
Recent Advances in Monitoring Stem Cell Status and Differentiation Using Nano-Biosensing Technologies


 ,
,  and
and
Abstract
:
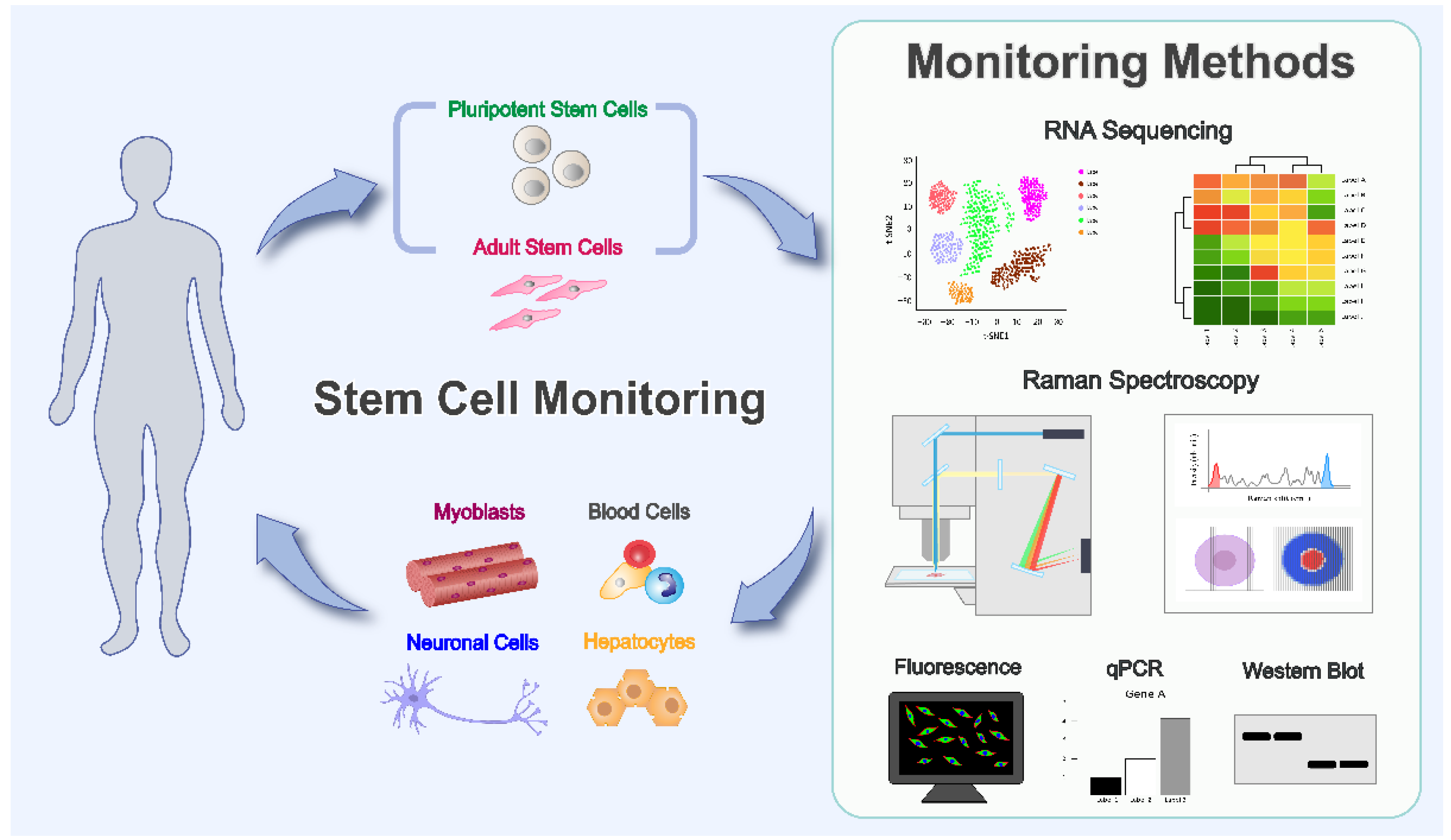
1. Introduction
2. Clinical Application of Various Stem Cells and Necessity of Monitoring Differentiation Process
2.1. Mesenchymal Stem Cells
2.2. Embryonic Stem Cells and Induced Pluripotent Stem Cells
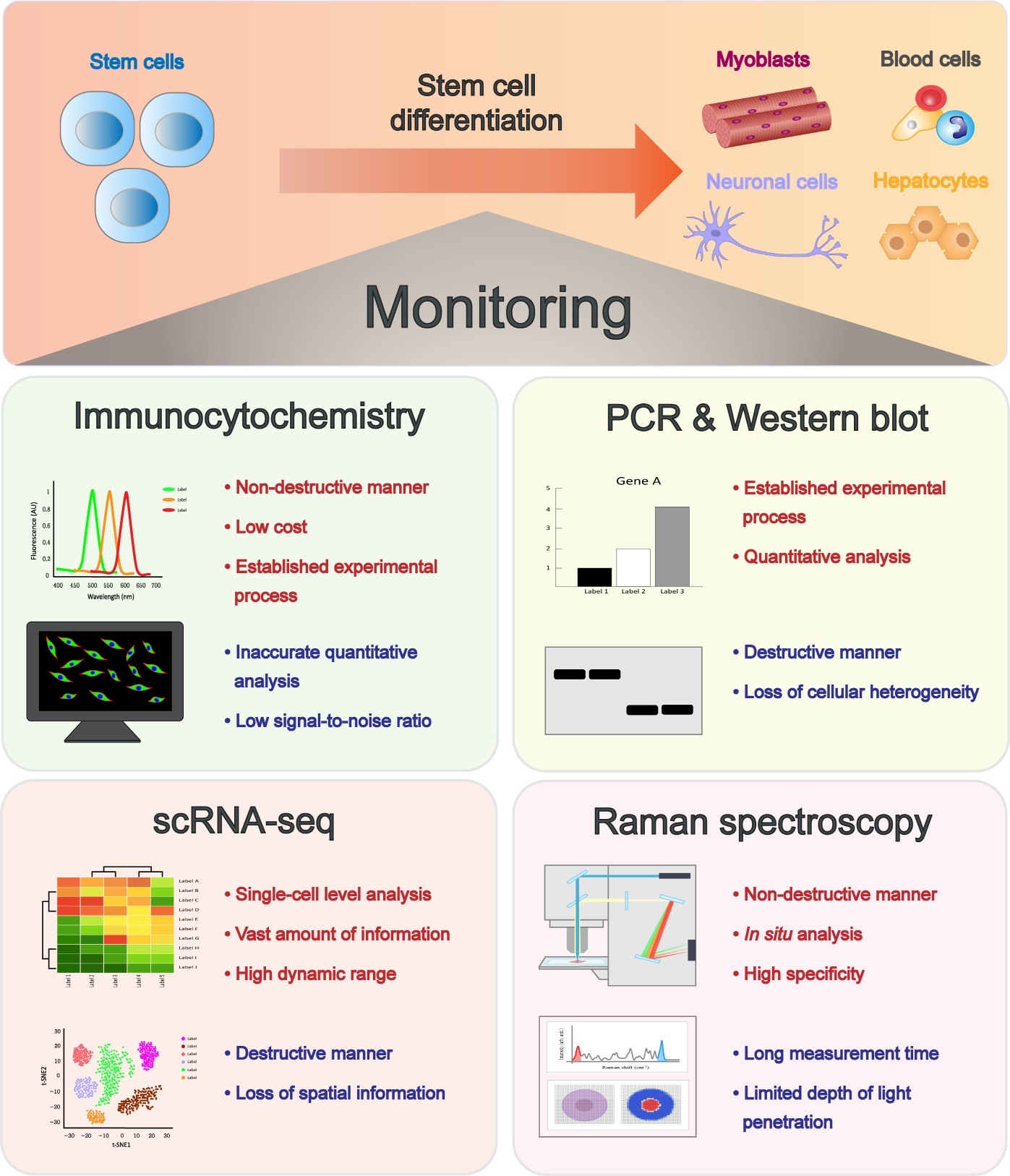
3. Latest Technological Advances in Monitoring Stem Cell Status and Differentiation
3.1. Tracing Differentiation of Stem Cells at the Single-Cell Level Using Single-Cell RNA Sequencing (scRNA-seq)
3.1.1. Cell Fate Mapping by Cellular Barcoding
3.1.2. Challenges of scRNA-seq-Based Differentiation Tracing
3.2. Fluorescence Spectroscopy to Monitor Stem Cell Differentiation
3.2.1. Immunocytochemistry (ICC)
3.2.2. Nanomaterials in Fluorescence-Based Biosensing
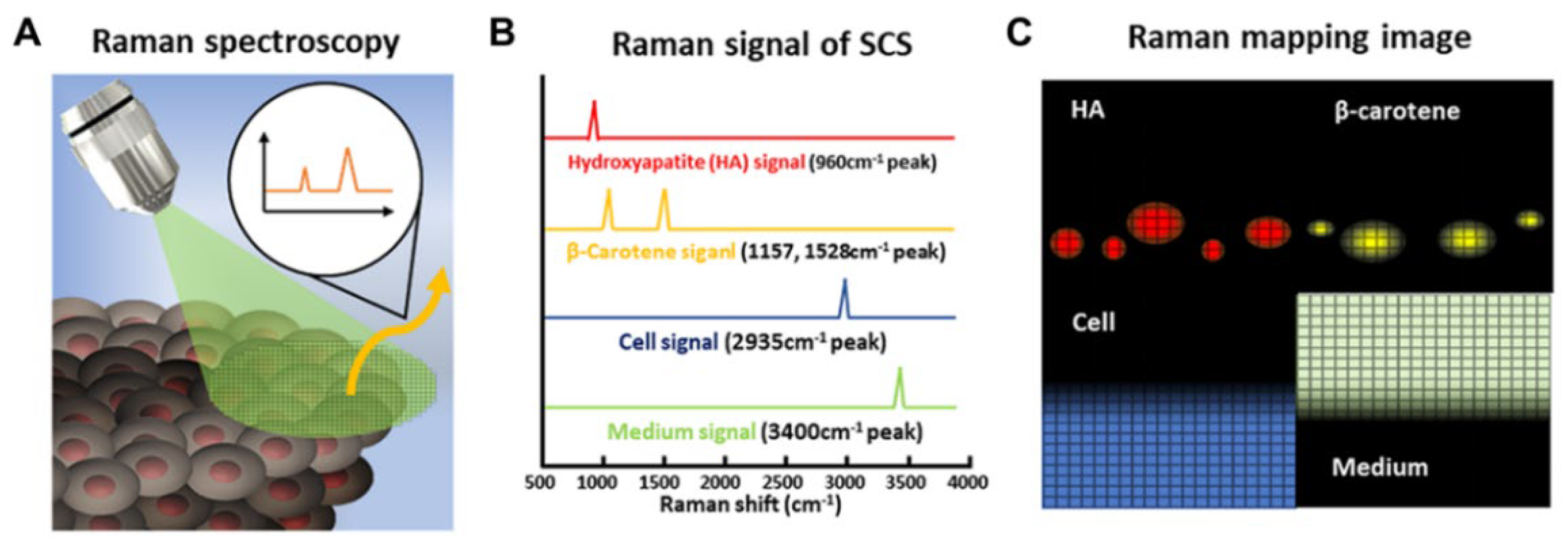
3.3. Profiling and Tracing of Stem Cell Differentiation Using Raman Spectroscopy
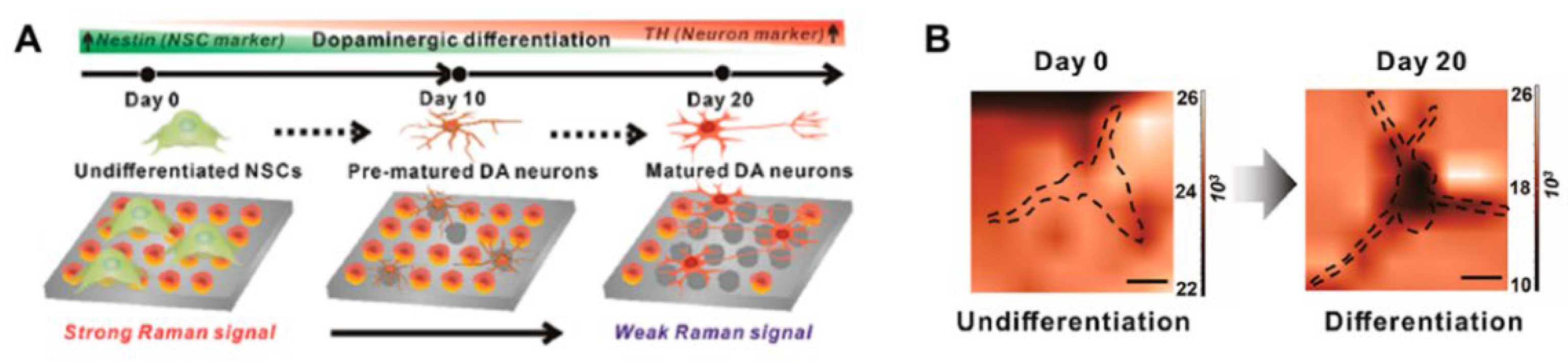
3.3.1. Raman Spectroscopy for Identification of Stem Cell Differentiation
3.3.2. Raman Spectroscopy for Imaging Stem Cell Differentiation
3.3.3. Surface-Enhanced Raman Spectroscopy (SERS)
3.4. Other Methods for Monitoring of Stem Cell Differentiation
3.4.1. Infrared (IR) Spectroscopy
3.4.2. Second Harmonic Generation (SHG) Scanning
3.4.3. Hyperspectral Spectroscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burdick, J.A.; Vunjak-Novakovic, G. Engineered microenvironments for controlled stem cell differentiation. Tissue Eng. Part A 2009, 15, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Mao, A.S.; Shin, J.-W.; Mooney, D.J. Effects of substrate stiffness and cell-cell contact on mesenchymal stem cell differentiation. Biomaterials 2016, 98, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Almalki, S.G.; Agrawal, D.K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, D.M.; Kalisz, M.; Nielsen, J.H. Cytokine signalling in embryonic stem cells. APMIS 2005, 113, 756–772. [Google Scholar] [CrossRef]
- Sato, Y.; Bando, H.; Di Piazza, M.; Gowing, G.; Herberts, C.; Jackman, S.; Leoni, G.; Libertini, S.; MacLachlan, T.; McBlane, J.W.; et al. Tumorigenicity assessment of cell therapy products: The need for global consensus and points to consider. Cytotherapy 2019, 21, 1095–1111. [Google Scholar] [CrossRef]
- Barkholt, L.; Flory, E.; Jekerle, V.; Lucas-Samuel, S.; Ahnert, P.; Bisset, L.; Büscher, D.; Fibbe, W.; Foussat, A.; Kwa, M. Risk of tumorigenicity in mesenchymal stromal cell–based therapies—Bridging scientific observations and regulatory viewpoints. Cytotherapy 2013, 15, 753–759. [Google Scholar] [CrossRef]
- Kircher, M.F.; Gambhir, S.S.; Grimm, J. Noninvasive cell-tracking methods. Nat. Rev. Clin. Oncol. 2011, 8, 677–688. [Google Scholar] [CrossRef]
- Nguyen, P.K.; Riegler, J.; Wu, J.C. Stem cell imaging: From bench to bedside. Cell Stem Cell 2014, 14, 431–444. [Google Scholar] [CrossRef]
- Martí, M.; Mulero, L.; Pardo, C.; Morera, C.; Carrió, M.; Laricchia-Robbio, L.; Esteban, C.R.; Belmonte, J.C.I. Characterization of pluripotent stem cells. Nat. Protoc. 2013, 8, 223–253. [Google Scholar] [CrossRef]
- Nardi, N.B.; Meirelles, L. Mesenchymal stem cells: Isolation, in vitro expansion and characterization. Stem Cells 2008, 249–282. [Google Scholar]
- Ragni, E.; Viganò, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What is beyond aq RT-PCR study on mesenchymal stem cell differentiation properties: How to choose the most reliable housekeeping genes. J. Cell. Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef]
- Suhito, I.R.; Angeline, N.; Choo, S.-S.; Woo, H.Y.; Paik, T.; Lee, T.; Kim, T.-H. Nanobiosensing platforms for real-time and non-invasive monitoring of stem cell pluripotency and differentiation. Sensors 2018, 18, 2755. [Google Scholar] [CrossRef]
- Kim, T.-H.; Yea, C.-H.; Chueng, S.-T.D.; Yin, P.T.-T.; Conley, B.; Dardir, K.; Pak, Y.; Jung, G.Y.; Choi, J.-W.; Lee, K.-B. Large-scale nanoelectrode arrays to monitor the dopaminergic differentiation of human neural stem cells. Adv. Mat. 2015, 27, 6356–6362. [Google Scholar] [CrossRef]
- Friedenstein, A.; Chailakhjan, R.; Lalykina, K. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Gorskaja, J.; Kulagina, N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Lee, O.K.; Kuo, T.K.; Chen, W.-M.; Lee, K.-D.; Hsieh, S.-L.; Chen, T.-H. Isolation of multipotent mesenchymal stem cells from umbilical cord blood. Blood 2004, 103, 1669–1675. [Google Scholar] [CrossRef]
- Bieback, K.; Kern, S.; Klüter, H.; Eichler, H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells 2004, 22, 625–634. [Google Scholar] [CrossRef]
- Zvaifler, N.J.; Marinova-Mutafchieva, L.; Adams, G.; Edwards, C.J.; Moss, J.; Burger, J.A.; Maini, R.N. Mesenchymal precursor cells in the blood of normal individuals. Arthritis Res. Ther. 2000, 2, 477. [Google Scholar] [CrossRef]
- Villaron, E.M.; Almeida, J.; López-Holgado, N.; Alcoceba, M.; Sánchez-Abarca, L.I.; Sanchez-Guijo, F.M.; Alberca, M.; Pérez-Simon, J.A.; San Miguel, J.F.; Del Cañizo, M.C. Mesenchymal stem cells are present in peripheral blood and can engraft after allogeneic hematopoietic stem cell transplantation. Haematologica 2004, 89, 1421–1427. [Google Scholar]
- Young, H.E.; Steele, T.A.; Bray, R.A.; Hudson, J.; Floyd, J.A.; Hawkins, K.; Thomas, K.; Austin, T.; Edwards, C.; Cuzzourt, J. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat. Rec. 2001, 264, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Helm, K.; Ruegg, P.; Varella-Garcia, M.; Burnham, E.; Majka, S. Adult lung side population cells have mesenchymal stem cell potential. Cytotherapy 2008, 10, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Wagner, W.; Wein, F.; Seckinger, A.; Frankhauser, M.; Wirkner, U.; Krause, U.; Blake, J.; Schwager, C.; Eckstein, V.; Ansorge, W. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp. Hematol. 2005, 33, 1402–1416. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Karp, J.M.; Teo, G.S.L. Mesenchymal stem cell homing: The devil is in the details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef]
- Lin, F.; Moran, A.; Igarashi, P. Intrarenal cells, not bone marrow–derived cells, are the major source for regeneration in postischemic kidney. J. Clin. Investig. 2005, 115, 1756–1764. [Google Scholar] [CrossRef]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.-F.; Lian, Q. Paracrine mechanisms of mesenchymal stem cell-based therapy: Current status and perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef]
