
Identification of Key Habitats of Bowhead and Blue Whales in the OSPAR Area of the North-East Atlantic—A Modelling Approach towards Effective Conservation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
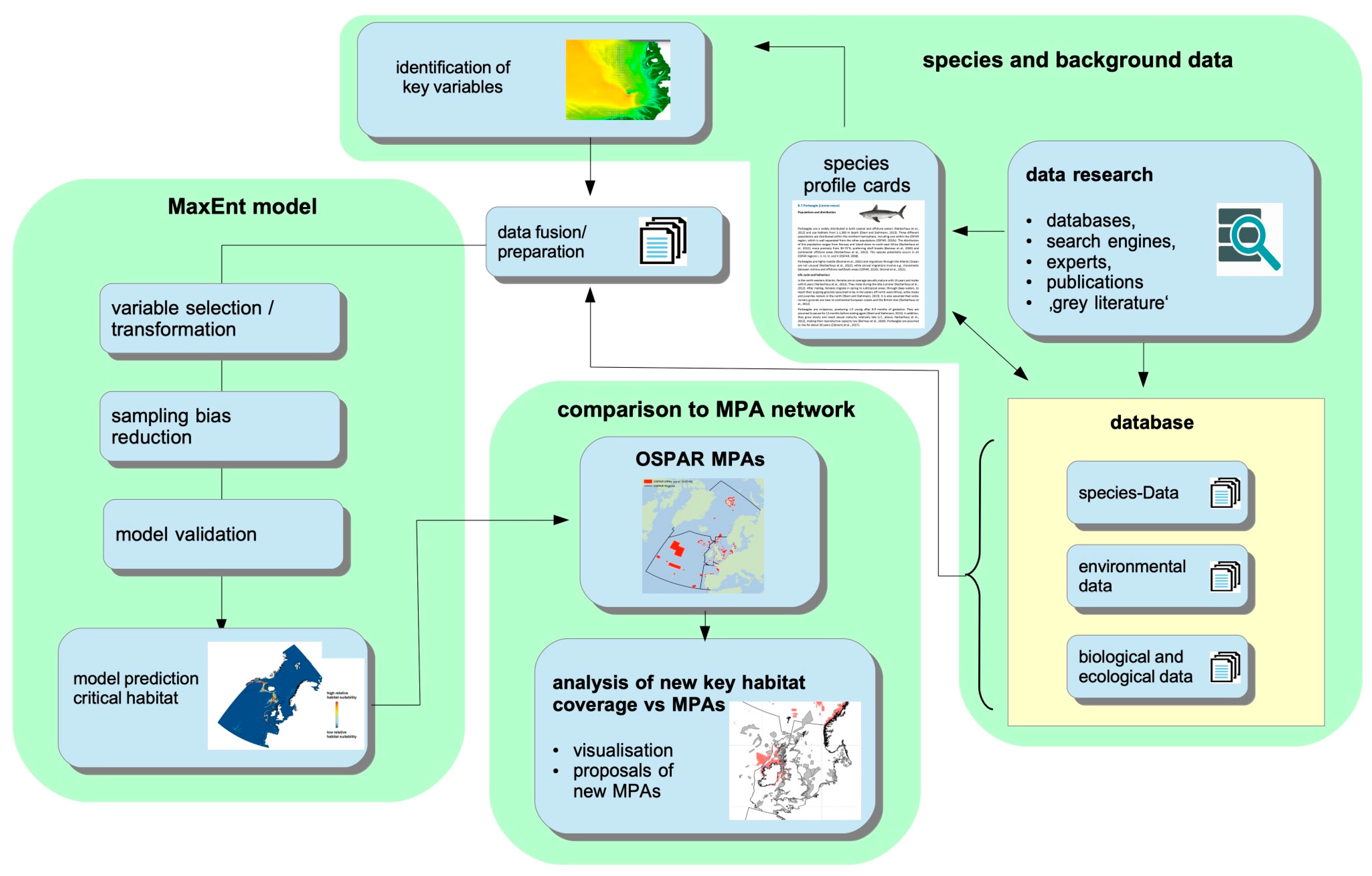
2. Materials and Methods
2.1. Species Data: Sources, Preparation and Suitability for Modelling
2.2. Definition of “Key Habitat” in the Context of the Project
2.3. Predictor Variables: Sources and Preparation
2.4. Statistical Methods
- Different species show different degrees of “patchiness” with respect to the distribution of (and strength of association with) key habitats. Thus, different sizes of protected areas are required to achieve the same strength/percentage of protection;
- the percentage of key habitats already protected by MPAs, and thus, the need for additional MPAs varies among species;
- our predicted key habitats differ in their quality of prediction, depending on both species and location (Supporting Material S2);
- for some of the species/identified locations for MPAs, protection measures may suffice for certain periods of the year (migratory species, breeding season);
- predicted key habitats of species can overlap, which increases the overall contribution of potential MPAs at those locations;
- MPAs should not be too fragmented and/or small (e.g., comprising only a few km2).
