimens
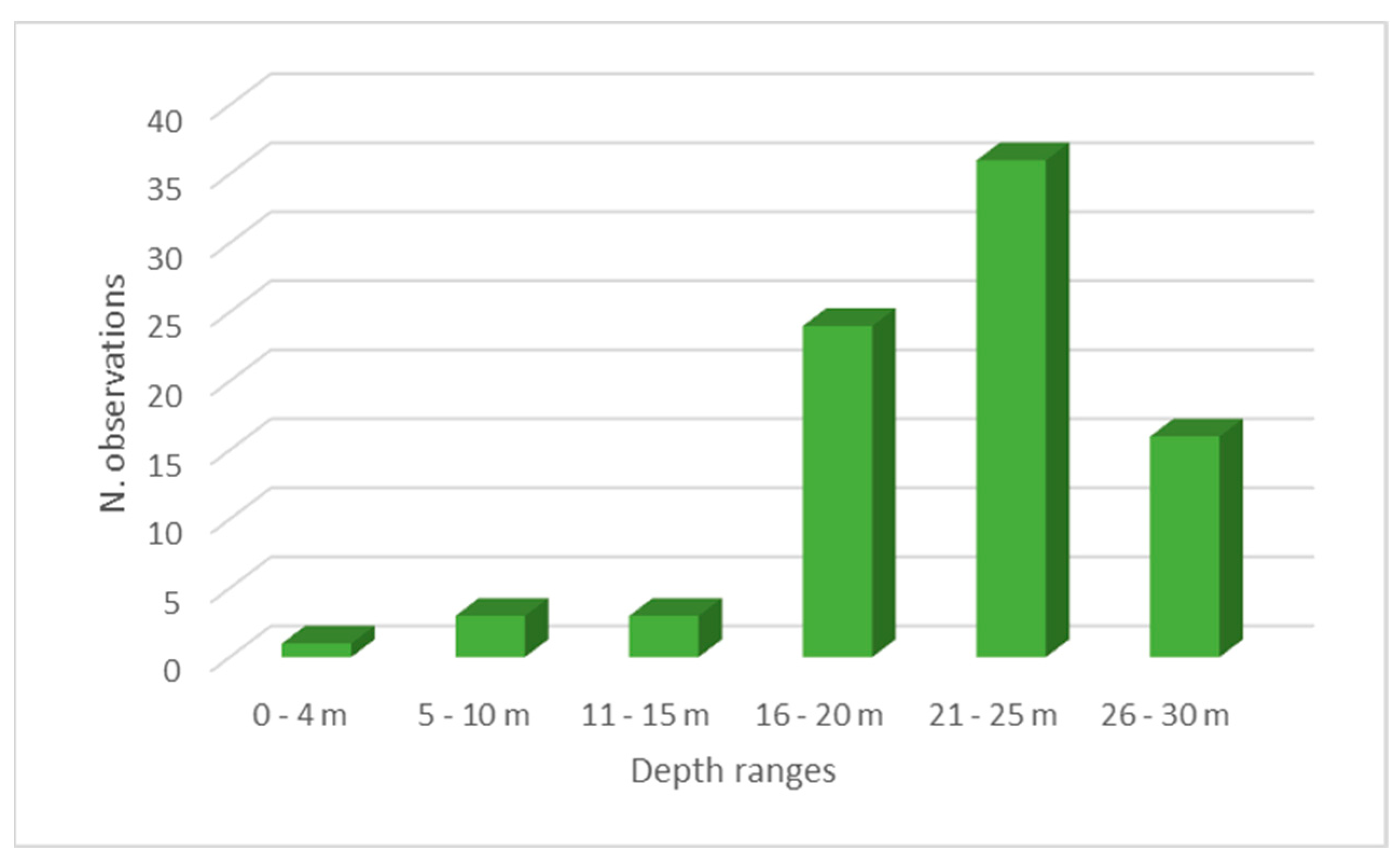
3.2. Habitat and Distribution
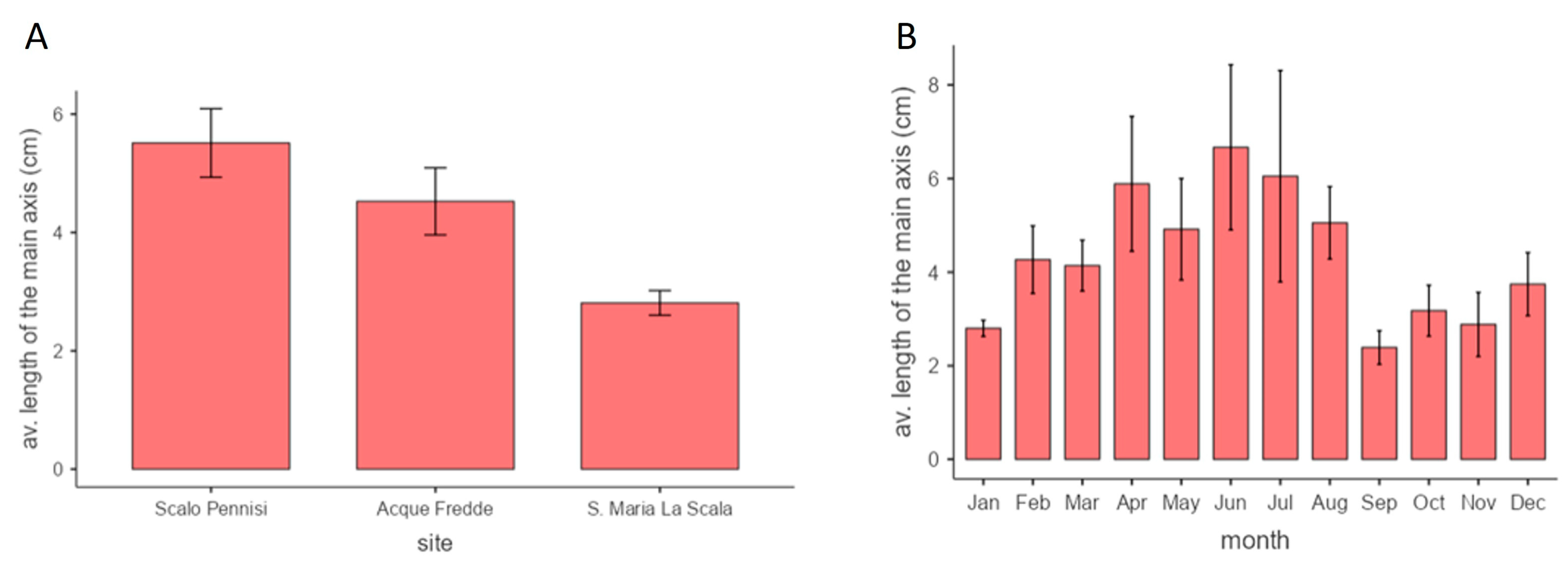
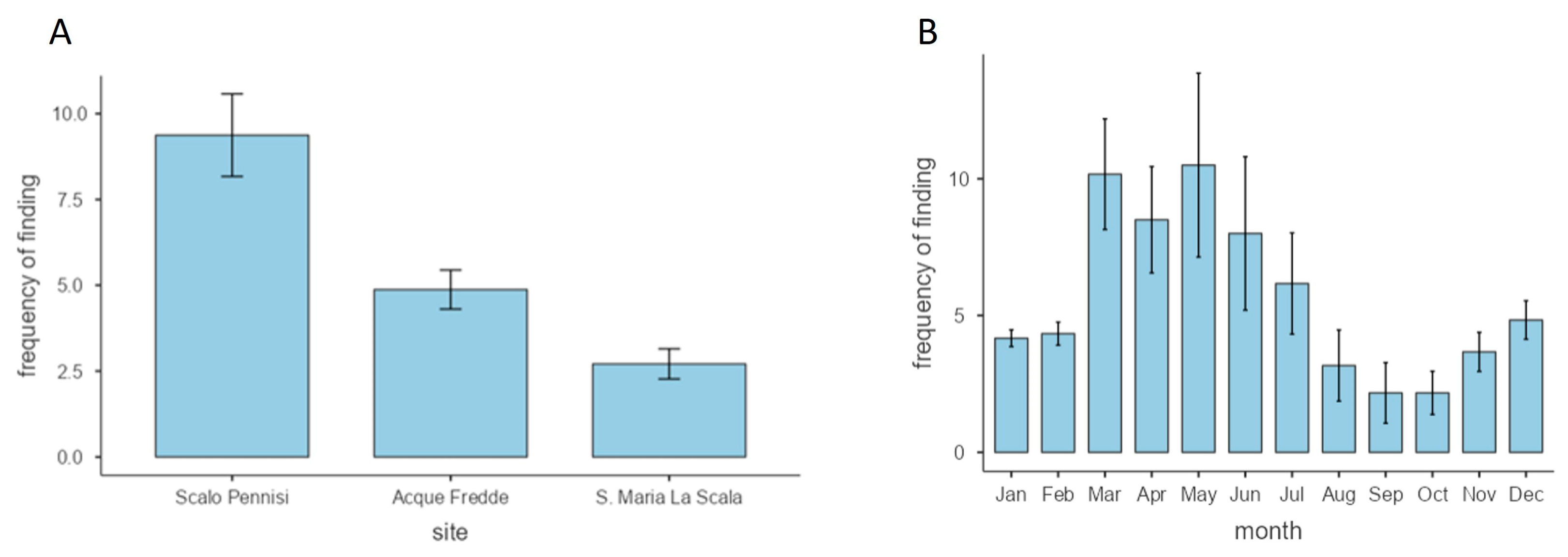
3.3. Phenology of Sargassum furcatum during the Year
3.4. Frequency of Sargassum furcatum during the Year
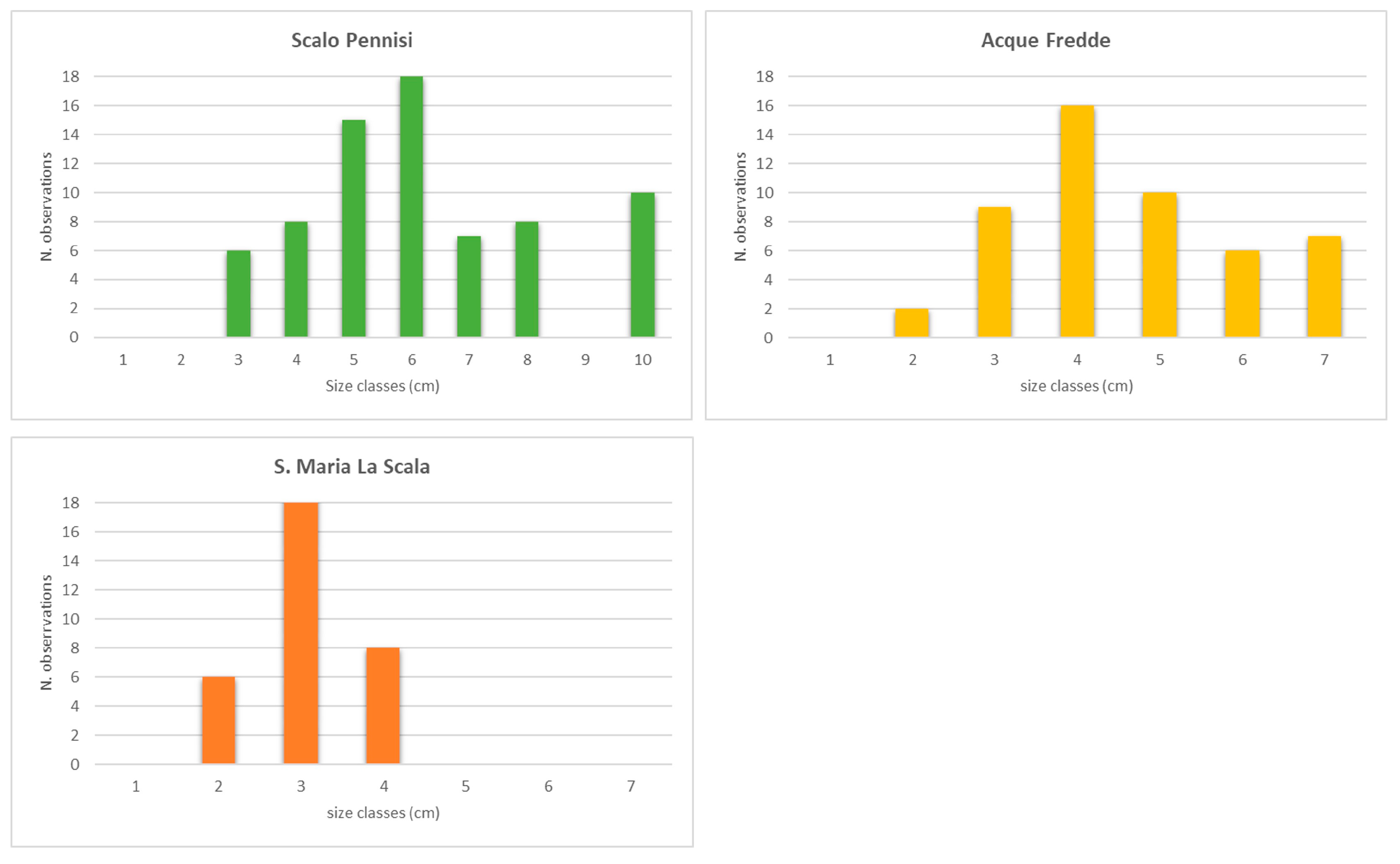
3.5. Size Distribution Frequencies
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dayton, P.K. Ecology of kelp communities. Annu. Rev. Ecol. Evol. Syst. 1985, 16, 215–245. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp Forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Schiel, D.R.; Foster, M.S. The population biology of large brown seaweeds: Ecological consequences of multiphase life histories in dynamic coastal environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Verlaque, M.; Boudouresque, C.F.; Ruitton, S. The Sargassum conundrum: Very rare, threatened or locally extinct in the NW Mediterranean and still lacking protection. Hydrobiologia 2016, 781, 3–23. [Google Scholar] [CrossRef]
- Reed, D.C.; Rassweiler, A.; Arkema, K.K. Biomass rather than growth rate determines variation in net primary production by giant kelp. Ecology 2008, 89, 2493–2505. [Google Scholar] [CrossRef] [PubMed]
- Wanders, J.B.W. The role of benthic algae in the shallow reef of Curaçao (Netherlands Antilles). II. Primary productivity of the Sargassum beds on the northeast coast submarine plateau. Aquat. Bot. 1976, 2, 327–335. [Google Scholar] [CrossRef]
- Guimaraens, M.A.; Combells, C.; Corbett, C. Species diversity and richness of reef building corals and macroalgae of reef communities in Discovery Bay, Jamaica. Acta Biol. Leopol. 1994, 16, 41–50. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 14 February 2024).
