
Fluid–Structure Interaction Analysis of Manta-Bots with Self-Induced Vertical Undulations during Fin-Based Locomotion


Abstract
1. Introduction
1.1. Fin Structures of Manta-like AUVs
1.2. Numerical Simulations
1.3. Research Objectives
2. Method and Validation
2.1. Flexible Multibody Dynamics
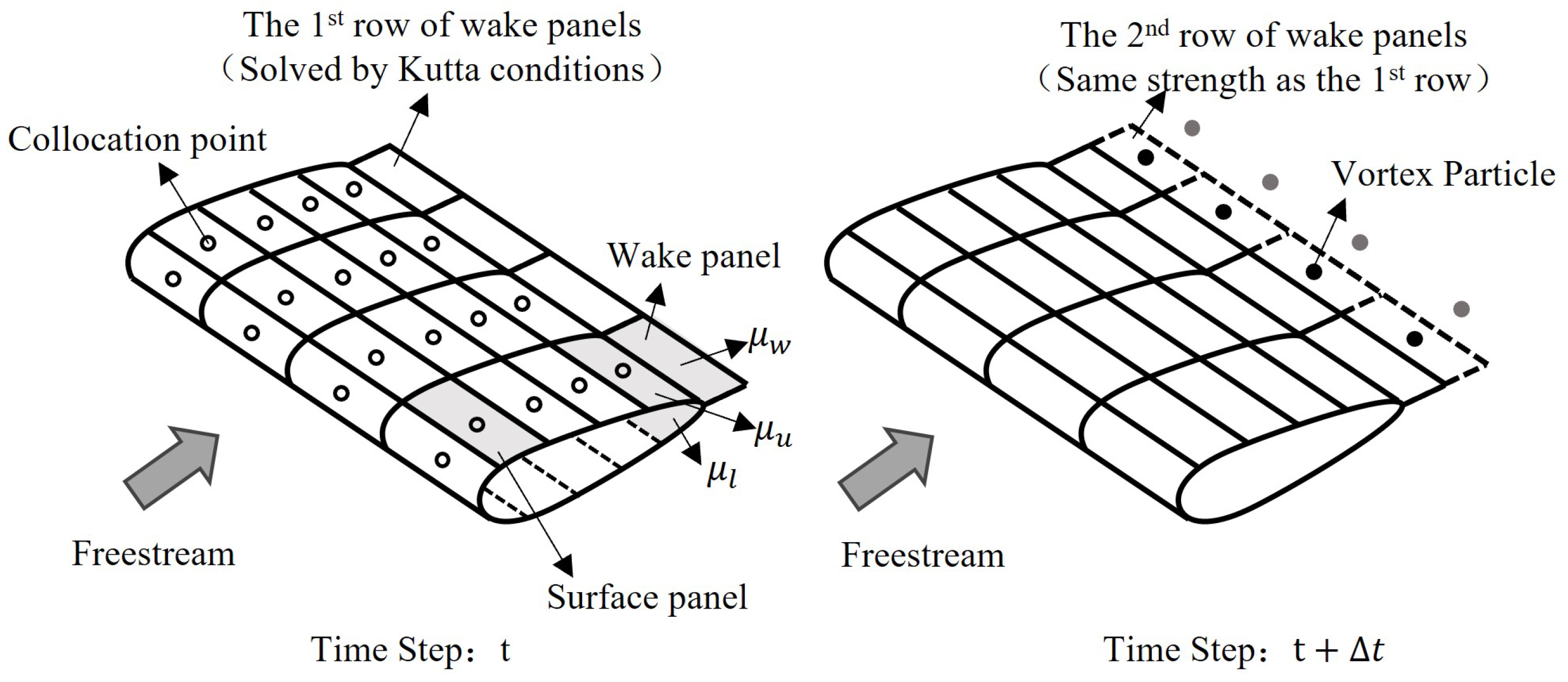
