
Engineered and Mimicked Extracellular Nanovesicles for Therapeutic Delivery

Abstract
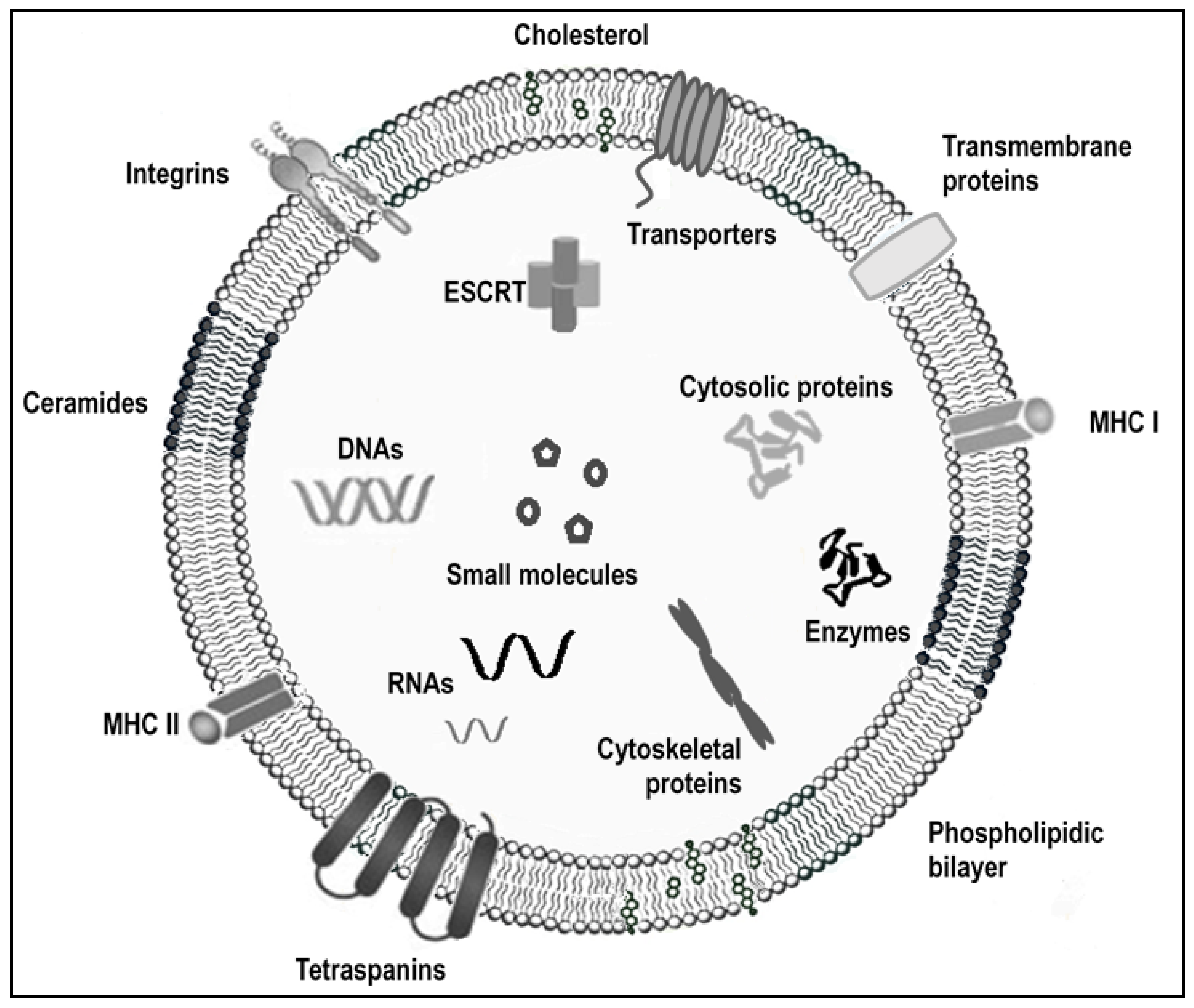
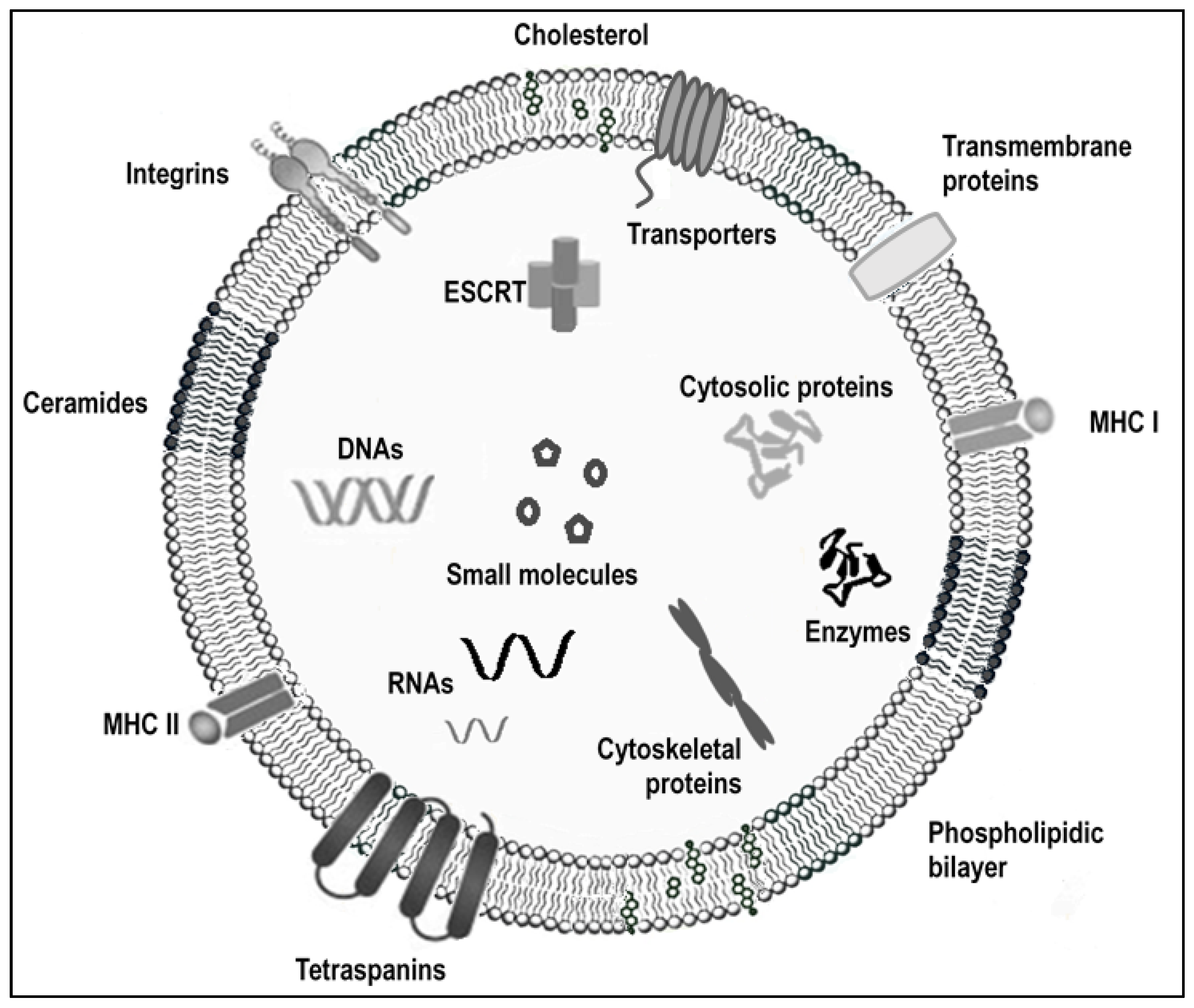
:1. Introduction
2. Clinical Trials Based on Exosomes
3. Artificially Loaded Exosomes
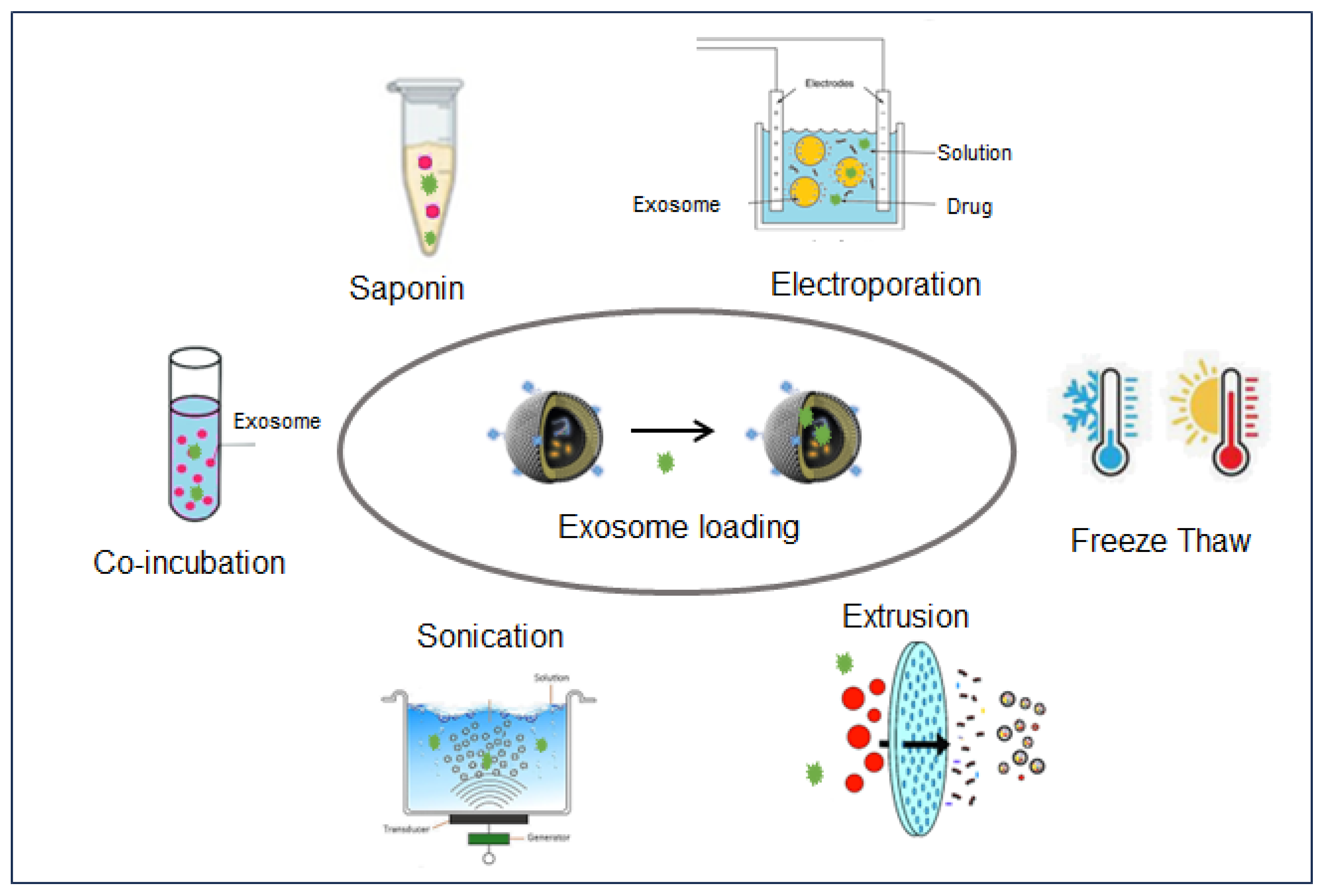
3.1. Loading Methods
3.2. Loaded Exosomes Preclinical Applications
3.2.1. Loading for Tracking and Labelling Purposes
3.2.2. RNA-Loaded EVs
3.2.3. Protein-Loaded EVs
3.2.4. Drug-Loaded EVs
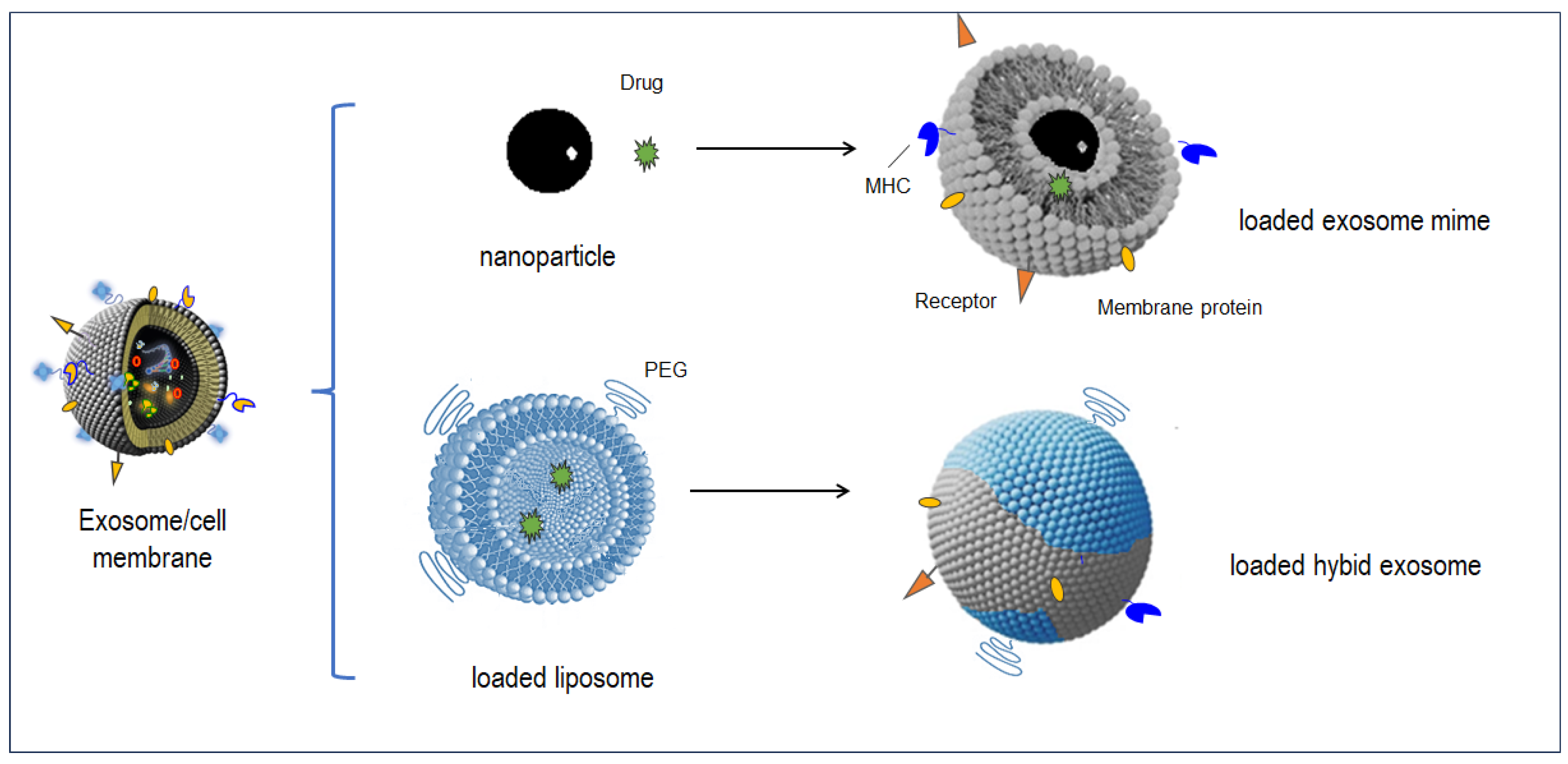
4. Artificial Mimes for Exosomes
4.1. Cell-Membrane Covered Napoparticles
4.1.1. Single-Membrane Covering

{kind=link}
{kind=link}
{kind=link}
| Loaded Molecules | Exosome Cell Line | Loading Methods | Max Loading Efficiency/Capacity | Targeted Pathology/Fonction | Ref. |
|---|---|---|---|---|---|
| siRNA MAPK1 | HeLa, HTB 177, human blood cells | Electroporation: 150 V-100 µF | NS | Cancer therapy | [84] |
| siRNA luciferase | Primary endothelial cell mice | Electroporation: 400 V-200 µF | 25% | Drug absorption | [85] |
| siRNA RAD51 and RAD52 | HeLa, ascites | Electroporation: 700 V-350 µF | NS | Drug absorption | [86] |
| siRNA FGL siRNA TGF-β1 | RAW 264.7 | Transfection with commercial reagent | 97% | Cancer immunotherapy | [87] |
| siRNA KRASG12D | Human MSC | Electroporation: 400 V-150 µF | NS | Pancreatic cancer | [88] |
| siRNA CTGF | Rat MSC | Electroporation: 400 V-200 to 500 µF | 33% | Cancer therapy | [89] |
| siRNA CCL2 | Induced neural SC, surface modified with CAQK | Electroporation: 400 mV-125 μF (pulse time 10–15 ms) | 17% | Spinal cord injury | [90] |
| siRNA DNSL1; SND1; ENDD1; RINI; 5NTD; ENPP1; CN37 | Umbilical cord Wharton’s jelly mesenchymal stem cells | Co-incubation, 37 °C, 1 h | 18% | Huntington’s disease | [93] |
| miRNA 484 | HEK293T | Electroporation: 700 V-150 µF | NS | Metastatic ovarian cancer | [94] |
| miRNA 21-5p (conjugated to CPP) | Mouse embryonic fibroblasts (MF) Human primar dermal fibroblasts (HDFa) | Co-incubation, 37 °C, 8 h | 1600 copies/exo | Loading optimization | [95] |
| miRNA 159 doxorubicine | Human monocytes THP-1 | Co-incubation, 37 °C, 1 h 30 | 16% | Breast cancer | [96] |
| miRNA 21 inhibitor 5-FU (5-fluorouracile) | Transfected HEK293T cells | Electroporation: 1000 V-15 µF, 10 ms | 3% | Colorectal caecinima | [97] |
| miRNA 21 antisens | Transfected HEK293T cells | Electroporation: 400 mV-125 μF | 1.7% | Glioblastoma | [98] |
| TPP1-encoding pDNA | Macrophages | Sonication; saponin-permeation | NS | Late-infantile neuronal ceroid lipofuscinosis | [99] |
| Dapagliflozin | iPS-derived ECs | Extrusion | 28% | Diabetic wound healing | [100] |
| β-glucuronidase | Human MSCs | Saponin permeation: 0.1 mg/mL, 10 min, RT | NS | EV-based hydrogel-developments | [101] |
| Human insulin | Hepatocellular carcinoma (HepG2) Primary dermal fibroblasts (HDFa) | Co-incubation, RT, 1 h Electroporation: 200 V-50 µF | 3% 50% | Diabetes melitus treatment | [102] |
| Yap1 protein | Rat plasma platelets | Electroporation: 200 V-500 µF, 26 ms | NS | Achilles tendon injury | [103] |
| Human chorionic gonadotrophin | Human uterine fluid | Freeze-thaw cycle and sonication | 40% | Assisted reproductive technology | [104] |
| Doxorubicin | Human breast cancer cells (MDA-MB-231, MCF 7) | Electroporation: 350 V-150 µF | NS | Tumor therapy | [68] |
| Doxorubicin | Huma CRC cells LIM1215 | Incubation: 5 min Dialyse PBS overnight | 9% | Colorectal cancer | [105] |
| Paclitaxel | M1 macrophages RAW 264.7 | Sonication: 20% amplitude; 6 cycles of 30 s | NS | Cancer therapy | [106] |
| Paclitaxel | ESC H9 cells | Incubation: 2H; RT | NS | Glioblatoma | [107] |
| Paclitaxel | PANC-1 (pancreatic ductal carcinoma) cells; U937 | Co-incubation, RT, 1 h and sonication | 37% | Pancreatic cancer | [108] |
| Paclitaxel | Human umbilical cord-derived mesenchymal stem cells | Extrusion | 14% | Spinal cord injury | [109] |
| Gemcitabine | Pancreatic cancer cells | Incubation: 2H; 37 °C Sonication: 20% amplitude; 3 cycles of 30 s | 12% | Pancreatic cancer | [111] |
| siRNA galectin-9 oxaliplatin | BM MSC cells | Electroporation: 400 V-125 µF | 13% | Pancreatic cancer | [112] |
| Erastin rose bengal | CD47-overexpressing HEK293T | Sonication | 60% 84% | Hepatocarcinoma | [113] |
| Vit A + quercetin | Mice MSC | Incubation: 2 h; 22 °C | Vit A: 0.279 µg/mL exo Que: 0.19 µg/mL exo | Acute liver injury | [114] |
| Curcuma resveratrol | Raw bovine milk | Incubation: 1 h, 37 °C darkness Sonication: 1 Hz; 150 W; 4 or 6 cycles of 20 s. Electroporation: 400 V; 2 pulses | Cur 2 µg/0.5 ng prot exo; Res 15 µg/0.8 ng prot exo | Breast cancer | [119] |
| Curcuma | Cow skimmed milk Intestinal cells (Caco-2) | Incubation: overnight; RT Size exclusion chromatography | Cur:9% Caco-2: 3% | Drug absorption | [149] |
4.1.2. Hybrid Membrane Covering
4.1.3. Membrane Covering of Organometallic Cores
4.2. Exosome–Liposome Hybrids
4.2.1. Hybridization Techniques
4.2.2. RNA-Loaded HEs
4.2.3. Protein-Loaded HEs
4.2.4. Drug-Loaded HEs
| Exosome-Donor Cells | Liposomes | Cargo | Fusion Technique | Application | Ref. |
|---|---|---|---|---|---|
| RAW 264.7 macrophages CMS7 sarcoma fibroblast | DOPC or DOPS | Fluorescent DMPE | Freeze–thaw | Setting-up hybridation | [170] |
| HEK293 T embryon cells | Not mentioned | Crispr/Cas9 | Co-incubation | Gene therapy | [172] |
| A549 and 3C3 cancer cells | DOPC/DOPS | Anti siRNA GFP | Membrane extrusion | Setting-up for gene delivery | [173] |
| K-HEP1 cancer cells | DPPC | siRNA CDK1 | Membrane extrusion | Hepatocarcinoma | [174] |
| NIH-3T3 CD24++ cancer cells | DOTAP/cholesterol | Anti miRNA 188 | Membrane extrusion | Osteoporosis | [175] |
| SKOv3 cancer cells CP cardiomyoblasts | DLin-MC3-MA/DPPC/cholesterol/18:1-BiotinylCapPE/DMG-PEG | siRNA AF647or AF488 | Membrane extrusion | Cardiac injury | [176] |
| 4T1 breast cancer cells B16F10 melanoma | DOPC/cholesterol/mPEG2000/DSPE | Doxorubicin | Sonication and extrusion | Cancer immunotherapy | [177] |
| J774A.1 macrophages | Egg-PC and cholesterol | Doxorubicin | Membrane extrusion | [179] | |
| Rat blood-serum | DSPE-PEG2000-tLyp-1 | SAB in exosome tLP1 and CTP | Membrane extrusion | Glioma | [180] |
| L-929 fibroblast raw 264.7 macrophages | Cholesterol/DOPC/DSPE-PEG2000 | CLD and Nin | Membrane extrusion | Pulmonary fibrosis | [181] |
| L-929 fibroblast | Cholesterol/DOPC/DSPE-PEG2000 | CLD and Nin | Membrane extrusion | Liver fibrosis | [182] |
| SCOV3-CDDP CD24++ cancer cells | DSPE-PEG1000-COOH | cRGT, tripotolide and miRNA-497 | Membrane extrusion | Ovarian cancer | [183] |
| Mouse blood platelets | Thermosensitive liposome: DPPC/DPPG/cypate | Glucoxidase and ferric ammonium | PEG mediation | Hepatocarcinoma | [184] |
| 3T3 CD24++ cancer cells | Thermosensitive liposome: CFL/DPPC/MSPC/DSPE-PEG2000 | GM-CSF and docetaxel | Freeze–thaw | Metastatic peritoneal carcinoma | [50] |
5. Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Johnstone, R.M.; Adam, M.; Hammond, J.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Aroc, C.J.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell. Dev. Bio. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Ristorcelli, E.; Beraud, E.; Mathieu, S.; Lombardo, D.; Verine, A. Essential role of Notch signaling in apoptosis of human pancreatic tumoral cells mediated by exosomal nanoparticles. Int. J. Cancer 2009, 125, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Desai, N.N.; Qureshi, M.Z.; Librelotto, D.R.N.; Gasparri, M.L.; Bishayee, A.; Nabavi, S.M.; Curti, V.; Daglia, M. Exosome biogenesis, bioactivities and functions as new delivery systems of natural compounds. Biotechnol. Adv. 2018, 36, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Maeki, M.; Kimura, N.; Sato, Y.; Harashima, H.; Tokeshi, M. Advances in microfluidics for lipid nanoparticles and extracellular vesicles and applications in drug delivery systems. Adv. Drug Deliv. Rev. 2018, 128, 84–100. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, C.; Melo, S.A.; Protopopov, A.; Tang, J.; Seth, S.; Koch, M.; Zhang, J.; Weitz, J.; Chin, L.; Futreal, A.; et al. Identification of double-stranded genomic DNA spanning all chromosomes with mutated KRAS and p53 DNA in the serum exosomes of patients with pancreatic cancer. J. Biol. Chem. 2014, 289, 3869–3875. [Google Scholar] [CrossRef]
- de Jong, O.G.; Verhaar, M.C.; Chen, Y.; Vader, P.; Gremmels, H.; Posthuma, G.; Schiffelers, R.M.; Gucek, M.; van Balkom, B.W. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 2012, 1, 18396. [Google Scholar] [CrossRef]
- Willms, E.; Cabanas, C.; Mager, I.; Wood, M.J.A.; Vader, P. Extracellular vesicle heterogeneity: Subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Greening, D.W.; Simpson, R.J. Understanding extracellular vesicle diversity—Current status. Expert Rev. Proteom. 2018, 15, 887–910. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Brennan, M.; Lötvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, 8521. [Google Scholar] [CrossRef] [PubMed]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef]
- Zhou, H.; Pisitkun, T.; Aponte, A.; Yuen, P.S.; Hoffert, J.D.; Yasuda, H.; Hu, X.; Chawla, L.; Shen, R.F.; Knepper, M.A.; et al. Exosomal Fetuin-A identified by proteomics: A novel urinary biomarker for detecting acute kidney injury. Kidney Int. 2006, 70, 1847–1857. [Google Scholar] [CrossRef]
- Miranda, K.C.; Bond, D.T.; McKee, M.; Skog, J.; Paunescu, T.G.; Da Silva, N.; Brown, D.; Russo, L.M. Nucleic acids within urinary exosomes/microvesicles are potential biomarkers for renal disease. Kidney Int. 2010, 78, 191–199. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D. Minimal information for studies of extracellular vesicles, (MISEV 2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV 2014 guidelines. J. Extracell. Vesicles 2017, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.A.; de Jong, O.G.; Schiffelers, R.M. Exploring interactions between extracellular vesicles and cells for innovative drug delivery system design. Adv. Drug Deliv. Rev. 2021, 173, 252–278. [Google Scholar] [CrossRef]
- Record, M.; Silvente-Poirot, S.; Poirot, M.; Wakelam, M.O. Extracellular vesicles: Lipids as key components of their biogenesis and functions. J. Lipid Res. 2018, 59, 1316–1324. [Google Scholar] [CrossRef]
- Llorente, A.; Skotland, T.; Sylvänne, T.; Kauhanen, D.; Rog, T.; Orłowski, A.; Vattulainen, I.; Ekroos, K.; Sandvig, K.; Bo, C. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Lipids 2013, 1831, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Ristorcelli, E.; Beraud, E.; Verrando, P.; Villard, C.; Lafitte, D.; Sbarra, V.; Lombardo, D.; Verine, A. Human tumor nanoparticles induce apoptosis of pancreat cancer cells. FASEB J. 2008, 22, 3358–3369. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; Gonzàlez-King, H.; Grueso, E.; Sànchez, R.; Martinez-Romero, A.; Jàvega, B.; O’connor, J.; Simons, P.; Handberg, A.; Sepúlveda, P. Circulating exosomes deliver free fatty acids from the bloodstream to cardiac cells: Possible role of CD36. PLoS ONE 2019, 14, e0217546. [Google Scholar] [CrossRef] [PubMed]
- Ghadami, S.; Dellinger, K. The lipid composition of extracellular vesicles: Applications in diagnostics and therapeutic delivery. Front. Mol. Biosci. 2023, 10, 1198044. [Google Scholar] [CrossRef] [PubMed]
- Czubowicz, K.; Jésko, H.; Wencel, P.; Lukiw, W.J.; Strosznajder, R.P. The role of ceramide and sphingosine-1-phosphate in Alzheimer’s disease and other neurodegenerative disorders. Mol. Neurobiol. 2019, 56, 5436–5455. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.-I.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J. Biol. Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.I.; Cashikar, A. Multivesicular body morphogenesis. Annu. Rev. Cell Dev. Biol. 2012, 28, 337–362. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, R.S.; Kulkarni, A.; Becker, M.W.; Lei, X.; Sarkar, S.; Ramanadham, S.; Phelps, E.A.; Nakayasu, E.S.; Sims, E.K.; Mirmira, R.G. Extracellular vesicles in β cell biology: Role of lipids in vesicle biogenesis, cargo, and intercellular signaling. Mol. Metab. 2022, 63, 101545. [Google Scholar] [CrossRef]
- Li, S.-P.; Lin, Z.-X.; Jiang, X.-Y.; Yu, X.-Y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharm. Sin. 2018, 39, 542–551. [Google Scholar] [CrossRef]
- Egea-Jimenez, A.L.; Zimmermann, P. Phospholipase D and phosphatidic acid in the biogenesis and cargo loading of extracellular vesicles: Thematic review series: Exosomes and microvesicles: Lipids as key components of their biogenesis and functions. J. Lipid Res. 2018, 59, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Mazzeo, C.; Rodríguez, M.C.; Marsh, M.; Fraile-Ramos, A.; Calvo, V.; Avila-Flores, A.; Merida, I.; Izquierdo, M. Diacylglycerol kinase α regulates the formation and polarisation of mature multivesicular bodies involved in the secretion of Fas ligand-containing exosomes in T lymphocytes. Cell Death Differ. 2011, 18, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Ghossoub, R.; Lembo, F.; Rubio, A.; Gaillard, C.B.; Bouchet, J.; Vitale, N.; Slavík, J.; Machala, M.; Zimmermann, P.J. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat. Commun. 2014, 5, 3477. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Witte, D.P.; Qi, X. Saposin C–LBPA interaction in late-endosomes/lysosomes. Exp. Cell Res. 2005, 303, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.M.; Lasiecka, Z.M.; Xu, Y.; Neufeld, J.; Shahriar, S.; Simoes, S.; Chan, R.B.; Oliveira, T.G.; Small, S.A.; Di Paolo, G. Neuronal lysosomal dysfunction releases exosomes harboring APP C-terminal fragments and unique lipid signatures. Nat. Commun. 2018, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Young, L.N.; Cho, K.; Lawrence, R.; Zoncu, R.; Hurley, J.H. Dynamics and Architecture of NRBF2-Containing phosphatidylinositol 3-kinase Complex I of autophagy. Proc. Natl. Acad. Sci. USA 2016, 113, 8224–8229. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, N.; Yan, T.; Shi, Y.-N.; Chen, J.; Zhang, C.-J.; Xie, X.-J.; Liao, D.-F.; Qin, L. The crosstalk: Exosomes and lipid metabolism. Cell Commun. Signal 2020, 18, 119. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Ding, W.-Q. Intercellular Communication By Exosome-Derived MicroRNAs In Cancer. Int. J. Mol. Sci. 2013, 14, 14240–14269. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Llorente, A. Proteomic analysis of microvesicles released by the human prostate cancer cell line PC-3, M111.012914. Mol. Cell Proteom. 2012, 11, M111.012914–1–M111.012914-11. [Google Scholar] [CrossRef]