- Xu, P.; Yang, X. The efficacy and safety of mesenchymal stem cell transplantation for spinal cord injury patients: A meta-analysis and systematic review. Cell Transplant. 2019, 28, 36–46. [Google Scholar] [CrossRef]
- Connick, P.; Kolappan, M.; Crawley, C.; Webber, D.J.; Patani, R.; Michell, A.W.; Du, M.-Q.; Luan, S.-L.; Altmann, D.R.; Thompson, A.J. Autologous mesenchymal stem cells for the treatment of secondary progressive multiple sclerosis: An open-label phase 2a proof-of-concept study. Lancet Neurol. 2012, 11, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Hong, J.M.; Moon, G.J.; Lee, P.H.; Ahn, Y.H.; Bang, O.Y. A long-term follow-up study of intravenous autologous mesenchymal stem cell transplantation in patients with ischemic stroke. Stem Cells 2010, 28, 1099–1106. [Google Scholar] [CrossRef]
- Gupta, P.K.; Das, A.K.; Chullikana, A.; Majumdar, A.S. Mesenchymal stem cells for cartilage repair in osteoarthritis. Stem Cell Res. Ther. 2012, 3, 25. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Cong, X.; Liu, G.; Zhou, J.; Bai, B.; Li, Y.; Bai, W.; Li, M.; Ji, H. Human umbilical cord mesenchymal stem cell therapy for patients with active rheumatoid arthritis: Safety and efficacy. Stem Cells Dev. 2013, 22, 3192–3202. [Google Scholar] [CrossRef]
- Chou, S.-H.; Lin, S.-Z.; Kuo, W.-W.; Pai, P.; Lin, J.-Y.; Lai, C.-H.; Kuo, C.-H.; Lin, K.-H.; Tsai, F.-J.; Huang, C.-Y. Mesenchymal stem cell insights: Prospects in cardiovascular therapy. Cell Transplant. 2014, 23, 513–529. [Google Scholar] [CrossRef]
- Tögel, F.; Westenfelder, C. Adult bone marrow–derived stem cells for organ regeneration and repair. Dev. Dyn. 2007, 236, 3321–3331. [Google Scholar] [CrossRef]
- Ide, C.; Nakai, Y.; Nakano, N.; Seo, T.-B.; Yamada, Y.; Endo, K.; Noda, T.; Saito, F.; Suzuki, Y.; Fukushima, M. Bone marrow stromal cell transplantation for treatment of sub-acute spinal cord injury in the rat. Brain Res. 2010, 1332, 32–47. [Google Scholar] [CrossRef]
- Chimenti, I.; Smith, R.R.; Li, T.-S.; Gerstenblith, G.; Messina, E.; Giacomello, A.; Marbán, E. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 2010, 106, 971–980. [Google Scholar] [CrossRef]
- Park, W.S.; Ahn, S.Y.; Sung, S.I.; Ahn, J.-Y.; Chang, Y.S. Strategies to enhance paracrine potency of transplanted mesenchymal stem cells in intractable neonatal disorders. Pediatr. Res. 2018, 83, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Qu, X.; Zhao, R.C. Clinical applications of mesenchymal stem cells. J. Hematol. Oncol. 2012, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Yuan, Z.; Weng, J.; Pei, D.; Du, X.; He, C.; Lai, P. Challenges and advances in clinical applications of mesenchymal stromal cells. J. Hematol. Oncol. 2021, 14, 24. [Google Scholar] [CrossRef]
- Phinney, D.G.; Kopen, G.; Righter, W.; Webster, S.; Tremain, N.; Prockop, D.J. Donor variation in the growth properties and osteogenic potential of human marrow stromal cells. J. Cell. Biochem. 1999, 75, 424–436. [Google Scholar] [CrossRef]
- McLeod, C.; Mauck, R. On the origin and impact of mesenchymal stem cell heterogeneity: New insights and emerging tools for single cell analysis. Eur. Cells Mater. 2017, 34, 217. [Google Scholar] [CrossRef]
- Ryan, A.E.; Lohan, P.; O’flynn, L.; Treacy, O.; Chen, X.; Coleman, C.; Shaw, G.; Murphy, M.; Barry, F.; Griffin, M.D. Chondrogenic differentiation increases antidonor immune response to allogeneic mesenchymal stem cell transplantation. Mol. Ther. 2014, 22, 655–667. [Google Scholar] [CrossRef]
- Nicolay, N.H.; Perez, R.L.; Debus, J.; Huber, P.E. Mesenchymal stem cells—A new hope for radiotherapy-induced tissue damage? Cancer Lett. 2015, 366, 133–140. [Google Scholar] [CrossRef]
- Costa, L.A.; Eiro, N.; Fraile, M.; Gonzalez, L.O.; Saá, J.; Garcia-Portabella, P.; Vega, B.; Schneider, J.; Vizoso, F.J. Functional heterogeneity of mesenchymal stem cells from natural niches to culture conditions: Implications for further clinical uses. Cell. Mol. Life Sci. 2021, 78, 447–467. [Google Scholar] [CrossRef]
- Herberts, C.A.; Kwa, M.S.; Hermsen, H.P. Risk factors in the development of stem cell therapy. J. Transl. Med. 2011, 9, 29. [Google Scholar] [CrossRef]
- Drela, K.; Stanaszek, L.; Nowakowski, A.; Kuczynska, Z.; Lukomska, B. Experimental strategies of mesenchymal stem cell propagation: Adverse events and potential risk of functional changes. Stem Cells Int. 2019, 2019, 7012692. [Google Scholar] [CrossRef]
- Suhito, I.R.; Han, Y.; Min, J.; Son, H.; Kim, T.-H. In situ label-free monitoring of human adipose-derived mesenchymal stem cell differentiation into multiple lineages. Biomaterials 2018, 154, 223–233. [Google Scholar] [CrossRef]
- Lee, J.; Cha, H.; Park, T.H.; Park, J.H. Enhanced osteogenic differentiation of human mesenchymal stem cells by direct delivery of Cbfβ protein. Biotechnol. Bioeng. 2020, 117, 2897–2910. [Google Scholar] [CrossRef]
- Honarpardaz, A.; Irani, S.; Pezeshki-Modaress, M.; Zandi, M.; Sadeghi, A. Enhanced chondrogenic differentiation of bone marrow mesenchymal stem cells on gelatin/glycosaminoglycan electrospun nanofibers with different amount of glycosaminoglycan. J. Biomed. Mater. Res. A 2019, 107, 38–48. [Google Scholar] [CrossRef]
- Yoon, H.H.; Bhang, S.H.; Kim, T.; Yu, T.; Hyeon, T.; Kim, B.S. Dual roles of graphene oxide in chondrogenic differentiation of adult stem cells: Cell-adhesion substrate and growth factor-delivery carrier. Adv. Funct. Mater. 2014, 24, 6455–6464. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Yamanaka, S. Pluripotent stem cell-based cell therapy—Promise and challenges. Cell Stem Cell 2020, 27, 523–531. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Seki, T.; Fukuda, K. Methods of induced pluripotent stem cells for clinical application. World J. Stem Cells 2015, 7, 116. [Google Scholar] [CrossRef]
- Baker, M. Why hES cells make teratomas. Nat. Rep. Stem Cells 2009. [Google Scholar] [CrossRef]
- Nishizawa, M.; Chonabayashi, K.; Nomura, M.; Tanaka, A.; Nakamura, M.; Inagaki, A.; Nishikawa, M.; Takei, I.; Oishi, A.; Tanabe, K.; et al. Epigenetic variation between human induced pluripotent stem cell lines is an indicator of differentiation capacity. Cell Stem Cell 2016, 19, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R.; Ishikawa, Y.; Katori, R.; Sasamoto, Y.; Taniwaki, Y.; Takayanagi, H.; Tsujikawa, M.; Sekiguchi, K.; Quantock, A.J.; Nishida, K. Coordinated generation of multiple ocular-like cell lineages and fabrication of functional corneal epithelial cell sheets from human iPS cells. Nat. Protoc. 2017, 12, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R.; Ishikawa, Y.; Katayama, T.; Quantock, A.J.; Nishida, K. CD200 facilitates the isolation of corneal epithelial cells derived from human pluripotent stem cells. Sci. Rep. 2018, 8, 16550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef]
