
Mechanical Properties, Tissue Structure, and Elemental Composition of the Walking Leg Tips of Coconut Crabs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
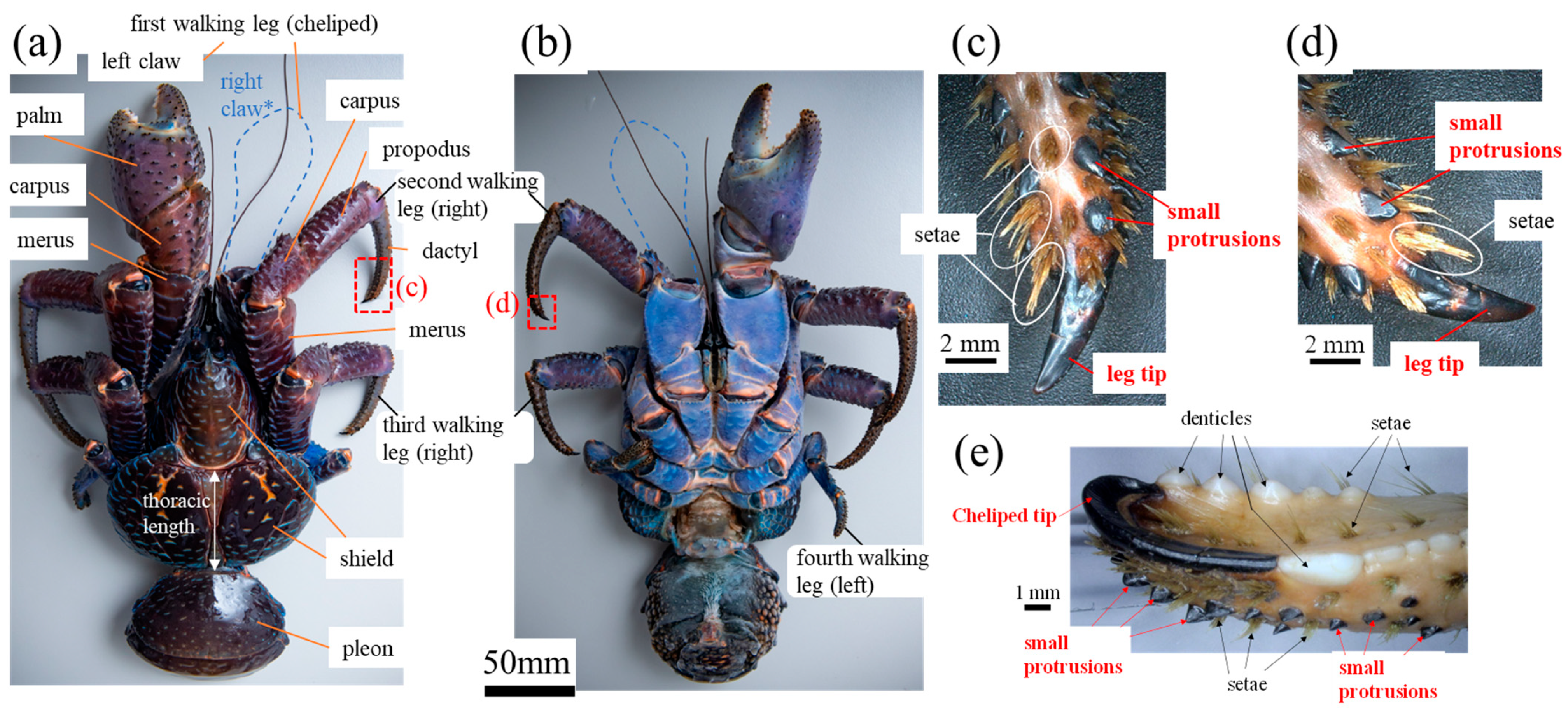
2.1. Sample Preparation
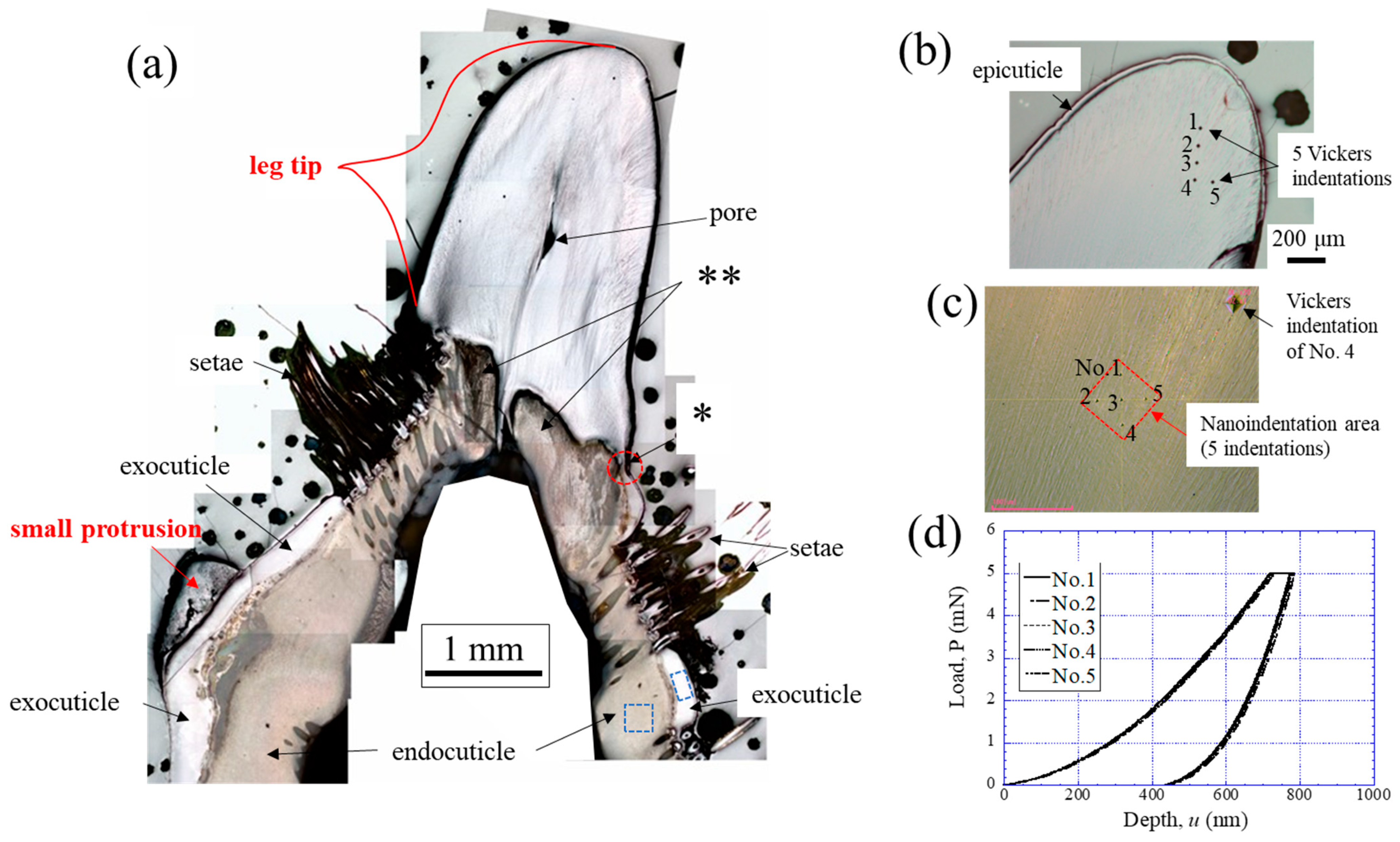
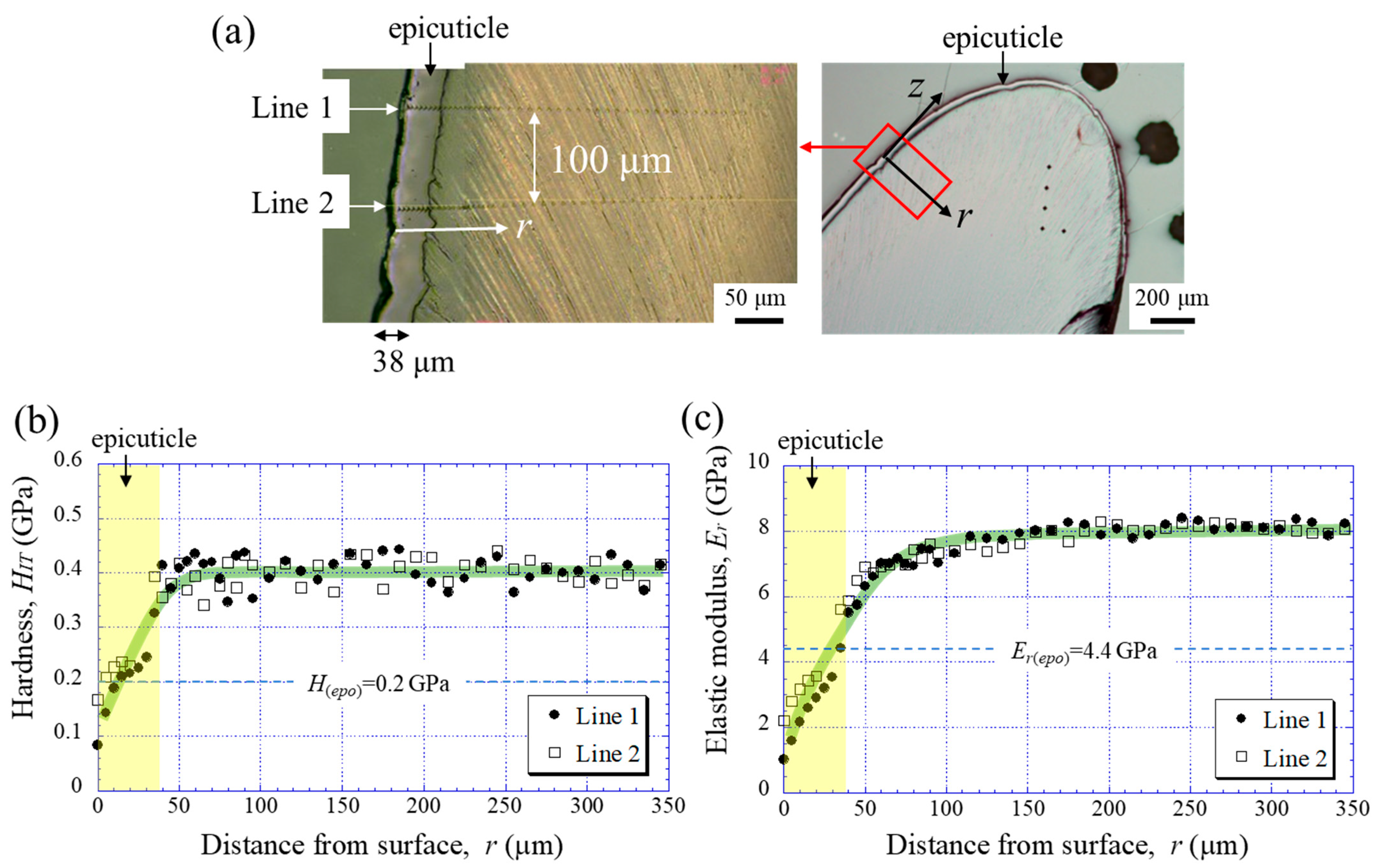
2.2. Vickers Hardness and Nanoindentation Testing
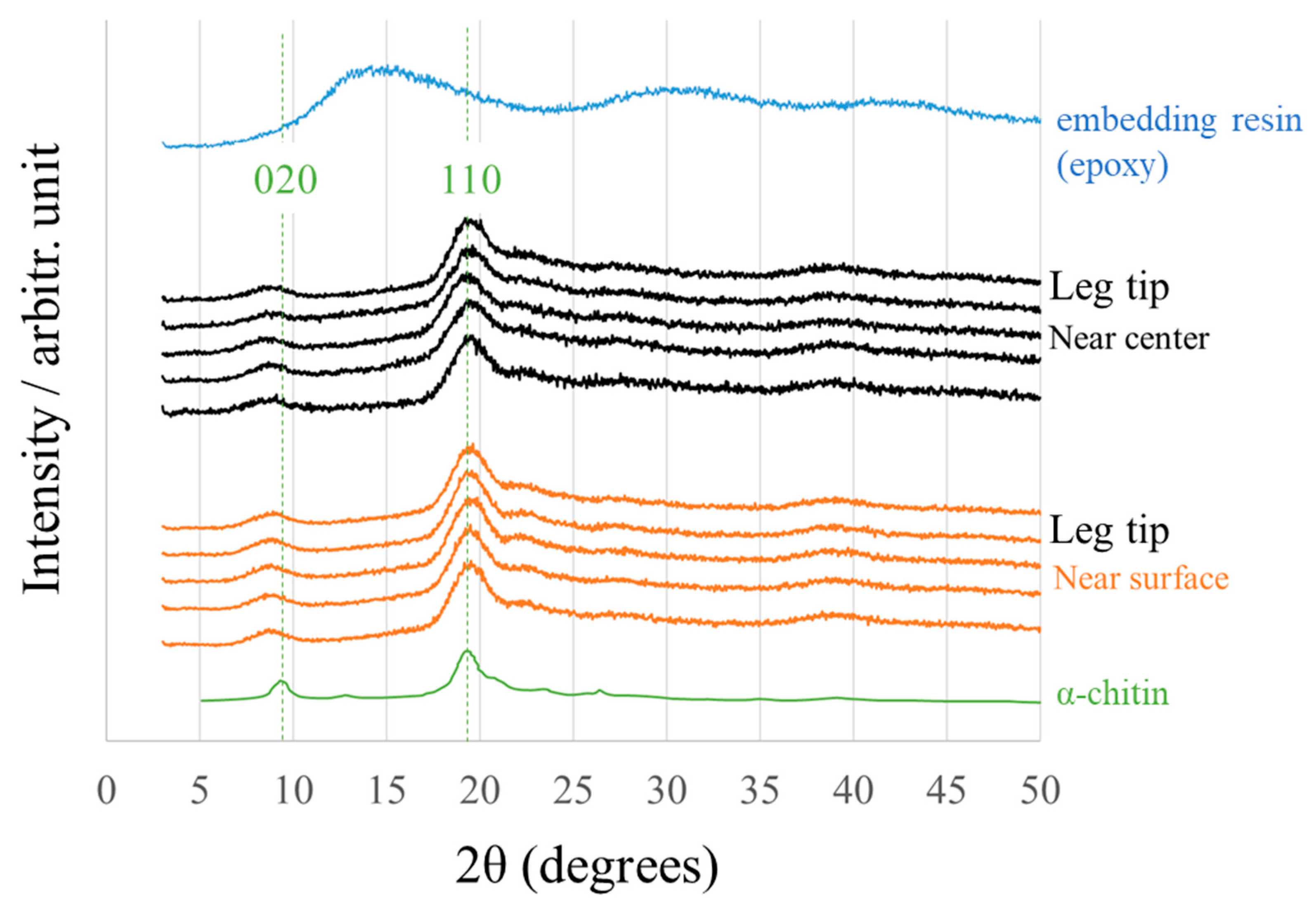
2.3. X-ray Diffraction
2.4. Microstructure and Elemental Compositions
2.5. Fracture Surfaces
3. Results
3.1. Mechanical Properties

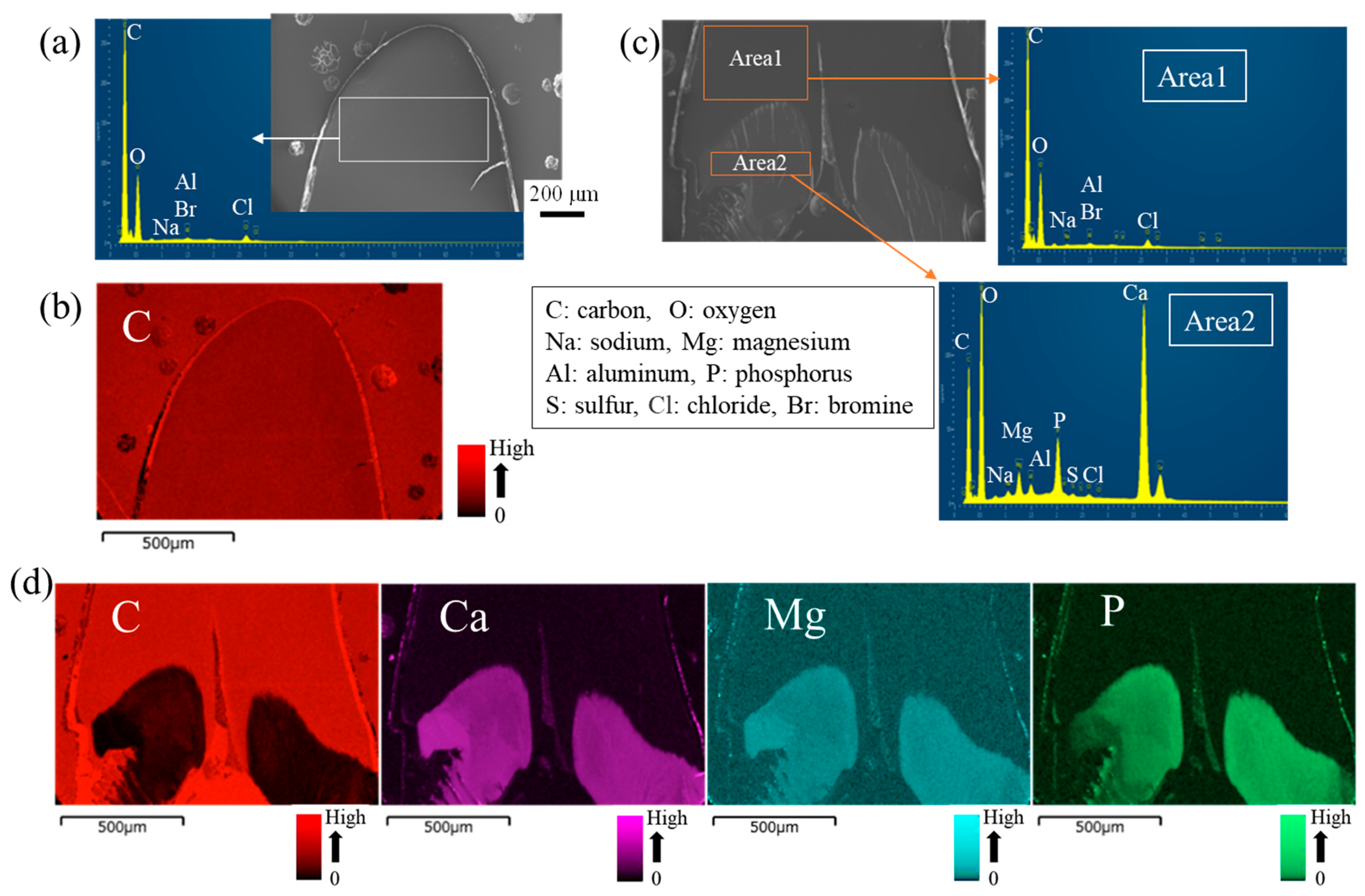
3.2. Crystal Structures and Elemental Compositions
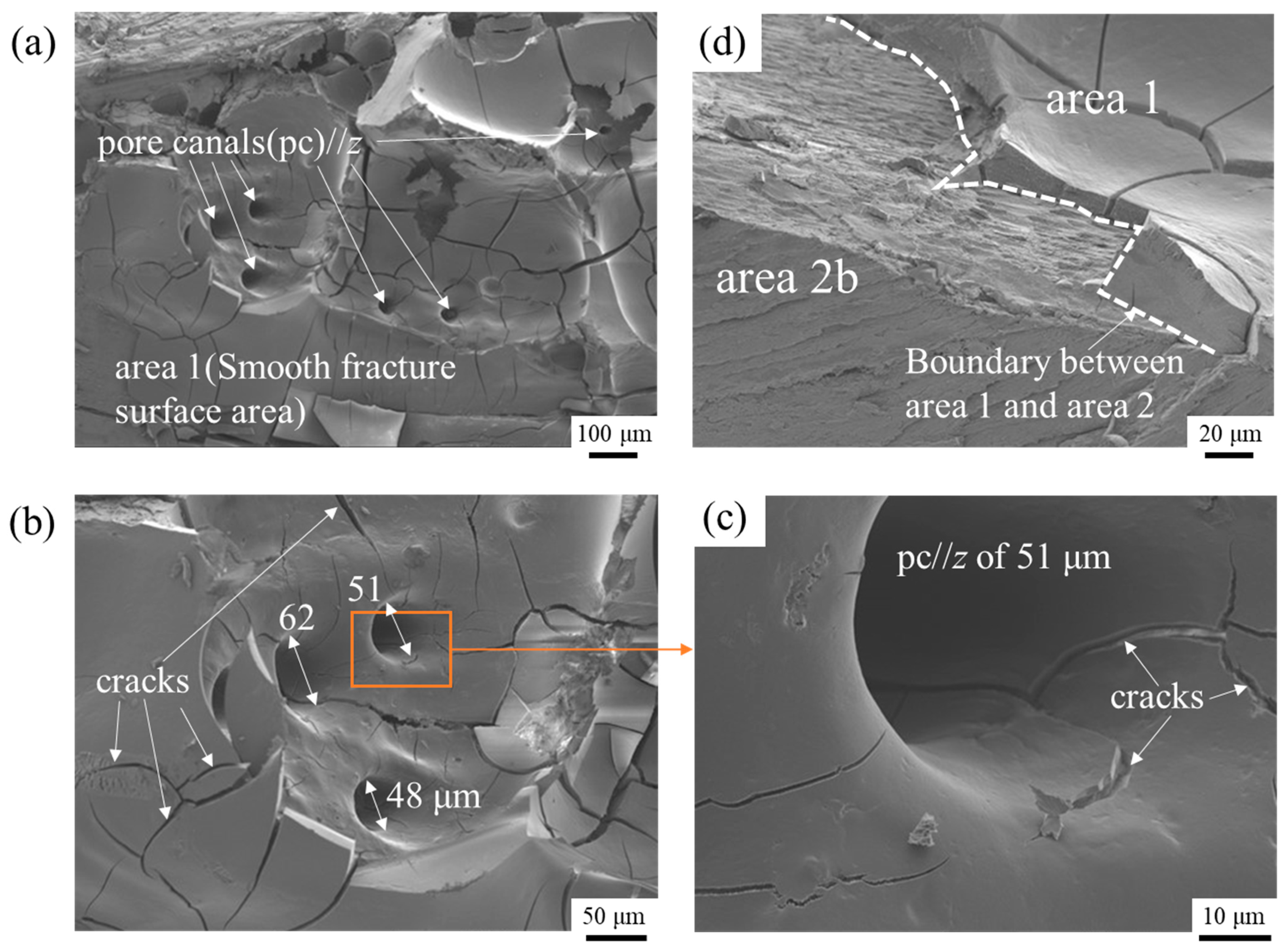
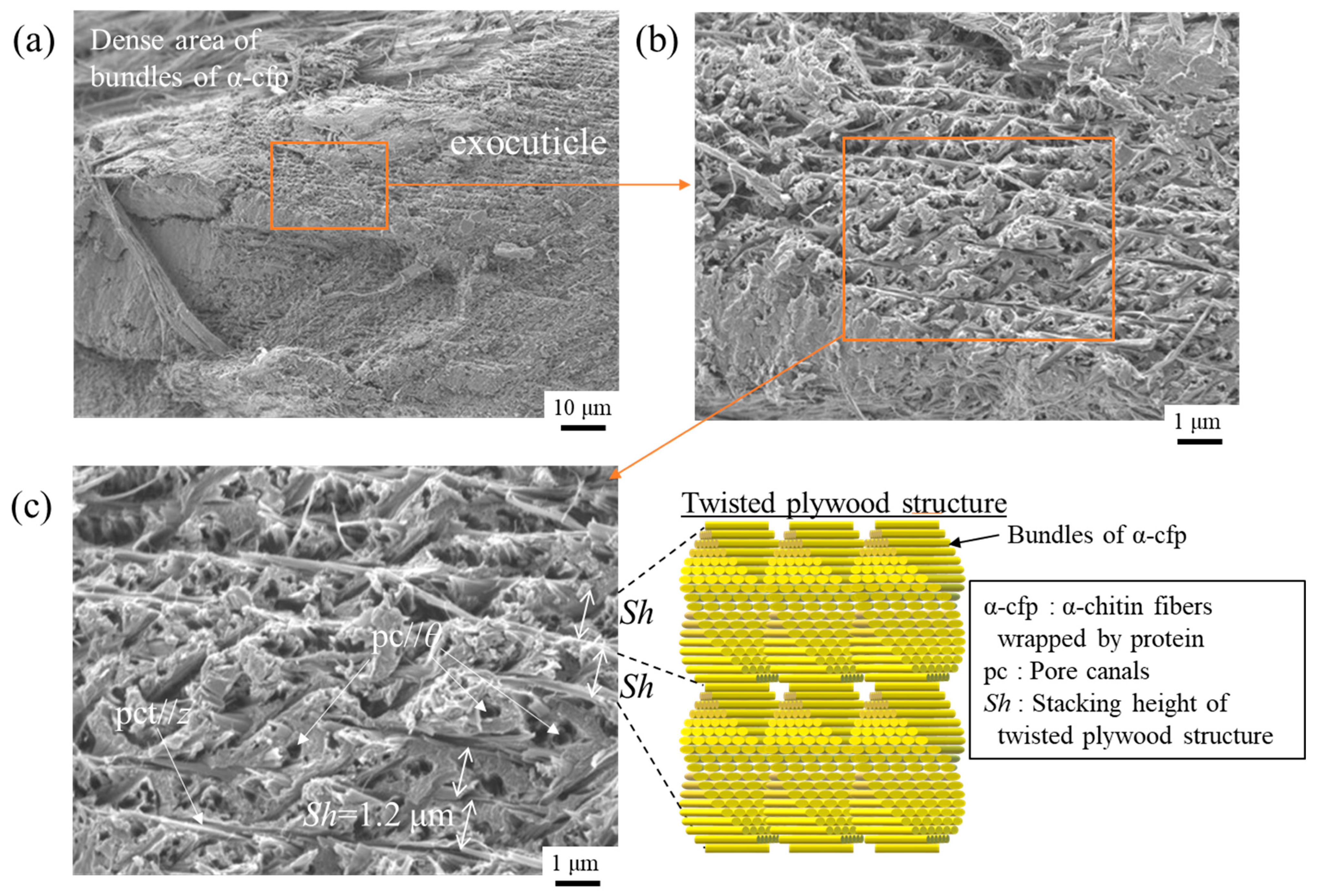
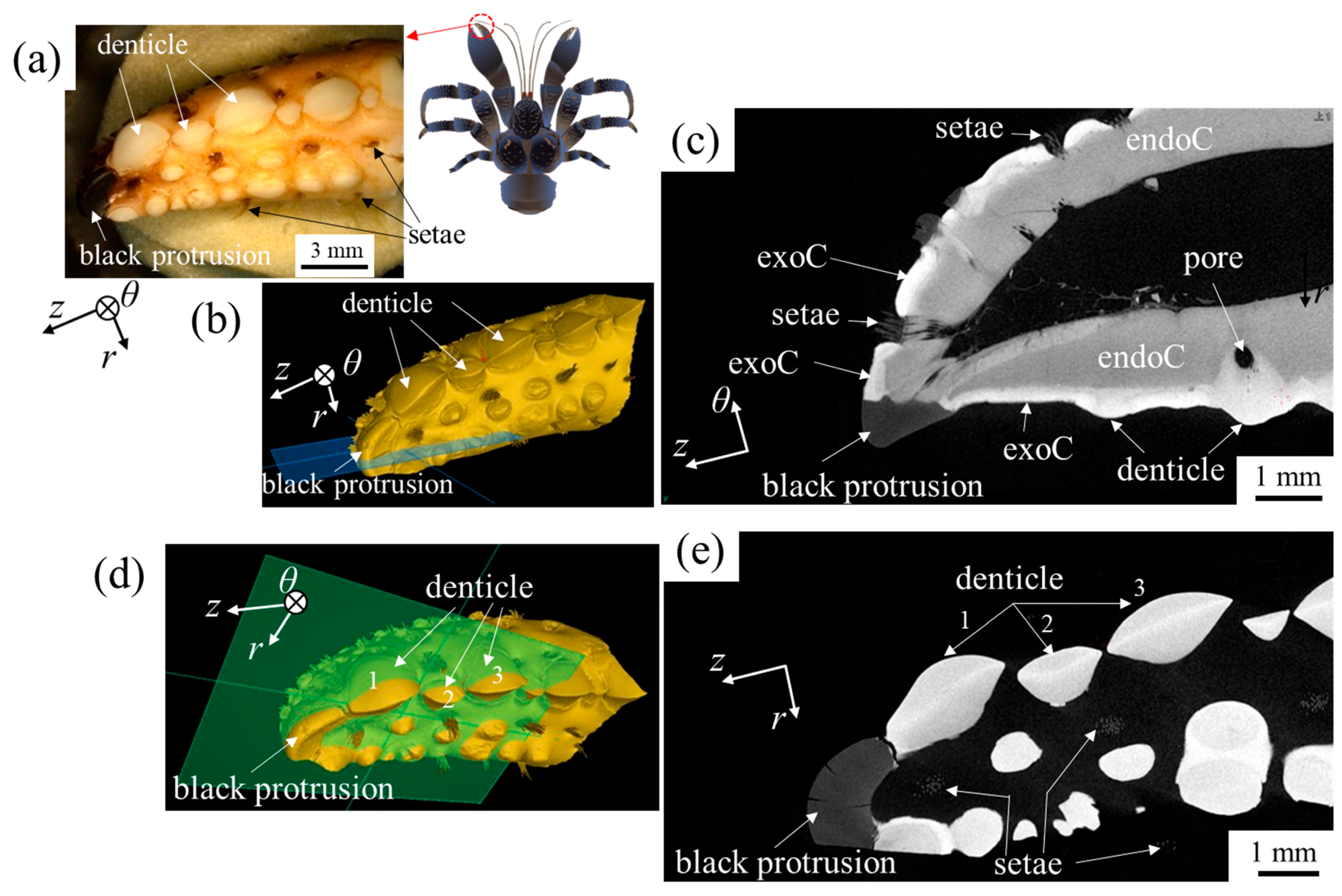
3.3. Microstructures
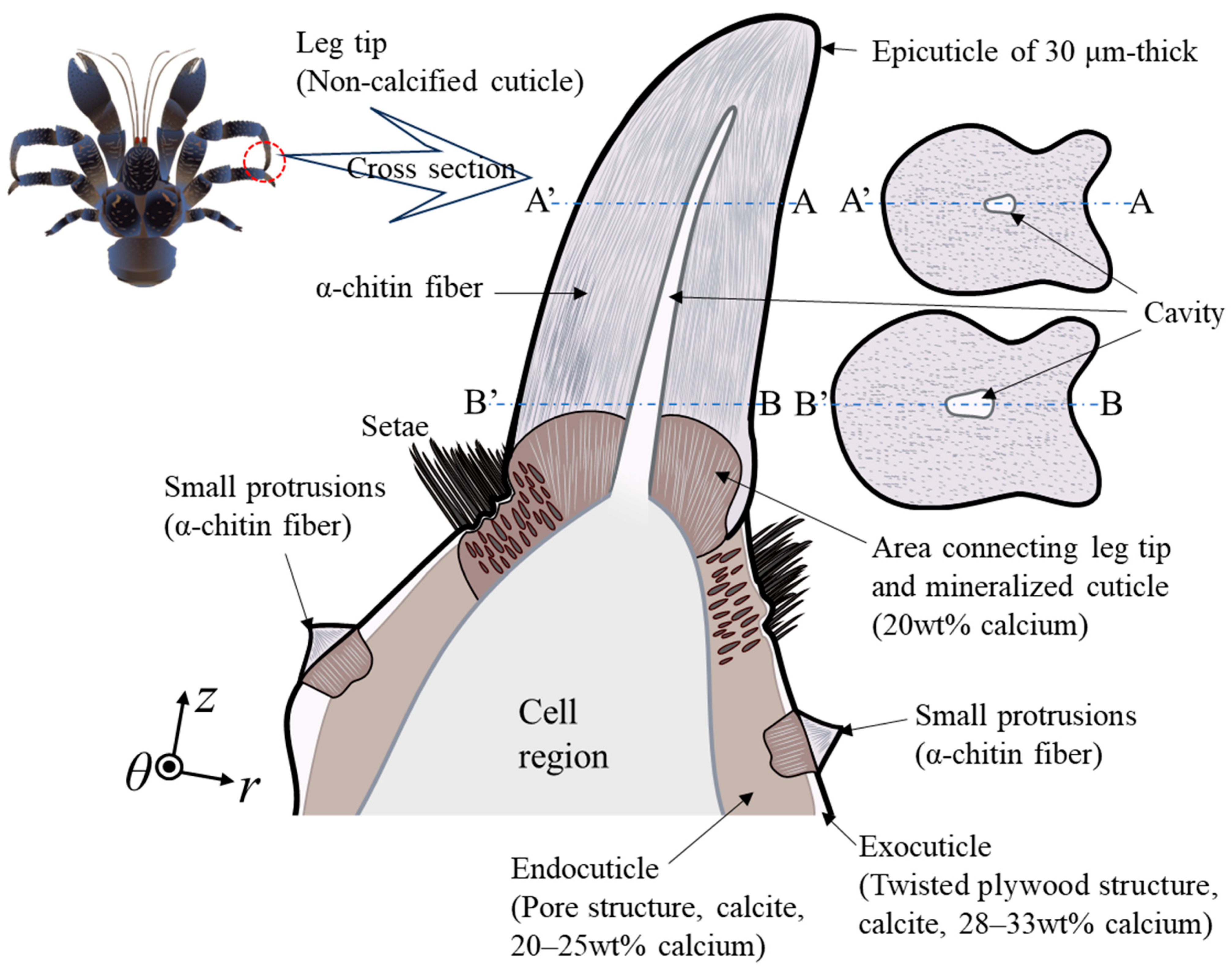
4. Discussion
4.1. The Black Protrusion and Small Black Protrusion
4.2. Role of Black Protrusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bai, L.; Liu, L.; Esquivel, M.; Tardy, B.L.; Huan, S.; Niu, X.; Liu, S.; Yang, G.; Fan, Y.; Rojas, O.J. Nanochitin: Chemistry, structure, assembly, and applications. Chem. Rev. 2022, 122, 11604–11674. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, B.; Gilman, J.W. Bioinspired Bouligand cellulose nanocrystal composites: A review of mechanical properties. Philos. Trans. R. Soc. A 2017, 376, 20170050. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B. Biomimetics: Lessons from nature—An overview. Philos. Trans. R. Soc. A 2009, 367, 1445–1486. [Google Scholar] [CrossRef] [PubMed]
- Amorim, L.; Santos, A.; Nunes, J.P.; Viana, J.C. Bioinspired approaches for toughening of fibre reinforced polymer composites. Mater. Des. 2021, 199, 109336. [Google Scholar] [CrossRef]
- Zhang, B.; Han, Q.; Zhang, J.; Han, Z.; Niu, S.; Ren, L. Advanced bio-inspired structural materials: Local properties determine overall performance. Mater. Today 2020, 41, 177–199. [Google Scholar] [CrossRef]
- Zhou, F.; Wu, Z.; Wang, M.; Chen, K. Structure and mechanical properties of pincers of lobster (Procambarus clarkii) and crab (Eriocheir Sinensis). J. Mech. Behav. Biomed. Mater. 2010, 3, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Bouligand, Y. Twisted fibrous arrangements in biological materials and cholesteric mesophases. Tissue Cell 1972, 4, 189–217. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Song, Z.; Zhang, S.; Ni, Y.; Cai, S.; Gong, X.; He, L.; Yu, S. Discontinuous fibrous Bouligand architecture enabling formidable fracture resistance with crack orientation insensitivity. Proc. Natl. Acad. Sci. USA 2020, 117, 15465–15472. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Pugno, N.M. Bio-mimetic mechanisms of natural hierarchical materials: A review. J. Mech. Behav. Biomed. Mater. 2013, 19, 3–33. [Google Scholar] [CrossRef]