- Choi, D.S.; Kim, D.K.; Kim, Y.K.; Gho, Y.S. Proteomics, transcriptomics and lipidomics of exosomes and ectosomes. Proteomics 2013, 13, 1554–1571. [Google Scholar] [CrossRef]
- Laulagnier, K.; Motta, C.; Hamdi, S.; Roy, S.; Fauvelle, F.; Pageaux, J.-F.; Kobayashi, T.; Salles, J.-P.; Perret, B.; Bonnerot, C.; et al. Mast cell- and dendritic cell-derived exosomes display a specific lipid composition and an unusual membrane organization. Biochem. J. 2004, 380, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Aoki, N.; Izumi, Y.; Takeda, H.; Takahashi, M.; Ochiya, T.; Bamba, T. Lipidomic Analysis of Cells and Extracellular Vesicles from High- and Low-Metastatic Triple-Negative Breast Cancer. Metabolites 2020, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Haraszti, R.A.; Didiot, M.-C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J. Extra Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef] [PubMed]
- Pizzinat, N.; Ong-Meang, V.; Bourgailh-Tortosa, F.; Blanzat, M.; Perquis, L.; Cussac, D.; Parini, A.; Poinsot, V. Extracellular vesicles of MSCs and cardiomyoblasts are vehicles for lipid mediators. Biochimie 2020, 178, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Subra, C.; Grand, D.; Laulagnier, K.; Stella, A.; Lambeau, G.; Paillasse, M.; De Medina, P.; Monsarrat, B.; Perret, B.; Silvente-Poirot, B.; et al. Exosomes account for vesicle-mediated transcellular transport of activatable phospholipases and prostaglandins. J. Lip. Res. 2010, 51, 2105–2120. [Google Scholar] [CrossRef] [PubMed]
- Esser, J.; Gehrmann, U.; D’Alexandri, F.L.; Hidalgo-Estévez, A.M.; Wheelock, C.E.; Scheynius, A.; Gabrielsson, S.; Rådmark, O.J. Exosomes from human macrophages and dendritic cells contain enzymes for leukotriene biosynthesis and promote granulocyte migration. J. Allergy Clin. Immunol. 2010, 126, 1032–1040. [Google Scholar] [CrossRef]
- Ong-Meang, V.; Blanzat, M.; Savchenko, L.; Perquis, L.; Guardia, M.; Pizzinat, N.; Poinsot, V. Extracellular Vesicles Produced by the Cardiac Microenvironment Carry Functional Enzymes to Produce Lipid Mediators In Situ. Int. J. Mol. Sci. 2023, 24, 5866. [Google Scholar] [CrossRef] [PubMed]
- Khoushab, S.; Aghmiuni, M.H.; Esfandiari, N.; Sarvandani, M.R.R.; Rashidi, M.; Taheriazam, A.; Entezari, M.; Hashemi, M. Unlocking the Potential of Exosomes in Cancer Research: A Paradigm Shift in Diagnosis, Treatment, and Prevention. Pathol. Res. Pract. 2024, 255, 155214. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.; Shin, S.; Baek, M.C.; Yea, K. Modification of immune cell-derived exosomes for enhanced cancer immunotherapy: Current advances and therapeutic applications. Exp. Mol. Med. 2024, 56, 19–31. [Google Scholar] [CrossRef]
- Huda, M.N.; Nurunnabi, M. Potential Application of Exosomes in Vaccine Development and Delivery. Pharm. Res. 2022, 39, 2635–2671. [Google Scholar] [CrossRef]
- Cheng, L.; Jin, C.; Lv, W.; Ding, Q.; Han, X. Developing a highly stable PLGA-mPEG nanoparticle loaded with cisplatin for chemotherapy of ovarian cancer. PLoS ONE 2011, 6, e25433. [Google Scholar] [CrossRef] [PubMed]
- Lajunen, T.; Viitala, L.; Kontturi, L.-S.; Laaksonen, T.; Liang, H.; Vuorimaa-Laukkanen, E.; Viitala, T.; Le Guével, X.; Yliperttula, M.; Murtomäki, L.; et al. Light induced cytosolic drug delivery from liposomes with gold nanoparticles. J. Control. Release 2015, 203, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Akita, H.; Ishida, R.; Tange, K.; Arai, M.; Kubo, K.; Harashima, H. Neutral biodegradable lipid-envelope-type nanoparticle using vitamin A-scaffold for nuclear targeting of plasmid DNA. Biomaterials 2014, 35, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Thi, E.P.; Mire, C.E.; Lee, A.C.H.; Geisbert, J.B.; Zhou, J.Z.; Agans, K.N.; Snead, N.M.; Deer, D.J.; Barnard, T.R.; Fenton, K.A.; et al. Lipid nanoparticle siRNA treatment of Ebola-virus-Makona-infected nonhuman primates. Nature 2015, 521, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Lee, R.J. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv. Drug Deliv. Rev. 2016, 99, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Dawidczyk, C.M.; Kim, C.; Park, J.H.; Russell, L.M.; Lee, K.H.; Pomper, M.G.; Searson, P.C. State-of-the-art in design rules for drug delivery platforms: Lessons learned from FDA-approved nanomedicines. J. Control. Release 2014, 187, 133–144. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and challenges of liposome assisted drug delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef]
- Xu, X.; Ho, W.; Zhang, X.; Bertrand, N.; Farokhzad, O. Cancer nanomedicine: From targeted delivery to combination therapy. Trends Mol. Med. 2015, 21, 223–232. [Google Scholar] [CrossRef]
- Noble, G.T.; Stefanick, J.F.; Ashley, J.D.; Kiziltepe, T.; Bilgicer, B. Ligand-targeted liposome design: Challenges and fundamental considerations. Trends Biotechnol. 2014, 32, 32–45. [Google Scholar] [CrossRef]
- Yingchoncharoen, P.; Kalinowski, D.S.; Richardson, D.R. Lipid-based drug delivery systems in cancer therapy: What is available and what is yet to come. Pharmacol. Rev. 2016, 68, 701–787. [Google Scholar] [CrossRef] [PubMed]
- Bastos, N.; Ruivo, C.F.; da Silva, S.; Melo, S.A. Exosomes in cancer: Use them or target them? Semin. Cell Dev. Biol. 2017, 78, 13–21. [Google Scholar] [CrossRef]
- Moon, B.; Chang, S. Exosome as a Delivery Vehicle for Cancer Therapy. Cells 2022, 11, 316. [Google Scholar] [CrossRef]
- Mizrak, A.; Bolukbasi, M.F.; Ozdener, G.B.; Brenner, G.J.; Madlener, S.; Erkan, E.P.; Strobel, T.; Breakefield, X.O.; Saydam, O. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol. Ther. 2013, 21, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Ouyang, S.; Li, Y.; Xiao, B.; Yang, H. Immature dendritic cell-derived exosomes: A promise subcellular vaccine for autoimmunity. Inflammation 2013, 36, 232–240. [Google Scholar] [CrossRef] [PubMed]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347. [Google Scholar] [CrossRef]
- Tian, Y.H.; Li, S.P.; Song, J.; Ji, T.J.; Zhu, M.T.; Anderson, G.J.; Wei, J.Y.; Nie, G.J. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014, 35, 2383–2390. [Google Scholar] [CrossRef]