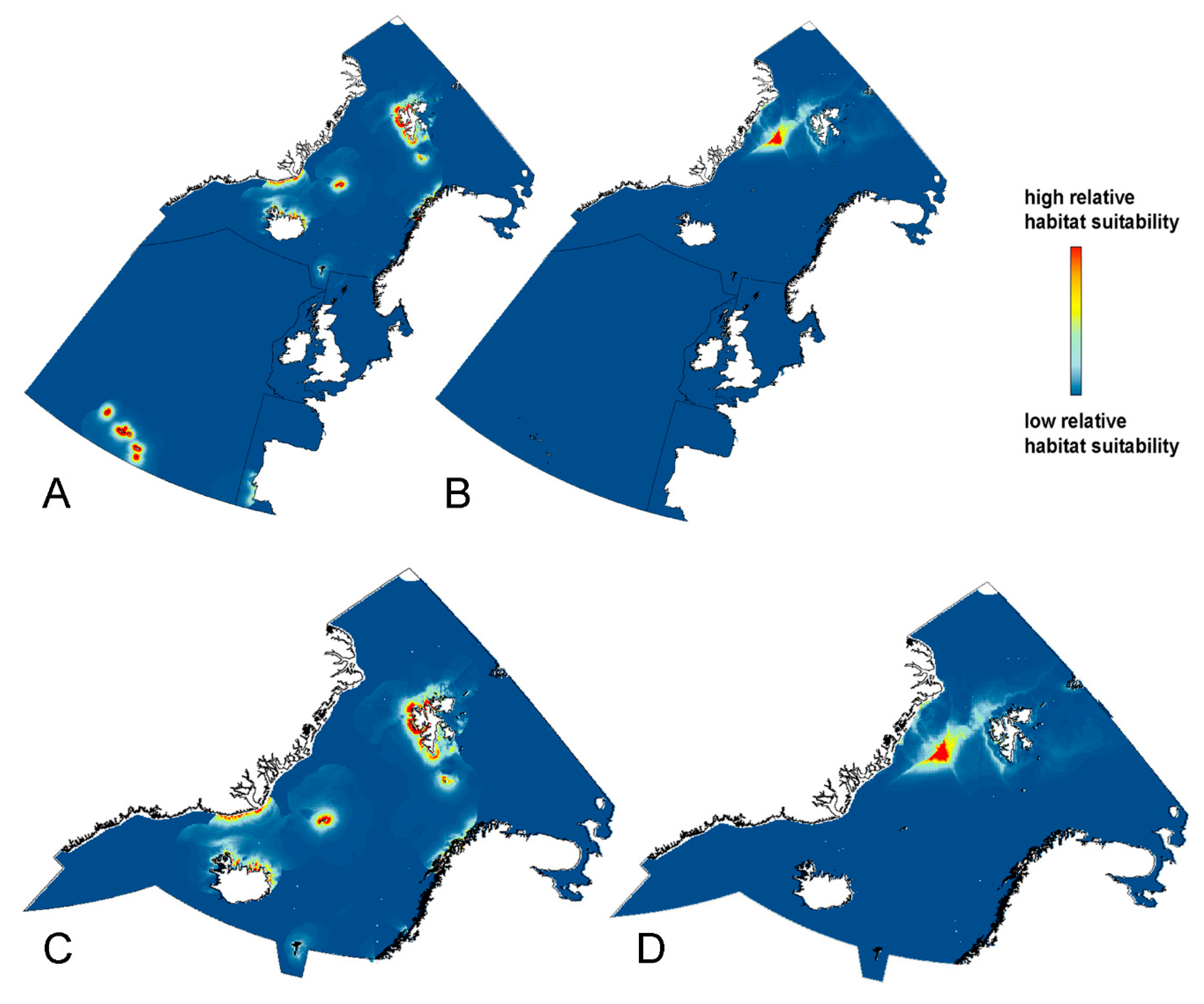
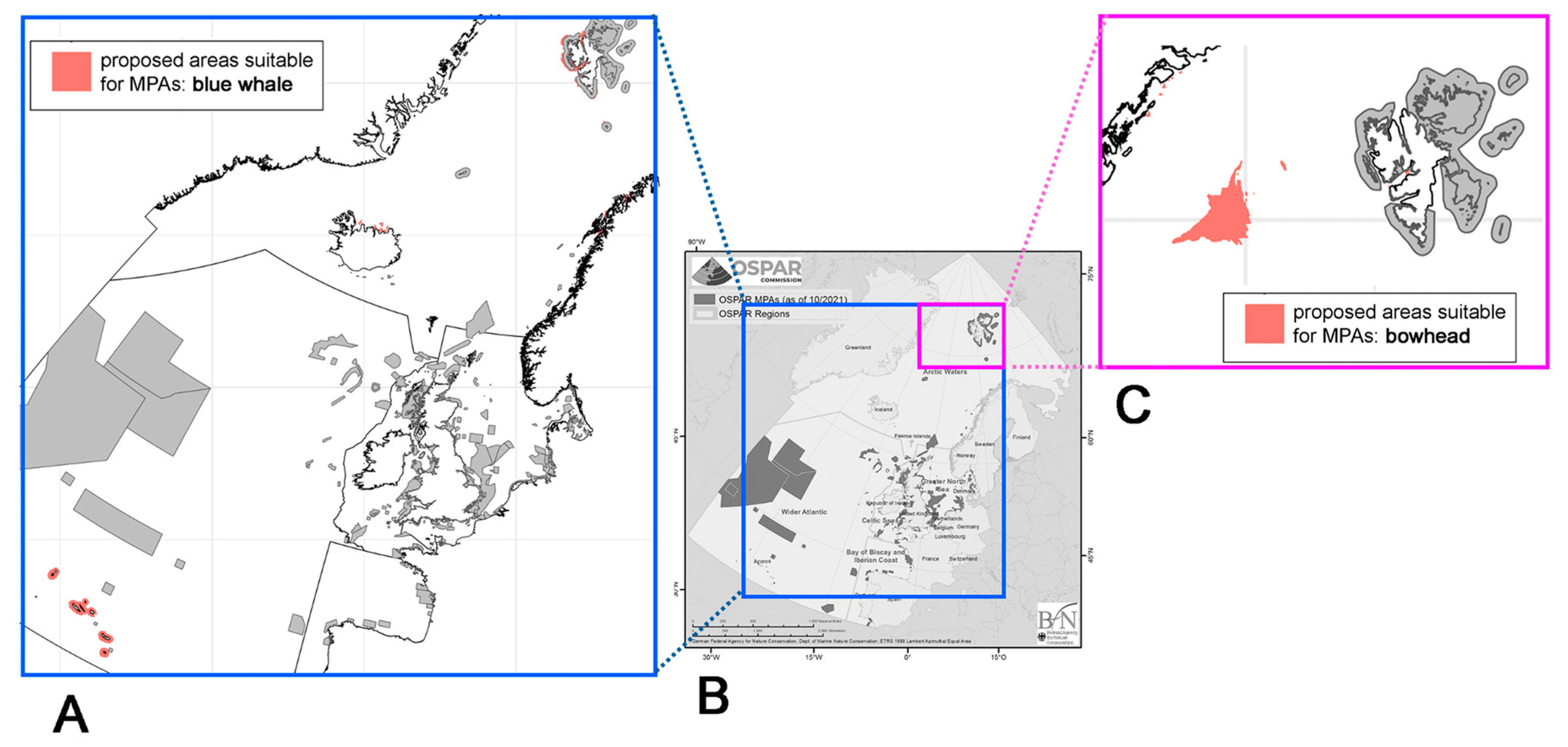
3. Results
3.1. Blue Whale (Balaenoptera musculus)
3.2. Bowhead Whale (Balaena mysticetus)
4. Discussion
5. Summary and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OSPAR, 2022. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic. OSPAR Commission. Available online: https://www.ospar.org/convention (accessed on 7 February 2022).