- Yamagata, N.; Randall, G.; Lavoie, E.; Arola, D.; Wang, J. Microstructure, mechanical properties and elemental composition of the terrestrial isopod Armadillidium vulgare cuticle. J. Mech. Behav. Biomed. Mater. 2022, 132, 105299. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, C.; Dai, N.; Shen, J.; Guan, J. Structural characterization and regulation of the mechanical properties of the carapace cuticle in tri-spine horseshoe crab (Tachypleus tridentatus). J. Mech. Behav. Biomed. Mater. 2022, 125, 104954. [Google Scholar] [CrossRef]
- Kundanati, L.; Signetti, S.; Gupta, H.S.; Menegon, M.; Pugno, N.M. Multilayer stag beetle elytra perform better under external loading via nonsymmetric bending properties. J. R. Soc. Interface 2018, 15, 20180427. [Google Scholar] [CrossRef]
- Tan, T.; Ribbans, B. A bioinspired study on the compressive resistance of helicoidal fibre structures. Proc. R. Soc. A 2017, 473, 20170538. [Google Scholar] [CrossRef]
- Drew, M.M.; Harzsch, S.; Stensmyr, M.; Erland, S.; Hansson, B.S. A review of the biology and ecology of the Robber Crab, Birgus latro (Linnaeus, 1767) (Anomura: Coenobitidae). Zoo. Anz. 2010, 249, 45–67. [Google Scholar] [CrossRef]
