Abstract
Agar oligosaccharides from the degradation of agar harbor great potential in the food and pharmaceutical industries. An agar-degrading bacterium, Vibrio natriegens WPAGA4, was isolated from the deep sea in our previous work. However, the agar-degrading activity of WPAGA4 remains to be improved for more production benefits of this strain. The aim of this study was to enhance the agar-degrading activity of WPAGA4 by using atmospheric and room temperature plasma (ARTP) mutagenesis. Three mutant strains, including T1, T2, and T3, with good genetic stability were obtained, and the agar-degrading activities of these strains increased by 136%, 141%, and 135%, respectively. The optimal temperature and pH for agar degradation were slightly changed in the mutant strains. No sequence mutation was detected in all the agarase genes of WPAGA4, including agaW3418, agaW3419, agaW3420, and agaW3472. However, ARPT mutagenesis increased the relative expression levels of agaW3418, agaW3419, and agaW3420 in the mutant strains, which could be the reason for the improvement of degradation activities in the mutant strains. Furthermore, T3 had the lowest consumption rate of agar oligosaccharide, which was 21% less than the wild-type strain. Therefore, T3 possessed a preferable production value due to its higher degrading activity and lower consumption of agar oligosaccharides. The current work enhanced the agar-degrading activity of WPAGA4 and offered strains with greater potential for agar oligosaccharide production, thereby laying the foundation for industrial applications.
1. Introduction
Agar is one of the world’s three major seaweed polysaccharides, which is mainly extracted from red algae, such as Gracilaria sp. and Gelidium sp. [1]. The application of agar in food and other industries is greatly limited because of its high viscosity and insolubility at room temperature [2]. The oligosaccharides from agar degradation with a low molecular weight, better water solubility, and high absorbency [3] have biological activities such as anti-inflammation [4], antitumor [5], whitening [6], and anti-obesity [7], which offers agar oligosaccharides a wide range of applications in beauty, food, medicine, and other industries [8]. Microbial hydrolysis is a low-pollution and environmentally friendly method for agar degradation compared to chemical degradation methods [3]; therefore, it is receiving more and more attention for agar degradation and oligosaccharide preparation.
Atmospheric and room temperature plasma (ARTP) is a novel, efficient, and safe genome-wide mutagenesis tool based on plasma jets that alters the structure and permeability of cell walls and membranes and could cause DNA damage in microorganisms [9]. ARTP mutagenesis can be operated flexibly at atmospheric pressure and room temperature (25–40 °C), which is simpler and more economical than conventional physico-chemical mutagenesis while keeping the operator safe and the environment pollution-free [10,11]. Recently, the ARTP mutagenesis technique has been applied to a number of different microorganisms to improve their enzyme activity [12], such as protease [13], amylase [14], xylanase [15], cellulase [16], and phospholipase D [17].
In a previous study, we isolated a bacterium, Vibrio natriegens WPAGA4, from deep-sea sediments. Four agarase genes, namely agaW3418, agaW3419, agaW3420, and agaW3472, were found through genome sequencing and enzyme activity determination, which proved that this bacterium has the potential to be applied in agar degradation [18,19,20]. However, the enzyme activity of the bacteria remains to be improved to further lower the cost of agar degradation and oligosaccharide preparation. Therefore, the present study aims to improve the activity of agar degradation of WPAGA4 by using ARTP technology, thereby laying the foundation for in-depth investigation of agar degradation and providing efficient and stable germplasm resources for the industrial production of agar oligosaccharide.
2. Materials and Methods
2.1. Strain, Medium, and Growth Conditions
The bacterium V. natriegens WPAGA4 was isolated from the deep-sea sediment in the Western Pacific (157°24′31″ E, 19°30′30″ N) at a depth of 5378 m in our previous work [20]. The strain was stored in an ultra-cold storage freezer at −80 °C in our laboratory.
The 2216E liquid medium (pH 7.4) contained the following components: 5.0 g peptone, 1.0 g yeast extract, 0.01 g iron phosphate, and 1 L seawater. The agar medium was made by adding 20 g of agar to 1 L 2216E liquid medium. All media used in this study were sterilized at 121 °C for 30 min.
To determine the optimized temperature for growth, the strain was cultured in liquid 2216E medium at 180 rpm at various temperatures (25, 28, 31, 34, and 37 °C) for 12 h. The absorbance of culture broth was measured by using a spectrophotometer at a wavelength of 600 nm (OD600). To determine the growth curve, the strain was cultured in liquid 2216E medium at 180 rpm at 28 °C for 0, 2, 4, 6, 8, 10, 12, 14, 16, 18, 24, 30, 36, and 48 h, and the OD600 of the broth was measured.
A total of 150 μL of culture broth with an OD600 value of 0.8 was spread on the solid 2216E plate and was incubated at 28 °C for 12 h. The number of colony-forming units (CFUs) per mL was counted after the incubation.
