
Novel Chitosan-Gelatin Scaffold with Valproic Acid Augments In Vitro Osteoblast Differentiation of Mesenchymal Stem Cells

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Fabrication of Chitosan-Gelatin Scaffolds
2.3. Preparation of VPA
2.4. VPA Loading and Incubation
2.5. Determination of VPA Releases from CH-G Scaffold
2.6. Cell Culture and Seeding
2.7. Cell Viability and Proliferation
2.8. Live/Dead Staining
2.9. Characterization of hBMSCs in CH-G Scaffolds by Scanning Electron Microscopy
2.10. Alkaline Phosphatase (ALP) Quantification Assay and Staining
2.11. Alizarin Red-S Staining Assay
2.12. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) for Gene Analysis
2.13. Statistical Analysis
3. Results
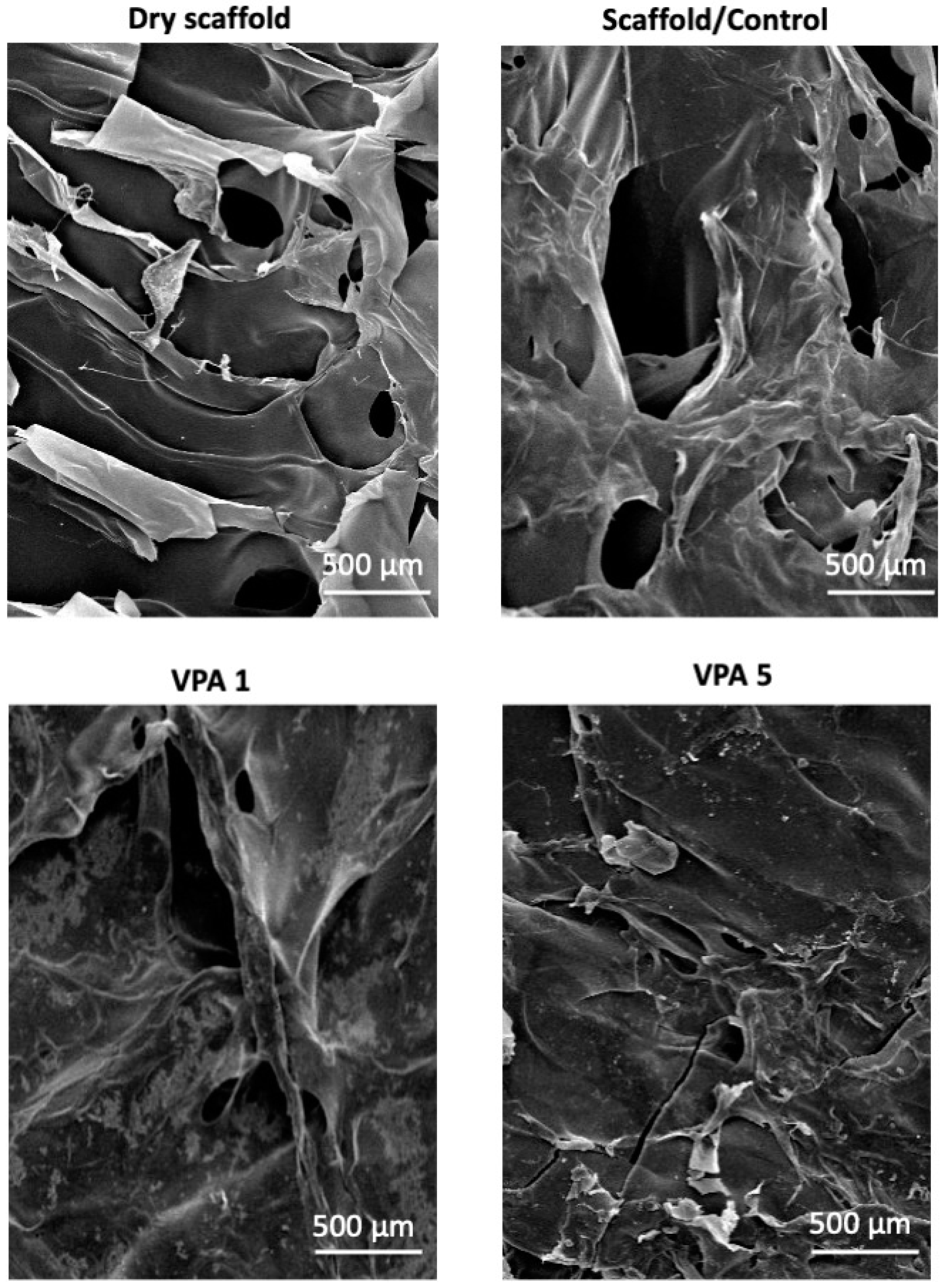
3.1. CH-G Scaffold Porosity and Morphology
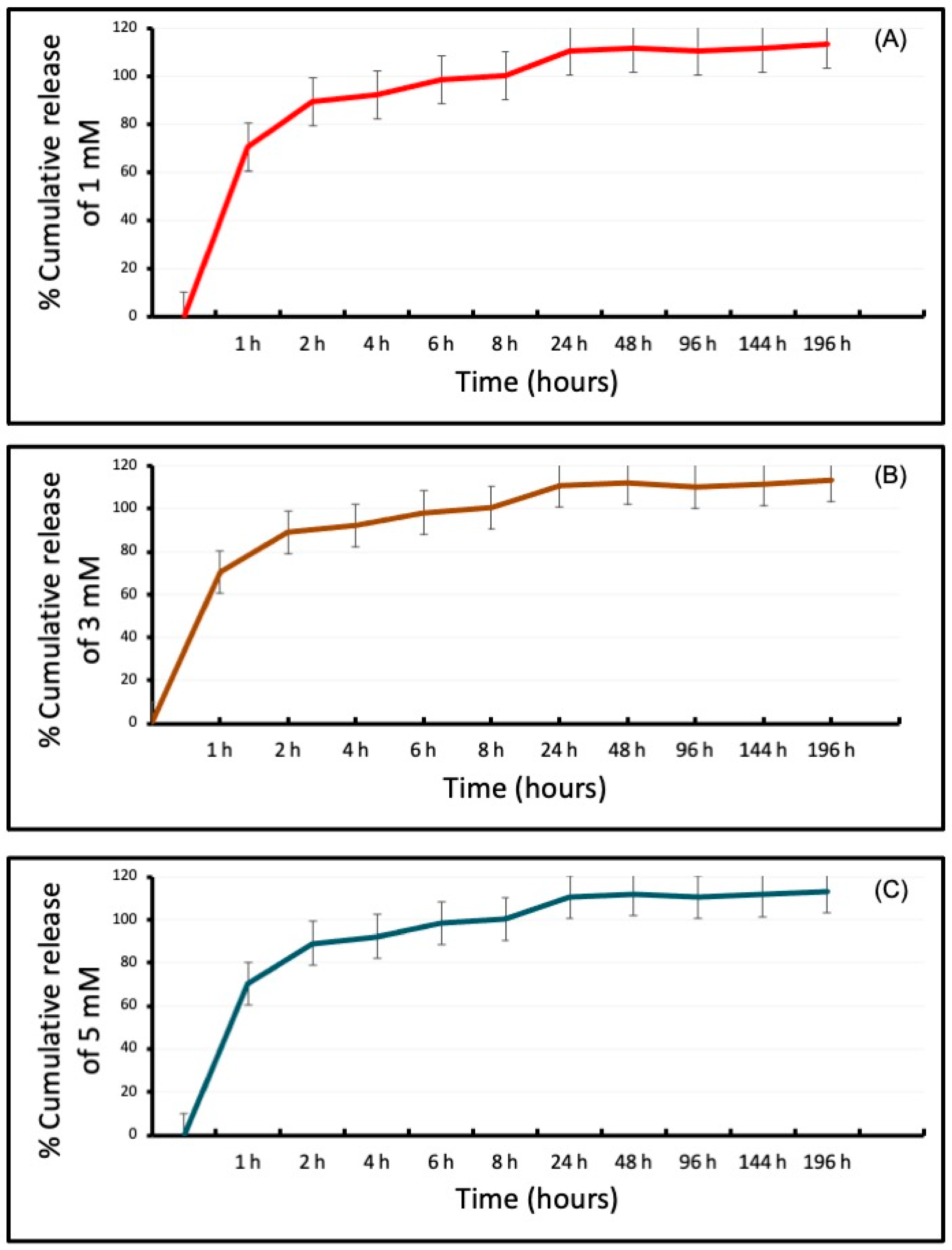
3.2. Differential Release of VPA from the CH-G Scaffold
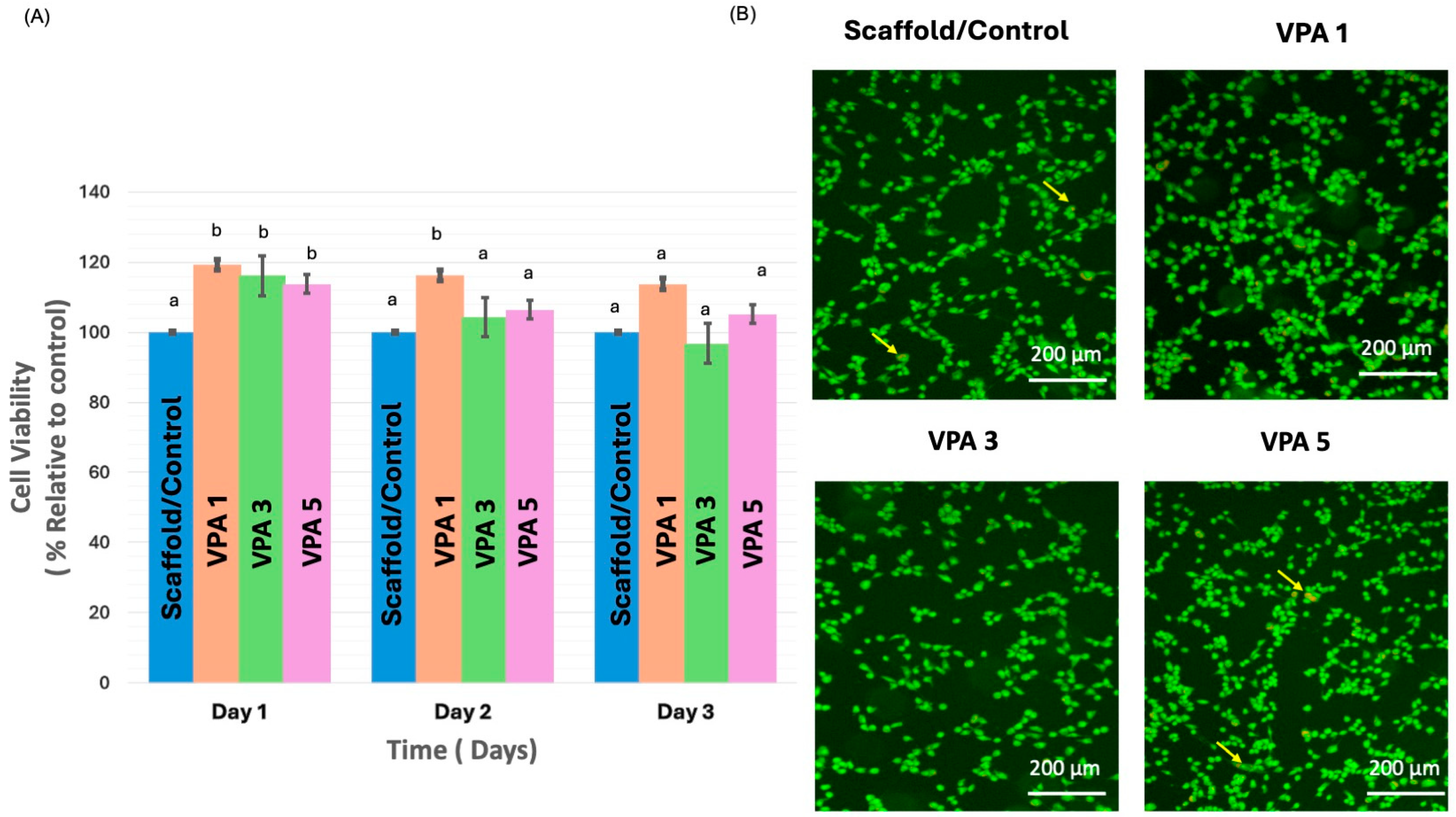
3.3. VPA Enhances Cell Proliferation and Improves Viability
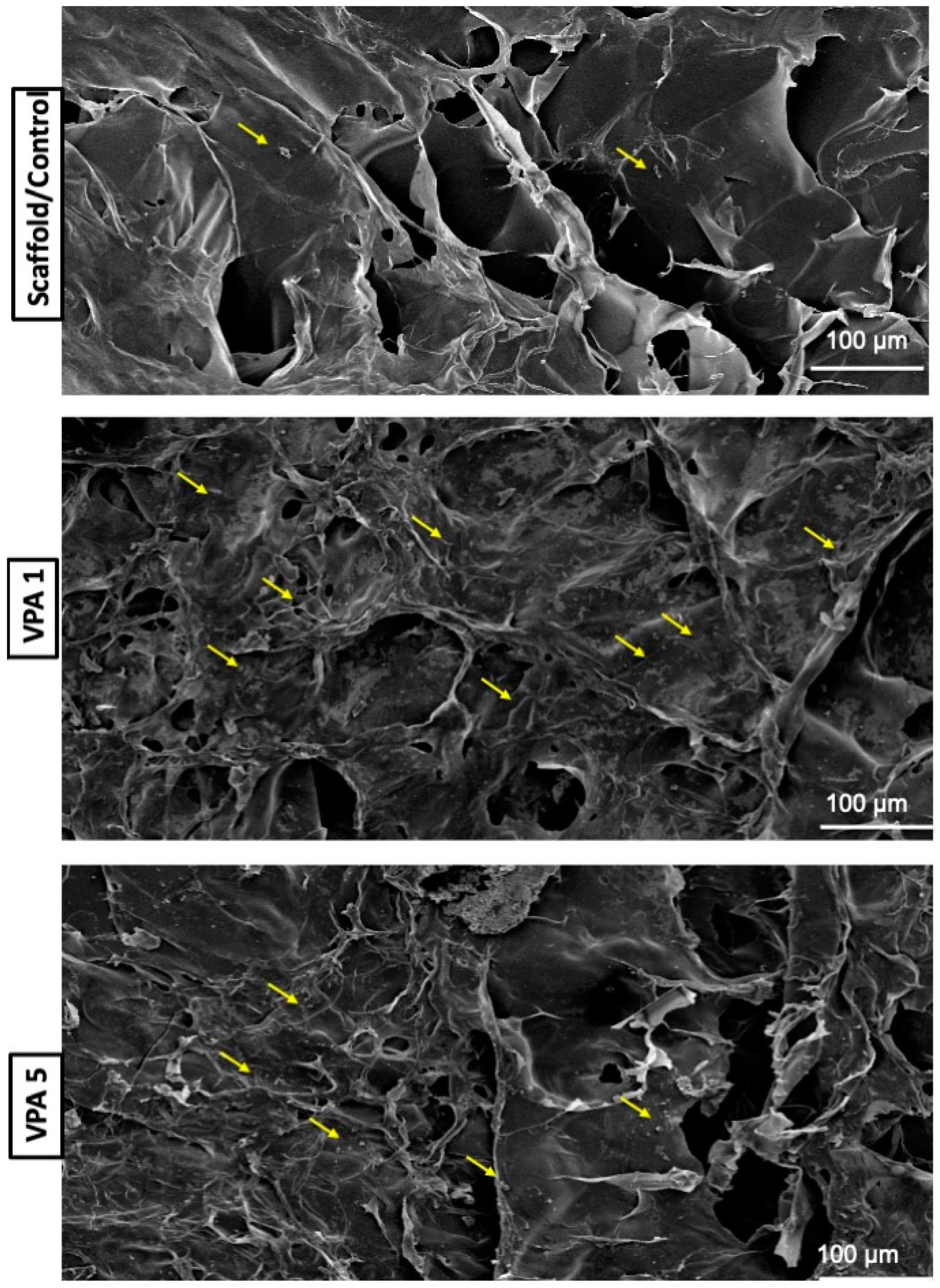
3.4. Implications of VPA on Cell Morphology
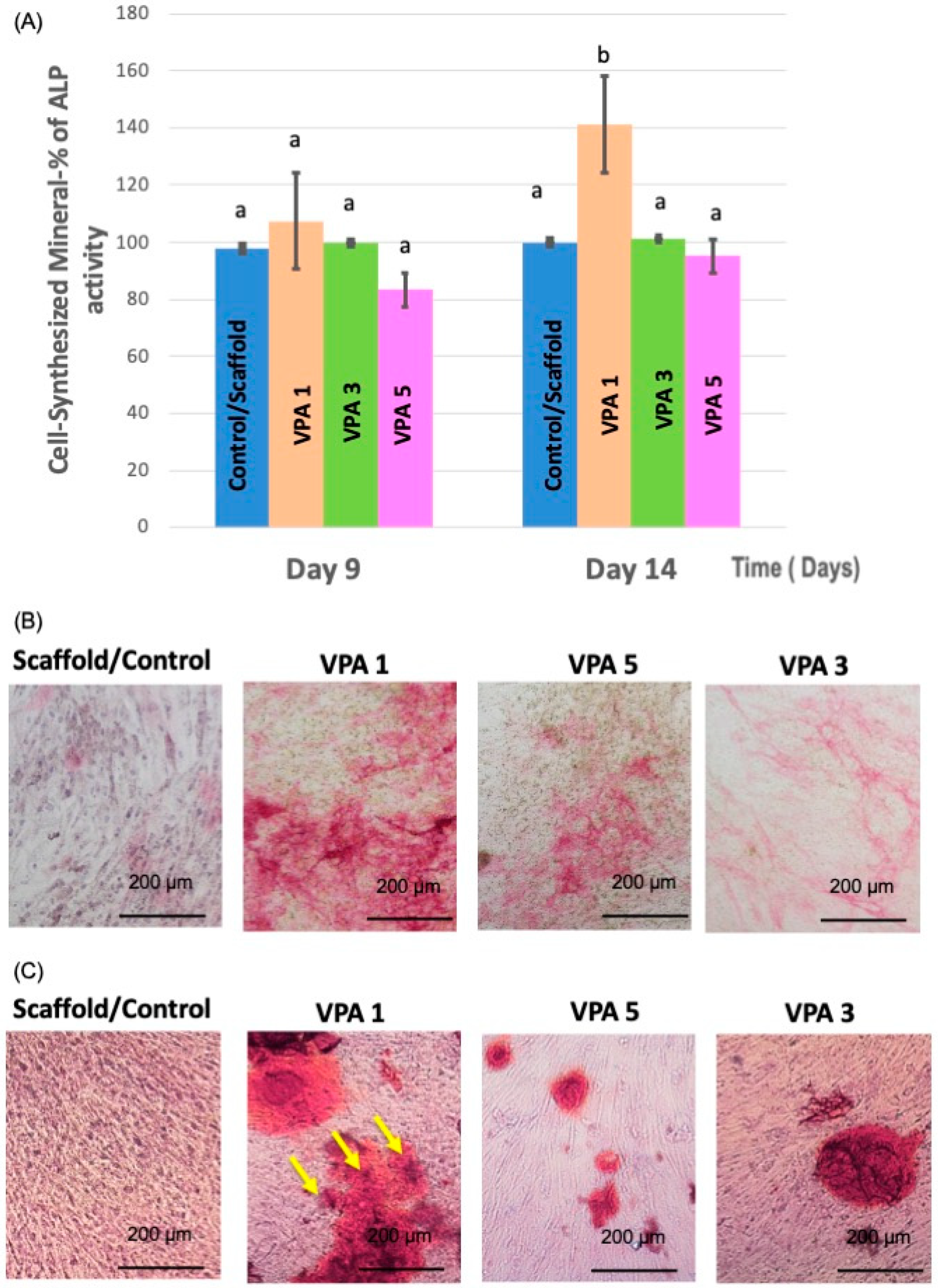
3.5. VPA Enhances Osteoblast Differentiation
3.6. VPA Modulates Osteogenic Gene Expression in hBMSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teven, C.M.; Liu, X.; Hu, N.; Tang, N.; Kim, S.H.; Huang, E.; Yang, K.; Li, M.; Gao, J.L.; Liu, H.; et al. Epigenetic regulation of mesenchymal stem cells: A focus on osteogenic and adipogenic differentiation. Stem Cells Int. 2011, 2011, 201371. [Google Scholar] [CrossRef]