- Hooker, S.K.; Canadas, A.M.; Hyrenbach, K.D.; Corrigan, C.; Polovina, J.J.; Reeves, R.R. Making Protected Area Networks Effective for Marine Top Predators. Endanger. Species Res. 2011, 13, 203–218. [Google Scholar] [CrossRef]
- Hoyt, E. Marine Protected Areas for Whales, Dolphins and Porpoises: A World Handbook for Cetacean Habitat Conservation and Planning; Earthscan: Oxford, UK; Routledge: London, UK; Taylor & Francis: New York, NY, USA, 2011. [Google Scholar]
- Reeves, R.R. Dolphins, Whales, and Porpoises: 2002–2010 Conservation Action Plan for the World’S…: 2002–2010 Conservation Action Plan for the World’s Cetaceans, 2nd ed.; Union Internationale pour la Conservation de la Nature et de ses Ressources: Gland, Switzerland; Cambridge, UK, 2003; ISBN 978-2-8317-0656-6. [Google Scholar]
- OSPAR MPAs. Available online: http://mpa.ospar.org/home_ospar (accessed on 13 May 2023).
- OSPAR List of Threatened and/or Declining Species and Habitats. Available online: https://www.ospar.org/work-areas/bdc/species-habitats/list-of-threatened-declining-species-habitats (accessed on 23 February 2023).
- POSH Roadmap. Available online: https://www.ospar.org/news/implementing-collective-actions-on-threatened-and-or-declining-species (accessed on 23 June 2023).
- Mercker, M.; Markones, N.; Borkenhagen, K.; Schwemmer, H.; Wahl, J.; Garthe, S. An Integrated Framework to Estimate Seabird Population Numbers and Trends. J. Wildl. Manag. 2021, 85, 751–771. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Korner-Nievergelt, F.; Roth, T.; von Felten, S.; Guelat, J.; Almasi, B.; Korner-Nievergelt, P. Bayesian Data Analysis in Ecology Using Linear Models with R, BUGS, and Stan; Elsevier: London, UK, 2015. [Google Scholar]
- Wood, S. Generalized Additive Models: An Introduction with R; Chapman & Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]
- Zuur, A.; Ieno, E.; Smith, G.M. Analysing Ecological Data; Springer Science+Business Media, LLC: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Saveliev, A.A. Spatial, Temporal and Spatial-Temporal Ecological Data Analysis with R-INLA, Volume I–II; Highland Statistics Ltd.: Newburgh, UK, 2017. [Google Scholar]
- Gilles, A.; Authier, M.; Ramirez-Martinez, N.; Araújo, H.; Blanchard, A.; Carlström, J.; Eira, C.; Dorémus, G.; Fernández-Maldonado, C.; Geelhoed, S.C.V.; et al. Scans-IV. Small Cetaceans in Euopean Atlantic Water and the North Sea 2022. In Proceedings of the 34th Annual Conference of the European Cetacean Society, O Grove, Spain, 16–20 April 2023. [Google Scholar]
- Philipps, S.J.; Dudik, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar]
- Elith, J.; Hastie, T.; Dudik, M.; Chee, Y.; Yates, C.J.; Phillips, S.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar]
- Bost, C.A.; Cotté, C.; Bailleul, F.; Cherel, Y.; Charrassin, J.B.; Guinet, C.; Ainley, D.G.; Weimerskirch, H. The Importance of Oceanographic Fronts to Marine Birds and Mammals of the Southern Oceans. J. Mar. Syst. 2009, 78, 363–376. [Google Scholar] [CrossRef]
- Schneider, D.C. Seabirds and Fronts: A Brief Overview. Polar Res. 1990, 8, 17–21. [Google Scholar] [CrossRef]
- Renner, I.W.; Elith, J.; Fithian, W.; Hastie, T.; Phillips; Warton, D.I.; Baddeley, A.; Popovic, G. Point Process Models for Presence-Only Analysis. Methods Ecol. Evol. 2015, 6, 366–379. [Google Scholar]