- Sun, C.; Wang, L.; Wang, H.; Huang, T.; Yao, W.; Li, J.; Zhang, X. Single-cell RNA-seq highlights heterogeneity in human primary Wharton’s jelly mesenchymal stem/stromal cells cultured in vitro. Stem Cell Res. Ther. 2020, 11, 149. [Google Scholar] [CrossRef]
- Camp, J.G.; Wollny, D.; Treutlein, B. Single-cell genomics to guide human stem cell and tissue engineering. Nat. Methods 2018, 15, 661–667. [Google Scholar] [CrossRef]
- Chu, L.-F.; Leng, N.; Zhang, J.; Hou, Z.; Mamott, D.; Vereide, D.T.; Choi, J.; Kendziorski, C.; Stewart, R.; Thomson, J.A. Single-cell RNA-seq reveals novel regulators of human embryonic stem cell differentiation to definitive endoderm. Genome Biol. 2016, 17, 173. [Google Scholar] [CrossRef]
- Kee, N.; Volakakis, N.; Kirkeby, A.; Dahl, L.; Storvall, H.; Nolbrant, S.; Lahti, L.; Björklund, Å.K.; Gillberg, L.; Joodmardi, E.; et al. Single-cell analysis reveals a close relationship between differentiating dopamine and subthalamic nucleus neuronal lineages. Cell Stem Cell 2017, 20, 29–40. [Google Scholar] [CrossRef]
- Yao, Z.; Mich, J.K.; Ku, S.; Menon, V.; Krostag, A.-R.; Martinez, R.A.; Furchtgott, L.; Mulholland, H.; Bort, S.; Fuqua, M.A.; et al. A single-cell roadmap of lineage bifurcation in human ESC models of embryonic brain development. Cell Stem Cell 2017, 20, 120–134. [Google Scholar] [CrossRef]
- Kebschull, J.M.; Zador, A.M. Cellular barcoding: Lineage tracing, screening and beyond. Nat. Methods 2018, 15, 871–879. [Google Scholar] [CrossRef]
- Kester, L.; van Oudenaarden, A. Single-cell transcriptomics meets lineage tracing. Cell Stem Cell 2018, 23, 166–179. [Google Scholar] [CrossRef]
- Weinreb, C.; Klein, A.M. Lineage reconstruction from clonal correlations. Proc. Natl. Acad. Sci. USA. 2020, 117, 17041–17048. [Google Scholar] [CrossRef]
- Biddy, B.A.; Kong, W.; Kamimoto, K.; Guo, C.; Waye, S.E.; Sun, T.; Morris, S.A. Single-cell mapping of lineage and identity in direct reprogramming. Nature 2018, 564, 219–224. [Google Scholar] [CrossRef]
- Pei, W.; Feyerabend, T.B.; Rössler, J.; Wang, X.; Postrach, D.; Busch, K.; Rode, I.; Klapproth, K.; Dietlein, N.; Quedenau, C.; et al. Polylox barcoding reveals haematopoietic stem cell fates realized in vivo. Nature 2017, 548, 456–460. [Google Scholar] [CrossRef]
- Weinreb, C.; Rodriguez-Fraticelli, A.; Camargo, F.D.; Klein, A.M. Lineage tracing on transcriptional landscapes links state to fate during differentiation. Science 2020, 367, eaaw3381. [Google Scholar] [CrossRef]
- Loulier, K.; Barry, R.; Mahou, P.; Le Franc, Y.; Supatto, W.; Matho, K.S.; Ieng, S.; Fouquet, S.; Dupin, E.; Benosman, R. Multiplex cell and lineage tracking with combinatorial labels. Neuron 2014, 81, 505–520. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Kalhor, R.; Mali, P.; Church, G.M. Rapidly evolving homing CRISPR barcodes. Nat. Methods 2017, 14, 195–200. [Google Scholar] [CrossRef]
- Strzelecka, P.M.; Ranzoni, A.M.; Cvejic, A. Dissecting human disease with single-cell omics: Application in model systems and in the clinic. Dis. Model Mech. 2018, 11, dmm036525. [Google Scholar] [CrossRef]
- Paik, D.T.; Cho, S.; Tian, L.; Chang, H.Y.; Wu, J.C. Single-cell RNA sequencing in cardiovascular development, disease and medicine. Nat. Rev. Cardiol. 2020, 17, 457–473. [Google Scholar] [CrossRef]
- Morris, S.A. The evolving concept of cell identity in the single cell era. Development 2019, 146, dev169748. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Liu, H.; Wei, Z.; Roy, S.B.; Gao, N. An active learning approach for clustering single-cell RNA-seq data. Lab. Investig. 2022, 102, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Biddy, B.A.; Kamimoto, K.; Amrute, J.M.; Butka, E.G.; Morris, S.A. CellTagging: Combinatorial indexing to simultaneously map lineage and identity at single-cell resolution. Nat. Protoc. 2020, 15, 750–772. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.E.; Klein, A.M. Lineage tracing meets single-cell omics: Opportunities and challenges. Nat. Rev. Genet. 2020, 21, 410–427. [Google Scholar] [CrossRef]
- Specht, E.A.; Braselmann, E.; Palmer, A.E. A critical and comparative review of fluorescent tools for live-cell imaging. Annu. Rev. Physiol. 2017, 79, 93–117. [Google Scholar] [CrossRef]
- Tsien, R.Y.; Ernst, L.; Waggoner, A. Fluorophores for confocal microscopy: Photophysics and photochemistry. In Handbook of Biological Confocal Microscopy; Pawley, J.B., Ed.; Springer: Boston, MA, USA, 2006; pp. 338–352. [Google Scholar]
- Meng, F.; Wang, J.; Ping, Q.; Yeo, Y. Quantitative assessment of nanoparticle biodistribution by fluorescence imaging, revisited. ACS Nano 2018, 12, 6458–6468. [Google Scholar] [CrossRef]
- Cox, G. Fundamentals of Fluorescence Imaging; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Rao, J.; Dragulescu-Andrasi, A.; Yao, H. Fluorescence imaging in vivo: Recent advances. Curr. Opin. Biotechnol. 2007, 18, 17–25. [Google Scholar] [CrossRef]
- Thorp-Greenwood, F.L. An introduction to organometallic complexes in fluorescence cell imaging: Current applications and future prospects. Organometallics 2012, 31, 5686–5692. [Google Scholar] [CrossRef]
- Flanagan, L.A.; Ziaeian, B.; Palmer, T.; Schwartz, P.H. CHAPTER 9-Immunocytochemical analysis of stem cells. In Human Stem Cell Manual; Loring, J.F., Wesselschmidt, R.L., Schwartz, P.H., Eds.; Academic Press: Oxford, UK, 2007; pp. 108–126. [Google Scholar]
- Van Hoof, D.; Passier, R.; Ward-Van Oostwaard, D.; Pinkse, M.W.; Heck, A.J.; Mummery, C.L.; Krijgsveld, J. A quest for human and mouse embryonic stem cell-specific proteins* S. Mol. Cell. Proteom. 2006, 5, 1261–1273. [Google Scholar] [CrossRef]
- Fong, C.Y.; Peh, G.S.; Gauthaman, K.; Bongso, A. Separation of SSEA-4 and TRA-1–60 labelled undifferentiated human embryonic stem cells from a heterogeneous cell population using magnetic-activated cell sorting (MACS) and fluorescence-activated cell sorting (FACS). Stem Cell Rev. Rep. 2009, 5, 72–80. [Google Scholar] [CrossRef]
- Brivanlou, A.H.; Gage, F.H.; Jaenisch, R.; Jessell, T.; Melton, D.; Rossant, J. Setting standards for human embryonic stem cells. Science 2003, 300, 913–916. [Google Scholar] [CrossRef]