- Cumberlidge, N. Birgus latro. In The IUCN Red List of Threatened Species 2020; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2020; Available online: https://doi.org/10.2305/IUCN.UK.2020-2.RLTS.T2811A126813586.en (accessed on 4 April 2024).
- Inoue, T.; Oka, S.; Hara, T. Three-dimensional microstructure of robust claw of coconut crab, one of the largest terrestrial crustaceans. Mater. Des. 2021, 206, 109765. [Google Scholar] [CrossRef]
- Oka, T.; Tomita, K.; Miyamoto, A. Mighty Claw: Pinching Force of the Coconut Crab, the Largest Terrestrial Crustacean. PLoS ONE 2016, 11, e0166108. [Google Scholar] [CrossRef]
- Inoue, T.; Hara, T.; Nakazato, K.; Oka, S. Superior mechanical resistance in the exoskeleton of the coconut crab, Birgus latro. Mater. Today Bio 2021, 12, 100132. [Google Scholar] [CrossRef]
- Inoue, T.; Oka, S.; Nakazato, K.; Hara, T. Columnar structure of claw denticles in the coconut crab, Birgus latro. Minerals 2022, 12, 274. [Google Scholar] [CrossRef]
- Inoue, T.; Oka, S.; Nakazato, K.; Hara, T. Structural changes and mechanical resistance of claws and denticles in coconut crabs of different sizes. Biology 2021, 10, 1304. [Google Scholar] [CrossRef]
- Oliver, W.C.; Pharr, G.M. An improved technique for determining hardness and elastic modulus using load and displacement sensing indentation experiments. J. Mater. Res. 1992, 7, 1564–1583. [Google Scholar] [CrossRef]