- Renner, I.W.; Warton, D.I. Equivalence of MAXENT and Poisson Point Process Models for Species Distribution Modeling in Ecology. Biometrics 2013, 69, 274–281. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping Species Distributions with MAXENT Using a Geographically Biased Sample of Presence Data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Botella, C.; Joly, A.; Monestiez, P.; Bonnet, P.; Munoz, F. Bias in Presence-Only Niche Models Related to Sampling Effort and Species Niches: Lessons for Background Point Selection. PLoS ONE 2020, 15, e0232078. [Google Scholar] [CrossRef]
- Ready, J.; Kaschner, K.; South, A.B.; Eastwood, P.D.; Rees, T.; Rius, J.; Agbayani, E.; Kullander, S.; Froese, R. Predicting the Distributions of Marine Organisms at the Global Scale. Ecol. Model. 2010, 221, 467–478. [Google Scholar] [CrossRef]
- Vollering, J.; Halvorsen, R.; Mazzoni, S. The MIAmaxent R Package: Variable Transformation and Model Selection for Species Distribution Models. Ecol. Evol. 2019, 9, 12051–12068. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org (accessed on 5 February 2024).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 2.5-8. 2016. Available online: https://cran.r-project.org/web/packages/raster/raster.pdf (accessed on 15 March 2023).
- Hijmans, R.J.; van Etten, J. Raster: Geographic Analysis and Modeling with Raster Data. R Package Version 2.0-12. 2012. Available online: http://CRAN.R-project.org/package=raster (accessed on 15 March 2023).
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar]
- Baddeley, A.; Turner, R. spatstat: An R Package for Analyzing Spatial Point Patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar] [CrossRef]
- Grolemund, G.; Wickham, H. Dates and Times Made Easy with Lubridate. J. Stat. Softw. 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Arya, S.; Kemp, S.E.; Jefferis, G. Mount RANN: Fast Nearest Neighbour Search (Wraps ANN Library) Using L2 Metric 2019. Available online: https://jefferislab.r-universe.dev/RANN (accessed on 12 August 2024).
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef]
- Carwardine, M. Handbook of Whales, Doplhins, and Porpoises of the World; Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2020. [Google Scholar]
- OSPAR Biodiversity Committee Assessments—Status. 2020. Available online: https://oap.ospar.org/en/ospar-assessments/committee-assessments/biodiversity-committee/status-assesments/blue-whale/ (accessed on 15 March 2023).
- Silva, M.A.; Prieto, R.; Jonsen, I.; Baumgartner, M.F.; Santos, R.S. North Atlantic Blue and Fin Whales Suspend Their Spring Migration to Forage in Middle Latitudes: Building up Energy Reserves for the Journey? PLoS ONE 2013, 8, e76507. [Google Scholar] [CrossRef]
- Pike, D.G.; Víkingsson, G.A.; Gunnlaugsson, T.; Øien, N. A Note on the Distribution and Abundance of Blue Whales (Blalaenoptera musculus) in the Central and Northeast North Atlantic. NAMMCO Sci. Entific. Publ. 2009, 7. [Google Scholar] [CrossRef]
- Joiris, C.R. Considerable Increase in Bowhead, Blue, Humpback and Fin Whales Numbers in the Greenland Sea and Fram Strait between 1979 and 2014. Adv. Polar Sci. 2016, 2, 117–125. [Google Scholar]
- OSPAR Abundance and Distribution of Cetaceans—Sheet Reference: BDC15/D103 2018. Available online: https://oap.ospar.org/en/ospar-assessments/intermediate-assessment-2017/biodiversity-status/marine-mammals/abundance-distribution-cetaceans/abundance-and-distribution-cetaceans/ (accessed on 11 August 2024).