- Man, K.; Lawlor, L.; Jiang, L.H.; Yang, X.B. The Selective Histone Deacetylase Inhibitor MI192 Enhances the Osteogenic Differentiation Efficacy of Human Dental Pulp Stromal Cells. Int. J. Mol. Sci. 2021, 22, 5224. [Google Scholar] [CrossRef]
- Hussain, A.; Tebyaniyan, H.; Khayatan, D. The Role of Epigenetic in Dental and Oral Regenerative Medicine by Different Types of Dental Stem Cells: A Comprehensive Overview. Stem Cells Int. 2022, 2022, 5304860. [Google Scholar] [CrossRef] [PubMed]
- Jamaladdin, S.; Kelly, R.D.; O’Regan, L.; Dovey, O.M.; Hodson, G.E.; Millard, C.J.; Portolano, N.; Fry, A.M.; Schwabe, J.W.; Cowley, S.M. Histone deacetylase (HDAC) 1 and 2 are essential for accurate cell division and the pluripotency of embryonic stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 9840–9845. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yin, S.; Zeng, H.; Li, H.; Wan, X. Regulation of embryonic stem cell self-renewal. Life 2022, 12, 1151. [Google Scholar] [CrossRef]
- Zhao, Q.; Ji, K.; Wang, T.; Li, G.; Lu, W.; Ji, J. Effect of the Histone Deacetylases Inhibitors on the Differentiation of Stem Cells in Bone Damage Repairing and Regeneration. Curr. Stem Cell Res. Ther. 2020, 15, 24–31. [Google Scholar] [CrossRef]
- Jin, H.; Park, J.Y.; Choi, H.; Choung, P.H. HDAC inhibitor trichostatin A promotes proliferation and odontoblast differentiation of human dental pulp stem cells. Tissue Eng. Part A 2013, 19, 613–624. [Google Scholar] [CrossRef]