- Yeo, R.W.; Lai, R.C.; Zhang, B.; Tan, S.S.; Yin, Y.; The, B.J.; Lim, S.K. Mesenchymal stem cell: An efficient mass producer of exosomes for drug delivery. Adv. Drug Deliv. Rev. 2013, 65, 336–341. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T.A. review on exosomes application in clinical trials: Perspective, questions, and challenges. Cell Commun. Signal. 2022, 20, 145. [Google Scholar] [CrossRef]
- Mohammadi, M.; Zargartalebi, H.; Salahandish, R.; Aburashed, R.; Yong, K.W.; Sanati-Nezhad, A. Emerging technologies and commercial products in exosome-based cancer diagnosis and prognosis. Biosens. Bioelectron. 2021, 183, 113176. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Sagini, K.; Ferrara, G.; Lanni, M.; Emiliani, C. Exosome-based strategies for Diagnosis and Therapy. Recent Patents CNS Drug Discov. 2015, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Makler, A.; Asghar, W. Exosomal biomarkers for cancer diagnosis and patient monitoring. Expert Rev. Mol. Diagn. 2020, 20, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Yousefpour, P.; Chilkoti, A. Co-opting biology to deliver drugs. Biotechnol. Bioeng. 2014, 111, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Sutaria, D.; Badawi, M.; Phelps, M.; Schmittgen, T. Achieving the promise of therapeutic extracellular vesicles: The devil is in details of therapeutic loading. Pharm. Res. 2017, 34, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Ayala-Mar, S.; González-Valdez, J. State-of-the-art exosome loading and functionalization techniques for enhanced therapeutics: A review. Crit. Rev. Biotechnol. 2020, 40, 804–820. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; Eijndhoven, M.A.J.V.; Hopmans, E.S.; Lindenberg, J.L.; Gruijl, T.D.D.; Wuerdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNA by exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.A.; Larregina, A.T.; Shufesky, W.J.; Sullivan, M.L.G.; Beer Stolz, D.; Papworth, G.D.; Zahorchak, A.F.; Logar, A.J.; Wang, Z.; Watkins, S.C.; et al. Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood 2004, 104, 3257–3266. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Ale Kovi, M.A.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; Garcıà-Santos, G.; Ghajar, C.M. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef]
- Yukawa, H.; Suzuki, K.; Aoki, K.; Arimoto, T.; Yasui, T.; Kaji, N.; Ishikawa, T.; Ochiya, T.; Baba, Y. Imaging of angiogenesis of human umbilical vein endothelial cells by uptake of exosomes secreted from hepatocellular carcinoma cells. Sci. Rep. 2018, 8, 6765. [Google Scholar] [CrossRef]
- Suetsugu, A.; Honma, K.; Saji, S.; Moriwaki, H.; Ochiya, T.; Hoffman, R.M. Imaging exosome transfer from breast cancer cells to stroma at metastatic sites in orthotopic nude-mouse models. Adv. Drug Deliv. Rev. 2013, 65, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zong, S.; Wang, Z.; Lu, J.; Zhu, D.; Zhang, Y.; Cui, Y. Imaging and Intracellular Tracking of Cancer-Derived Exosomes Using Single-Molecule Localization-Based Super-Resolution Microscope. ACS Appl. Mater. Interfaces 2016, 8, 25825–25833. [Google Scholar] [CrossRef] [PubMed]
- Andaloussi, S.E.L.; Lakhal, S.; Mäger, I.; Wood, M.J.A. Exosomes for targeted siRNA delivery across biological barriers. Adv. Drug Deliv. Rev. 2013, 65, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Wahlgren, J.; Karlson, T.D.L.; Brisslert, M.; Vaziri Sani, F.; Telemo, E.; Sunnerhagen, P.; Valadi, H. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res. 2012, 40, e130. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Banizs, A.B.; Huang, T.; Dryden, K.; Berr, S.S.; Stone, J.R.; Nakamoto, R.K.; Shi, W. In vitro evaluation of endothelial exosomes as carriers for small interfering ribonucleic acid delivery. Int. J. Nanomed. 2014, 9, 4223–4230. [Google Scholar] [CrossRef] [PubMed]
- Shtam, T.A.; Kovalev, R.A.; Varfolomeeva, E.Y.; Makarov, E.M.; Kil, Y.V.; Filatov, M.V. Exosomes are natural carriers of exogenous siRNA to human cells in vitro. Cell Commun. Signal. 2013, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Zhang, X.; Zhang, L.; Yuan, M.; Sun, L.; Yu, F.; Wang, B.; Zhao, J.; He, H.; Yang, V.C. Targeted exosomes for co-delivery of siFGL1 and siTGF-β1 trigger combined cancer immunotherapy by remodeling immunosuppressive tumor microenvironment. Chem. Eng. J. 2021, 421, 129774. [Google Scholar] [CrossRef]
- Kamerkar, S.; LeBleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef]
- Huang, W.; Qu, M.; Li, L.; Liu, T.; Lin, M.; Yu, X. SiRNA in MSC derived exosomes silences CTGF gene for locomotor recovery in spinal cord injury rats. Stem Cell Res. Ther. 2021, 12, 334. [Google Scholar] [CrossRef]
- Rong, Y.; Wang, Z.; Tang, P.; Wang, J.; Ji, C.; Chang, J.; Zhu, Y.; Ye, W.; Bai, J.; Liu, W.; et al. Engineered extracellular vesicles for delivery of siRNA promoting targeted repair of traumatic spinal cord injury. Bioact. Mater. 2023, 23, 328–342. [Google Scholar] [CrossRef]
- Haraszt, R.A.; Miller, R.; Didiot, M.; Biscans, A.; Alterman, J.F.; Hassler, M.R.; Roux, L.; Echeverria, D.; Sapp, E.; DiFiglia, M.; et al. Optimized Cholesterol-siRNA Chemistry Improves Productive Loading onto Extracellular Vesicles. Mol. Ther. 2018, 26, 1973–1982. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Cai, G.; Liu, K.; Zhuang, Z.; Jia, K.; Pei, S.; Wang, X.; Wang, H.; Xu, S.; Cui, C.; et al. Microglia exosomal miRNA-137 attenuates ischemic brain injury through targeting Notch1. Aging 2021, 13, 4079–4095. [Google Scholar] [CrossRef]
- Ohno, S.-I.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Shuang, T.; Gao, Y.; Lu, F.; Zhang, J.; He, W.; Qu, L.; Chen, B.; Hao, Q. Targeted delivery of exosomal miR-484 reprograms tumor vasculature for chemotherapy sensitization. Cancer Lett. 2022, 530, 45–58. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Suo, Z. An Effective Peptide-Based Platform for Efficient Exosomal Loading and Cellular Delivery of a microRNA. ACS Appl. Mater. Interfaces 2023, 15, 3851–3866. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Tian, J.; Wang, Z.; Gao, Y.; Wu, X.; Ding, X.; Qiang, L.; Li, G.; Han, Z.; Yuan, Y.; et al. Functional exosome-mediated codelivery of doxorubicin and hydrophobically modified microRNA 159 for triple-negative breast cancer therapy. J. Nanobiotechnol. 2019, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnol. 2020, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Kim, M.; Lee, Y.; Byun, J.W.; Hwang, D.W.; Lee, M. Systemic delivery of microRNA-21 antisense oligonucleotides to the brain using T7-peptide decorated exosomes. J. Control. Release 2020, 317, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Harrison, E.B.; Zhao, Y.; Kabanov, A.V.; Batrakova, E.V. TPP1 Delivery to Lysosomes with Extracellular Vesicles and their Enhanced Brain Distribution in the Animal Model of Batten Disease. Adv. Health Mater. 2019, 8, e1801271. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Guo, H.; Chen, L.; Huang, X. Dapagliflozin-Loaded Exosome Mimetics Facilitate Diabetic Wound Healing by HIF-1a-Mediated Enhancement of Angiogenesis. Adv. Healthc. Mater. 2022, 12, 2202751. [Google Scholar] [CrossRef]