- Reid, J.B.; Evans, P.G.H.; Northridge, S.P. Atlas of Cetacean: Distribution in North-West European Waters; Joint Nature Conservation Committee: Peterborough, UK, 2003.
- Wall, D.; O’kelly, I.; Whooley, P.; Tyndall, P. New records of blue whales (Balaenoptera musculus) with evidence of possible feeding behaviour from the continental shelf slopes to the west of Ireland. Mar. Biodivers. Rec. 2009, 2, e128. [Google Scholar] [CrossRef]
- Visser, F.; Hartman, K.L.; Pierce, G.J.; Valavanis, V.D.; Huisman, J. Timing of Migratory Baleen Whales at the Azores in Relation to the North Atlantic Spring Bloom. Mar. Ecol. Prog. Ser. 2011, 440, 267–279. [Google Scholar] [CrossRef]
- Storrie, L.; Lydersen, C.; Andersen, M.; Wynn, R.B.; Kovacs, K.M. Determining the Species Assemblage and Habitat Use of Cetaceans in the Svalbard Archipelago, Based on Observations from 2002 to 2014. Polar Res. 2018, 37, 1463065. [Google Scholar] [CrossRef]
- Gilles, A.; Viquerat, S.; Becker, A.A.; Forney, K.A.; Geelhoed, S.C.V.; Helters, J.; Nabe-Nielsen, J.; Scheidat, M.; Siebert, U.; Sveegard, S.; et al. Seasonal habitat-based density models for a marine top predator, the harbor porpoise, in a dynamic environment. Ecosphere 2016, 7, e01367. [Google Scholar]
- Vacquié-Garcia, J.; Lydersen, C.; Marques, T.A.; Aars, J.; Ahonen, H.; Skern-Mauritzen, M.; Øien, N.; Kovacs, K.M. Late Summer Distribution and Abundance of Ice-Associated Whales in the Norwegian High Arctic. Endanger. Species Res. 2017, 32, 59–70. [Google Scholar] [CrossRef]
- Shirihai, H.; Jarret, B. Whales, Dolphins And Seals: A Field Guide to the Marine Mammals of the World; A & C Black: London, UK, 2006. [Google Scholar]
- Reeves, R.R.; Ewins, P.J.; Agbayani, S.; Heide-Jørgensen, M.P.; Kovacs, K.M.; Lydersen, C.; Suydam, R.; Elliott, W.; Polet, G.; van Dijk, Y.; et al. Distribution of Endemic Cetaceans in Relation to Hydrocarbon Development and Commercial Shipping in a Warming Arctic. Mar. Policy 2014, 44, 375–389. [Google Scholar] [CrossRef]
- de Boer, M.N.; Jones, D.; Jones, H. Ocean Wanderers: Extralimital Encounters with Bowhead Whales (Balaena mysticetus) in Temperate European Shallow Waters. Aquat. Mamm. 2017, 43, 279–288. [Google Scholar] [CrossRef]
- de Boer, M.N.; Janinhoff, N.; Nijs, G.; Verdaat, H. Encouraging Encounters: Unusual Aggregations of Bowhead Whales Balaena mysticetus in the Western Fram Strait. Endanger. Species Res. 2019, 39, 51–62. [Google Scholar] [CrossRef]
- Wiig, Ø.; Bachmann, L.; Kovacs, K.; Swift, R.; Lydersen, C. Survey of Bowhead Whales (Balaena mysticetus) in the Northeast Atlantic in 2010. 2010. Available online: https://www.researchgate.net/publication/242579602_Survey_of_bowhead_whales_Balaena_mysticetus_in_the_Northeast_Atlantic_in_2010 (accessed on 15 March 2023).