- Duncan, H.F.; Smith, A.J.; Fleming, G.J.; Cooper, P.R. Histone deacetylase inhibitors induced differentiation and accelerated mineralization of pulp-derived cells. J. Endod. 2012, 38, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Shnayder, N.A.; Grechkina, V.V.; Khasanova, A.K.; Bochanova, E.N.; Dontceva, E.A.; Petrova, M.M.; Asadullin, A.R.; Shipulin, G.A.; Altynbekov, K.S.; Al-Zamil, M.; et al. Therapeutic and Toxic Effects of Valproic Acid Metabolites. Metabolites 2023, 13, 134. [Google Scholar] [CrossRef]
- Angus-Leppan, H.; Arkell, R.; Watkins, L.; Heaney, D.; Cooper, P.; Shankar, R. New valproate regulations, informed choice and seizure risk. J. Neurol. 2024, 271, 5671–5686. [Google Scholar] [CrossRef]
- Bug, G.; Gül, H.; Schwarz, K.; Pfeifer, H.; Kampfmann, M.; Zheng, X.; Beissert, T.; Boehrer, S.; Hoelzer, D.; Ottmann, O.G.; et al. Valproic acid stimulates proliferation and self-renewal of hematopoietic stem cells. Cancer Res. 2005, 65, 2537–2541. [Google Scholar] [CrossRef]
- Monti, B.; Polazzi, E.; Contestabile, A. Biochemical, molecular and epigenetic mechanisms of valproic acid neuroprotection. Curr. Mol. Pharmacol. 2009, 2, 95–109. [Google Scholar] [CrossRef]
- Akshaya, N.; Prasith, P.; Abinaya, B.; Ashwin, B.; Chandran, S.V.; Selvamurugan, N. Valproic acid, A Potential Inducer of Osteogenesis in Mouse Mesenchymal Stem Cells. Curr. Mol. Pharmacol. 2021, 14, 27–35. [Google Scholar] [CrossRef]