- Inoue, T.; Hiroto, T.; Hara, Y.; Nakazato, K.; Oka, S. Tissue structure and mechanical properties of the exoskeleton of the huge claws of the mud crab, Scylla serrata. J. Mater. Sci. 2023, 58, 1099–1115. [Google Scholar] [CrossRef]
- Vittori, M.; Srot, V.; Korat, L.; Rejec, M.; Sedmak, P.; Bussmann, B.; Predel, F.; Aken, P.A.; Štrus, J. The mechanical consequences of the interplay of mineral distribution and organic matrix orientation in the claws of the sea slater Ligia pallasii. Minerals 2021, 11, 1373. [Google Scholar] [CrossRef]
- Fabritius, H.-O.; Karsten, E.S.; Balasundaram, K.; Hild, S.; Huemer, K.; Raabe, D. Correlation of structure, composition and local mechanical properties in the dorsal carapace of the edible crab Cancer pagurus. Z. Krist. 2012, 227, 766–776. [Google Scholar] [CrossRef]
- Sachs, C.; Fabritius, H.; Raabe, D. Hardness and elastic properties of dehydrated cuticle from the lobster Homarus americanus obtained by nanoindentation. J. Mater. Res. 2006, 21, 1987–1995. [Google Scholar] [CrossRef]
- Kellersztein, I.; Cohen, S.R.; Bar-On, B.; Wagner, H.D. The exoskeleton of scorpions’ pincers: Structure and micro-mechanical properties. Acta Biomater. 