- Moreira, L.; Suárez, A.M. Study of the Genus Sargassum C. Agardh, 1820 (Phaeophyta, Fucales, Sargassaceae) in Cuban Waters. 1. Sargassum furcatum Kützing, 1843, New Record. Rev. Invest. Mar. 2002, 23, 53–54. [Google Scholar]
- Mattio, L.; Payri, C.E. 190 years of Sargassum taxonomy, facing the advent of DNA phylogenies. Bot. Rev. 2011, 77, 31–70. [Google Scholar] [CrossRef]
- Yip, Z.T.; Quek, Z.B.R.; Huang, D. Historical biogeography of the widespread macroalga Sargassum (Fucales, Phaeophyceae). J. Phycol. 2020, 56, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Cormaci, M.; Furnari, G.; Catra, M.; Alongi, G.; Giaccone, G. Flora marina bentonica del Mediterraneo: Phaeophyceae. Boll. Accad. Gioenia Sci. Nat. 2012, 45, 1–508. [Google Scholar]
- Aouissi, M.; Sellam, L.; Boudouresque, C.; Blanfuné, A.; Derbal, F.; Frihi, H.; Perretboudouresque, M.; Rebzani-Zahaf, C.; Verlaque, M.; Thibaut, T. Insights into the species diversity of the genus Sargassum (Phaeophyceae) in the Mediterranean Sea, with a focus on a previously unnoticed taxon from Algeria. Mediterr. Mar. Sci. 2018, 19, 48–57. [Google Scholar] [CrossRef]
- Tiralongo, F.; Akyol, O.; Al Mabruk, S.A.; Battaglia, P.; Beton, D.; Bitlis, B.; Borg, J.A.; Bouchoucha, M.; Çinar, M.E.; Crocetta, F.; et al. New Alien Mediterranean Biodiversity Records (August 2022). Mediterr. Mar. Sci. 2022, 23, 725–747. [Google Scholar] [CrossRef]
- Kützing, F.T. Phycologia Generalis Oder Anatomie, Physiologie und Systemkunde der Tange; Brockhaus: Leipzig, Germany, 1843; pp. 143–458. [Google Scholar]
- Flores-Moya, A.; Conde, F. Fragmentos taxonómicos, corológicos, nomenclaturales y fitosociológicos. Acta Bot. Malacit. 1998, 23, 197–228. [Google Scholar] [CrossRef]
- Blanfuné, A.; Verlaque, M.; Boudouresque, C.F.; Rozis, E.; Thibaut, T. Les Forêts Marines de France et de Méditerranée. Guide de Détermination des Espèces-Ingénieurs. Sargassaceae, Fucales, Phaeophyceae; Presses Universitaries de Provence: Marseille, France, 2022; pp. 1–207. [Google Scholar]
- Marletta, G.; Lombardo, A.; Serio, D.; Bianchelli, S. Diversity of Fucales (Ochrophyta, Phaeophyceae) along the Coasts of Lipari and Vulcano (Aeolian Archipelago), Tyrrhenian Sea (Central Mediterranean Sea). J. Mar. Sci. Eng. 2023, 11, 2222. [Google Scholar] [CrossRef]
- Godoy, E.A.S.; Coutinho, R. Can artificial beds of plastic mimics compensate for seasonal absence of natural beds of Sargassum furcatum? ICES J. Mar. Sci. 2002, 59, S111–S115. [Google Scholar] [CrossRef]
- Taylor, W.R. Marine Algae of the Eastern Tropical and Subtropical Coasts of the Americas; The University of Michigan Press: Ann Arbor, MI, USA, 1960; pp. 1–870. [Google Scholar]
- Ganesan, E.K. A Catalog of Benthic Marine Algae and Seagrasses of Venezuela; Fondo Editorial Conicit: Caracas, Venezuela, 1990; pp. 1–237. [Google Scholar]
- Parente, M.I.; Gil-Rodríguez, M.C.; Haroun, R.J.; Neto, A.I.; de Smedt, G.; Hernández-González, C.L.; Berecibar Zugasti, E. Flora marina de las Ilhas Selvagens: Resultados preliminares de la expedición “Macronesia 2000”. Rev. Acad. Canar. Cienc. 2000, 12, 9–20. [Google Scholar]
- Suárez, A.M.; Martínez-Daranas, B.; Alfonso Sánchez, Y.; Moreira-González, A.R.; Capote, J. Lista actualizada de las macroalgas marinas cubanas. Acta Bot. Mex. 2023, 130, e2196. [Google Scholar] [CrossRef]