- Fuhrmann, G.; Chandrawati, R.; Parmar, P.A.; Keane, T.J.; Maynard, S.A.; Bertazzo, S.; Stevens, M.M. Engineering Extracellular Vesicles with the Tools of Enzyme Prodrug Therapy. Adv. Mater. 2018, 30, 1706616. [Google Scholar] [CrossRef]
- Rodríguez-Morales, B.; Antunes-Ricardo, M.; González-Valdez, J. Exosome-Mediated Insulin Delivery for the Potential Treatment of Diabetes Mellitus. Pharmaceutics 2021, 13, 1870. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yang, X.; He, C.; Chen, Y.; Li, C.; Li, S.; Chen, Y.; Wu, Y.; Xiang, Z.; Kang, J.; et al. Rejuvenation of tendon stem/progenitor cells for functional tendon regeneration through platelet-derived exosomes loaded with recombinant Yap1. Acta Biomater. 2023, 161, 80–99. [Google Scholar] [CrossRef]
- Hajipour, H.; Farzadi, L.; Roshangar, L.; Latifi, Z.; Kahroba, H.; Shahnazi, V.; Hamdi, K.; Ghasemzadeh, A.; Fattahi, A.; Nouri, M. A human chorionic gonadotropin (hCG) delivery platform using engineered uterine exosomes to improve endometrial receptivity. Life Sci. 2021, 275, 119351. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Gong, C.; Wang, Z.; Xia, Q.; Gu, F.; Hu, C.; Zhang, L.; Guo, H.; Gao, S. A33 antibody functionalized exosomes for targeted delivery of doxorubicin against colorectal cancer. Nanomedicine 2018, 14, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Huang, Q.; Peng, C.; Yao, L.; Chen, H.; Qiu, Z.; Wu, Y.; Wang, L.; Chen, W. Exosomes from M1-polarized macrophages enhance paclitaxel antitumor activity by activating macrophages-mediated inflammation. Theranostics 2019, 9, 1714–1727. [Google Scholar] [CrossRef]
- Zhu, Q.; Ling, X.; Yang, Y.; Zhang, J.; Li, Q.; Niu, X.; Hu, G.; Chen, B.; Li, H.; Wang, Y.; et al. Embryonic stem cells-derived exosomes endowed with targeting properties as chemotherapeutics delivery vehicles for glioblastoma therapy. Adv. Sci. 2019, 6, 1801899. [Google Scholar] [CrossRef] [PubMed]
- Al Faruque, H.; Choi, E.; Kim, J.; Kim, E. Enhanced effect of autologous EVs delivering paclitaxel in pancreatic cancer. J. Control. Release 2022, 347, 330–346. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, C.; Hao, W.; Zhuang, Y.; Liu, X.; Zhao, Y.; Chen, B.; Xiao, Z.; Chen, Y.; Dai, J. NSCs Migration Promoted and Drug Delivered Exosomes-Collagen Scaffold via a Bio-Specific Peptide for One-Step Spinal Cord Injury Repair. Adv. Healthc. Mater. 2021, 10, 2001896. [Google Scholar] [CrossRef]
- Li, Y.-J.; Wu, J.-Y.; Wang, J.-M.; Hu, X.-B.; Cai, J.-X.; Xiang, D.-X. Gemcitabine loaded autologous exosomes for effective and safe chemotherapy of pancreatic cancer. Acta Biomater. 2020, 101, 519–530. [Google Scholar] [CrossRef]
- Zhou, W.; Zhou, Y.; Chen, X.; Ning, T.; Chen, H.; Guo, Q.; Zhang, Y.; Liu, P.; Zhang, Y.; Li, C.; et al. Pancreatic cancer-targeting exosomes for enhancing immunotherapy and reprogramming tumor microenvironment. Biomaterials 2021, 268, 120546. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wan, Z.; Wang, C.; Lu, F.; Wei, M.; Wang, D.; Hao, Q. Designer exosomes for targeted and efficient ferroptosis induction in cancer via chemo-photodynamic therapy. Theranostics 2021, 11, 8185–8196. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Liang, W. ASCs-derived exosomes loaded with vitamin A and quercetin inhibit rapid senescence-like response after acute liver injury. Biochem. Biophys. Res. Commun. 2021, 572, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.M.; Paramonik, A.P.; Sedykh, S.S.; Nevinsky, G.A. Milk Exosomes: Next-Generation Agents for Delivery of Anticancer Drugs and Therapeutic Nucleic Acids. Int. J. Mol. Sci. 2023, 24, 10194. [Google Scholar] [CrossRef] [PubMed]
- Ngu, A.; Wang, S.; Wang, H.; Khanam, A.; Zempleni, J. Milk exosomes in nutrition and drug delivery. Am. J. Physiol. Physiol. 2022, 322, C865–C874. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Xia, B.; Shan, S.; Zheng, A.; Zhang, S.; Chen, J.; Liang, X.J. High-quality milk exosomes as oral drug delivery system. Biomaterials 2021, 277, 121126. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Iglesias-Aguirre, C.E.; Cortés-Martín, A.; Vallejo, F.; Cattivelli, A.; del Pozo-Acebo, L.; Del Saz, A.; Hazas, M.C.L.d.L.; Dávalos, A.; Espín, J.C. Milk-derived exosomes as nanocarriers to deliver curcumin and resveratrol in breast tissue and enhance their anticancer activity. Int. J. Mol. Sci. 2022, 23, 2860. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Han, Y.; An, Y.; Ding, Y.; He, C.; Wang, X.; Tang, Q. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials 2018, 178, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Carobolante, G.; Mantaj, J.; Ferrari, E.; Vllasaliu, D. Cow milk and intestinal epithelial cell-derived extracellular vesicles as systems for enhancing oral drug delivery. Pharmaceutics 2020, 12, 226. [Google Scholar] [CrossRef]
- Yang, X.; Shi, G.; Guo, J.; Wang, C.; He, Y. Exosome-encapsulated antibiotic against intracellular infections of methicillin-resistant Staphylococcus aureus. Int. J. Nanomed. 2018, 13, 8095–8104. [Google Scholar] [CrossRef]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell type-specific and common characteristics of exosomes derived from mouse cell lines: Yield, physico-chemical properties, and pharmacokinetics. Eur. J. Pharm. Sci. 2017, 96, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Toward exosome-based therapeutics: Isolation, heterogeneity, and fit-for-purpose potency. Front. Cardiovasc. Med. 2017, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Riches, A.; Campbell, E.; Borger, E.; Powis, S. Regulation of exosome release from mammary epithelial and breast cancer cells—A new regulatory pathway. Eur. J. Cancer 2014, 50, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.C.; Bayik, D.; Srivatsan, A.; Bergamaschi, C.; Valentin, A.; Niu, G.; Bear, J.; Monninger, M.; Sun, M.; Morales-Kastresana, A.; et al. Efficient production and enhanced tumor delivery of engineered extracellular vesicles. Biomaterials 2016, 105, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Jeske, R.; Liu, C.; Duke, L.; Castro, M.L.C.; Muok, L.; Arthur, P.; Singh, M.; Jung, S.; Sun, L.; Li, Y. Upscaling human mesenchymal stromal cell production in a novel vertical-wheel bioreactor enhances extracellular vesicle secretion and cargo profile. Bioact. Mat. 2023, 25, 732–747. [Google Scholar] [CrossRef] [PubMed]
- Haraszti, R.A.; Miller, R.; Stoppato, M.; Sere, Y.Y.; Coles, A.; Didiot, M.C.; Wollacott, R.; Sapp, E.; Dubuke, M.L.; Li, X.; et al. Exosomes produced from 3D cultures of MSCs by tangential flow filtration show higher yield and improved activity. Mol. Ther. 2018, 26, 2838–2847. [Google Scholar] [CrossRef] [PubMed]