2019, 94, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Cribb, B.W.; Rathmell, A.; Charters, R.; Rasch, R.; Huang, H.; Tibbetts, I.R. Structure, composition and properties of naturally occurring non-calcified crustacean cuticle. Arthropod Struct. Dev. 2009, 38, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.; Dillaman, R. The structure and calcification of the crustacean cuticle. Am. Zool. 1984, 24, 893–909. [Google Scholar] [CrossRef]
- Yano, I. A histochemical study on the exocuticle with respect to its calcification and associated epidermal cells in a shore crab. Bull. Jpn. Soc. Sci. Fish 1972, 38, 733–739. [Google Scholar] [CrossRef]
- Qian, Z.; Yang, M.; Zhou, L.; Liu, J.; Akhtar, R.; Liu, C.; Liu, Y.; Ren, L.; Ren, L. Structure, mechanical properties and surface morphology of the snapping shrimp claw. J. Mater. Sci. 2018, 53, 10666–10678. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, J.; Liu, L.; Zheng, K.; Yu, S.; Fan, Y. Preparation, assessment, and comparison of α-chitin nano-fiber films with different surface charges. Nanoscale Res. Lett. 2015, 10, 226. [Google Scholar] [CrossRef]
- Boßelmann, F.; Romano, P.; Fabritius, H.; Raabe, D.; Epple, M. The composition of the exoskeleton of two crustacea: The American lobster Homarus americanus and the edible crab Cancer pagurus. Thermochim. Acta 2007, 463, 65–68. [Google Scholar] [CrossRef]