- Pereira, R.C.; Bianco, É.M.; Bueno, L.B.; De Oliveira, M.A.L.; Pamplona, O.S.; Da Gama, B.A.P. Associational defense against herbivory between brown seaweeds. Phycologia 2010, 49, 424–428. [Google Scholar] [CrossRef]
- Paula, J.C.; Coracao, A.; Lopes-Filho, E.A.; Silva, R.P.; Santos, L.N.; Carvalho, W.F. Diversity and turnover in a rocky shore intertidal community of an upwelling region (Arraial do Cabo, Brazil). An. Acad. Bras. Cienc. 2020, 92, e20181096. [Google Scholar] [CrossRef]
- Duncan, E.J.; Lee Lum, L.M. A checklist of the marine macroalgae of the Republic of Trinadad and Tobago. Caribb. Mar. Stud. 2006, 7, 1–96. [Google Scholar]
- Robinson, N.M.; Galicia-García, C.; Okolodkov, Y.B. New records of green (Chlorophyta) and brown algae (Phaeophyceae) for Cabezo Reef, National Park Sistema Arrecifal Veracruzano, Gulf of Mexico. Acta Bot. Mex. 2012, 101, 11–48. [Google Scholar]
- Pedroche, F.F.; Sentíes, A. Diversidad de macroalgas marinas en México. Una actualización florística y nomenclatural. Cymbella 2020, 6, 4–55. [Google Scholar]
- Afonso-Carrillo, J. Lista Actualizada de las Algas Marinas de las Islas Canarias; Elaborada para la Sociedad Española de Ficología (SEF): Las Palmas, Spain, 2014; pp. 1–64. [Google Scholar]
- Ferreira, S.J.; Gonçalves Silva, J.J.; Araujo, R. Marine algae collection in the Herbarium of the Funchal Natural History Museum (MADM) with new records from the archipelago of Madeira. Bol. Mus. Munic. Funchal 2018, 68, 31–52. [Google Scholar]
- Cabrera, R.; Díaz-Larrea, J.; Umanzor, S. New Records of Marine Macroalgae on the Caribbean on Coast of Costa Rica. Am. J. Plant Sci. 2019, 10, 1708–1728. [Google Scholar] [CrossRef]
- Neto, A.I.A.; Parente, M.I.; Botelho, A.Z.; Prestes, A.C.L.; Resendes, R.; Afonso, P.; Álvaro, N.V.; Milla-Figueras, D.; Neto, R.M.A.; Tittley, I.; et al. Marine algal flora of Graciosa Island, Azores. Biodivers. Data J. 2020, 8, e57201. [Google Scholar] [CrossRef]
- Ballantine, D.L.; Norris, J.N.; Ruiz, H. The Marine Benthic Algal Flora of Puerto Rico, I. Ochrophyta: Phaeophyceae, Pelagophyceae, and Xanthophyceae. Smithson. Contrib. Bot. 2021, 114, 1–111. [Google Scholar] [CrossRef]
- Ang, P.O., Jr.; Leung, S.M.; Choi, M.M. A verification of reports of marine algal species from the Philippines. Philipp. J. Sci. 2014, 142, 5–49. [Google Scholar]
- Catra, M.; Giaccone, T.; Giardina, S.; Nicastro, A. Il patrimonio naturale marino bentonico della Timpa di Acireale (Catania). Boll. Accad. Gioenia Sci. Nat. 2006, 39, 129–158. [Google Scholar]
- Sciuto, F.; Sanflippo, R.; Alongi, G.; Catra, M.; Serio, D.; Bejaoui, S.; Leonardi, R.; Viola, A.; Rosso, A. First data on ostracods and foraminifera living in Cystoseira communities in western Ionian Sea (southern Italy, Mediterranean Sea). Mediterr. Mar. Sci. 2017, 18, 393–405. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Chauvet, C.; Duval, C.; Galzin, R.; Lejeune, P.; Barnabe, G.; Blanc, F.; Chevalier, R.; Duclerc, J.; et al. Evaluation visuelle des peuplements et populations de poissons: Methodes et problems. Rev. Ecol.-Terre Vie 1985, 40, 467–539. [Google Scholar] [CrossRef]
- Marletta, G.; Lombardo, A. The Fucales (Ochrophyta, Phaeophyceae) of the Island of Pantelleria (Sicily Channel, Mediterranean Sea): A new contribution. Ital. Bot. 2023, 15, 137–163. [Google Scholar] [CrossRef]
- Navarro, L.; Ballesteros, E.; Linares, C.; Hereu, B. Spatial and temporal variability of deepwater algal assemblages in the Northwestern Mediterranean: The effects of an exceptional storm. Est. Coast. Shelf. Sci. 2011, 95, 52–58. [Google Scholar] [CrossRef]