- Harmati, M.; Tarnai, Z.; Decsi, G.; Kormondi, S.; Szegletes, Z.; Janovak, L.; Dekany, I.; Saydam, O.; Gyukity-Sebestyen, E.; Dobra, G.; et al. Stressors alter intercellular communication and exosome profile of nasopharyngeal carcinoma cells. J. Oral Pathol. Med. 2017, 46, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, M.L.; Ilmer, M.; Silva, L.P.; Hawke, D.H.; Recio, A.; Vorontsova, M.A.; Alt, E.; Vykoukal, J. Benchtop isolation and characterization of functional exosomes by sequential filtration. J. Chromatogr. A 2014, 1371, 125–135. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.O.; Saleem, A.A. Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Weng, Y.; Sui, Z.; Shan, Y.; Hu, Y.; Chen, Y.; Zhang, L.; Zhang, Y. Effective isolation of exosomes with polyethylene glycol from cell culture supernatant for in-depth proteome profiling. Analyst 2016, 141, 4640–4646. [Google Scholar] [CrossRef]
- Wang, J.; Ma, P.; Kim, D.H.; Liu, B.-F.; Demirci, U. Towards microfluidic-based exosome isolation and detection for tumor therapy. Nano Today 2021, 37, 101066. [Google Scholar] [CrossRef] [PubMed]
- Willms, E.; Johansson, H.J.; Mäger, I.; Lee, Y.; Blomberg, K.E.M.; Sadik, M.; Alaarg, A.; Smith, C.E.; Lehtiö, J.; EL Andaloussi, S.; et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci. Rep. 2016, 6, 22519. [Google Scholar] [CrossRef] [PubMed]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, A.; Fenaroli, F.; Monopoli, M.P.; Aberg, C.; Dawson, K.A.; Salvati, A. Effects of the presence or absence of a protein corona on silica nanoparticle uptake and impact on cells. ACS Nano 2012, 6, 5845–5857. [Google Scholar] [CrossRef] [PubMed]
- Digiacomo, L.; Cardarelli, F.; Pozzi, D.; Palchetti, S.; Digman, M.A.; Gratton, E.; Capriotti, A.L.; Mahmoudi, M.; Caracciolo, G. An apolipoprotein-enriched biomolecular corona switches the cellular uptake mechanism and trafficking pathway of lipid nanoparticles. Nanoscale 2017, 9, 17254–17262. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.C.; Kim, O.Y.; Yoon, C.M.; Choi, D.-S.; Roh, T.-Y.; Park, J.; Nilsson, J.; Lötvall, J.; Kim, Y.-K.; Gho, Y.S. BCorrection to bioinspired exosome-mimetic nanovesicles for targeted delivery of chemotherapeutics to malignant tumors. ACS Nano 2013, 7, 7698–7710. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, R.; Corbo, C.; Martinez, J.O.; Taraballi, F.; Evangelopoulos, M.; Minardi, S.; Yazdi, I.K.; Zhao, P.; De Rosa, E.; Sherman, M.B.; et al. Biomimetic proteolipid vesicles for targeting inflamed tissues. Nat. Mater. 2016, 15, 1037–1046. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, Y.; Zeng, Y.; Shen, C.; Li, R.; Guo, Z.; Li, S.; Zheng, Q.; Chu, C.; Wang, Z.; et al. Virus-mimetic nanovesicles as a versatile antigen-delivery system. Proc. Natl. Acad. Sci. USA 2015, 112, 6129–6138. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Fang, R.H.; Wang, K.-C.; Luk, B.T.; Thamphiwatana, S.; Dehaini, D.; Nguyen, P.; Angsantikul, P.; Wen, C.H.; Kroll, A.V.; et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature 2015, 526, 118–121. [Google Scholar] [CrossRef]
- Rao, L.; Meng, Q.; Huang, Q.; Wang, Z.; Yu, G.; Li, A.; Ma, W.; Zhang, N.; Guo, S.; Zhao, X.; et al. Platelet–leukocyte hybrid membrane-coated immunomagnetic beads for highly efficient and highly specific isolation of circulating tumor cells. Adv. Funct. Mater. 2018, 28, 1803531. [Google Scholar] [CrossRef]
- Jing, L.; Qu, H.; Wu, D.; Zhu, C.; Yang, Y.; Jin, X.; Zheng, J.; Shi, X.; Yan, X.; Wang, Y. Platelet-camouflaged nanococktail: Simultaneous inhibition of drug-resistant tumor growth and metastasis via a cancer cells and tumor vasculature dual-targeting strategy. Theranostics 2018, 8, 2683–2695. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.W.; Kang, M.; Kim, H.Y.; Han, J.; Kang, S.; Lee, J.-R.; Jeong, G.-J.; Kwon, S.P.; Song, S.Y.; Go, S.; et al. M1 macrophage-derived nanovesicles potentiate the anticancer efficacy of immune checkpoint inhibitors. ACS Nano 2018, 12, 8977–8993. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yu, M.; Xi, Z.; Nie, D.; Dai, Z.; Wang, J.; Qian, K.; Weng, H.; Gan, Y.; Xu, L. Cancer cell membrane-camouflaged nanorods with endoplasmic reticulum targeting for improved antitumor therapy. ACS Appl. Mater. Interfaces 2019, 11, 46614–46625. [Google Scholar] [CrossRef]
- Yanga, J.; Wanga, F.; Lua, Y.; Qia, J.; Denga, L.; Sousac, F.; Sarmentoc, B.; Xua, X.; Cuia, W. Recent advance of erythrocyte-mimicking nanovehicles: From bench to bedside. J. Control. Rel. 2019, 314, 81–91. [Google Scholar] [CrossRef]
- Aryal, S.; Hu, C.M.; Fang, R.H.; Dehaini, D.; Carpenter, C.; Zhang, D.E.; Zhang, L. Erythrocyte membrane-cloaked polymeric nanoparticles for controlled drug loading and release. Nanomedicine 2013, 8, 1271–1280. [Google Scholar] [CrossRef]
- Luk, B.T.; Fang, R.H.; Hu, C.M.; Copp, J.A.; Thamphiwatana, S.; Dehaini, D.; Gao, W.; Zhang, K.; Li, S.; Zhang, L. Safe and Immunocompatible nanocarriers cloaked in RBC membranes for drug delivery to treat solid tumors. Theranostics 2016, 6, 1004–1011. [Google Scholar] [CrossRef]
- Fu, Q.; Lv, P.; Chen, Z.; Ni, D.; Zhang, L.; Yue, H.; Yue, Z.; Wei, W.; Ma, G. Programmed co-delivery of paclitaxel and doxorubicin boosted by camouflaging with erythrocyte membrane. Nanoscale 2015, 7, 4020–4030. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Xu, P.; Yao, J.; Li, R.; Gong, L.; Yin, Y.; Lin, Z. Erythrocyte-mimicking paclitaxel nanoparticles for improving biodistributions of hydrophobic drugs to enhance antitumor efficacy. Drug Deliv. 2020, 27, 387–399. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Gnanasammandhan, M.K.; Xie, C.; Huang, K.; Cui, M.Y.; Chan, J.M. Monocyte cell membrane-derived nanoghosts for targeted cancer therapy. Nanoscale 2016, 8, 6981–6985. [Google Scholar] [CrossRef]
- Kang, T.; Zhu, Q.; Wei, D.; Feng, J.; Yao, J.; Jiang, T.; Song, Q.; Wei, X.; Chen, H.; Gao, X.; et al. Nanoparticles Coated with Neutrophil Membranes Can Effectively Treat Cancer Metastasis. ACS Nano 2017, 11, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Wang, H.; Williams, G.R.; Yang, Y.; Zheng, Y.; Wu, J.; Zhu, L.M. Erythrocyte membrane cloaked curcumin-loaded nanoparticles for enhanced chemotherapy. Pharmaceutics 2019, 11, 429. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, G.; Ran, D.; Krishnan, N.; Fang, R.H.; Gao, W.; Spector, S.A.; Zhang, L. T-Cell-Mimicking nanoparticles can neutralize HIV infectivity. Adv. Mater. 2018, 30, e1802233. [Google Scholar] [CrossRef] [PubMed]