- Cho, H.H.; Park, H.T.; Kim, Y.J.; Bae, Y.C.; Suh, K.T.; Jung, J.S. Induction of osteogenic differentiation of human mesenchymal stem cells by histone deacetylase inhibitors. J. Cell. Biochem. 2005, 96, 533–542. [Google Scholar] [CrossRef]
- Sharma, P.K.; Halder, M.; Srivastava, U.; Singh, Y. Antibacterial PEG-Chitosan Hydrogels for Controlled Antibiotic/Protein Delivery. ACS Appl. Bio Mater. 2019, 2, 5313–5322. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, C.; Shrestha, S.; Torneck, C.; Kishen, A. Effects of a Bioactive Scaffold Containing a Sustained Transforming Growth Factor-β1-releasing Nanoparticle System on the Migration and Differentiation of Stem Cells from the Apical Papilla. J. Endod. 2016, 42, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Alghofaily, M.; Almana, A.; Alrayes, J.; Lambarte, R.; Weir, M.D.; Alsalleeh, F. Chitosan–Gelatin Scaffolds Loaded with Different Antibiotic Formulations for Regenerative Endodontic Procedures Promote Biocompatibility and Antibacterial Activity. J. Funct. Biomater. 2024, 15, 186. [Google Scholar] [CrossRef] [PubMed]
- Thein-Han, W.W.; Saikhun, J.; Pholpramoo, C.; Misra, R.D.; Kitiyanant, Y. Chitosan-gelatin scaffolds for tissue engineering: Physico-chemical properties and biological response of buffalo embryonic stem cells and transfectant of GFP-buffalo embryonic stem cells. Acta Biomater. 2009, 5, 3453–3466. [Google Scholar] [CrossRef] [PubMed]