- Marletta, G.; Lombardo, A. Population dynamics of Ericaria zosteroides (Ochrophyta, Fucales) in the central Mediterranean. Mediterr. Bot. 2023, 44, e79885. [Google Scholar] [CrossRef]
- Di Stefano, A.; De Pietro, R.; Monaco, C.; Zanini, A. Anthropogenic influence on coastal evolution: A case history from the Catania Gulf shoreline (eastern Sicily, Italy). Ocean Coast. Manag. 2013, 80, 133–148. [Google Scholar] [CrossRef]
- Einav, R.; Guiry, M.D.; Israel, A. A revised list of seaweeds from the Red Sea (1756–2020). Isr. J. Plant Sci. 2021, 68, 175–247. [Google Scholar] [CrossRef]
- Turner, D. Fuci Sive Plantarum Fucorum Generi a Botanicis Ascriptarum Icones Descriptiones et Historia. Fuci, or Coloured Figures and Descriptions of the Plants Referred by Botanists to the Genus Fucus; Typis J. M’Creery, Impensis J. et A. Arch: London, UK, 1808; Volume II, pp. 1–164. [Google Scholar]
- Boudouresque, C.-F.; Perret-Boudouresque, M.; Blanfuné, A. Diversity of marine and brackish macrophytes in the Port-Cros National Park (Provence, France, Mediterranean Sea): Taxa and research effort over space and time. Diversity 2022, 14, 329. [Google Scholar] [CrossRef]
- Grunow, A. Additamenta ad cognitionem Sargassorum. Verh. Zool.-Bot. Ges. Wien 1916, 66, 136–185. [Google Scholar]
- Lima, F.P.; Ribeiro, P.A.; Queiroz, N.; Hawkins, S.J.; Santos, A.M. Do distributional shifts of northern and southern species of algae match the warming pattern? Glob. Chang. Biol. 2007, 13, 2592–2604. [Google Scholar] [CrossRef]
- Russell, B.D.; Thompson, J.A.I.; Falkenberg, L.J.; Connell, S.D. Synergistic effects of climate change and local stressors: CO2, and nutrient-driven change in subtidal rocky habitats. Glob. Chang. Biol. 2009, 15, 2153–2162. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in French Riviera: The harbinger of future extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Faga, F.; Gurgel, C.F.D. Distributional range shifts of Western Atlantic benthic Sargassum species (Fucales, Phaeophyceae) under future climate change scenarios. Aquat. Bot. 2024, 190, 103705. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marletta, G.; Lombardo, A.; Serio, D. Ecology and Phenology of the Subtidal Brown Alga Sargassum furcatum (Ochrophyta, Fucales), a Likely Non-Indigenous Species from the Mediterranean Sea. J. Mar. Sci. Eng. 2024, 12, 640. https://doi.org/10.3390/jmse12040640
Marletta G, Lombardo A, Serio D. Ecology and Phenology of the Subtidal Brown Alga Sargassum furcatum (Ochrophyta, Fucales), a Likely Non-Indigenous Species from the Mediterranean Sea. Journal of Marine Science and Engineering. 2024; 12(4):640. https://doi.org/10.3390/jmse12040640
Chicago/Turabian StyleMarletta, Giuliana, Andrea Lombardo, and Donatella Serio. 2024. "Ecology and Phenology of the Subtidal Brown Alga Sargassum furcatum (Ochrophyta, Fucales), a Likely Non-Indigenous Species from the Mediterranean Sea" Journal of Marine Science and Engineering 12, no. 4: 640. https://doi.org/10.3390/jmse12040640
APA StyleMarletta, G., Lombardo, A., & Serio, D. (2024). Ecology and Phenology of the Subtidal Brown Alga Sargassum furcatum (Ochrophyta, Fucales), a Likely Non-Indigenous Species from the Mediterranean Sea. Journal of Marine Science and Engineering, 12(4), 640. https://doi.org/10.3390/jmse12040640