2.2. Unsteady Panel and Vortex Particle Hybrid Method
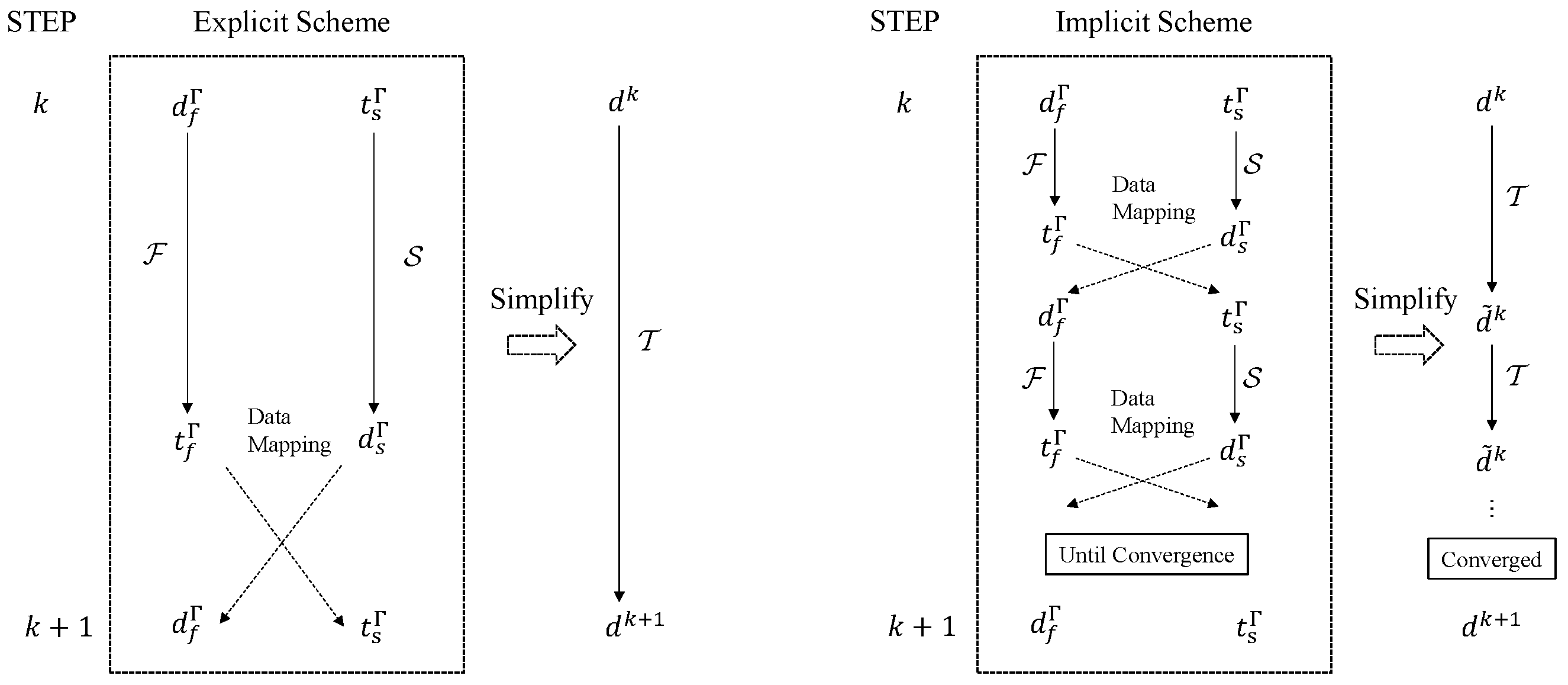
2.3. Partitioned FSI Coupling Techniques
2.3.1. Mapping Techniques
2.3.2. Coupling Algorithms
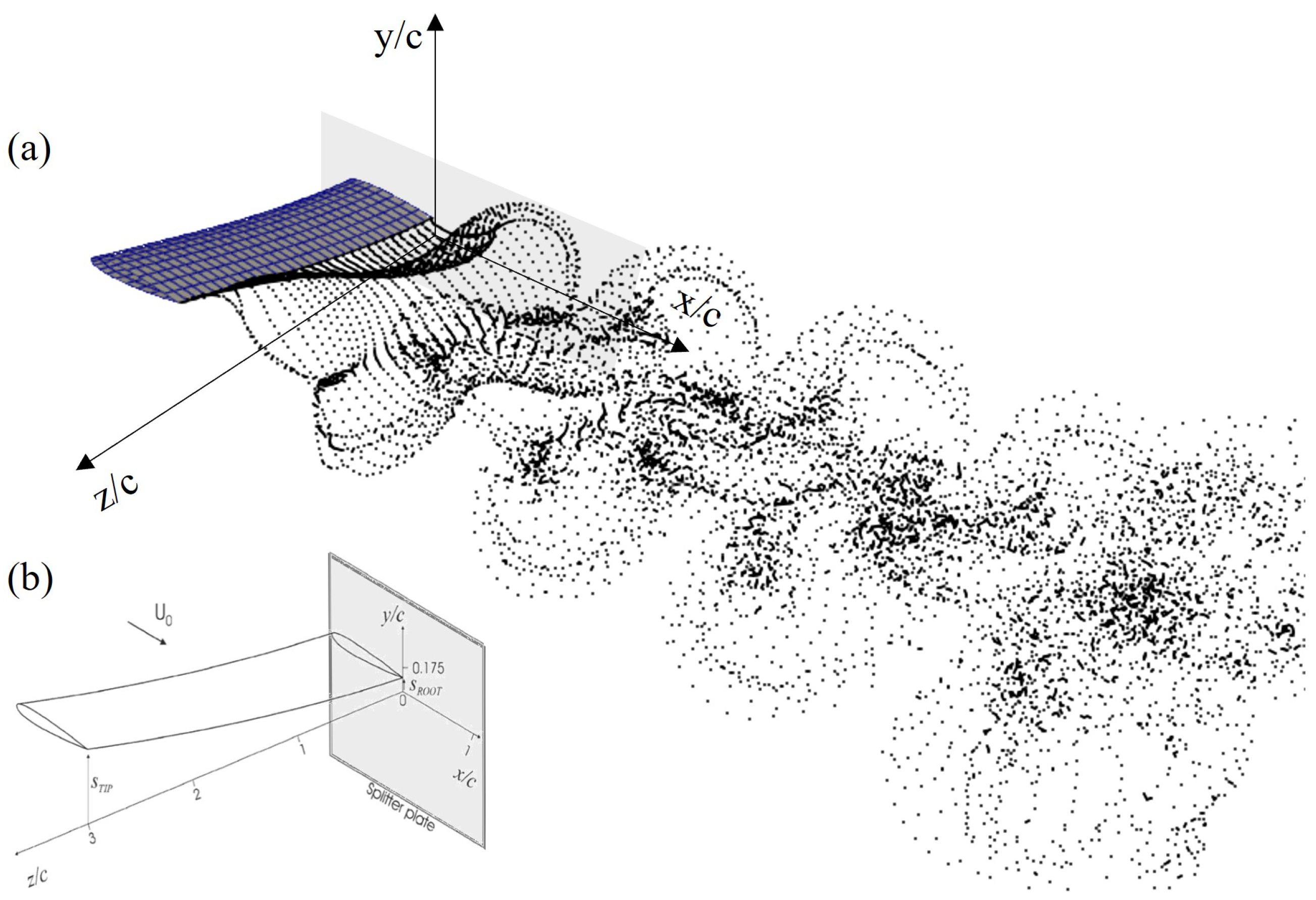
2.4. Validation
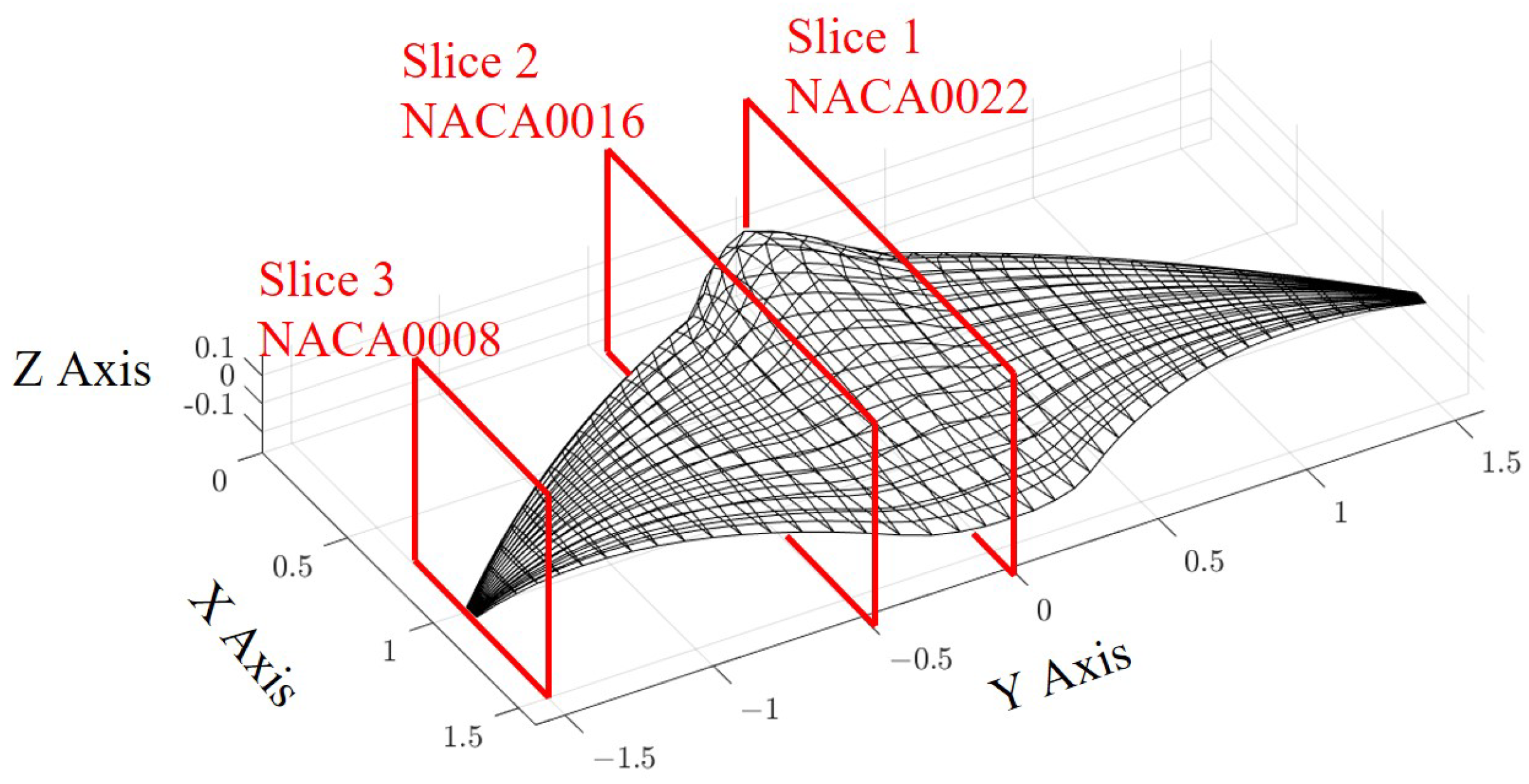
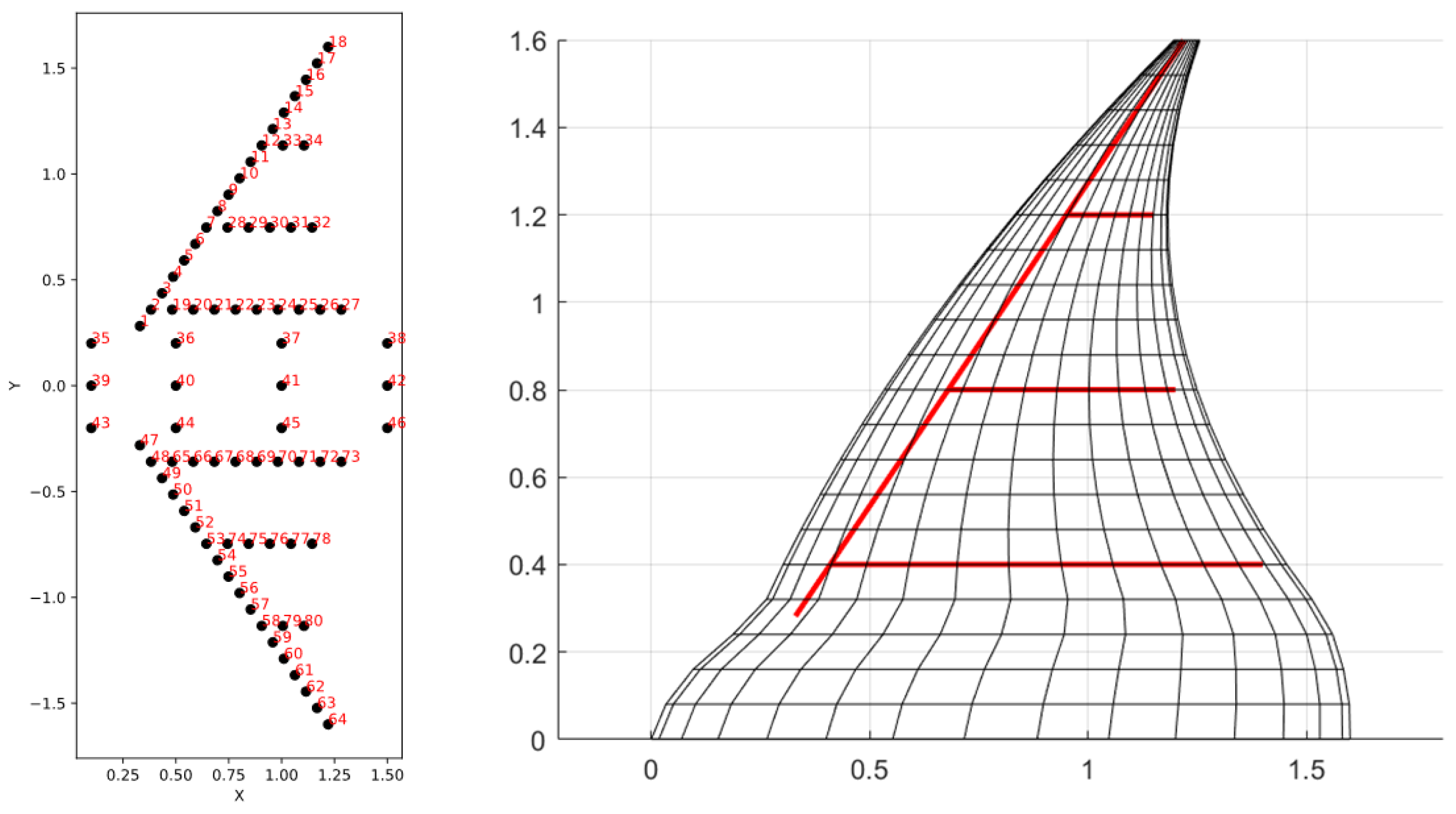