- Abe, R.; Yamauchi, K.; Kuniyoshi, K.; Suzuki, T.; Matsuura, Y.; Ohtori, S.; Takahashi, K. Neural crest stem cells can be induced in vitro from human-induced pluripotent stem cells using a novel protocol free of feeder cells. J. Rural Med. 2021, 16, 143–147. [Google Scholar] [CrossRef]
- Niwa, H.; Miyazaki, J.-I.; Smith, A.G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nature Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]
- Yuan, H.; Corbi, N.; Basilico, C.; Dailey, L. Developmental-specific activity of the FGF-4 enhancer requires the synergistic action of Sox2 and Oct-3. Genes Dev. 1995, 9, 2635–2645. [Google Scholar] [CrossRef]
- Bredenkamp, N.; Stirparo, G.G.; Nichols, J.; Smith, A.; Guo, G. The cell-surface marker sushi containing domain 2 facilitates establishment of human naive pluripotent stem cells. Stem Cell Rep. 2019, 12, 1212–1222. [Google Scholar] [CrossRef]
- Yuan, H.; Hu, H.; Chen, R.; Mu, W.; Wang, L.; Li, Y.; Chen, Y.; Ding, X.; Xi, Y.; Mao, S. Premigratory neural crest stem cells generate enteric neurons populating the mouse colon and regulating peristalsis in tissue-engineered intestine. Stem Cells Trans. Med. 2021, 10, 922–938. [Google Scholar] [CrossRef]
- Tian, Y.I.; Zhang, X.; Torrejon, K.; Danias, J.; Gindina, S.; Nayyar, A.; Du, Y.; Xie, Y. A bioengineering approach to Schlemm’s canal-like stem cell differentiation for in vitro glaucoma drug screening. Acta Biomater. 2020, 105, 203–213. [Google Scholar] [CrossRef]
- Darvishi, M.; Hamidabadi, H.G.; Bojnordi, M.N.; Zahiri, M.; Niapour, A.; Alizadeh, R. Differentiation of human dental pulp stem cells into functional motor neuron: In vitro and ex vivo study. Tissue Cell 2021, 72, 101542. [Google Scholar] [CrossRef]
- Yang, X.; Han, G.; Pang, X.; Fan, M. Chitosan/collagen scaffold containing bone morphogenetic protein-7 DNA supports dental pulp stem cell differentiation in vitro and in vivo. J. Biomed. Mater. Res. A 2020, 108, 2519–2526. [Google Scholar] [CrossRef]
- Okajcekova, T.; Strnadel, J.; Pokusa, M.; Zahumenska, R.; Janickova, M.; Halasova, E.; Skovierova, H. A comparative in vitro analysis of the osteogenic potential of human dental pulp stem cells using various differentiation conditions. Int. J. Mol. Sci. 2020, 21, 2280. [Google Scholar] [CrossRef]
- Perczel-Kovách, K.; Hegedűs, O.; Földes, A.; Sangngoen, T.; Kálló, K.; Steward, M.C.; Varga, G.; Nagy, K.S. STRO-1 positive cell expansion during osteogenic differentiation: A comparative study of three mesenchymal stem cell types of dental origin. Arch. Oral Biol. 2021, 122, 104995. [Google Scholar] [CrossRef]
- Mennen, R.H.G.; Pennings, J.L.A.J.; Piersma, A.H.A. Neural crest related gene transcript regulation by valproic acid analogues in the cardiac embryonic stem cell test. Reprod. Toxicol. 2019, 90, 44–52. [Google Scholar] [CrossRef]
- Zhang, S.; Hang, Y.; Wu, J.; Tang, Z.; Li, X.; Zhang, S.; Wang, L.; Brash, J.L.; Chen, H. Dual pathway for promotion of stem cell neural differentiation mediated by gold nanocomposites. ACS Appl. Mater. Interfaces 2020, 12, 22066–22073. [Google Scholar] [CrossRef]
- Ku, T.; Hao, F.; Yang, X.; Rao, Z.; Liu, Q.S.; Sang, N.; Faiola, F.; Zhou, Q.; Jiang, G. Graphene quantum dots disrupt embryonic stem cell differentiation by interfering with the methylation level of Sox2. Environ. Sci. Technol. 2021, 55, 3144–3155. [Google Scholar] [CrossRef]
- Regent, F.; Morizur, L.; Lesueur, L.; Habeler, W.; Plancheron, A.; Ben M’Barek, K.; Monville, C. Automation of human pluripotent stem cell differentiation toward retinal pigment epithelial cells for large-scale productions. Sci. Rep. 2019, 9, 10646. [Google Scholar] [CrossRef]
- Duong, A.; Evstratova, A.; Sivitilli, A.; Hernandez, J.J.; Gosio, J.; Wahedi, A.; Sondheimer, N.; Wrana, J.L.; Beaulieu, J.-M.; Attisano, L. Characterization of mitochondrial health from human peripheral blood mononuclear cells to cerebral organoids derived from induced pluripotent stem cells. Sci. Rep. 2021, 11, 4523. [Google Scholar] [CrossRef]
- Chen, C.X.Q.; Abdian, N.; Maussion, G.; Thomas, R.A.; Demirova, I.; Cai, E.; Tabatabaei, M.; Beitel, L.K.; Karamchandani, J.; Fon, E.A.; et al. Standardized quality control workflow to evaluate the reproducibility and differentiation potential of human iPSCs into neurons. bioRxiv 2021. [Google Scholar] [CrossRef]
- Urrutia-Cabrera, D.; Hsiang-Chi Liou, R.; Lin, J.; Shi, Y.; Liu, K.; Hung, S.S.C.; Hewitt, A.W.; Wang, P.-Y.; Ching-Bong Wong, R. Combinatorial approach of binary colloidal crystals and CRISPR activation to improve induced pluripotent stem cell differentiation into neurons. ACS Appl. Mater. Interfaces 2022, 14, 8669–8679. [Google Scholar] [CrossRef]
- Kobolak, J.; Teglasi, A.; Bellak, T.; Janstova, Z.; Molnar, K.; Zana, M.; Bock, I.; Laszlo, L.; Dinnyes, A. Human induced pluripotent stem cell-derived 3D-neurospheres are suitable for neurotoxicity screening. Cells 2020, 9, 1122. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, H.Y.; Kwon, A.; Park, H.; Park, M.-H.; Kim, J.-W.; Kim, M.J.; Kim, Y.-O.; Kwak, S.; Koo, S.K. Generation of a PDX1–EGFP reporter human induced pluripotent stem cell line, KSCBi005-A-3, using the CRISPR/Cas9 system. Stem Cell Res. 2019, 41, 101632. [Google Scholar] [CrossRef]
- Fleischer, S.; Jahnke, H.-G.; Fritsche, E.; Girard, M.; Robitzki, A.A. Comprehensive human stem cell differentiation in a 2D and 3D mode to cardiomyocytes for long-term cultivation and multiparametric monitoring on a multimodal microelectrode array setup. Biosens. Bioelectron. 2019, 126, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Suryawan, I.G.R.; Andrianto; Anggaraeni, A.D.; Agita, A.; Nugraha, R.A. The role of human platelet-rich plasma to enhance the differentiation from adipose derived mesenchymal stem cells into cardiomyocyte: An experimental study. bioRxiv 2021. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Kumar, V.B.; Wen, Y.-T.; Huang, C.-Y.; Tsai, R.-K.; Ding, D.-C. Induction of human umbilical mesenchymal stem cell differentiation into retinal pigment epithelial cells using a transwell-based co-culture system. Cell Transplat. 2022, 31, 09636897221085901. [Google Scholar] [CrossRef] [PubMed]
- Ekram, S.; Khalid, S.; Bashir, I.; Salim, A.; Khan, I. Human umbilical cord-derived mesenchymal stem cells and their chondroprogenitor derivatives reduced pain and inflammation signaling and promote regeneration in a rat intervertebral disc degeneration model. Mol. Cell. Biochem. 2021, 476, 3191–3205. [Google Scholar] [CrossRef]