- Nie, D.; Dai, Z.; Li, J.; Yang, Y.; Xi, Z.; Wang, J.; Zhang, W.; Qian, K.; Guo, S.; Zhu, C.; et al. Cancer-cell membrane- coated nanoparticles with a yolk–shell structure augment cancer chemotherapy. Nano Lett. 2019, 20, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Cui, Y.; Hao, W.; Chen, M.; Liu, Q.; Wang, Y.; Yang, M.; Li, Z.; Gong, W.; Song, S.; et al. Carrier-free highly drug-loaded biomimetic nanosuspensions encapsulated by cancer cell membrane based on homology and active targeting for the treatment of glioma. Bioact. Mater. 2021, 6, 4402–4414. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Mei, Q.; Li, J.; Zhai, Y.; Chen, Y.; Wang, R.; Lu, E.; Zhang, X.-Y.; Zhang, Z.; Sha, X. Preferential targeting cerebral ischemic lesions with cancer cell-inspired nanovehicle for ischemic stroke treatment. Nano Lett. 2021, 21, 3033–3043. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chen, X.; Bu, W.; Meng, L.; Wang, C.; Jin, N.; Chen, Y.; Ren, C.; Zhang, K.; Sun, H. Modification of Metal-Organic Framework Nanoparticles Using Dental Pulp Mesenchymal Stem Cell Membranes to Target Oral Squamous Cell Carcinoma. J Colloid Interface Sci. 2021, 601, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Pisano, S.; Pierini, I.; Gu, J.; Gazze, A.; Francis, L.W.; Gonzalez, D.; Conlan, R.S.; Corradetti, B. Immune (cell) derived exosome mimetics (IDEM) as a treatment for ovarian cancer. Front. Cell Dev. Biol. 2020, 8, 553576. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Deng, J.; Yao, X.; He, Y.; Li, H.; Jian, Z.; Tang, Y.; Zhang, X.; Zhang, J.; Dai, H. Bone-targeted erythrocyte-cancer hybrid membrane-camouflaged nanoparticles for enhancing photothermal and hypoxia-activated chemotherapy of bone invasion by OSCC. J. Nanobiotechnol. 2021, 19, 342. [Google Scholar] [CrossRef]
- Rao, L.; Wu, L.; Liu, Z.; Tian, R.; Yu, G.; Zhou, Z.; Yang, K.; Xiong, H.-G.; Zhang, A.; Yu, G.-T.; et al. Hybrid cellular membrane nanovesicles amplify macrophage immune responses against cancer recurrence and metastasis. Nat. Commun. 2020, 11, 4909. [Google Scholar] [CrossRef]
- Rao, L.; Bu, L.; Ma, L.; Wang, W.; Liu, H.; Wan, D.; Liu, J.; Li, A.; Guo, S.; Zhang, L.; et al. Platelet -Facilitated Photothermal Therapy of Head and Neck Squamous Cell Carcinoma. Angew. Chem. Int. 2018, 130, 998–1003. [Google Scholar] [CrossRef]
- Bu, L.; Rao, L.; Yu, G.; Chen, L.; Deng, W.; Liu, J.; Wu, H.; Meng, Q.; Guo, S.; Zhao, X.; et al. Cancer Stem Cell-Platelet Hybrid Membrane-Coated Magnetic Nanoparticles for Enhanced Photothermal Therapy of Head and Neck Squamous Cell Carcinoma. Adv. Funct. Mater. 2019, 29, 1807733. [Google Scholar] [CrossRef]
- Rao, L.; He, Z.; Meng, Q.-F.; Zhou, Z.; Bu, L.-L.; Guo, S.-S.; Liu, W.; Zhao, X.Z. Effective cancer targeting and imaging using macrophage membrane-camouflaged upconversion nanoparticles. J. Biomed. Mater. Res. A 2017, 105, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, J.; Cai, X.; Ding, P.; Lv, H.; Pei, R. Metal–Organic Frameworks with Enhanced Photodynamic Therapy: Synthesis, Erythrocyte Membrane Camouflage, and Aptamer-Targeted Aggregation. ACS Appl. Mater. Interfaces 2020, 12, 23697–23706. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhong, Y.; Fan, W.; Xiang, J.; Wang, G.; Zhou, Q.; Wang, J.; Geng, Y.; Sun, R.; Zhang, Z.; et al. Enhanced tumour penetration and prolonged circulation in blood of polyzwitterion–drug conjugates with cell-membrane affinity. Nat. Biomed. Eng. 2021, 5, 1019–1037. [Google Scholar] [CrossRef] [PubMed]
- van Dommelen, S.M.; Vader, P.; Lakhal, S.; Kooijmans, S.; van Solinge, W.W.; Wood, M.J.; Schiffelers, R.M. Microvesicles and exosomes: Opportunities for cell-derived membrane vesicles in drug delivery. J. Control. Release 2012, 161, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in exosome isolation techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Yang, G.; Liu, Y.; Liu, T. Research progress in membrane fusion-based hybrid exosomes for drug delivery systems, Front. Bioeng. Biotechnol. 2022, 10, 939441. [Google Scholar] [CrossRef]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.-A.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, X.; Tang, J.; Lv, Q.; Liu, J. Gene-engineered exosomes-thermosensitive liposomes hybrid nanovesicles by the blockade of CD47 signal for combined photothermal therapy and cancer immunotherapy. Biomaterials 2021, 275, 120964. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, J.; Gu, W.; Huang, Y.; Tong, Z.; Huang, L.; Tan, J. Exosome-liposome hybrid nanoparticles deliver CRISPR/Cas9 system in MSCs. Adv. Sci. 2018, 5, 1700611. [Google Scholar] [CrossRef] [PubMed]
- Jhan, Y.-Y.; Prasca-Chamorro, D.; Zuniga, G.P.; Moore, D.M.; Kumar, S.A.; Gaharwar, A.K.; Bishop, C.J. Engineered extracellular vesicles with synthetic lipids via membrane fusion to establish efficient gene delivery. Int. J. Pharm. 2020, 573, 118802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Miao, Y.; Wang, Y.; He, S.; Guo, L.; Mao, J.; Chen, M.; Yang, Y.; Zhang, X.; Gan, Y. Tumour-derived extracellular vesicle membrane hybrid lipid nanovesicles enhance siRNA delivery by tumour-homing and intracellular freeway transportation. J. Extracell. Vesicle 2022, 11, e12198. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, X.; Zhang, Q.; Gu, Z.; Luo, Y.; Guo, J.; Wang, X.; Jing, Y.; Chen, X.; Su, J. Exosome-guided bone targeted delivery of Antagomir-188 as an anabolic therapy for bone loss. Bioact. Mater. 2021, 6, 2905–2913. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.J.W.; van de Wakker, S.I.; de Groot, E.M.; de Jong, O.G.; Gitz-François, J.J.J.; Seinen, C.S.; Sluijter, J.P.G.; Schiffelers, R.M.; Vader, P. Functional siRNA delivery by extracellular vesicle-liposome hybrid nanoparticles. Adv. Healthc. Mater. 2021, 11, 2101202. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhang, J.; Kong, L.; Yu, Y.; Hu, Q.; Yang, T.; Wang, Y.; Tu, K.; Qiao, Q.; Qin, X.; et al. Immunogenic hybrid nanovesicles of liposomes and tumor-derived nanovesicles for cancer immunochemotherapy. ACS Nano 2021, 15, 3123–3138. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, Y.; Shi, J. Exosome biochemistry and advanced nanotechnology for next-generation theranostic platforms. Adv. Mater. 2019, 31, 1802896. [Google Scholar] [CrossRef] [PubMed]
- Rayamajhi, S.; Nguyen, T.D.T.; Marasini, R.; Aryal, S. Macrophage-derived exosome-mimetic hybrid vesicles for tumor targeted drug delivery. Acta Biomater. 2019, 94, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, X.; Zhao, H.; Li, N.; Li, J.; Zhang, H.; Di, L. Targeted delivery of hybrid nanovesicles for enhanced brain penetration to achieve synergistic therapy of glioma. J. Control. Release 2024, 365, 331–347. [Google Scholar] [CrossRef]
- Sun, L.; Fan, M.; Huang, D.; Li, B.; Xu, R.; Gao, F.; Chen, Y. Clodronate-loaded liposomal and fibroblast-derived exosomal hybrid system for enhanced drug delivery to pulmonary fibrosis. Biomaterials 2021, 271, 120761. [Google Scholar] [CrossRef]