2.2. Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis
The strains were inoculated into 2216E liquid medium with 1% inoculum volume (v/v) and were cultured in a shaker (180 rpm) at 28 °C for 12 h. After that, 1 mL culture solution was sampled and centrifuged at 4266× g for 10 min at 4 °C, and the bacteria precipitates were resuspended with 5% sterilized glycerin (v/v). The suspended bacterial solution was uniformly coated on the sterile metal slide and placed in the ARTP mutator apparatus (Wuxi Yuanqing Tianmu Biological Technology Co., Ltd., Wuxi, China) for a mutation test (120 W, 10 SML). The exposure time under the plasma jet was set as 0, 5, 10, 15, and 20 s. After mutagenesis, the slide was placed back into a 1.5 mL centrifuge tube, and the bacteria were suspended by swirling oscillation for 1 min. A total of 100 µL treated bacterial suspension was coated on the 2216E solid medium, and the optimal induction intensity was calculated according to the fatality rate. The fatality rate is calculated as follows:
where Nc and Ns are the colony numbers of the untreated and treated samples by using ARTP mutagenesis, respectively.
2.3. Positive Clone Screening
The bacterial solution after ARTP treatment was spread on a fresh 2216E plate and was cultured at 28 °C for 24 h. The clear zones around the colonies were observed after pooling Lugo’s iodine solution on the plates. The agar degradation abilities of the mutated colonies were initially assayed based on the degradation zone, which is defined as the rates of the diameters of transparent zones to the colony. The colonies with higher rate values compared to the wild-type strain were selected and inoculated into 2216E liquid medium for further enzyme activity measurement to confirm the increase in the agar degradation activities of the mutant strains compared to the wild one.
2.4. Measurement of Enzymatic Activity
The selected strains were inoculated into 2216E liquid medium and cultured at 180 rpm and 28 °C until the OD600 values reached 0.8 and the number of mutant strains was the same as that of the wild-type strain by using a microplate reader (Biotek, Winooski, VT, USA). The supernatant was sampled to determine the agar degradation activity by centrifuging at 4266× g and 10 °C for 10 min. The determination of the degradation activity of the fermentation broth was performed by the 3,5-dinitrosalicylic acid (DNS) method [21] as follows: a total of 25 μL of supernatant was mixed with 225 μL of 0.4% agarose solution (w/v), and the mixture was incubated at a certain temperature for 50 min. Subsequently, 750 μL of DNS solution was added into the mixture and was heated at 100 °C for 10 min, and the absorbance value of the reaction solution was measured at OD550. The enzyme activity unit (U) was defined as the amount of enzyme required to hydrolyze agarose to produce 1 μmol of reducing sugars per minute. The activities of the mutant strains were statistically compared with the wild-type strain by using the t-test.
2.5. Measurement of Optimal Degradation Conditions
To determine the optimal temperature and pH of the mutant and the wild-type strains, the supernatant was reacted with 0.4% (w/v) agarose solution at various temperatures (0, 10, 20, 30, 40, 50, 60, and 70 °C) and pHs (pH 3.0–8.0, 50 mmol/L citrate buffer; pH 8.0–9.0, 50 mmol/L Tris-HCl buffer; pH 10.0–10.6, 50 mmol/L glycine-NaOH buffer), and then the enzyme activity was measured based on the method in Section 2.4. The maximum agar degradation activity in the experimental group was set to 100%.
The effects of the metal ions and chemical reagents on the degradation activity were measured by using the method in Section 2.4 with the metal ions (MgCl2, CuSO4, CaCl2, NaCl, ZnSO4, MnSO4, FeSO4, NH4Fe(SO4)2, and KCl) and chemical reagents (EDTA, SDS, β-mercaptoethanol, and DTT) at a final concentration of 1 mmol/L. The control group without any metal ions or chemical reagents was set to 100%.
All experiments in this section were performed with three biological replicates.
2.6. Stability of the Mutant Strains
The mutant strains were cultured in 2216E liquid medium at 180 rpm and 28 °C until the OD600 value reached 0.8. Next, the culture was transferred to fresh 2216E liquid medium at 1% inoculum volume (v/v) and cultured in a shaker under the same conditions for five generations. The enzyme activity of each generation was measured by using the method in Section 2.4. The stable activity of the mutants was evaluated by comparing the activities across the generations.
2.7. Genome Sequencing and Insertion-Deletion (InDel) Analysis
The genome sequences of mutant strains were sequenced by Wuhan Benagen Technology Company (Wuhan, China) using the platforms of Nanopore GridION (Oxford Nanopore Technologies, Oxford, UK) and Illumina Hiseq (Illumina, California, CA, USA). The genome sequences of mutant strains were aligned against the wild-type strain by using Nucmer (version 3.1, default parameters), and the SNP information on the genome alignment sequences was output by using the tool show-snps (parameter-C: only the uniquely matched SNPs in the alignment results) in Mummer (version 3.23). Finally, we annotate these SNPs through Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) and Cluster of Orthologous Groups (COG).
2.8. Neoagarooligosaccharide (NAOS) Consumption in Degrading Strains
The wild-type and mutant strains were inoculated into 2216E liquid medium (1% inoculum size) with additional NAOS at a final concentration of 0.5 g/L and were cultured at 28 °C and 180 rpm. A total of 1 mL broth was sampled at 0, 3, 6, 9, and 12 h and was centrifuged for 15 min at 4266× g and 4 °C for sampling the supernatant to determine the remaining NAOS concentrations in the fermentation broth by using the DNS method outlined in Section 2.4.