- Wu, S.-H.; Liao, Y.-T.; Hsueh, K.-K.; Huang, H.-K.; Chen, T.-M.; Chiang, E.-R.; Hsu, S.-H.; Tseng, T.-C.; Wang, J.-P. Adipose-derived mesenchymal stem cells from a hypoxic culture improve neuronal differentiation and nerve repair. Front. Cell. Dev. Biol. 2021, 9, 1110. [Google Scholar] [CrossRef]
- Swanson, W.B.; Omi, M.; Zhang, Z.; Nam, H.K.; Jung, Y.; Wang, G.; Ma, P.X.; Hatch, N.E.; Mishina, Y. Macropore design of tissue engineering scaffolds regulates mesenchymal stem cell differentiation fate. Biomaterials 2021, 272, 120769. [Google Scholar] [CrossRef]
- Kadkhodaeian, H.A.; Salati, A.; Ansari, M.; Nooshabadi, V.T. Tracking the transplanted neurosphere in retinal pigment epithelium degeneration model. Basic Clin. Neurosci. 2021, 12, 523. [Google Scholar] [CrossRef]
- Yu, M.; Lei, B.; Gao, C.; Yan, J.; Ma, P.X. Optimizing surface-engineered ultra-small gold nanoparticles for highly efficient miRNA delivery to enhance osteogenic differentiation of bone mesenchymal stromal cells. Nano Res. 2017, 10, 49–63. [Google Scholar] [CrossRef]
- Niu, Y.-T.; Xie, L.; Deng, R.-R.; Zhang, X.-Y. In the presence of TGF-β1, Asperosaponin VI promotes human mesenchymal stem cell differentiation into nucleus pulposus like- cells. BMC Complement. Med. Ther. 2021, 21, 32. [Google Scholar] [CrossRef]
- Dadheech, N.; Srivastava, A.; Vakani, M.; Shrimali, P.; Bhonde, R.; Gupta, S. Direct lineage tracing reveals Activin—A potential for improved pancreatic homing of bone marrow mesenchymal stem cells and efficient ß-cell regeneration in vivo. Stem Cell Res. 2020, 11, 327. [Google Scholar] [CrossRef]
- Ojaghi, M.; Soleimanifar, F.; Kazemi, A.; Ghollasi, M.; Soleimani, M.; Nasoohi, N.; Enderami, S.E. Electrospun poly-l-lactic acid/polyvinyl alcohol nanofibers improved insulin-producing cell differentiation potential of human adipose-derived mesenchymal stem cells. J. Cell. Biochem. 2019, 120, 9917–9926. [Google Scholar] [CrossRef]
- Patil, S.; Singh, N. Silk fibroin-alginate based beads for human mesenchymal stem cell differentiation in 3D. Biomater. Sci. 2019, 7, 4687–4697. [Google Scholar] [CrossRef]
- Sokolowska, P.; Zukowski, K.; Lasocka, I.; Szulc-Dabrowska, L.; Jastrzebska, E. Human mesenchymal stem cell (hMSC) differentiation towards cardiac cells using a new microbioanalytical method. Analyst 2020, 145, 3017–3028. [Google Scholar] [CrossRef]
- Parodi, V.; Jacchetti, E.; Bresci, A.; Talone, B.; Valensise, C.M.; Osellame, R.; Cerullo, G.; Polli, D.; Raimondi, M.T. Characterization of mesenchymal stem cell differentiation within miniaturized 3D scaffolds through advanced microscopy techniques. Int. J. Mol. Sci. 2020, 21, 8498. [Google Scholar] [CrossRef]
- Sharma, K.D.; Alghazali, K.M.; Hamzah, R.N.; Pandanaboina, S.C.; Nima Alsudani, Z.A.; Muhi, M.; Watanabe, F.; Zhou, G.-L.; Biris, A.S.; Xie, J.Y. Gold nanorod substrate for rat fetal neural stem cell differentiation into oligodendrocytes. Nanomaterials 2022, 12, 929. [Google Scholar] [CrossRef]
- Qu, A.; Sun, M.; Kim, J.-Y.; Xu, L.; Hao, C.; Ma, W.; Wu, X.; Liu, X.; Kuang, H.; Kotov, N.A. Stimulation of neural stem cell differentiation by circularly polarized light transduced by chiral nanoassemblies. Nat. Biomed. Eng. 2021, 5, 103–113. [Google Scholar] [CrossRef]
- Menéndez-Méndez, A.; Paniagua-Herranz, L.; Olivos-Oré, L.A.; Gómez-Villafuertes, R.; Pérez-Sen, R.; Delicado, E.G.; Artalejo, A.R.; Ortega, F. Combining low-density cell culture, single-cell tracking, and patch-clamp to monitor the behavior of postnatal murine cerebellar neural stem cells. STAR Protoc. 2021, 2, 100964. [Google Scholar] [CrossRef]
- Zhang, C.; Tu, H.-L.; Jia, G.; Mukhtar, T.; Taylor, V.; Rzhetsky, A.; Tay, S. Ultra-multiplexed analysis of single-cell dynamics reveals logic rules in differentiation. Sci. Adv. 2019, 5, eaav7959. [Google Scholar] [CrossRef]
- Jaberi, R.; Mirsadeghi, S.; Kiani, S. In vitro characterization of subventricular zone isolated neural stem cells, from adult monkey and rat brain. Mol. Biol. Rep. 2021, 48, 1311–1321. [Google Scholar] [CrossRef]
- Jung, S.; Harris, N.; Niyonshuti, I.I.; Jenkins, S.V.; Hayar, A.M.; Watanabe, F.; Jamshidi-Parsian, A.; Chen, J.; Borrelli, M.J.; Griffin, R.J. Photothermal response induced by nanocage-coated artificial extracellular matrix promotes neural stem cell differentiation. Nanomaterials 2021, 11, 1216. [Google Scholar] [CrossRef]
- Marchini, A.; Favoino, C.; Gelain, F. Multi-functionalized self-assembling peptides as reproducible 3D cell culture systems enabling differentiation and survival of various human neural stem cell lines. Front. Neurosci. 2020, 14, 413. [Google Scholar] [CrossRef]
- Lee, I.C.; Wu, H.J.; Liu, H.L. Dual-frequency ultrasound induces neural stem/progenitor cell differentiation and growth factor utilization by enhancing stable cavitation. ACS Chem. Neurosci. 2019, 10, 1452–1461. [Google Scholar] [CrossRef]
- Sui, Y.; Zhang, W.; Tang, T.; Gao, L.; Cao, T.; Zhu, H.; You, Q.; Yu, B.; Yang, T. Insulin-like growth factor-II overexpression accelerates parthenogenetic stem cell differentiation into cardiomyocytes and improves cardiac function after acute myocardial infarction in mice. Stem Cell Res. Ther. 2020, 11, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W. Nanomaterials in fluorescence-based biosensing. Anal. Bioanal. Chem. 2009, 394, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Song, S.H.; Jin, Y.; Park, H.-J.; Yoon, H.; Jeon, S.; Cho, S.-W. Multiphoton luminescent graphene quantum dots for in vivo tracking of human adipose-derived stem cells. Nanoscale 2016, 8, 8512–8519. [Google Scholar] [CrossRef] [PubMed]
- Niaraki, A.; Shirsavar, M.A.; Aykar, S.S.; Taghavimehr, M.; Montazami, R.; Hashemi, N.N. Minute-sensitive real-time monitoring of neural cells through printed graphene microelectrodes. Biosens. Bioelectron. 2022, 210, 114284. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Li, Y.-M.; Seo, J.G.; Jang, T.-S.; Knowles, J.C.; Song, S.H.; Lee, J.-H. Biological potential of polyethylene glycol (PEG)-functionalized graphene quantum dots in in vitro neural stem/progenitor cells. Nanomaterials 2021, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tay, L.M.; Anggara, R.; Chuah, Y.J.; Kang, Y. Long-term tracking mesenchymal stem cell differentiation with photostable fluorescent nanoparticles. ACS Appl. Mater. Interfaces 2016, 8, 11925–11933. [Google Scholar] [CrossRef]
- Wang, L.; Xu, K.; Hou, X.; Han, Y.; Liu, S.; Wiraja, C.; Yang, C.; Yang, J.; Wang, M.; Dong, X. Fluorescent poly (glycerol-co-sebacate) acrylate nanoparticles for stem cell labeling and longitudinal tracking. ACS Appl. Mater. Interfaces 2017, 9, 9528–9538. [Google Scholar] [CrossRef]