- Schofield, R.M.S.; Niedbala, J.C.; Nesson, M.H.; Tao, Y.; Shokes, J.E.; Scott, R.A.; Latimer, M.J. Br-rich tips of calcified crab claws are less hard but more fracture resistant: A comparison of mineralized and heavy-element biological materials. J. Struct. Biol. 2009, 166, 272–287. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Lu, X.; Sun, N.; Rogers, R. Dissolution or extraction of crustacean shells using ionic liquids to obtain high molecular weight purified chitin and direct production of chitin films and fibers. Green Chem. 2010, 12, 968–971. [Google Scholar] [CrossRef]
- Shamshina, J.L.; Gurau, G.; Block, L.E.; Hansen, L.K.; Dingee, C.; Walters, A.; Rogers, R.D. Chitin–calcium alginate composite fibers for wound care dressings spun from ionic liquid solution. J. Mater. Chem. B 2014, 2, 3924–3936. [Google Scholar] [CrossRef] [PubMed]
- Nekvapil, F.; Pinzaru, S.C.; Barbu–Tudoran, L.; Suciu, M.; Glamuzina, B.; Tamas, T.; Chis, V. Color-specific porosity in double pigmented natural 3D-nanoarchitectures of blue crab shell. Sci. Rep. 2020, 10, 3019. [Google Scholar] [CrossRef] [PubMed]
- Altevogt, R.; Davis, T.A. B Birgus latro, India’s monstrous crab. A study and an appeal. Bull. Dep. Mar. Sci. Univ. Cochin 1975, 7, 11–24. [Google Scholar]