2.9. Real-Time Quantitative PCR (RT-qPCR)
The strain was cultured in liquid 2216E medium with 0.4% agarose at 180 rpm and 28 °C for 14 h, and the total RNA was extracted by using RNAprep Pure Cell/Bacteria Kit (Transgen, Beijing, China). The RNA concentration was determined by using NanoDrop ONE (ThermFisher, Waltham, MA, USA). The total RNA was reversely transcribed into cDNA by using Applied Biosystems 7500 Fast Real-Time PCR Systemy (ThermFisher) by strictly following the instructions of Takara PrimeScript™ RT reagent kit (Takara, Kusatsu, Japan). We chose the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) genes as the housekeeping genes. The primers used for RT-qPCR are listed in Table 1. Relative gene expression was calculated based on the expression level of housekeeping genes GAPDH: ΔCT (Target) = CT (Target) − CT (GAPDH), ΔCT (control) = CT (control) − CT (GAPDH), ΔΔCT = ΔCT (Target) − ΔCT (control), gene expression = 2−ΔΔCT. All experiments were repeated three times.

Table 1.
Primer sequences.
2.10. Heterologous Expression and Purification of the Agarase Genes
The genes agaW3418, agaW3419, agaW3420, and agaW3472 were expressed in E. coli BL21(DE3) cells using a pEAZY®-Blunt E2 expression vector (TransGen Biotech., Beijing, China). The recombinant protein was obtained by ultrasonication and then purified by Ni-nitrilotriacetic acid (NTA) Sefinose™ Resin (Tiangen Biotech, Beijing, China). The molecular weights and purities of recombinant AgaW3418, AgaW3419, AgaW3420, and AgaW3472 were subsequently estimated using SDS-PAGE and Coomassie brilliant blue staining. The recombinant proteins were mixed according to their proportions in RT-qPCR results.
3. Results
3.1. Lethality Rate of WPAGA4 after ARTP Treatment
The rejuvenated strain that was cultured at 28 °C for 12 h was selected for further ARTP mutagenesis according to the results of optimal temperature (Figure 1A) and growth curve (Figure 1B) analyses. A lethality rate of 95% was chosen as the threshold for obtaining a maximum frequency of mutation based on former work [17,22]. The result showed that a lethality rate of 95% of the WPAGA4 strain required an exposure time of 15 s (Figure 1C). Therefore, 15 s was selected as the most appropriate time for ARTP treatment in this study.
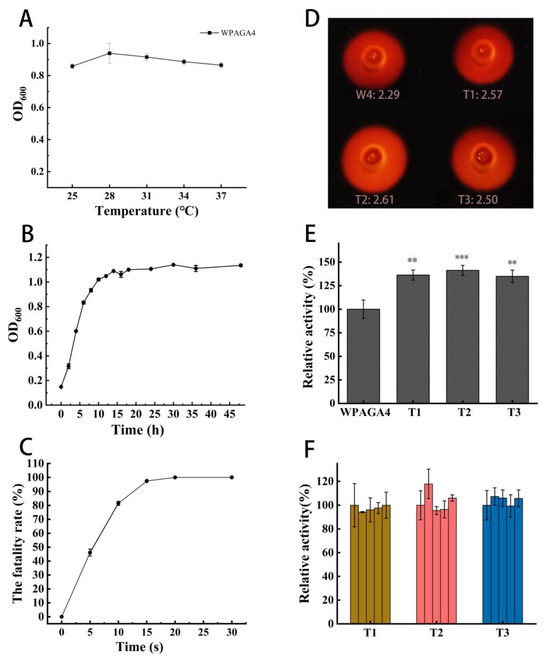
Figure 1.
The growth and the agar-degrading activity analyses of WPAGA4. (A) The effect of temperature on the growth of WPAGA4; (B) the growth curve of WPAGA4; (C) the effect of ARTP treatment time on the lethality rate of WPAGA4; (D) the clean zones of the wild-type (W4) and mutant strains (T1, T2, and T3). The numbers following the strain names in the figure are the diameter ratios of transparent circles/colonies; (E) the relative agar degradation activities of WPAGA4, T1, T2, and T3. The symbols “**” and “***” represent the p values lower than 0.05 and 0.01, respectively; (F) the genetic stability of mutants T1, T2, and T3 for degradation activity increase.
3.2. Screening and Verification of the Mutants
Three mutant strains, including T1, T2, and T3, had larger diameter ratios of transparent circles/colonies than the wild-type strain (Figure 1D). The CFUs per mL of mutant strains at OD600 of 0.8 had no significant difference from that of the wild-type strain (t-test; p > 0.05; Table S1). The growth curves of the mutant strains were not obviously changed (Figure S1). Furthermore, the enzyme activities of the T1, T2, and T3 strains in the broth were determined. The results showed that the relative enzyme activities of T1, T2, and T3 strains in the broth significantly increased to 136%, 141%, and 135% compared to the wild-type strain, respectively (t-test; p < 0.05) (Figure 1E). After five-generation cultivation, the agar-degrading activities of the three mutant strains were maintained (Figure 1F) and were still significantly higher than the wild-type strain (t test; p < 0.05; Table S2), indicating high genetic stability.