- Jang, S.E.; Qiu, L.; Cai, X.; Lee, J.W.L.; Zhang, W.; Tan, E.-K.; Liu, B.; Zeng, L. Aggregation-induced emission (AIE) nanoparticles labeled human embryonic stem cells (hESCs)-derived neurons for transplantation. Biomaterials 2021, 271, 120747. [Google Scholar] [CrossRef]
- Choi, C.K.K.; Li, J.; Wei, K.; Xu, Y.J.; Ho, L.W.C.; Zhu, M.; To, K.K.; Choi, C.H.J.; Bian, L. A gold@ polydopamine core–shell nanoprobe for long-term intracellular detection of microRNAs in differentiating stem cells. J. Am. Chem. Soc. 2015, 137, 7337–7346. [Google Scholar] [CrossRef]
- Yue, S.; Li, Y.; Qiao, Z.; Song, W.; Bi, S. Rolling circle replication for biosensing, bioimaging, and biomedicine. Trends Biotechnol. 2021, 39, 1160–1172. [Google Scholar] [CrossRef]
- Shi, P.; Wang, Y. Synthetic DNA for cell-surface engineering. Angew. Chem. Int. Ed. 2021, 133, 11684–11695. [Google Scholar] [CrossRef]
- Bi, S.; Yue, S.; Zhang, S. Hybridization chain reaction: A versatile molecular tool for biosensing, bioimaging, and biomedicine. Chem. Soc. Rev. 2017, 46, 4281–4298. [Google Scholar] [CrossRef]
- Samia, A.C.; Chen, X.; Burda, C. Semiconductor quantum dots for photodynamic therapy. J. Am. Chem. Soc. 2003, 125, 15736–15737. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Xu, L.; Feng, L.; Ji, Y.; Tao, L.; Li, S.; Wei, Y. Biocompatible polydopamine fluorescent organic nanoparticles: Facile preparation and cell imaging. Nanoscale 2012, 4, 5581–5584. [Google Scholar] [CrossRef]
- Li, X.; Que, L. Fluorescence enhancement enabled by nanomaterials and nanostructured substrates: A brief review. Rev. Nanosci. Nanotechnol. 2014, 3, 161–176. [Google Scholar] [CrossRef]
- Li, J.; Leung, C.W.T.; Wong, D.S.H.; Xu, J.; Li, R.; Zhao, Y.; Yung, C.Y.Y.; Zhao, E.; Tang, B.Z.; Bian, L. Photocontrolled siRNA delivery and biomarker-triggered luminogens of aggregation-induced emission by up-conversion NaYF4:Yb3+Tm3+@SiO2 nanoparticles for inducing and monitoring stem-cell differentiation. ACS Appl. Mater. Interfaces 2019, 11, 22074–22084. [Google Scholar] [CrossRef]
- Sharifi, M.; Attar, F.; Saboury, A.A.; Akhtari, K.; Hooshmand, N.; Hasan, A.; El-Sayed, M.A.; Falahati, M. Plasmonic gold nanoparticles: Optical manipulation, imaging, drug delivery and therapy. J. Control. Release 2019, 311–312, 170–189. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, K.; Wang, X.; Liang, G.; Li, J. Delivering siRNA to control osteogenic differentiation and real-time detection of cell differentiation in human mesenchymal stem cells using multifunctional gold nanoparticles. J. Mat. Chem. B 2020, 8, 3016–3027. [Google Scholar] [CrossRef]
- Palonpon, A.F.; Sodeoka, M.; Fujita, K. Molecular imaging of live cells by Raman microscopy. Curr. Opin. Chem. Biol. 2013, 17, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.W.; Lieu, D.K. Label-free biochemical characterization of stem cells using vibrational spectroscopy. J. Biophotonics 2009, 2, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Ghita, A.; Pascut, F.C.; Sottile, V.; Denning, C.; Notingher, I. Applications of Raman micro-spectroscopy to stem cell technology: Label-free molecular discrimination and monitoring cell differentiation. EPJ Tech. Instrum. 2015, 2, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sil, S.; Mukherjee, R.; Kumbhar, D.; Reghu, D.; Shrungar, D.; Kumar, N.S.; Singh, U.K.; Umapathy, S. Raman spectroscopy and artificial intelligence open up accurate detection of pathogens from DNA-based sub-species level classification. J. Raman Spectrosc. 2021, 52, 2648–2659. [Google Scholar] [CrossRef]
- Camp, C.H., Jr.; Cicerone, M.T. Chemically sensitive bioimaging with coherent Raman scattering. Nat. Photonics 2015, 9, 295–305. [Google Scholar] [CrossRef]
- Smith, R.; Wright, K.L.; Ashton, L. Raman spectroscopy: An evolving technique for live cell studies. Analyst 2016, 141, 3590–3600. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, P.; Daengngam, C.; Peng, S.; Chongcheawchamnan, M. A review of artificial intelligence methods combined with Raman spectroscopy to identify the composition of substances. J. Raman Spectrosc. 2022, 53, 6–19. [Google Scholar] [CrossRef]
- Liu, Z.; Guo, Z.; Zhong, H.; Qin, X.; Wan, M.; Yang, B. Graphene oxide based surface-enhanced Raman scattering probes for cancer cell imaging. Phys. Chem. Chem. Phys. 2013, 15, 2961–2966. [Google Scholar] [CrossRef]
- Klein, K.; Gigler, A.M.; Aschenbrenner, T.; Monetti, R.; Bunk, W.; Jamitzky, F.; Morfill, G.; Stark, R.W.; Schlegel, J. Label-free live-cell imaging with confocal Raman microscopy. Biophys. J. 2012, 102, 360–368. [Google Scholar] [CrossRef]
- Geng, J.; Zhang, W.; Chen, C.; Zhang, H.; Zhou, A.; Huang, Y. Tracking the differentiation status of human neural stem cells through label-free Raman spectroscopy and machine learning-based analysis. Anal. Chem. 2021, 93, 10453–10461. [Google Scholar] [CrossRef]
- Kallepitis, C.; Bergholt, M.S.; Mazo, M.M.; Leonardo, V.; Skaalure, S.C.; Maynard, S.A.; Stevens, M.M. Quantitative volumetric Raman imaging of three dimensional cell cultures. Nat. Commun. 2017, 8, 14843. [Google Scholar] [CrossRef]
- Parrotta, E.; De Angelis, M.T.; Scalise, S.; Candeloro, P.; Santamaria, G.; Paonessa, M.; Coluccio, M.L.; Perozziello, G.; De Vitis, S.; Sgura, A. Two sides of the same coin? Unraveling subtle differences between human embryonic and induced pluripotent stem cells by Raman spectroscopy. Stem Cell Res. Ther. 2017, 8, 271. [Google Scholar] [CrossRef]
- Lazarević, J.J.; Kukolj, T.; Bugarski, D.; Lazarević, N.; Bugarski, B.; Popović, Z.V. Probing primary mesenchymal stem cells differentiation status by micro-Raman spectroscopy. Spectroc. Acta Pt. A-Molec. Biomolec. Spectr. 2019, 213, 384–390. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Xu, J.; Brinkhof, B.; Wang, H.; Cui, Z.; Huang, W.E.; Ye, H. A single-cell Raman-based platform to identify developmental stages of human pluripotent stem cell-derived neurons. Proc. Natl. Acad. Sci. USA. 2020, 117, 18412–18423. [Google Scholar] [CrossRef]
- Ralbovsky, N.M.; Dey, P.; Dey, B.K.; Lednev, I.K. Determining the stages of cellular differentiation using deep ultraviolet resonance Raman spectroscopy. Talanta 2021, 227, 122164. [Google Scholar] [CrossRef]
- Mandair, G.S.; Steenhuis, P.; Ignelzi, M.A., Jr.; Morris, M.D. Bone quality assessment of osteogenic cell cultures by Raman microscopy. J. Raman Spectrosc. 2019, 50, 360–370. [Google Scholar] [CrossRef]