- Kovacs, K.M.; Lydersen, C.; Vacquiè-Garcia, J.; Shpak, O.; Glazov, D.; Heide-Jørgensen, M.P. The Endangered Spitsbergen Bowhead Whales’ Secrets Revealed after Hundreds of Years in Hiding. Biol. Lett. 2020, 16, 20200148. [Google Scholar] [CrossRef]
- Boertmann, D.; Kyhn, L.A.; Witting, L.; Heide-Jørgensen, M.P. A hidden getaway for bowhead whales in the Greenland Sea. Polar Biol. 2015, 38, 1315–1319. [Google Scholar] [CrossRef]
- Boertmann, D.; Merkel, F.; Durinck, J. Bowhead Whales in East Greenland, Summers 2006–2008. Polar Biol. 2009, 32, 1805–1809. [Google Scholar] [CrossRef]
- Boertmann, D.; Nielsen, R.D. A bowhead whale calf observed in northeast Greenland waters. Polar Rec. 2010, 46, 373–375. [Google Scholar] [CrossRef]
- Gilg, O.; Born, E.W. Recent Sightings of the Bowhead Whale (Balaena mysticetus) in Northeast Greenland and the Greenland Sea. Polar Biol. 2005, 28, 796–801. [Google Scholar] [CrossRef]
- Lee, H.; Irvine, A.; Brito-Morales, I.; Fuller, S.; Davies, T.; Tittensor, D.; Reville, G.; Shackell, N.; Hennicke, J.; Stanley, R. To Save the High Seas, Plan for Climate Change. Nature 2024, 630, 298–301. [Google Scholar]
- ESAS. Available online: https://www.ices.dk/data/data-portals/Pages/European-Seabirds-at-sea.aspx (accessed on 5 June 2022).
- SCANS-III. Available online: https://scans3.wp.st-andrews.ac.uk/ (accessed on 1 May 2024).
- GBIF. Available online: https://www.gbif.org/ (accessed on 1 May 2024).
- Norwegian Biodiversity Centre & Polar Institute: Arctic Data. Available online: https://www.npolar.no/ (accessed on 1 March 2023).
- Kovacs, K.M.; Andersen, M. Marine Mammals Sightings in and around Svalbard 1995–2019 [Data Set]. Available online: https://data.npolar.no/dataset/246c1053-6fdc-5043-b7b4-284f4fa8c095 (accessed on 11 August 2024).
- Artsdatenbanken/Norwegian Biodiversity Centre Map Service. Available online: https://artsdatabanken.no/ (accessed on 3 May 2022).
- OBIS. Available online: https://obis.org/ (accessed on 3 June 2022).
- Vertnet. Available online: http://vertnet.org/ (accessed on 1 June 2022).
- RusMam. Available online: https://rusmam.ru/atlas/map (accessed on 3 May 2022).
- OSPAR Case Reports for the OSPAR List of Threatened and/or Declining Species and Habitats. 2008. Available online: https://qsr2010.ospar.org/media/assessments/p00358_case_reports_species_and_habitats_2008.pdf (accessed on 11 August 2024).
- OSPAR Background Document for Blue Whale: Balaenoptera musculus. 2010. Available online: https://www.ospar.org/documents?d=7232 (accessed on 11 August 2024).
- Lesage, V.; Gosselin, J.-F.; Lawson, J.W.; McQuinn, I.; Moors-Murphy, H.; Plourde, S.; Sears, R.; Simard, Y. Habitats Important to Blue Whales (Balaenoptera musculus) in the Western North Atlantic; Canadian Science Advisory Secretariat (CSAS): Ottawa, ON, USA, 2018; pp. 1–56. [Google Scholar]
- Barlow, D.R.; Torres, L.G. Planning Ahead: Dynamic Models Forecast Blue Whale Distribution with Applications for Spatial Management. J. Appl. Ecol. 2021, 58, 2493–2504. [Google Scholar] [CrossRef]
- George, J.C.; Theweissen, J.G.M. The Bowhead Whale: Balaena mysticetus: Biology and Human Interactions; Academic Press Inc.: London, UK, 2021. [Google Scholar]
- Citta, J.J.; Quakenbush, L.T.; Okkonen, S.R.; Druckenmiller, M.L.; Maslowski, W.; Clement-Kinney, J.; George, J.C.; Brower, H.; Small, R.J.; Ashjian, C.J.; et al. Ecological Characteristics of Core-Use Areas Used by Bering–Chukchi–Beaufort (BCB) Bowhead Whales, 2006–2012. Prog. Oceanogr. 2015, 136, 201–222. [Google Scholar] [CrossRef]