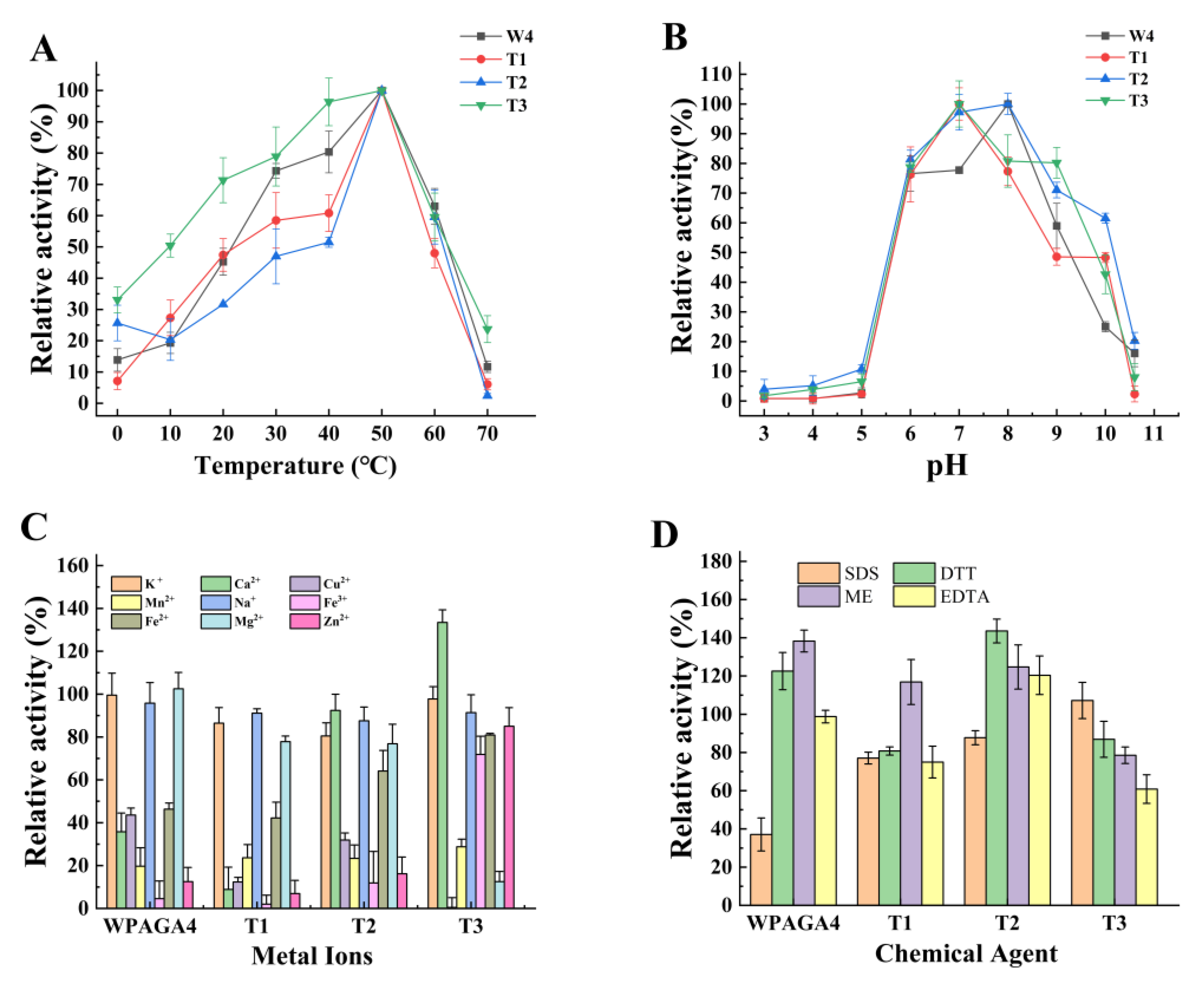
3.3. Enzymatic Properties of WPAGA4, T1, T2, and T3
The optimal temperature of the three mutant strains was the same as the wild-type strain (50 °C; Figure 2A). Compared with the wild-type strain, the optimal pH of T1 and T3 decreased to 7 (Figure 2B). The activities of the T2 and T3 strains increased with Ca2+, and the activities of the T3 strain with Fe3+, Fe2+, and Zn2+ were higher than the other strains (Figure 2C). The tolerance of T1, T2, and T3 strains to SDS was improved compared with the wild-type strain. Moreover, DTT and EDTA had inhibitory effects on the activities of T1 and T3, and EDTA had a promoting effect on the activity of T2 (Figure 2D).