3. Model Description
4. Fixed Vertical Position Analysis
4.1. Basic Analysis
4.2. Effects of Flexibility
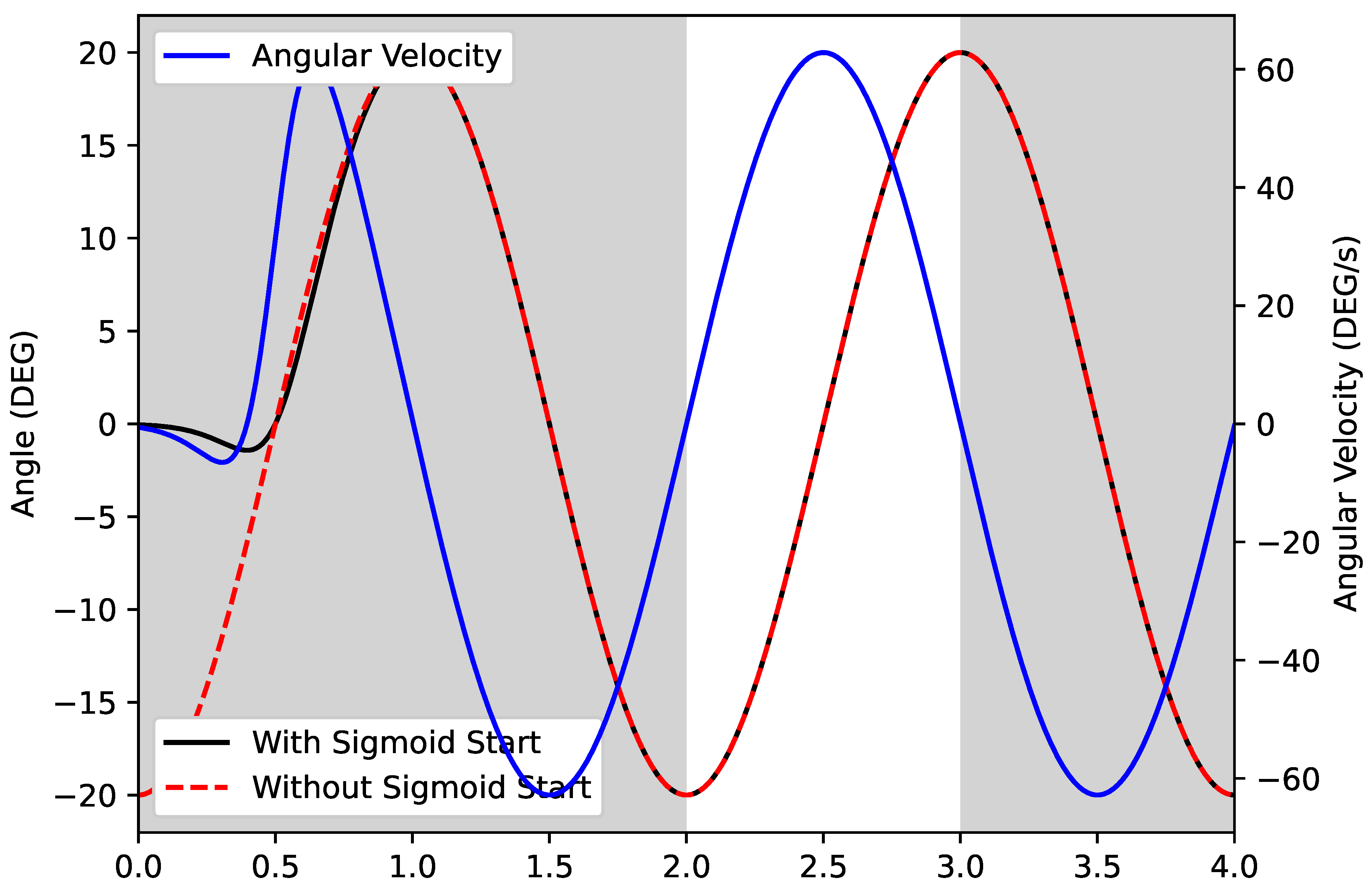
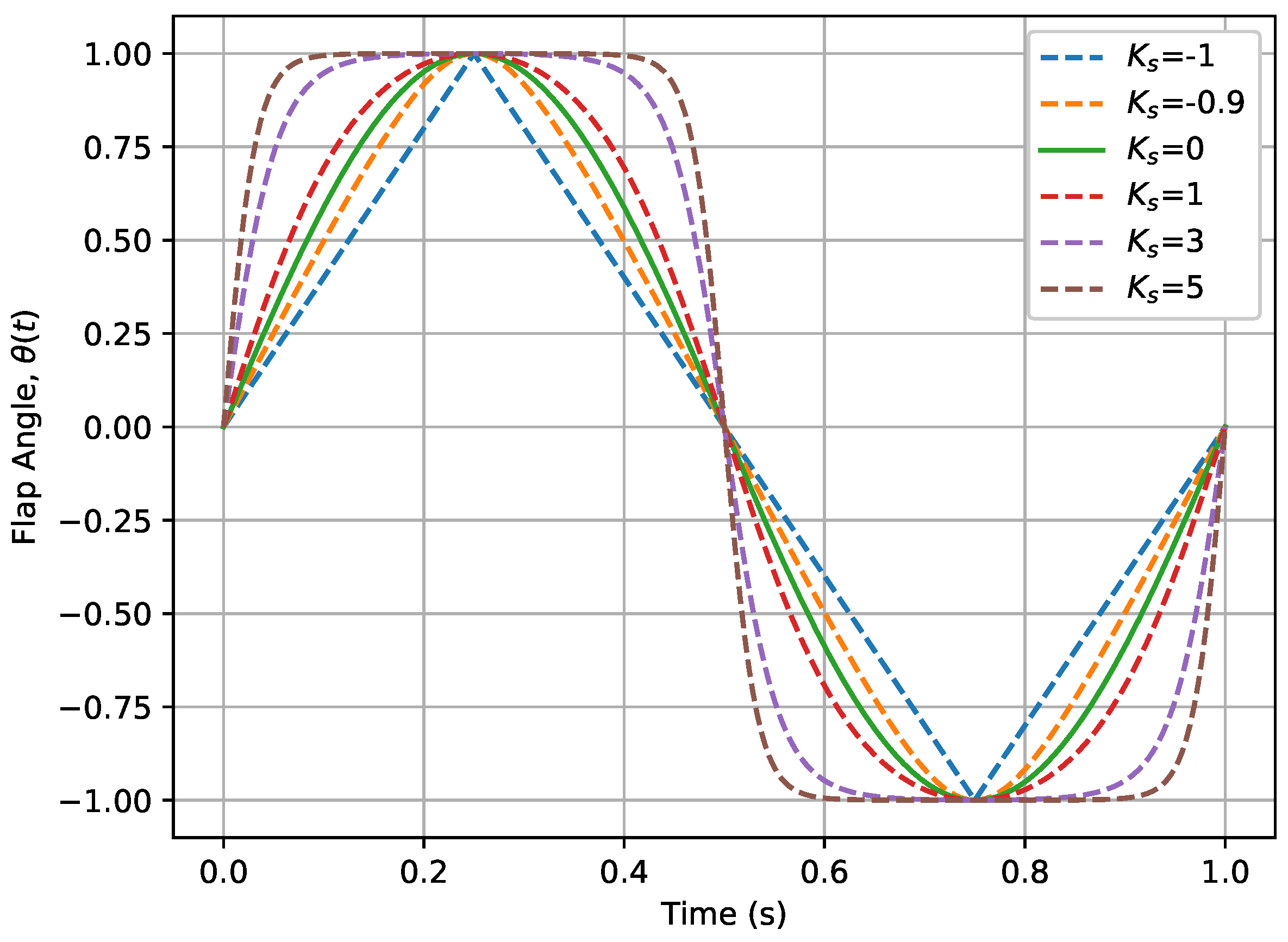
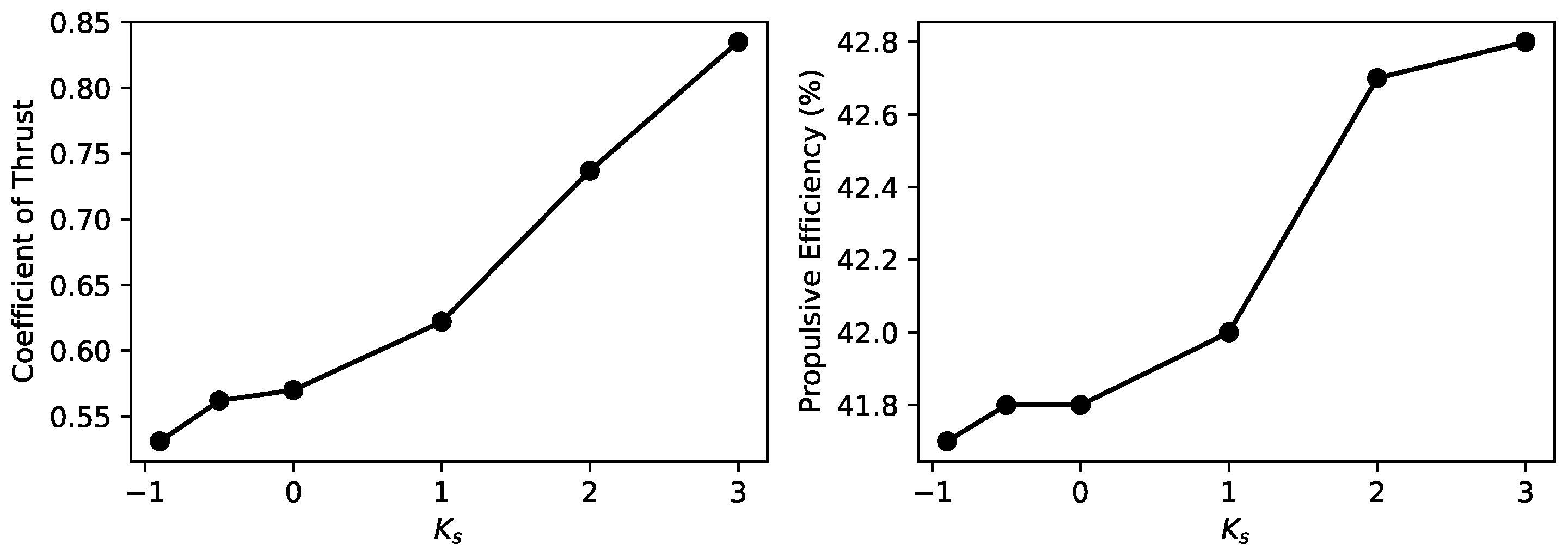
4.3. Effects of Non-Sinusoidal Flap Motion
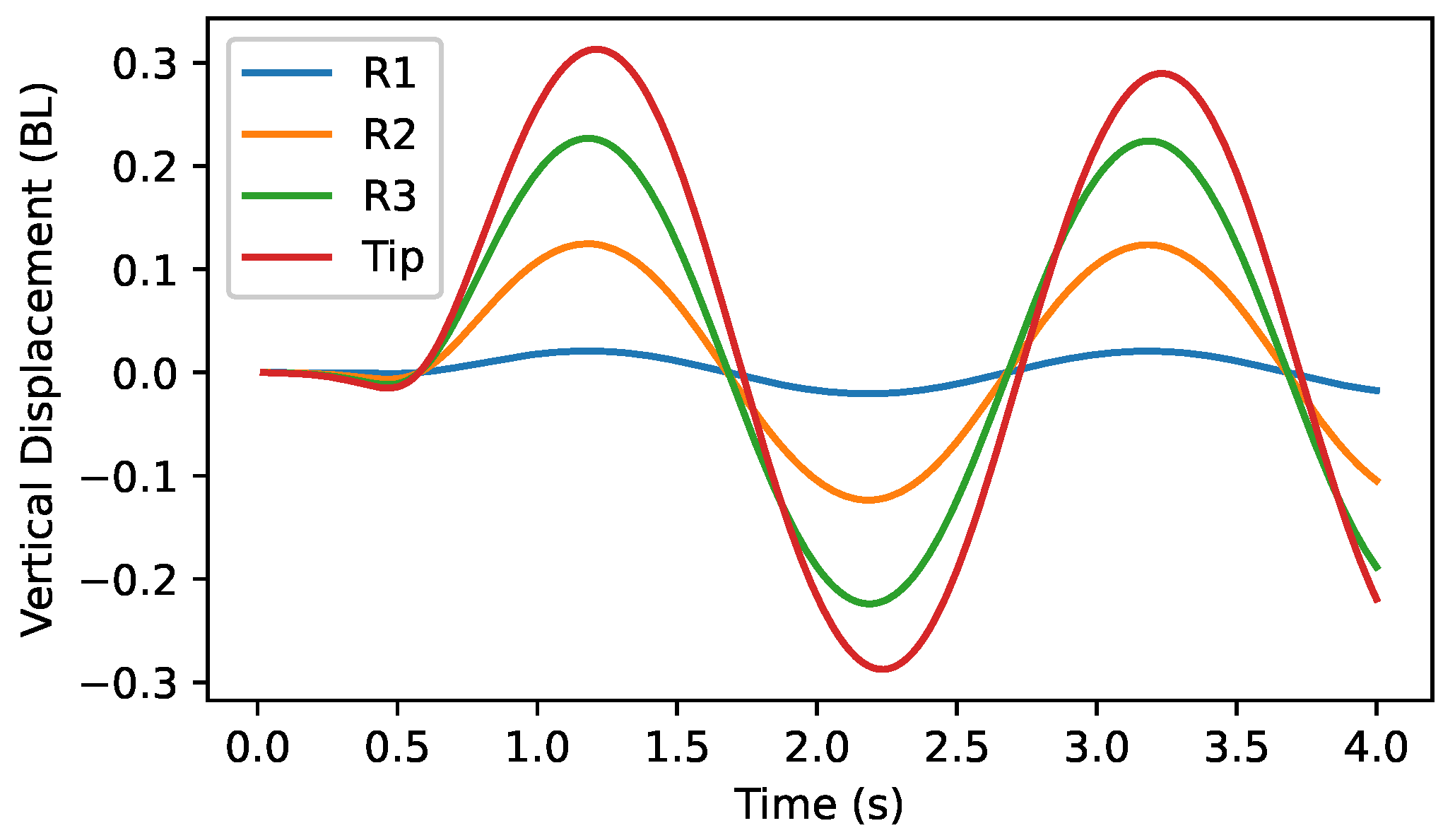
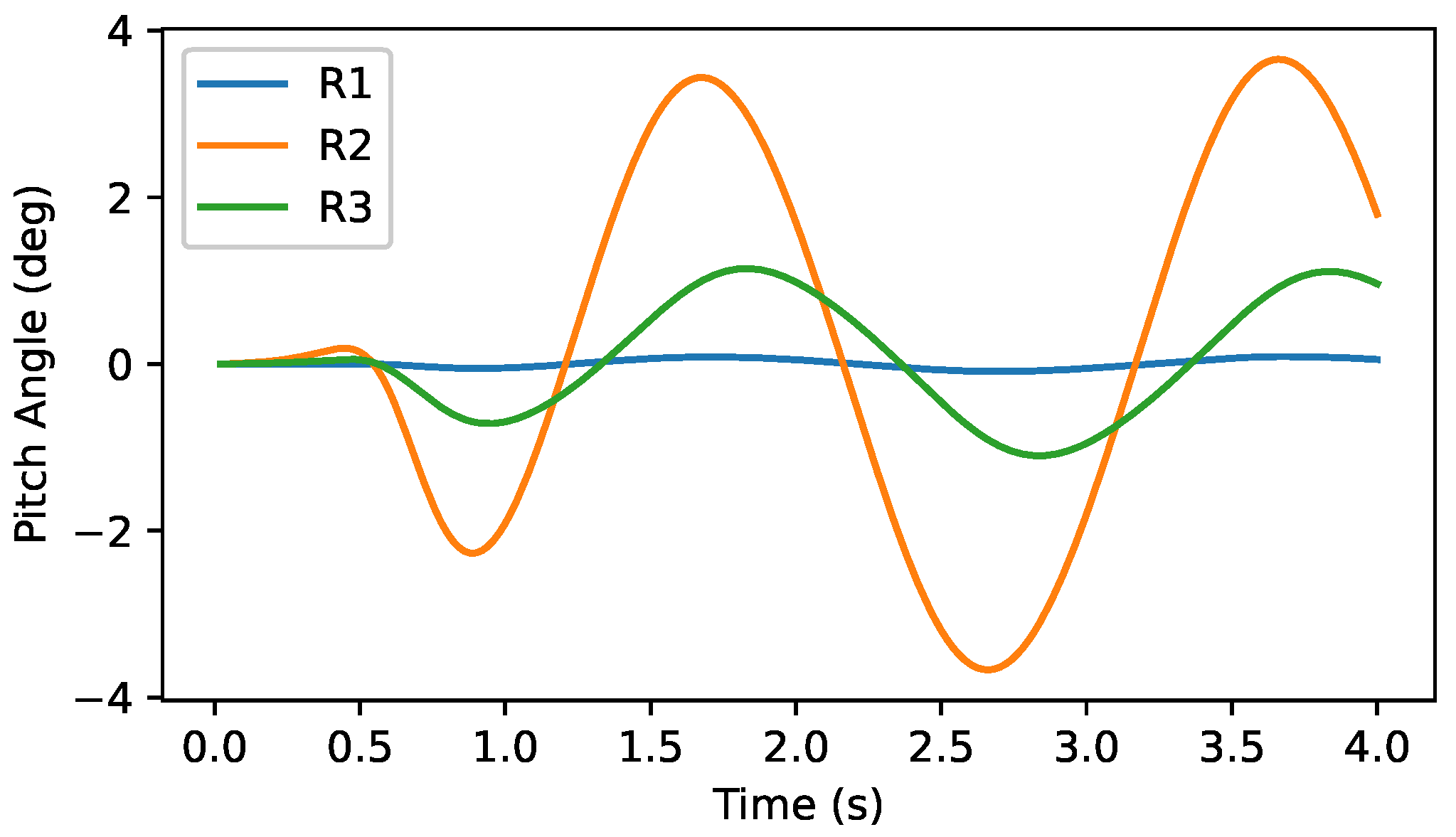