- Duncan, H.F.; Smith, A.J.; Fleming, G.J.; Cooper, P.R. Histone deacetylase inhibitors epigenetically promote reparative events in primary dental pulp cells. Exp. Cell Res. 2013, 319, 1534–1543. [Google Scholar] [CrossRef]
- Abdallah, B.M.; Haack-Sørensen, M.; Burns, J.S.; Elsnab, B.; Jakob, F.; Hokland, P.; Kassem, M. Maintenance of differentiation potential of human bone marrow mesenchymal stem cells immortalized by human telomerase reverse transcriptase gene despite [corrected] extensive proliferation. Biochem. Biophys. Res. Commun. 2005, 326, 527–538. [Google Scholar] [CrossRef]
- Simonsen, J.L.; Rosada, C.; Serakinci, N.; Justesen, J.; Stenderup, K.; Rattan, S.I.; Jensen, T.G.; Kassem, M. Telomerase expression extends the proliferative life-span and maintains the osteogenic potential of human bone marrow stromal cells. Nat. Biotechnol. 2002, 20, 592–596. [Google Scholar] [CrossRef]
- Manikandan, M.; Abuelreich, S.; Elsafadi, M.; Alsalman, H.; Almalak, H.; Siyal, A.; Hashmi, J.A.; Aldahmash, A.; Kassem, M.; Alfayez, M.; et al. NR2F1 mediated down-regulation of osteoblast differentiation was rescued by bone morphogenetic protein-2 (BMP-2) in human MSC. Differentiation 2018, 104, 36–41. [Google Scholar] [CrossRef]
- Yarahmadi, A.; Dousti, B.; Karami-Khorramabadi, M.; Afkhami, H. Materials based on biodegradable polymers chitosan/gelatin: A review of potential applications. Front. Bioeng. Biotechnol. 2024, 12, 1397668. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Deng, J.; Yang, F.; Gong, Y.; Zhao, N.; Zhang, X. Study on physical properties and nerve cell affinity of composite films from chitosan and gelatin solutions. Biomaterials 2003, 24, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Sendemir-Urkmez, A.; Jamison, R.D. The addition of biphasic calcium phosphate to porous chitosan scaffolds enhances bone tissue development in vitro. J. Biomed. Mater. Res. A 2007, 81, 624–633. [Google Scholar] [CrossRef]
- Hardy, I.J.; Windberg-Baarup, A.; Neri, C.; Byway, P.V.; Booth, S.W.; Fitzpatrick, S. Modulation of drug release kinetics from hydroxypropyl methyl cellulose matrix tablets using polyvinyl pyrrolidone. Int. J. Pharm. 2007, 337, 246–253. [Google Scholar] [CrossRef]
- Wang, Y.; Pei, Y.A.; Sun, Y.; Zhou, S.; Zhang, X.-B.; Pei, M. Stem cells immortalized by hTERT perform differently from those immortalized by SV40LT in proliferation, differentiation, and reconstruction of matrix microenvironment. Acta Biomater. 2021, 136, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Narai, T.; Watase, R.; Nakayama, Y.; Kodani, I.; Inoue, T.; Kokura, K. Establishment of human immortalized mesenchymal stem cells lines for the monitoring and analysis of osteogenic differentiation in living cells. Heliyon 2020, 6, e05398. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Suh, J.H.; Kim, A.Y.; Lee, Y.S.; Park, S.Y.; Kim, J.B. Histone deacetylase 1-mediated histone modification regulates osteoblast differentiation. Mol. Endocrinol. 2006, 20, 2432–2443. [Google Scholar] [CrossRef]