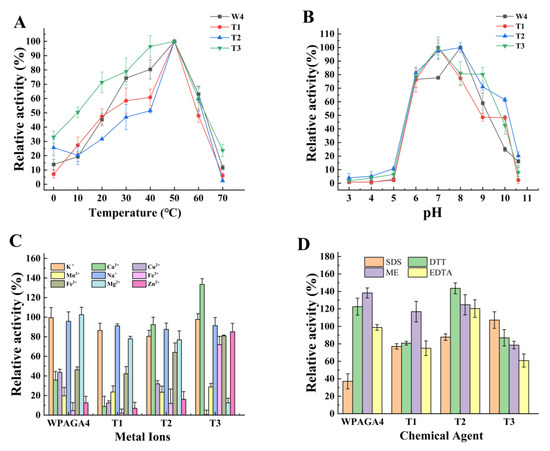
Figure 2.
The effects of temperature (A), pH (B), metal ions (C), and chemical agents (D) on the agar degradation activity of WPAGA4, T1, T2, and T3. SDS, sodium dodecyl sulfate; DTT, dithiothreitol; ME, β-mercaptoethanol; EDTA, ethylenediaminetetraacetic acid.
3.4. Genome Sequencing and Comparative Genomic Analysis
The genomes of the three mutant and wild-type strains were sequenced to further explain the mechanisms of agar degradation activity increase in mutant strains. A total of 24, 25, and 24 indels was found in the genomes of strains T1, T2, and T3, respectively. Moreover, 151, 155, and 180 SNPs were detected in the genomes of strains T1, T2, and T3, respectively (Table 2). However, no sequence mutation was discovered in agaW3418, agaW3419, agaW3420, and agaW3472 genes in all mutant strains.
Table 2.
The indels and SNPs.
3.5. Real-Time qPCR Analysis
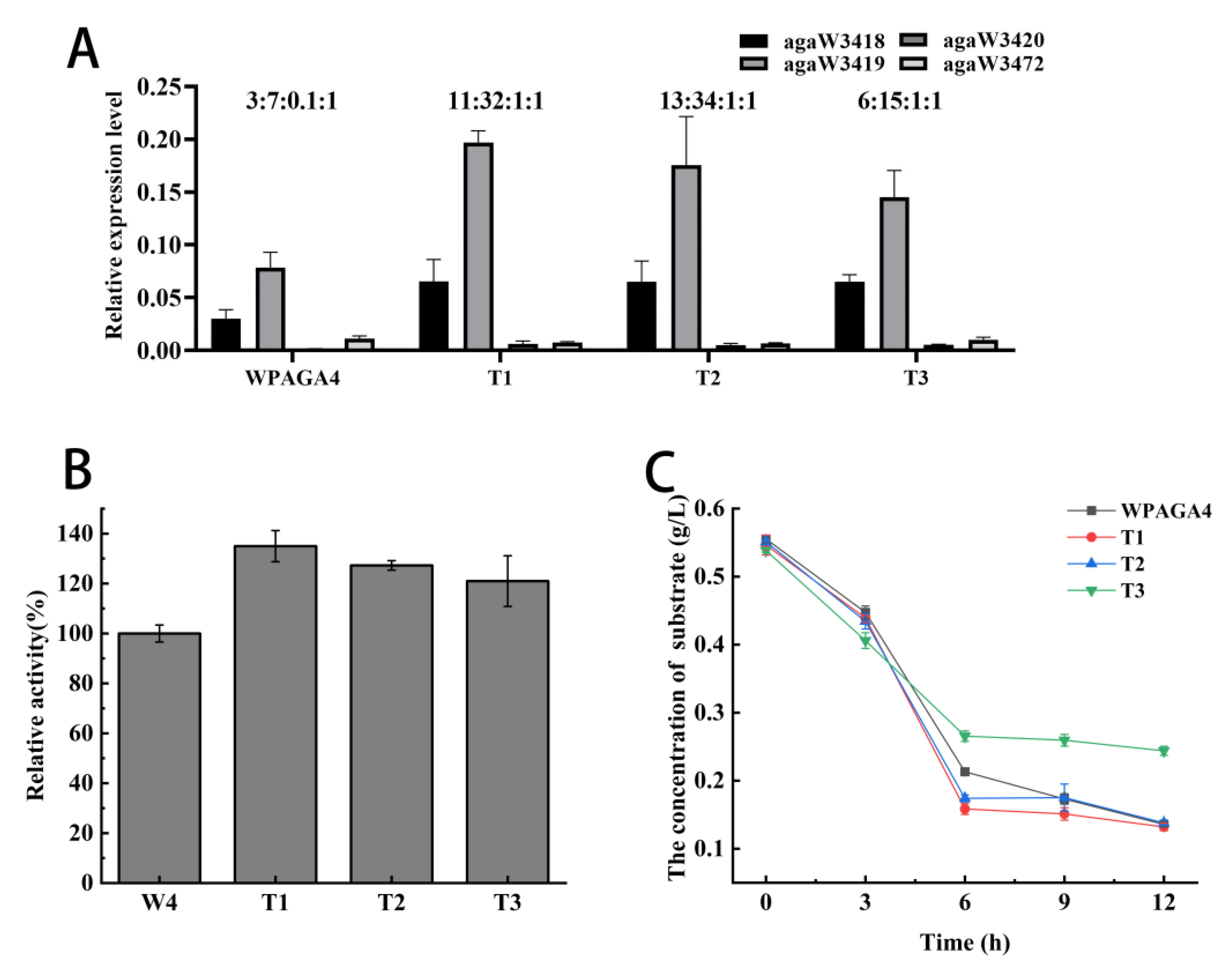
The ratio of gene expression levels of agaW3418, agaW3419, agaW3420, and agaW3472 in the wild-type strain was approximately 3:7:0.1:1. In the three mutant strains, the expression ratios of these four genes were approximately 11:32:1:1 (T1), 13:34:1:1 (T2), and 6:15:1:1 (T3), respectively (Figure 3A). Therefore, almost all the expression levels of the genes agaW3418, agaW3419, and agaW3420 were greatly increased in mutant strains. In detail, the expression level of agaW3418 in T1, T2, and T3 increased by 117%, 116%, and 115%, respectively. The expression level of agaW3419 in T1 and T2 increased by 151% and 124%, respectively; however, that of T3 decreased by 85%. The expression level of agaW3420 in T1, T2, and T3 increased by 304%, 235%, and 248%, respectively (Figure 3A).