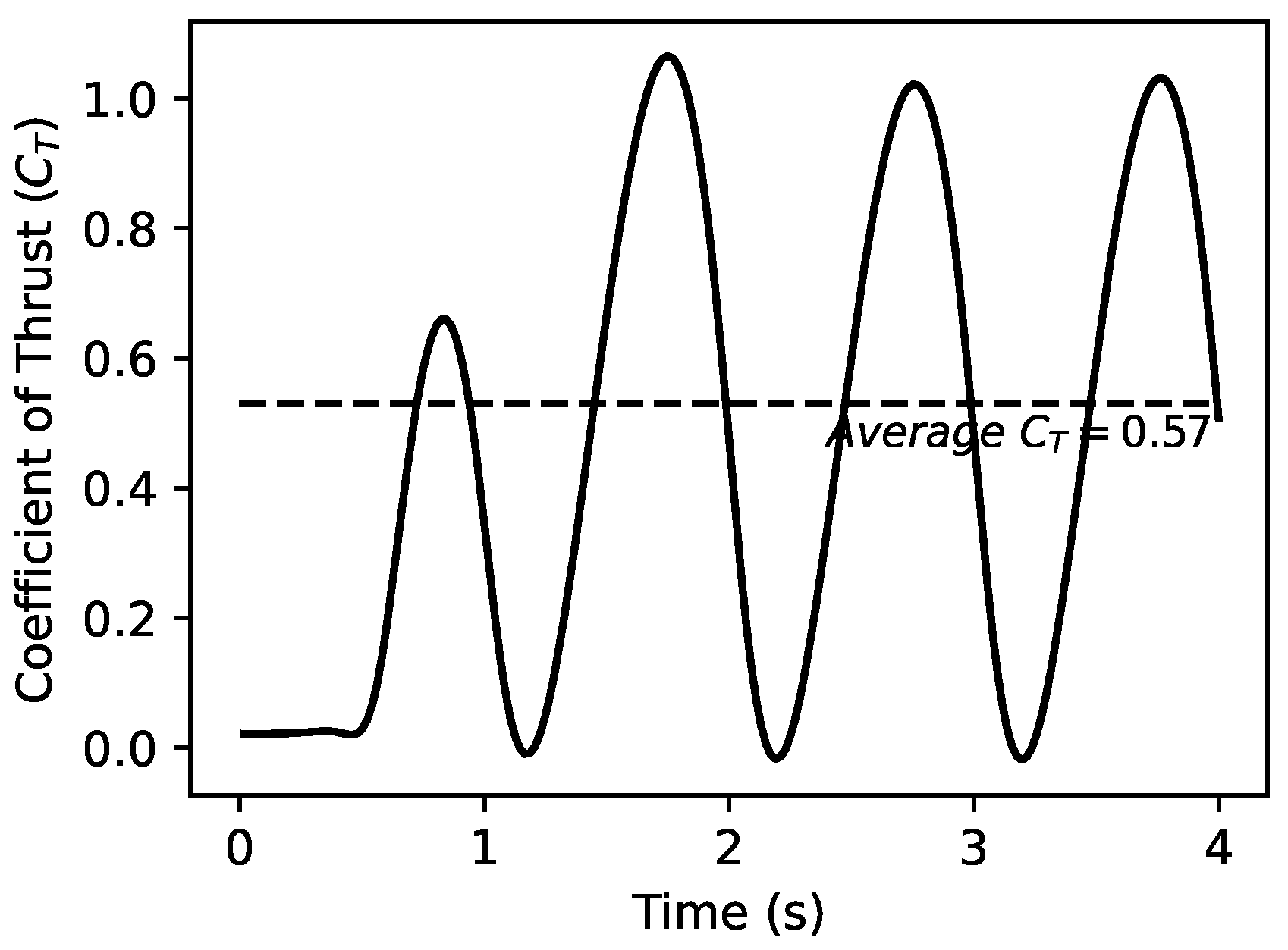
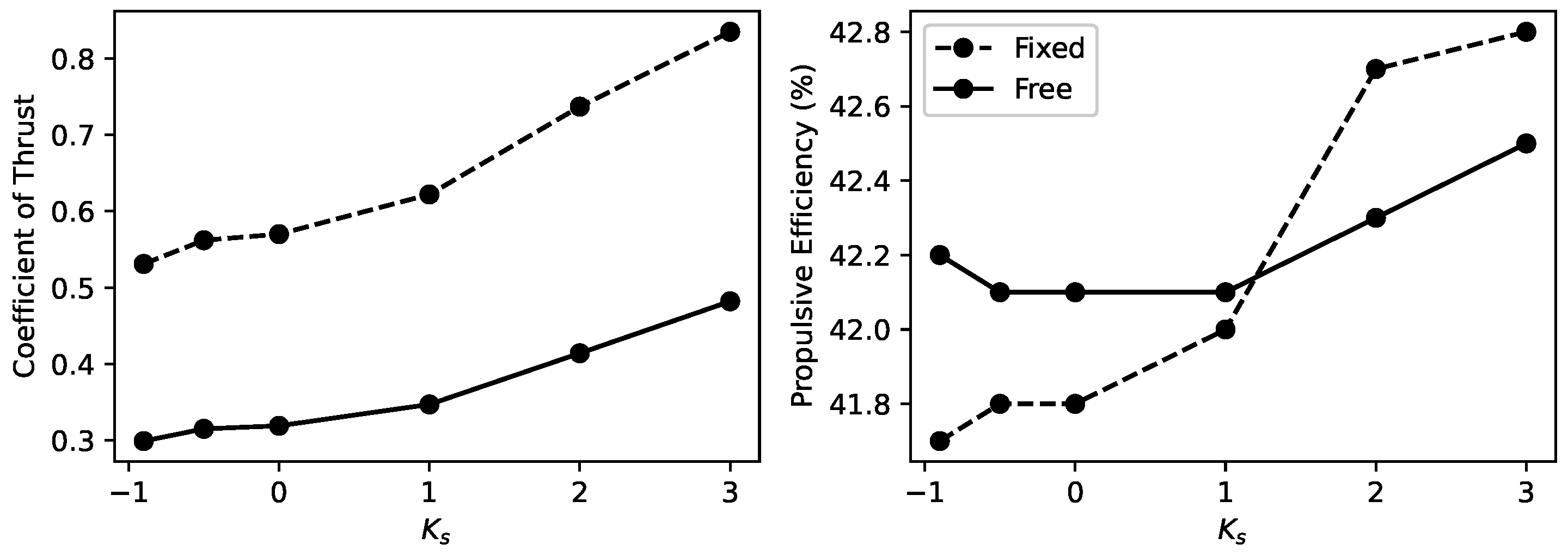
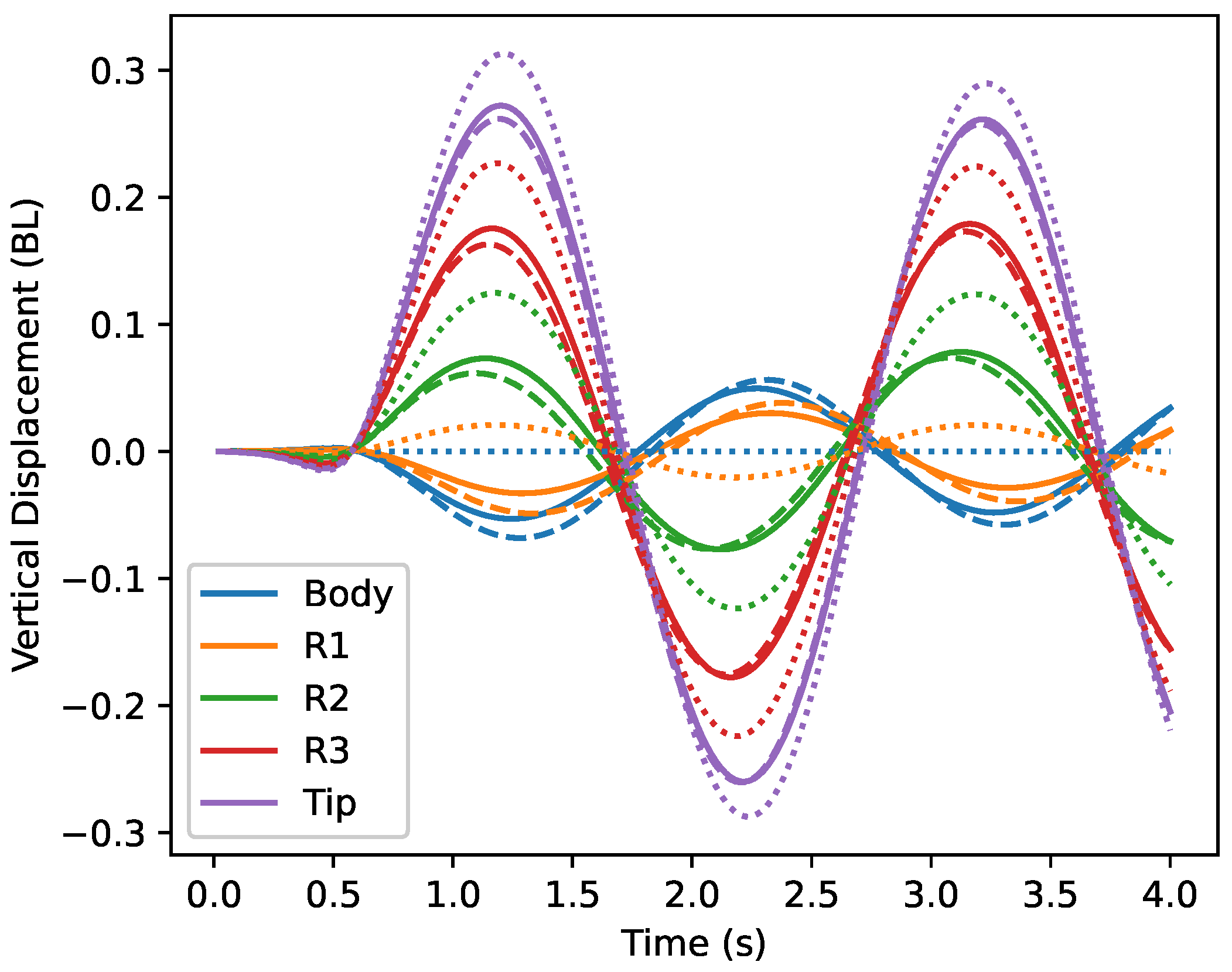
5. Self-Generated Vertical Undulations Analysis
5.1. Basic Analysis
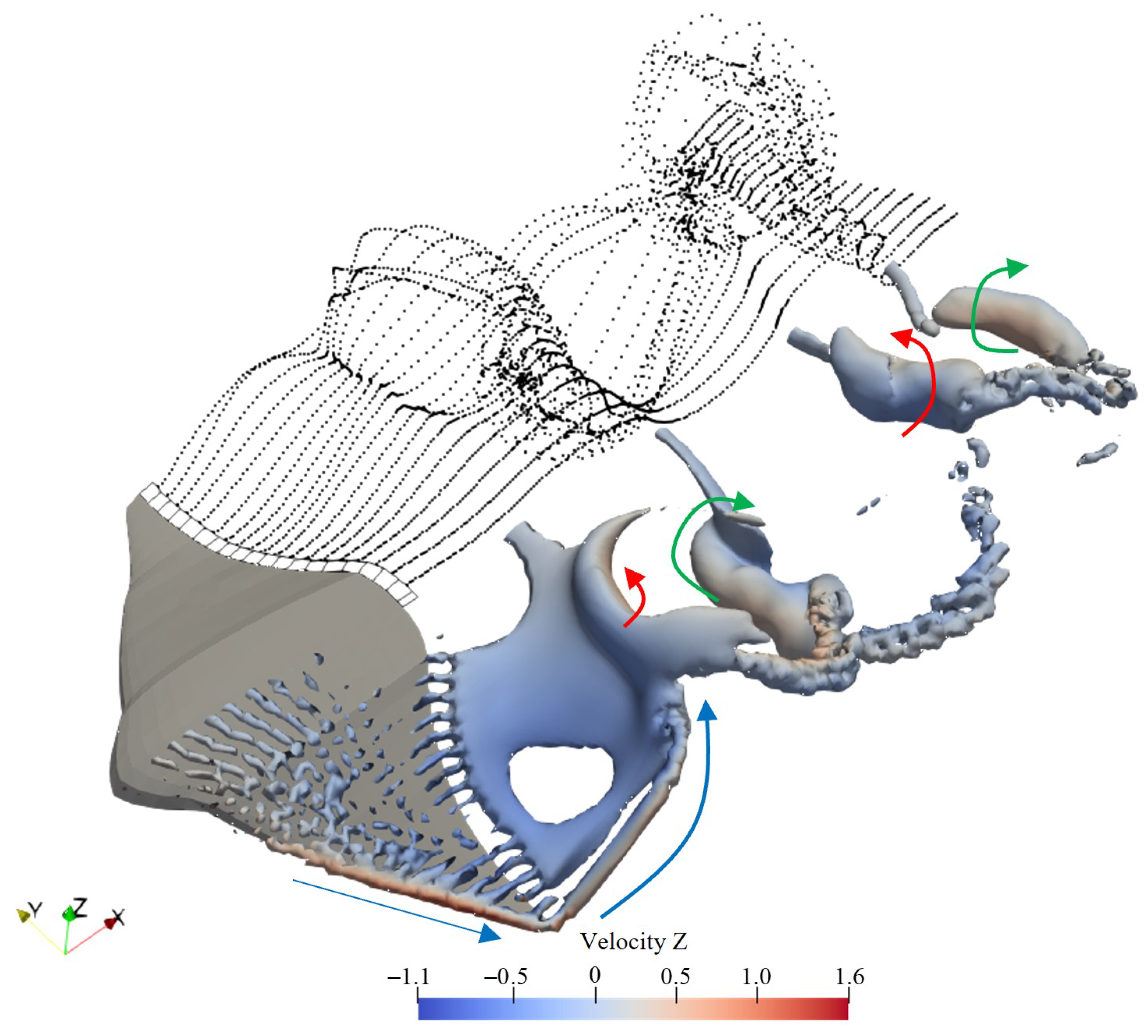
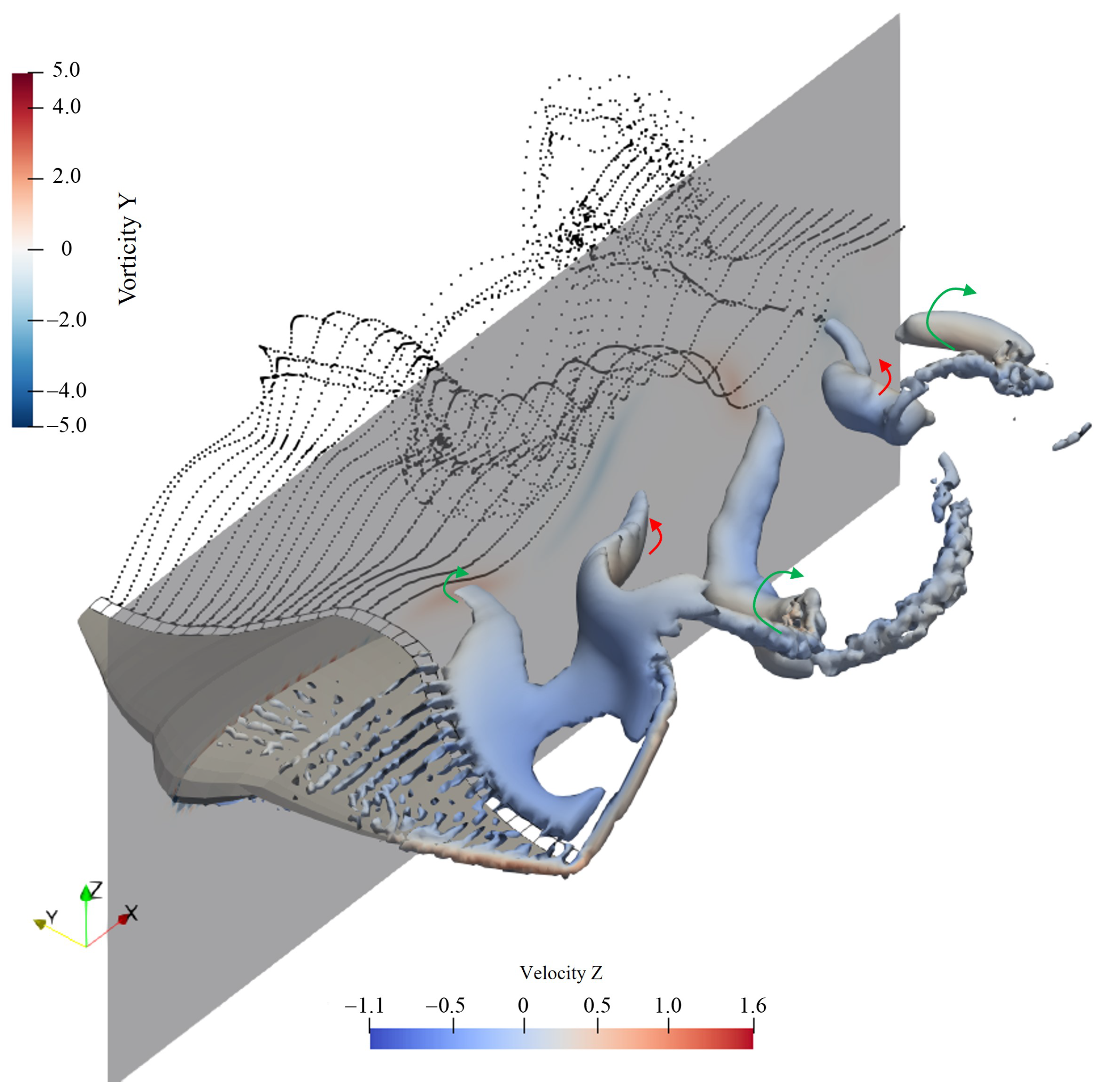
- The strength of the leading-edge vortex is significantly reduced, with a noticeable decrease in its coverage area near the body. This reduction leads to a diminished force exerted by the flow field on the structure;
- At the symmetry plane behind the body, the tail vortex exhibits stretching in the direction of the incoming flow, and its intensity is also diminished. This shift causes the location where the reverse Kármán vortex street typically forms to move backward. Given that the inductive effect of vortices follows an inverse square law, the farther from the body, the weaker the induced effect. As a result, the thrust generated by the reverse Kármán vortex street near the body is weakened.