- Alraies, A.; Canetta, E.; Waddington, R.J.; Moseley, R.; Sloan, A.J. Discrimination of dental pulp stem cell regenerative heterogeneity by single-cell Raman spectroscopy. Tissue Eng. Part C-Methods 2019, 25, 489–499. [Google Scholar] [CrossRef]
- Simonović, J.; Toljić, B.; Rašković, B.; Jovanović, V.; Lazarević, M.; Milošević, M.; Nikolić, N.; Panajotović, R.; Milašin, J. Raman microspectroscopy: Toward a better distinction and profiling of different populations of dental stem cells. Croat. Med. J. 2019, 60, 78–86. [Google Scholar] [CrossRef]
- Esmonde-White, F.W.L.; Morris, M.D. Raman imaging and Raman mapping. In Emerging Raman Applications and Techniques in Biomedical and Pharmaceutical Fields; Matousek, P., Morris, M.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 97–110. [Google Scholar]
- Ravera, F.; Efeoglu, E.; Byrne, H.J. Monitoring stem cell differentiation using Raman microspectroscopy: Chondrogenic differentiation, towards cartilage formation. Analyst 2021, 146, 322–337. [Google Scholar] [CrossRef]
- De Bleye, C.; Fontaine, M.; Dumont, E.; Sacré, P.Y.; Hubert, P.; Theys, N.; Ziemons, E. Raman imaging as a new analytical tool for the quality control of the monitoring of osteogenic differentiation in forming 3D bone tissue. J. Pharm. Biomed. Anal. 2020, 186, 113319. [Google Scholar] [CrossRef]
- Suhito, I.R.; Han, Y.; Ryu, Y.-S.; Son, H.; Kim, T.-H. Autofluorescence-Raman mapping integration analysis for ultra-fast label-free monitoring of adipogenic differentiation of stem cells. Biosens. Bioelectron. 2021, 178, 113018. [Google Scholar] [CrossRef]
- Kim, H.; Han, Y.; Suhito, I.R.; Choi, Y.; Kwon, M.; Son, H.; Kim, H.-R.; Kim, T.-H. Raman spectroscopy-based 3D analysis of odontogenic differentiation of human dental pulp stem cell spheroids. Anal. Chem. 2021, 93, 9995–10004. [Google Scholar] [CrossRef]
- Dou, X.; Zhao, Y.; Li, M.; Chen, Q.; Yamaguchi, Y. Raman imaging diagnosis of the early stage differentiation of mouse embryonic stem cell (mESC). Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 224, 117438. [Google Scholar] [CrossRef]
- Kukolj, T.; Lazarević, J.; Borojević, A.; Ralević, U.; Vujić, D.; Jauković, A.; Lazarević, N.; Bugarski, D. A single-cell Raman spectroscopy analysis of bone marrow mesenchymal stem/stromal cells to identify inter-individual diversity. Int. J. Mol. Sci. 2022, 23, 4915. [Google Scholar] [CrossRef]
- Zong, C.; Xu, M.; Xu, L.-J.; Wei, T.; Ma, X.; Zheng, X.-S.; Hu, R.; Ren, B. Surface-enhanced Raman spectroscopy for bioanalysis: Reliability and challenges. Chem. Rev. 2018, 118, 4946–4980. [Google Scholar] [CrossRef] [PubMed]
- Martines-Arano, H.; Palacios-Barreto, S.; Castillo-Cruz, J.; Meda-Campaña, J.A.; García-Pérez, B.E.; Torres-Torres, C. Fractional photodamage triggered by chaotic attractors in human lung epithelial cancer cells. Int. J. Therm. Sci. 2022, 181, 107734. [Google Scholar] [CrossRef]
- Langer, J.; Jimenez de Aberasturi, D.; Aizpurua, J.; Alvarez-Puebla, R.A.; Auguié, B.; Baumberg, J.J.; Bazan, G.C.; Bell, S.E.J.; Boisen, A.; Brolo, A.G.; et al. Present and future of surface-enhanced Raman scattering. ACS Nano 2020, 14, 28–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gao, X.; Xu, C.; Liu, D. SERS tags for biomedical detection and bioimaging. Theranostics 2022, 12, 1870. [Google Scholar] [CrossRef] [PubMed]
- Mosier-Boss, P.A. Review of SERS substrates for chemical sensing. Nanomaterials 2017, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Jin, S.; Guo, S.; Park, Y.; Chen, L.; Zhao, B.; Jung, Y.M. Recent development of SERS technology: Semiconductor-based study. ACS Omega 2019, 4, 20101–20108. [Google Scholar] [CrossRef]
- Song, C.; Guo, S.; Jin, S.; Chen, L.; Jung, Y.M. Biomarkers determination based on surface-enhanced Raman scattering. Chemosensors 2020, 8, 118. [Google Scholar] [CrossRef]
- El-Said, W.A.; Kim, S.U.; Choi, J.-W. Monitoring in vitro neural stem cell differentiation based on surface-enhanced Raman spectroscopy using a gold nanostar array. J. Mater. Chem. C 2015, 3, 3848–3859. [Google Scholar] [CrossRef]
- Shi, C.; Cao, X.; Chen, X.; Sun, Z.; Xiang, Z.; Zhao, H.; Qian, W.; Han, X. Intracellular surface-enhanced Raman scattering probes based on TAT peptide-conjugated Au nanostars for distinguishing the differentiation of lung resident mesenchymal stem cells. Biomaterials 2015, 58, 10–25. [Google Scholar] [CrossRef]
- Alattar, N.; Daud, H.; Al-Majmaie, R.; Zeulla, D.; Al-Rubeai, M.; Rice, J.H. Surface-enhanced Raman scattering for rapid hematopoietic stem cell differentiation analysis. Appl. Opt. 2018, 57, E184–E189. [Google Scholar] [CrossRef]
- Milewska, A.; Zivanovic, V.; Merk, V.; Arnalds, U.B.; Sigurjónsson, Ó.E.; Kneipp, J.; Leosson, K. Gold nanoisland substrates for SERS characterization of cultured cells. Biomed. Opt. Express 2019, 10, 6172–6188. [Google Scholar] [CrossRef]
- D’Acunto, M. In situ surface-enhanced Raman spectroscopy of cellular components: Theory and experimental results. Materials 2019, 12, 1564. [Google Scholar] [CrossRef]
- Wang, J.; Qi, G.; Qu, X.; Ling, X.; Zhang, Z.; Jin, Y. Molecular profiling of dental pulp stem cells during cell differentiation by surface enhanced Raman spectroscopy. Anal. Chem. 2020, 92, 3735–3741. [Google Scholar] [CrossRef]
- Milewska, A.; Sigurjonsson, O.E.; Leosson, K. SERS imaging of mesenchymal stromal cell differentiation. ACS Appl. Bio Mater. 2021, 4, 4999–5007. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Yang, H.; Xie, J.; Liu, A. Diagnosis and staging of diffuse large B-cell lymphoma using label-free surface-enhanced Raman spectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2022, 267, 120571. [Google Scholar] [CrossRef]
- Choi, J.-H.; Kim, T.-H.; El-said, W.A.; Lee, J.-H.; Yang, L.; Conley, B.; Choi, J.-W.; Lee, K.-B. In situ detection of neurotransmitters from stem cell-derived neural interface at the single-cell level via graphene-hybrid SERS nanobiosensing. Nano Lett. 2020, 20, 7670–7679. [Google Scholar] [CrossRef]
- Omrani, M.; Mohammadi, H.; Fallah, H. Ultrahigh sensitive refractive index nanosensors based on nanoshells, nanocages and nanoframes: Effects of plasmon hybridization and restoring force. Sci. Rep. 2021, 11, 2065. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Z.; Bi, L.; Bi, C.; Du, Q. Gold nanocage-based surface-enhanced Raman scattering probes for long-term monitoring of intracellular microRNA during bone marrow stem cell differentiation. Nanoscale 2020, 12, 1513–1527. [Google Scholar] [CrossRef]