- Rashid, S.; Qazi, R.E.; Malick, T.S.; Salim, A.; Khan, I.; Ilyas, A.; Haneef, K. Effect of valproic acid on the hepatic differentiation of mesenchymal stem cells in 2D and 3D microenvironments. Mol. Cell. Biochem. 2021, 476, 909–919. [Google Scholar] [CrossRef]
- de Boer, J.; Licht, R.; Bongers, M.; van der Klundert, T.; Arends, R.; van Blitterswijk, C. Inhibition of histone acetylation as a tool in bone tissue engineering. Tissue Eng. 2006, 12, 2927–2937. [Google Scholar] [CrossRef]
- Schroeder, T.M.; Westendorf, J.J. Histone deacetylase inhibitors promote osteoblast maturation. J. Bone Miner. Res. 2005, 20, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Kwon, A.; Park, H.J.; Baek, K.; Lee, H.L.; Park, J.C.; Woo, K.M.; Ryoo, H.M.; Baek, J.H. Suberoylanilide hydroxamic acid enhances odontoblast differentiation. J. Dental. Res. 2012, 91, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Uehara, O.; Yoshida, K.; Saito, T.; Abiko, Y. The histone deacetylase inhibitor, entinostat (MS-275), induces the odontogenic differentiation of an odontoblast-like cell line in the absence of an osteoblast mineralization medium. Odontology 2021, 109, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, J.R.; Seo, M.S.; Roh, K.H.; Park, S.B.; Hwang, J.W.; Sun, B.; Seo, K.; Lee, Y.S.; Kang, S.K.; et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009, 42, 711–720. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward | Reverse |