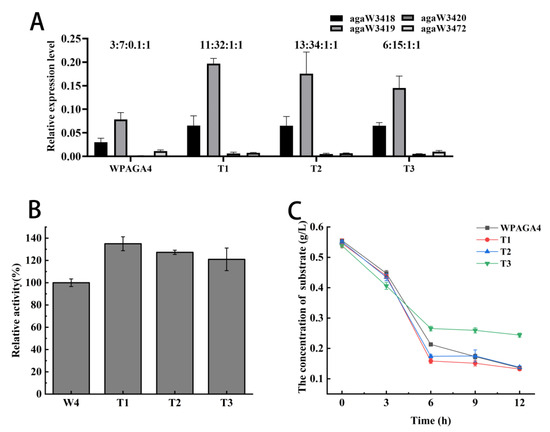
Figure 3.
(A) Analysis of gene expression levels of four agarases in the wild-type and mutant strains; (B) the relative activity of agarase mixtures with various mole ratios; (C) the effect of culture time on the consumption of oligosaccharides in the medium.
3.6. The Degradation Activities of the Agarase Mixtures with Different Mole Ratios
The four agarases were mixed based on the mole ratios according to the relative expression level proportions in strains T1, T2, and T3 in RT-qPCR results, and then the agar degradation activities of enzyme mixtures were measured to verify the effect of enzyme proportion on agar degradation. The result showed that the degradation activities of the agarase mixture with the mole ratios of T1, T2, and T3 increased to 135%, 127%, and 121%, respectively (Figure 3B).
3.7. The Determination of Agar Oligosaccharide Substrate Consumption
The oligosaccharide concentrations in the medium decreased with time by bacterial consumption before 6 h. The consumption of oligosaccharides in the mutant strains T1, T2, T3, and the wild-type strain were 71%, 68%, 51%, and 62% at 6 h, respectively. The consumption of oligosaccharides by T1 and T2 was higher than that of the wild-type strain. After 6 h, there was no obvious change in oligosaccharides consumption. And by 12 h, the oligosaccharides consumed by mutant strains T1, T2, T3, and the wild-type strain were 76%, 75%, 55%, and 76%, respectively. Therefore, the oligosaccharide consumption of strain T3 was the lowest among the strains (Figure 3C).
4. Discussion
In this study, we applied ARTP technology to WPAGA4 and obtained three mutants—T1, T2, and T3—with increased agar degradation activity to 136%, 141%, and 135%, respectively. The higher agar degradation activity is conducive to increasing the yield of agar-oligosaccharides, reducing the production cost of agar-oligosaccharides, and providing a basis for their application in food, cosmetics, and other industries.
The oligosaccharides from agar degradation can be further utilized by the degrading strains [23], thereby resulting in product consumption and increasing the cost of oligosaccharide preparation. The mutant strain T3 showed a 21% reduction in oligosaccharide consumption compared to the wild-type strain. This reduction in oligosaccharide consumption enhances the application values of T3 in oligosaccharide production.
The ARTP technique has been successfully applied to improve the enzyme activity of bacteria, fungi, and microalgae, including xylanase [15], cellulase [16], protease, and amylase [13]. It represents a novel and effective breeding strategy. As far as we know, this is the first performance of ARTP technology for increasing agar-degrading activity. In addition, the ARTP/EMS-combined mutagenesis method was used to improve starch-degrading enzyme production [24]. Therefore, the agar-degrading activity of WPAGA4 could be further improved by using co-mutagenesis or multiple rounds of mutagenesis in future experiments.
In this study, Cu2+ and Mn2+ strongly inhibited the agar-degrading activities of the three mutant strains and the wild-type strain. Therefore, both Cu2+ and Mn2+ should be avoided as much as possible in industrial production, which could improve the production efficiency of oligosaccharides.