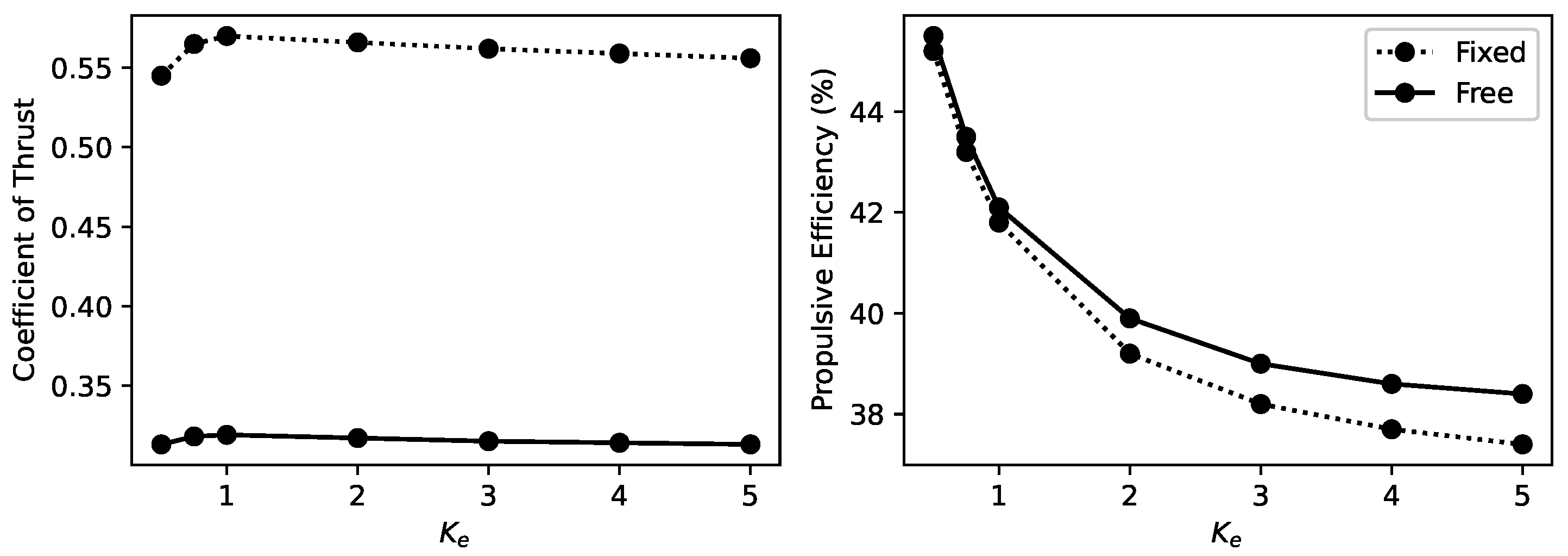
5.2. Effects of Flexibility
5.3. Effects of Non-Sinusoidal Flap Motion
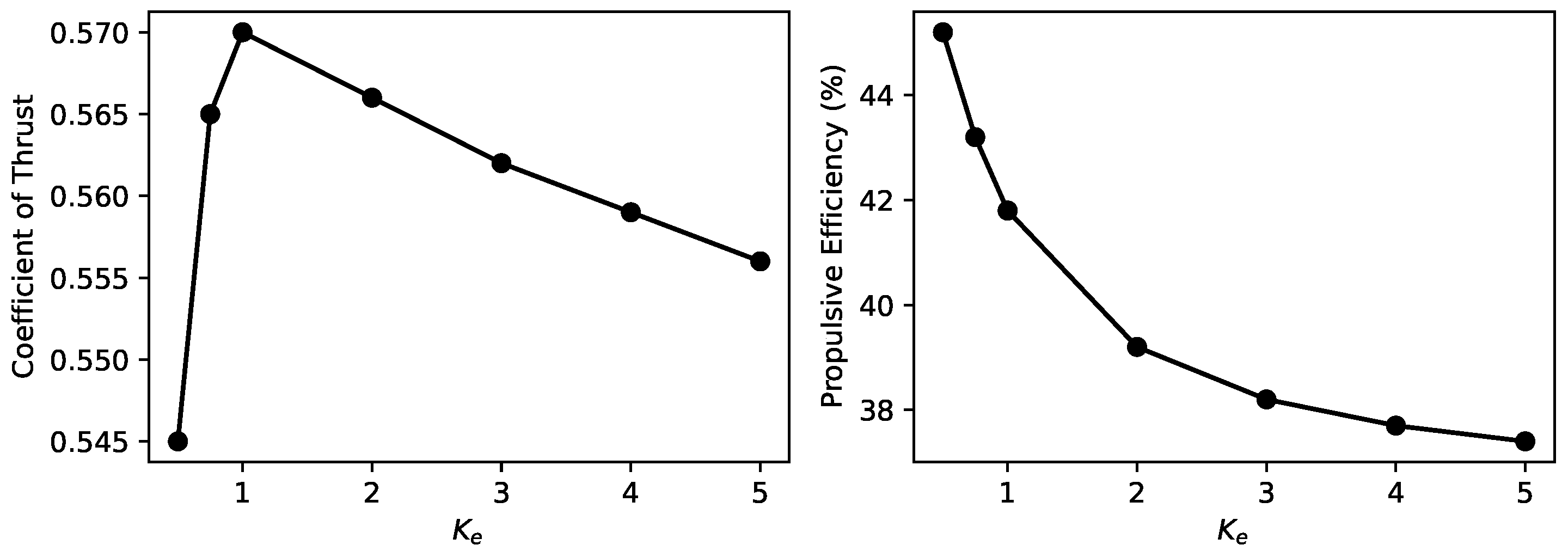
5.4. Effects of Mass Distribution
6. Conclusions
- The introduction of vertical freedom has notably diminished the influence of stiffness variations on thrust, resulting in a significant overall decrease in thrust. However, the propulsive efficiency shows a slight overall improvement, maintaining a consistent influence trend despite these changes.
- The adoption of a non-sinusoidal, square-wave flapping motion pattern brings us closer to the actual flapping motion of the manta ray, which significantly enhances thrust without substantially affecting propulsive efficiency. The impact of this motion pattern remains consistent even with the introduction of longitudinal degrees of freedom, suggesting a robust design strategy for enhancing propulsion.
- The distribution of mass is critically important in the design of biomimetic manta rays and has been largely overlooked in previous studies. An excessive focus on replicating the complex structure of the pectoral fins to mimic the real manta ray’s flapping motion has inadvertently increased the mass proportion of the pectoral fins. This, in turn, induces more pronounced longitudinal undulatory motions in the body, substantially reducing thrust.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salazar, R.; Campos, A.; Fuentes, V.; Abdelkefi, A. A review on the modeling, materials, and actuators of aquatic unmanned vehicles. Ocean. Eng. 2019, 172, 257–285. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Zhou, F.; Liang, Y.; Xiao, Y.; Cao, X.; Zhang, Z.; Zhang, M.; Wu, B.; Yin, S.; et al. Self-powered soft robot in the Mariana Trench. Nature 2021, 591, 66–71. [Google Scholar] [CrossRef]
- Lamas, M.; Rodriguez, C. Hydrodynamics of biomimetic marine propulsion and trends in computational simulations. J. Mar. Sci. Eng. 2020, 8, 479. [Google Scholar] [CrossRef]
- Babu Mannam, N.P.; Mahbub Alam, M.; Krishnankutty, P. Review of biomimetic flexible flapping foil propulsion systems on different planetary bodies. Results Eng. 2020, 8, 100183. [Google Scholar] [CrossRef]
- Wei, Q.P.; Wang, S.; Dong, X.; Shang, L.J.; Tan, M. Design and Kinetic Analysis of a Biomimetic Underwater Vehicle with Two Undulating Long-fins. Acta Autom. Sin. 2013, 39, 1330–1338. [Google Scholar] [CrossRef]
- He, J.; Cao, Y.; Huang, Q.; Pan, G.; Dong, X.; Cao, Y. Effects of bionic pectoral fin rays’ spanwise flexibility on forwarding propulsion performance. J. Mar. Sci. Eng. 2022, 10, 783. [Google Scholar] [CrossRef]
- Liu, K.; Liu, X.; Huang, H. Scaling the self-propulsive performance of pitching and heaving flexible plates. J. Fluid Mech. 2022, 936, A9. [Google Scholar] [CrossRef]
- Cao, Y.; Cao, Y.; Ma, S.; Li, X.; Qu, Y.; Cao, Y. Realization and Online Optimization for Gliding and Flapping Propulsion of a Manta Ray Robot. J. Mar. Sci. Eng. 2023, 11, 2173. [Google Scholar] [CrossRef]
- Zhou, C.; Low, K.H. Better Endurance and Load Capacity: An Improved Design of Manta Ray Robot (RoMan-II). J. Bionic Eng. 2010, 7, S137–S144. [Google Scholar] [CrossRef]
- Lu, Y.; Cao, Y.; Pan, G.; Huang, Q.; Dong, X.; Cao, Y. Effect of cross-joints fin on the thrust performance of bionic pectoral fins. J. Mar. Sci. Eng. 2022, 10, 869. [Google Scholar] [CrossRef]
- Chew, C.M.; Lim, Q.Y.; Yeo, K.S. Development of propulsion mechanism for Robot Manta Ray. In Proceedings of the 2015 IEEE International Conference on Robotics and Biomimetics (ROBIO), Zhuhai, China, 6–9 December 2015; pp. 1918–1923. [Google Scholar] [CrossRef]
- Chen, L.; Bi, S.; Cai, Y.; Cao, Y.; Pan, G. Design and experimental research on a bionic robot fish with tri-dimensional soft pectoral fins inspired by cownose ray. J. Mar. Sci. Eng. 2022, 10, 537. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, H.; Wang, Z.; He, Q.; Chen, L.; Li, W.; Li, R.; Cui, W. A manta ray robot with soft material based flapping wing. J. Mar. Sci. Eng. 2022, 10, 962. [Google Scholar] [CrossRef]
- Xing, C.; Cao, Y.; Cao, Y.; Pan, G.; Huang, Q. Asymmetrical Oscillating Morphology Hydrodynamic Performance of a Novel Bionic Pectoral Fin. J. Mar. Sci. Eng. 2022, 10, 289. [Google Scholar] [CrossRef]
- Fish, F.; Schreiber, C.; Moored, K.; Liu, G.; Dong, H.; Bart-Smith, H. Hydrodynamic Performance of Aquatic Flapping: Efficiency of Underwater Flight in the Manta. Aerospace 2016, 3, 20. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, L.; Guo, S.; Yin, H.; Li, A.; Bao, P.; Liu, M. Mechanism Design, Kinematic and Hydrodynamic Simulation of a Wave-driven Amphibious Robot. In Proceedings of the 2021 IEEE International Conference on Mechatronics and Automation (ICMA), Takamatsu, Japan, 8–11 August 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 1038–1043. [Google Scholar]
- Huang, H.; Sheng, C.; Jiannan, W.; Wu, G.; Zhou, C.; Wang, H. Hydrodynamic analysis and motion simulation of fin and propeller driven manta ray robot. Appl. Ocean. Res. 2021, 108, 102528. [Google Scholar] [CrossRef]
- Qu, Y.; Xie, X.; Zhang, S.; Xing, C.; Cao, Y.; Cao, Y.; Pan, G.; Song, B. A Rigid-Flexible Coupling Dynamic Model for Robotic Manta with Flexible Pectoral Fins. J. Mar. Sci. Eng. 2024, 12, 292. [Google Scholar] [CrossRef]
- Wang, W.; Huang, H.; Lu, X.Y. Interplay of chordwise stiffness and shape on performance of self-propelled flexible flapping plate. Phys. Fluids 2021, 33, 091904. [Google Scholar] [CrossRef]