- Oka, S.; Matsuzaki, S.; Miyamoto, K. Current status of the coconut crab, Birgus latro (Anomura: Coenobitidae) in Ocean Expo Park, Okinawa-jima Island, southwestern Japan. Biol. Mag. Okinawa 2014, 52, 11–19, (In Japanese with English abstract). [Google Scholar]
- Helfman, G.S. Agonistic behavior of the coconut crab, Birgus latro (L.). Z. Tierpsychol. 1977, 43, 425–438. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, T.; Oka, S.-i.; Hiroto, T. Mechanical Properties, Tissue Structure, and Elemental Composition of the Walking Leg Tips of Coconut Crabs. J. Mar. Sci. Eng. 2024, 12, 639. https://doi.org/10.3390/jmse12040639
Inoue T, Oka S-i, Hiroto T. Mechanical Properties, Tissue Structure, and Elemental Composition of the Walking Leg Tips of Coconut Crabs. Journal of Marine Science and Engineering. 2024; 12(4):639. https://doi.org/10.3390/jmse12040639
Chicago/Turabian StyleInoue, Tadanobu, Shin-ichiro Oka, and Takanobu Hiroto. 2024. "Mechanical Properties, Tissue Structure, and Elemental Composition of the Walking Leg Tips of Coconut Crabs" Journal of Marine Science and Engineering 12, no. 4: 639. https://doi.org/10.3390/jmse12040639
APA StyleInoue, T., Oka, S.-i., & Hiroto, T. (2024). Mechanical Properties, Tissue Structure, and Elemental Composition of the Walking Leg Tips of Coconut Crabs. Journal of Marine Science and Engineering, 12(4), 639. https://doi.org/10.3390/jmse12040639