- Harwood, L.A.; Quakenbush, L.T.; Small, R.J.; George, J.C.; Pokiak, J.; Pokiak, C.; Heide-Jørgensen, M.P.; Lea, E.V.; Brower, H. Movements and Inferred Foraging by Bowhead Whales in the Canadian Beaufort Sea during August and September, 2006–12. Arctic 2017, 70, 161–176. [Google Scholar] [CrossRef]
- Citta, J.J.; Okkonen, S.R.; Quakenbush, L.T.; Maslowski, W.; Osinski, R.; George, J.C.; Small, R.J.; Brower, H.; Heide-Jørgensen, M.P.; Harwood, L.A. Oceanographic Characteristics Associated with Autumn Movements of Bowhead Whales in the Chukchi Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2018, 152, 121–131. [Google Scholar] [CrossRef]
- Chambault, P.; Albertsen, C.M.; Patterson, T.A.; Hansen, R.G.; Tervo, O.; Laidre, K.L.; Heide-Jørgensen, M.P. Sea Surface Temperature Predicts the Movements of an Arctic Cetacean: The Bowhead Whale. Sci. Rep. 2018, 8, 9658. [Google Scholar] [CrossRef]
- CLS Maritime Intelligence. Available online: https://maritime-intelligence.groupcls.com/ (accessed on 1 June 2023).
- NASA Ocean Color Web—Distcoast. Available online: https://oceancolor.gsfc.nasa.gov/resources/docs/distfromcoast/ (accessed on 2 June 2022).
- NASA AQUA MODIS Web. Available online: https://neo.gsfc.nasa.gov/view.php?datasetId=MYD28M (accessed on 6 August 2022).
- Marine Conservation Institute (MCI). Available online: https://www.arcgis.com/home/item.html?id=ab0926890e444fd0a2ecd4f40fb318f7 (accessed on 8 August 2023).
- NASA Ocean Color Web-Chorophyll a. Available online: https://oceancolor.gsfc.nasa.gov/resources/atbd/chlor_a/ (accessed on 8 August 2022).
- GEBCO Compilation Group (2020) GEBCO 2020 Grid. Doi:10.5285/A29c5465-B138-234d-E053-6c86abc040b9. Available online: https://www.gebco.net/data_and_products/gridded_bathymetry_data/ (accessed on 8 August 2022).
- Global SMOS Level 4 Sea Surface Salinity File (Institution: Barcelona Expert Center (BEC)). Available online: https://erddap.emodnet.eu/erddap/index.html (accessed on 7 August 2022).
- National Snow and Ice Data Center Respectively NOAA. Available online: https://nsidc.org/data/seaice_index (accessed on 5 August 2022).
- Yi, D.; Zwally, H.J. Arctic Sea Ice Freeboard and Thickness; Version 1; Updated 2014-04-15; NASA National Snow and Ice Data Center Distributed Active Archive Center: Boulder, CO, USA, 2009. [Google Scholar]
- Swedish National Data Service (SND). Dataset. (Obst, M. (2017). Global Tidal Variables (1.0). University of Gothenburg. Available online: https://snd.gu.se/en/catalogue/study/ecds0243# (accessed on 5 August 2022).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercker, M.; Müller, M.; Werner, T.; Hennicke, J. Identification of Key Habitats of Bowhead and Blue Whales in the OSPAR Area of the North-East Atlantic—A Modelling Approach towards Effective Conservation. J. Mar. Sci. Eng. 2024, 12, 1445. https://doi.org/10.3390/jmse12081445
Mercker M, Müller M, Werner T, Hennicke J. Identification of Key Habitats of Bowhead and Blue Whales in the OSPAR Area of the North-East Atlantic—A Modelling Approach towards Effective Conservation. Journal of Marine Science and Engineering. 2024; 12(8):1445. https://doi.org/10.3390/jmse12081445
Chicago/Turabian StyleMercker, Moritz, Miriam Müller, Thorsten Werner, and Janos Hennicke. 2024. "Identification of Key Habitats of Bowhead and Blue Whales in the OSPAR Area of the North-East Atlantic—A Modelling Approach towards Effective Conservation" Journal of Marine Science and Engineering 12, no. 8: 1445. https://doi.org/10.3390/jmse12081445