|---|---|---|
| Alkaline phosphate | 5′-GGAACTCCTGACCCTTGACC-3′ | 5′-TCCTGTTCAGCTCGTACTGC-3′ |
| Osteocalcin | 5′-GGCAGCGAGGTAGTGAAGAG-3′ | 5′-CTCACACACCTCCCTCCTG-3′ |
| Runx-2 | 5′-GTA GAT GGA CCT CGG GAA CC3′ | 5′-GAG GCG GTC AGA GAA CAA AC-3′ |
| Osteonectin | 5′-GAGGAAACCGAAGAGGAGG-3′ | 5-GGGGTGTTGTTCTCATCCAG-3′ |
| Collagen Type 1 | 5′-GAGTGCTGTCCCGTCTGC-3′ | 5-TTTCTTGGTCGGTGGGTG-3′ |
| GAPDH | 5′-GAAGGTGAAGGTCGGAGT-3′ | 5′-GAAGATGGTGATGGGATTTC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alghofaily, M.; Alsalleeh, F.; Alssum, L.; Muthurangan, M.; Alfayez, M.; Weir, M.D.; Xu, H.H.K. Novel Chitosan-Gelatin Scaffold with Valproic Acid Augments In Vitro Osteoblast Differentiation of Mesenchymal Stem Cells. J. Funct. Biomater. 2024, 15, 252. https://doi.org/10.3390/jfb15090252
Alghofaily M, Alsalleeh F, Alssum L, Muthurangan M, Alfayez M, Weir MD, Xu HHK. Novel Chitosan-Gelatin Scaffold with Valproic Acid Augments In Vitro Osteoblast Differentiation of Mesenchymal Stem Cells. Journal of Functional Biomaterials. 2024; 15(9):252. https://doi.org/10.3390/jfb15090252
Chicago/Turabian StyleAlghofaily, Maha, Fahd Alsalleeh, Lamees Alssum, Manikandan Muthurangan, Musaad Alfayez, Michael D. Weir, and Hockin H. K. Xu. 2024. "Novel Chitosan-Gelatin Scaffold with Valproic Acid Augments In Vitro Osteoblast Differentiation of Mesenchymal Stem Cells" Journal of Functional Biomaterials 15, no. 9: 252. https://doi.org/10.3390/jfb15090252