Furthermore, this work investigated the underlying cause of the increased degrading activity. Initially, we hypothesized that mutations in the agarase sequences were responsible for this phenomenon. However, genome sequencing revealed that there were no mutations in the sequences of the four agarase genes. Subsequently, we employed RT-qPCR to determine the expression levels of the four agarase genes in the three mutant strains and detected the obvious increasing expression levels of three agarase genes therein. Therefore, we speculate that the increasing expression levels of the agarase genes were the possible reason for the improved agar-degrading activity of the mutant strains. Subsequently, the activity improvement of the agarase mixture with the mole ratios based on the RT-qPCR result verified our speculation.
The increasing expression levels of agarase genes resulted in a changed ratio of the expression levels of four agarase genes in the mutant strains compared to the wild one. Previous studies have demonstrated that cellulose degradation requires mixed cellulases, and the proportion of enzyme amounts in the mixture affects their synergistic effect and hydrolysis efficiency [25]. The expression levels of agarase genes not only increased but also showed a similar expression ratio for four agarases in the mutant strains. Therefore, an appropriate expression level ratio of agarases is also of value for efficient agar degradation in addition to the high expression levels of agarase genes.
5. Conclusions
Three mutant strains with increased activity were obtained based on the deep-sea bacterium V. natriegens WPAGA4 by using ARTP technology in this study. Their biochemical properties for agar degradation slightly varied compared to the wild WPAGA4 strain. However, the gene sequences of agarases in the mutant strains were not changed. The enhanced activity was attributed to the increasing expression levels of agarase genes and appropriate expression level ratios in the mutant strains. The current study provides efficient and stable germplasm resources for agar degradation and agar-oligosaccharide industrial production.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jmse12071154/s1. Table S1. The colony forming units (CFUs) per mL of the wild-type and mutant strains at the OD600 value of 0.8; Table S2. The agar-degrading activities of the mutant strains across generations; Figure S1. Growth curves of the mutant strains of T1, T2, and T3.
Author Contributions
Conceptualization, W.Q.; methodology, X.T., X.L., S.F., M.Z. and W.Q.; validation, J.W. and W.Q.; investigation, X.T., X.L., J.W. and W.Q.; data curation, X.T., X.L. and S.F.; writing—original draft preparation, X.T. and X.L.; writing—review and editing, J.W., S.F. and W.Q.; visualization, X.T. and S.F.; funding acquisition, J.W. and W.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the “Pioneer” and “Leading Goose” R&D Program of Zhejiang [grant number 2022C02040], the Science and Technology Program of Zhoushan [grant number 2022C13048], and the Technology Innovation Center for Exploitation of Marine Biological Resources, MNR [grant number TICMBR202406].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, W.-K.; Lim, Y.-Y.; Leow, A.T.-C.; Namasivayam, P.; Abdullah, J.O.; Ho, C.-L. Biosynthesis of agar in red seaweeds: A review. Carbohydr. Polym. 2017, 164, 23–30. [Google Scholar] [CrossRef]
- Mostafavi, F.S.; Zaeim, D. Agar-based edible films for food packaging applications—A review. Int. J. Biol. Macromol. 2020, 159, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Xiaodan, C.; Xiaoting, F.; Luqiang, H.; Jiachao, X.; Xin, G. Agar oligosaccharides: A review of preparation, structures, bioactivities and application. Carbohydr. Polym. 2021, 265, 118076. [Google Scholar]
- Wang, W.; Liu, P.; Hao, C.; Wu, L.; Wan, W.; Mao, X. Neoagaro-oligosaccharide monomers inhibit inflammation in LPS-stimulated macrophages through suppression of MAPK and NF-κB pathways. Sci. Rep. 2017, 7, 44252. [Google Scholar] [CrossRef]
- Enoki, T.; Tominaga, T.; Takashima, F.; Ohnogi, H.; Sagawa, H.; Kato, I. Anti-tumor-Promoting Activities of Agaro-Oligosaccharides on Two-Stage Mouse Skin Carcinogenesis. Biol. Pharm. Bull. 2012, 35, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Jang, M.K.; Lee, O.H.; Kim, N.Y.; Ju, S.A.; Lee, S.H. Over-production of a glycoside hydrolase family 50 beta-agarase from Agarivorans sp. JA-1 in Bacillus subtilis and the whitening effect of its product. Biotechnol. Lett. 2008, 30, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.J.; Lee, J.-H.; Kim, E.J.; Yang, H.J.; Park, J.-S.; Hong, S.-K.; Long, P. Anti-Obesity and Anti-Diabetic Effect of Neoagarooligosaccharides on High-Fat Diet-Induced Obesity in Mice. Mar. Drugs 2017, 15, 90. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lee, C.R.; Hong, S.K. Implications of agar and agarase in industrial applications of sustainable marine biomass. Appl. Microbiol. Biotechnol. 2020, 104, 2815–2832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, C.; Zhou, Q.Q.; Zhang, X.F.; Wang, L.Y.; Chang, H.B.; Li, H.P.; Oda, Y.; Xing, X.H. Quantitative evaluation of DNA damage and mutation rate by atmospheric and room-temperature plasma (ARTP) and conventional mutagenesis. Appl. Microbiol. Biotechnol. 2015, 99, 5639–5646. [Google Scholar] [CrossRef]