- Lapsansky, A.B.; Zatz, D.; Tobalske, B.W. Alcids ‘fly’at efficient Strouhal numbers in both air and water but vary stroke velocity and angle. eLife 2020, 9, e55774. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Wu, Z.; Yang, C. Strongly coupled fluid–structure interaction analysis of aquatic flapping wings based on flexible multibody dynamics and the modified unsteady vortex lattice method. Ocean. Eng. 2023, 281, 114921. [Google Scholar] [CrossRef]
- Tasora, A.; Serban, R.; Mazhar, H.; Pazouki, A.; Melanz, D.; Fleischmann, J.; Taylor, M.; Sugiyama, H.; Negrut, D. Chrono: An Open Source Multi-physics Dynamics Engine. In High Performance Computing in Science and Engineering; Kozubek, T., Blaheta, R., Šístek, J., Rozložník, M., Čermák, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 9611, pp. 19–49. [Google Scholar] [CrossRef]
- Recuero, A.; Negrut, D. Chrono Support for ANCF Finite Elements: Formulation and Validation Aspects; ResearchGate: Berlin, Germany, 2016. [Google Scholar]
- Lee, H.; Sengupta, B.; Araghizadeh, M.; Myong, R. Review of vortex methods for rotor aerodynamics and wake dynamics. Adv. Aerodyn. 2022, 4, 20. [Google Scholar] [CrossRef]
- Hess, J.L. The problem of three-dimensional lifting potential flow and its solution by means of surface singularity distribution. Comput. Methods Appl. Mech. Eng. 1974, 4, 283–319. [Google Scholar] [CrossRef]
- TAN, J.; WANG, H.; WU, C.; LIN, C. Rotor/Empennage Unsteady Aerodynamic Interaction with Unsteady Panel/Viscous Vortex Particle Hybrid Method. Acta Aeronaut. Astronaut. Sin. 2014, 35, 14. [Google Scholar]
- Tugnoli, M.; Montagnani, D.; Syal, M.; Droandi, G.; Zanotti, A. Mid-fidelity approach to aerodynamic simulations of unconventional VTOL aircraft configurations. Aerosp. Sci. Technol. 2021, 115, 106804. [Google Scholar] [CrossRef]
- Dettmer, W.; Perić, D. A computational framework for fluid–rigid body interaction: Finite element formulation and applications. Comput. Methods Appl. Mech. Eng. 2006, 195, 1633–1666. [Google Scholar] [CrossRef]
- Wendland, H. Piecewise polynomial, positive definite and compactly supported radial functions of minimal degree. Adv. Comput. Math. 1995, 4, 389–396. [Google Scholar] [CrossRef]
- Wendland, H. Scattered Data Approximation; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Chourdakis, G.; Davis, K.; Rodenberg, B.; Schulte, M.; Simonis, F.; Uekermann, B.; Abrams, G.; Bungartz, H.; Cheung Yau, L.; Desai, I.; et al. preCICE v2: A sustainable and User-Friendly Coupling Library [Version 2; peer Review: 2 Approved]. Open Research Europe. 2022, Volume 2. Available online: https://open-research-europe.ec.europa.eu/articles/2-51/v2 (accessed on 1 June 2024).
- Heathcote, S.; Wang, Z.; Gursul, I. Effect of spanwise flexibility on flapping wing propulsion. J. Fluids Struct. 2008, 24, 183–199. [Google Scholar] [CrossRef]
- Masarati, P.; Morandini, M.; Quaranta, G.; Chandar, D.; Roget, B.; Sitaraman, J. Tightly Coupled CFD/Multibody Analysis of Flapping-Wing Micro-Aerial Vehicles. In Proceedings of the 29th AIAA Applied Aerodynamics Conference, Honolulu, HI, USA, 27–30 June 2011. [Google Scholar] [CrossRef]
- Nakata, T.; Liu, H. A fluid–structure interaction model of insect flight with flexible wings. J. Comput. Phys. 2012, 231, 1822–1847. [Google Scholar] [CrossRef]
- Xue, D.; Song, B.; Song, W.; Yang, W.; Xu, W.; Wu, T. Computational simulation and free flight validation of body vibration of flapping-wing MAV in forward flight. Aerosp. Sci. Technol. 2019, 95, 105491. [Google Scholar] [CrossRef]
- Yang, S.b.; Qiu, J.; Han, X.y. Kinematics modeling and experiments of pectoral oscillation propulsion robotic fish. J. Bionic Eng. 2009, 6, 174–179. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Illustration | Features |
|---|---|---|
 |
| |
 |
| |
 |
|
| Parameter | Value | Units |
|---|---|---|
| Chord | 0.1 | m |
| Span | 0.3 | m |
| Young’s modulus | 70 | GPa |
| Density | kg/m3 | |
| Fundamental frequency | 1.517 | Hz |
| Mesh Level | Total Meshes | |||
|---|---|---|---|---|
| Ultra Fine | 20 | 60 | 2400 | 819.22 |
| Fine | 15 | 40 | 1200 | 821.93 |
| Medium | 10 | 30 | 600 | 823.64 |
| Coarse | 5 | 20 | 200 | 661.83 |
| Beam ID | Node Pair | w (cm) | h (cm) | E (GPa) | M (kg) |
|---|---|---|---|---|---|
| 1 | 1–7 | 5 | 8.5 | 150 | 6.442 |
| 2 | 7–12 | 3 | 3 | 70 | 1.137 |
| 3 | 12–18 | 1 | 1 | 50 | 0.152 |
| 4 | 2–27 | 3 | 3 | 100 | 2.187 |
| 5 | 7–32 | 1 | 1 | 50 | 0.135 |
| 6 | 12–34 | 1 | 1 | 50 | 0.054 |
| Feature Parameter | Value | Unit |
|---|---|---|
| Span Length, L | 3.2 | m |
| Chord Length, c | 1.6 | m |
| Flapping Frequency, f | 0.5 | Hz |
| Maximum Flapping Angle, | 30 | deg |
| Cruising Speed, | 1.0 | m/s |
| Number of Interpolation Points, N | 80 | count |
| Efficiency (%) | ||
|---|---|---|
| 0.02 | 0.319 | 42.1 |
| 0.25 | 0.302 | 42.3 |
| 0.66 | 0.287 | 42.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Wu, Z.; Zhou, M.; Yang, C. Fluid–Structure Interaction Analysis of Manta-Bots with Self-Induced Vertical Undulations during Fin-Based Locomotion. J. Mar. Sci. Eng. 2024, 12, 1165. https://doi.org/10.3390/jmse12071165
Luo M, Wu Z, Zhou M, Yang C. Fluid–Structure Interaction Analysis of Manta-Bots with Self-Induced Vertical Undulations during Fin-Based Locomotion. Journal of Marine Science and Engineering. 2024; 12(7):1165. https://doi.org/10.3390/jmse12071165
Chicago/Turabian StyleLuo, Ming, Zhigang Wu, Minghao Zhou, and Chao Yang. 2024. "Fluid–Structure Interaction Analysis of Manta-Bots with Self-Induced Vertical Undulations during Fin-Based Locomotion" Journal of Marine Science and Engineering 12, no. 7: 1165. https://doi.org/10.3390/jmse12071165
APA StyleLuo, M., Wu, Z., Zhou, M., & Yang, C. (2024). Fluid–Structure Interaction Analysis of Manta-Bots with Self-Induced Vertical Undulations during Fin-Based Locomotion. Journal of Marine Science and Engineering, 12(7), 1165. https://doi.org/10.3390/jmse12071165