- Hua, S.; Zhong, S.; Arami, H.; He, J.; Zhong, D.; Zhang, D.; Chen, X.; Qian, J.; Hu, X.; Zhou, M. Simultaneous deep tracking of stem cells by surface enhanced Raman imaging combined with single-cell tracking by NIR-II imaging in myocardial infarction. Adv. Funct. Mater. 2021, 31, 2100468. [Google Scholar] [CrossRef]
- Riley, R.S.; Day, E.S. Gold nanoparticle-mediated photothermal therapy: Applications and opportunities for multimodal cancer treatment. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1449. [Google Scholar] [CrossRef]
- Krafft, C.; Salzer, R.; Seitz, S.; Ern, C.; Schieker, M. Differentiation of individual human mesenchymalstem cells probed by FTIR microscopic imaging. Analyst 2007, 132, 647–653. [Google Scholar] [CrossRef]
- Downes, A.; Mouras, R.; Elfick, A. Optical spectroscopy for noninvasive monitoring of stem cell differentiation. J. Microbiol. Biotechnol. 2010, 2010, 101864. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W.; Liu, Z.; Liu, J.; Cheng, J.; Li, Y.; Li, X.; Hu, J.; Lü, J. Single-cell infrared microspectroscopy quantifies dynamic heterogeneity of mesenchymal stem cells during adipogenic differentiation. Anal. Chem. 2021, 93, 671–676. [Google Scholar] [CrossRef]
- Gieroba, B.; Przekora, A.; Kalisz, G.; Kazimierczak, P.; Song, C.L.; Wojcik, M.; Ginalska, G.; Kazarian, S.G.; Sroka-Bartnicka, A. Collagen maturity and mineralization in mesenchymal stem cells cultured on the hydroxyapatite-based bone scaffold analyzed by ATR-FTIR spectroscopic imaging. Mater. Sci. Eng. C 2021, 119, 111634. [Google Scholar] [CrossRef]
- Kato, N. Optical second harmonic generation microscopy: Application to the sensitive detection of cell membrane damage. Biophys. Rev. 2019, 11, 399–408. [Google Scholar] [CrossRef]
- Qi, X.; Liu, H.; Guo, W.; Lin, W.; Lin, B.; Jin, Y.; Deng, X. New opportunities: Second harmonic generation of boron-doped graphene quantum dots for stem cells imaging and ultraprecise tracking in wound healing. Adv. Funct. Mater. 2019, 29, 1902235. [Google Scholar] [CrossRef]
- Ibrahim, A.; Rodriguez-Florez, N.; Gardner, O.F.W.; Zucchelli, E.; New, S.E.P.; Borghi, A.; Dunaway, D.; Bulstrode, N.W.; Ferretti, P. Three-dimensional environment and vascularization induce osteogenic maturation of human adipose-derived stem cells comparable to that of bone-derived progenitors. Stem Cell Res. Ther. 2020, 9, 1651–1666. [Google Scholar] [CrossRef] [PubMed]
- Kourgiantaki, A.; Tzeranis, D.S.; Karali, K.; Georgelou, K.; Bampoula, E.; Psilodimitrakopoulos, S.; Yannas, I.V.; Stratakis, E.; Sidiropoulou, K.; Charalampopoulos, I.; et al. Neural stem cell delivery via porous collagen scaffolds promotes neuronal differentiation and locomotion recovery in spinal cord injury. Regen. Med. 2020, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Bertani, F.R.; Ferrari, L.; Mussi, V.; Botti, E.; Costanzo, A.; Selci, S. Living matter observations with a novel hyperspectral supercontinuum confocal microscope for VIS to near-IR reflectance spectroscopy. Sensors 2013, 13, 14523–14542. [Google Scholar] [CrossRef] [PubMed]
- Ogi, H.; Moriwaki, S.; Kokubo, M.; Hikida, Y.; Itoh, K. Label-free classification of neurons and glia in neural stem cell cultures using a hyperspectral imaging microscopy combined with machine learning. Sci. Rep. 2019, 9, 633. [Google Scholar] [CrossRef]
- Mehta, N.; Shaik, S.; Prasad, A.; Chaichi, A.; Sahu, S.P.; Liu, Q.; Hasan, S.M.A.; Sheikh, E.; Donnarumma, F.; Murray, K.K.; et al. Multimodal label-free monitoring of adipogenic stem cell differentiation using endogenous optical biomarkers. Adv. Funct. Mater. 2021, 31, 2103955. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stem Cell | Cell Source | Target Lineage |
| Adipose-derived stem cell (ADSC) | Human ADSC | Human Schlemm’s canal cell [100] |
| Dental pulp stem cell (DPSC) | Human DPSC | Motor neuron cell [101] |
| Human DPSC | Osteogenic cell [102,103] | |
| DPSC, dental follicle stem cells, periodontal ligament stem cell | Osteogenesis [104] | |
| Embryonic stem cell (ESC) | Mouse ESC | Neural crest cell [105] |
| Mouse ESC | Neuron [106] | |
| Mouse ESC | Embryoid body [107] | |
| Human ESC | Retinal pigment epithelial [108] | |
| Human ESC | Somatic cell [98] | |
| Induced pluripotent stem cell (iPSC) | Human iPSC | Neural crest stem cell [95] |
| Human peripheral blood mononuclear cell | iPSC [109] | |
| Human iPSC | Neuron [110,111] | |
| Human iPSC | Neuron [112] | |
| Human iPSC | β-cell [113] | |
| Human iPSC | Cardiomyocyte [114] | |
| Mesenchymal stem cell (MSC) | Human adipose-derived (AD)-MSC | Cardiomyocyte [115] |
| Human umbilical cord (UC)-MSC | Retinal pigment epithelial [116] | |
| Human UC-MSC | Chondroprogenitor [117] | |
| Human bone marrow (BM)-MSC | Neuron [118] | |
| Mouse MSC | Bone [119] | |
| Rat BM-MSC | Neurosphere [120] | |
| Rat BM-MSC | Bone [121] | |
| Human MSC | Nucleus pulposus-like cell [122] | |
| Mouse BM-MSC | β-cell into pancreatic lineage [123] | |
| Human AD-MSC | Pancreatic cell [124] | |
| Human MSC | Osteogenic and chondrogenic lineage [125] | |
| Human MSC | Cardiac cell [126] | |
| Rat BM-MSC | Adipogenic and chondrogenic cell [127] | |
| Neural stem cell (NSC) | Rat NSC | oligodendrocyte [128] |
| NSC | Neuron [112,129,130,131] | |
| Premigratory neural crest stem cell | Enteric neuron [99] | |
| Monkey NSC | Neuronal cell, glial cell [132] | |
| Rat NSC | Neuron [133] | |
| Human NSC | Neuron [134] | |
| NSC/progenitor cell | Neuron [135] | |
| Parthenogenetic stem cell | Mouse parthenogenetic stem cell | Cardiomyocyte [136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Park, E.; Yoo, H.S.; Park, J.; Jung, Y.M.; Park, J.H. Recent Advances in Monitoring Stem Cell Status and Differentiation Using Nano-Biosensing Technologies. Nanomaterials 2022, 12, 2934. https://doi.org/10.3390/nano12172934
Kim W, Park E, Yoo HS, Park J, Jung YM, Park JH. Recent Advances in Monitoring Stem Cell Status and Differentiation Using Nano-Biosensing Technologies. Nanomaterials. 2022; 12(17):2934. https://doi.org/10.3390/nano12172934
Chicago/Turabian StyleKim, Wijin, Eungyeong Park, Hyuk Sang Yoo, Jongmin Park, Young Mee Jung, and Ju Hyun Park. 2022. "Recent Advances in Monitoring Stem Cell Status and Differentiation Using Nano-Biosensing Technologies" Nanomaterials 12, no. 17: 2934. https://doi.org/10.3390/nano12172934
APA StyleKim, W., Park, E., Yoo, H. S., Park, J., Jung, Y. M., & Park, J. H. (2022). Recent Advances in Monitoring Stem Cell Status and Differentiation Using Nano-Biosensing Technologies. Nanomaterials, 12(17), 2934. https://doi.org/10.3390/nano12172934