- Zou, R.-S.; Li, S.; Zhang, L.-L.; Zhang, C.; Han, Y.-J.; Gao, G.; Sun, X.; Gong, X. Mutagenesis of Rhodobacter sphaeroides using atmospheric and room temperature plasma treatment for efficient production of coenzyme Q10. J. Biosci. Bioeng. 2019, 127, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-P.; Wang, Z.-B.; Ge, N.; Le, P.-S.; Wu, H.; Lu, Y.; Wang, L.-Y.; Zhang, C.; Bao, C.-Y.; Xing, X.-H. Studies on the Physical Characteristics of the Radio-Frequency Atmospheric-Pressure Glow Discharge Plasmas for the Genome Mutation of Methylosinus trichosporium. IEEE Trans. Plasma Sci. 2012, 40, 2853–2860. [Google Scholar]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Ma, Y.; Deng, Y.; Zhou, Z.; Cao, Y.; Yang, B.; Bai, J.; Sun, Q. Enhancing Protease and Amylase Activities in Bacillus licheniformis XS-4 for Traditional Soy Sauce Fermentation Using ARTP Mutagenesis. Foods 2023, 12, 2381. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, H.; Chen, X.; Sun, B.; Du, G.; Zhou, Z.; Song, J.; Fan, Y.; Shen, W. Significantly improving the yield of recombinant proteins in Bacillus subtilis by a novel powerful mutagenesis tool (ARTP): Alkaline α-amylase as a case study. Protein Expr. Purif. 2015, 114, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Yue, J.; Juan, S.Y.; Chun, J.J.; Juan, T.Y. Breeding of a thermostable xylanase-producing strain of Myceliophthora thermophila by atmospheric room temperature plasma (ARTP) mutagenesis. Front. Bioeng. Biotechnol. 2023, 10, 1095323. [Google Scholar]
- He, R.; Ding, R.; Heyman, J.A.; Zhang, D.; Tu, R. Ultra-high-throughput picoliter-droplet microfluidics screening of the industrial cellulase-producing filamentous fungus Trichoderma reesei. J. Ind. Microbiol. Biotechnol. 2019, 46, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xia, Y.; Li, M.; Zhang, T. ARTP mutagenesis of phospholipase D-producing strain Streptomyces hiroshimensis SK43.001, and its enzymatic properties. Heliyon 2022, 8, e12587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Tong, X.; Wang, W.; Wang, J.; Qu, W. Agarose biodegradation by deep-sea bacterium Vibrio natriegens WPAGA4 with the agarases through horizontal gene transfer. J. Basic Microbiol. 2023, 64, 2300521. [Google Scholar] [CrossRef]
- Tong, X.; Wang, N.; Zhang, M.; Wang, W.; Zhang, M.; Deng, S.; Wang, J.; Zeng, R.; Qu, W. Cloning, expression, and characterization of three agarases in Vibrio natriegens WPAGA4. Acta Microbiol. Sin. 2023, 63, 3667–3678. (In Chinese) [Google Scholar]
- Zhang, M.; Wang, J.; Zeng, R.; Wang, D.; Wang, W.; Tong, X.-C.; Qu, W. Agarose-Degrading Characteristics of a Deep-Sea Bacterium Vibrio Natriegens WPAGA4 and Its Cold-Adapted GH50 Agarase Aga3420. Mar. Drugs 2022, 20, 692. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 2002, 31, 426–428. [Google Scholar] [CrossRef]
- Cao, S.; Zhou, X.; Jin, W.; Wang, F.; Tu, R.; Han, S.; Chen, H.; Chen, C.; Xie, G.-J.; Ma, F. Improving of lipid productivity of the oleaginous microalgae Chlorella pyrenoidosa via atmospheric and room temperature plasma (ARTP). Bioresour. Technol. 2017, 244, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Duleepa, P.; Line, C.; Byeonghyeok, P.; Mikkel, S.; Geul, B.; Peter, S.; InGeol, C. A Novel Auxiliary Agarolytic Pathway Expands Metabolic Versatility in the Agar-Degrading Marine Bacterium Colwellia echini A3T. Appl. Environ. Microbiol. 2021, 87, e0023021. [Google Scholar]
- Gu, L.-S.; Tan, M.-Z.; Li, S.-H.; Zhang, T.; Zhang, Q.-Q.; Li, C.-X.; Luo, X.-M.; Feng, J.-X.; Zhao, S. ARTP/EMS-combined multiple mutagenesis efficiently improved production of raw starch-degrading enzymes in Penicillium oxalicum and characterization of the enzyme-hyperproducing mutant. Biotechnol. Biofuels 2020, 13, 187. [Google Scholar] [CrossRef] [PubMed]
- Colussi, F.; Serpa, V.; da Silva Delabona, P.; Manzine, L.R.; Voltatodio, M.L.; Alves, R.; Mello, B.L.; Pereira, N., Jr.; Farinas, C.S.; Golubev, A.M.; et al. Purification, and biochemical and biophysical characterization of cellobiohydrolase I from Trichoderma harzianum IOC 3844. J. Microbiol. Biotechnol. 2011, 21, 808–817. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).