- Ji, K.; Fan, M.; Huang, D.; Sun, L.; Li, B.; Xu, R.; Zhang, J.; Shao, X.; Chen, Y. Clodronate-nintedanib-loaded exosome-liposome hybridization enhances the liver fibrosis therapy by inhibiting Kupffer cell activity. Biomater. Sci. 2022, 10, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; He, D.; Guo, Q.; Zhang, Z.; Ru, D.; Wang, L.; Gong, K.; Liu, F.; Duan, Y.; Li, H. Exosome-liposome hybrid nanoparticle codelivery of TP and miR497 conspicuously overcomes chemoresistant ovarian cancer. J. Nanobiotechnol. 2022, 20, 50. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, Y.; Han, R.; Li, Y.; Zhai, Y.; Qian, Z.; Gu, Y.; Li, S. A cascade synergetic strategy induced by photothermal effect based on platelet exosome nanoparticles for tumor therapy. Biomaterials 2022, 282, 121384. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Cheng, L.; Lu, Y.; Zhang, X.; Wang, Y.; Deng, J.; Zhou, J.; Liu, B. Thermosensitive exosome-liposome hybrid nanoparticle-mediated chemoimmunotherapy for improved treatment of metastatic peritoneal cancer. Adv. Sci. 2020, 7, 2000515. [Google Scholar] [CrossRef] [PubMed]


| Pathology Type | Pathology | Study | Type | Start Date | NCT Number |
|---|---|---|---|---|---|
| Blood pathology | Sepsis | Function of circulating exosomes in sepsis-induced immunosuppression | O | 15/04/2021 | NCT04979767 |
| Brain disease | Alzheimer’s disease | Safety and efficacy evaluation of allogenic adipose MSC-Exos in patients with Alzheimer’s disease | I | 01/07/2020 | NCT04388982 |
| Brain disease | Parkinson’s disease | LRRK2 and other novel exosome proteins in Parkinson’s disease: biomarkers associated with Parkinson’s disease susceptibility and/or progression in exosome-proteomes derived | O | 01/01/2013 | NCT01860118 |
| Cancer | Bladder cancer | Characterization of the protein profile in tumor derived exosomes from the gallbladder carcinoma patients | O | 01/01/2018 | NCT03581435 |
| Cancer | Bladder cancer | Use of urinary exosome lncRNAs for preoperative diagnosis of lymphatic metastasis in patients with bladder cancer | O | 01/06/2023 | NCT05270174 |
| Cancer | Bladder cancer | Use of urine exosomal RNA for accurate diagnosis of urinary tract tumors and the development of kits | O | 01/06/2023 | NCT06193941 |
| Cancer | Bone cancer | Identification and characterization of miRNAs content in circulating tumor exosomes in bone metastases | O | 03/12/2018 | NCT03895216 |
| Cancer | Brain cancer | Clinical relevance of detecting molecular abnormalities in glial tumor exosomes (blood sampling) | I | 15/12/2023 | NCT06116903 |
| Cancer | Breast cancer | Omic technologies to track resistance to palbociclib in metastatic breast cancer: longitudinal changes in volatile organic components profile and exosomes according to response to treatment | I | 08/09/2020 | NCT04653740 |
| Cancer | Breast cancer | Development of a horizontal data integration classifier for noninvasive early diagnosis of breast cancer (from different radiomics analyses on baseline mammograms and molecular analyses on peripheral blood (miRNA sequencing from exosomes) and urine | I | 19/01/2021 | NCT04781062 |
| Cancer | Breast cancer | Exosome as prognostic and predictive biomarker in early breast cancer patients | I | 01/05/2021 | NCT05955521 |
| Cancer | Breast cancer | Feasibility of exosome analysis in cerebrospinal fluid during the diagnostic workup of metastatic meningitis (Exo-LCR) | I | 04/01/2023 | NCT05286684 |
| Cancer | Breast cancer | Study to measure the expression of the HER2-HER3 dimer in tumor and blood (exosomes) samples from patients with HER2 positive breast cancer receiving HER2 targeted therapies | O | 20/12/2019 | NCT04288141 |
| Cancer | Cancer | Pilot study with the aim to quantify a stress protein in the blood and in the urine for the monitoring and early diagnosis of malignant solid tumors: concentration of HSP70 exosomes in the blood and urine | I | 15/12/2015 | NCT02662621 |
| Cancer | Colon cancer | Study investigating the ability of plant exosomes to deliver curcumin to normal and colon cancer tissue | I | 01/01/2011 | NCT01294072 |
| Cancer | Colorectal cancer | Contents of circulating extracellular vesicles: prognostic role of exosomes and their contents on the survival of colorectal cancer patients | O | 01/07/2020 | NCT04523389 |
| Cancer | Colorectal cancer | Identification in blood sample of new diagnostic protein markers derived from circulating tumor exosomes for colorectal cancer | O | 07/01/2021 | NCT04394572 |
| Cancer | Epithelial cancer Bowel disease | Plant exosomes +/− curcumin to abrogate symptoms of inflammatory bowel disease | I | 01/03/2018 | NCT04879810 |
| Cancer | Gastric cancer | Circulating exosomes as potential prognostic and predictive biomarkers in advanced gastric cancer patients (“EXO-PPP study“): characterization of the molecular profile in tumor derived exosomes, correlation of plasma level and kinetics of gastric cancer derived exosomes | O | 01/01/2013 | NCT01779583 |
| Cancer | Liver cancer | Early plasmatic biomarkers of tumor response in high dose hypofractionated radiotherapy. Work Package 3: Immune response (analyses of immunological parameters, description of secreted markers and nanovesicles production, verification of the presence and evolution of activation markers and quantification of secreted exosomes) | I | 16/09/2015 | NCT02439008 |
| Cancer | Lung cancer | Prediction of immunotherapeutic effect of advanced non-small cell lung cancer (detection of the difference of miRNA expression profiles of exosomes in NSCLC patients before and after immunotherapy -pabolizumab, nafulizumab-) | I | 08/06/2020 | NCT04427475 |
| Cancer | Lung cancer | Molecular profiling of exosomes in tumor-draining vein of early-staged lung cancer | I | 21/06/2021 | NCT04939324 |
| Cancer | Lung cancer | Combined diagnosis of cancerous tissue and exosome in early lung cancer | O | 20/05/2018 | NCT03542253 |
| Cancer | Lung cancer | Early diagnosis of lung cancer using blood plasma derived exosome (evaluating the possibility of distinguishing between normal and lung cancer patients through the analysis of lung cancer-specific exosomal protein) | O | 09/04/2020 | NCT04529915 |
| Cancer | Lung cancer | Study of exosome EML4-ALK fusion in NSCLC clinical diagnosis and dynamic monitoring (the objective response rate of those NSCLC patients receiving ALK inhibitors treatment according to exosome ALK fusion diagnosis and fluorescence in situ hybridization examination) | O | 01/08/2020 | NCT04499794 |
| Cancer | Lung cancer | Serum exosomal miRNA predicting the therapeutic efficiency in lung squamous carcinoma | O | 01/04/2022 | NCT05854030 |
| Cancer | Lung cancer | Extracellular vesicles (isolate exosomes from bronchial washings) and particles as biomarkers of recurrence in non-small cell lung cancer | O | 14/06/2022 | NCT05424029 |
| Cancer | Lymphoma, B-cellaggressive non-Hodgkin (B-NHL) | Exosomes and immunotherapy in non-Hodgkin B-cell lymphomas (quantification of CD20 and PDL-1 in exosomes purified from cell cultures of DLBCL human cell lines and from healthy volunteers) | I | 02/07/2019 | NCT03985696 |
| Cancer | Melanoma | Analysis of circulating exosomes in plasma of melanoma patients (dosage of proteic biomarkers in circulating exosome) | O | 01/03/2019 | NCT05744076 |
| Cancer | Oropharyngeal cancer | Exosome testing as a screening modality for human papillomavirus-positive oropharyngeal squamous cell carcinoma | O | 25/02/2015 | NCT02147418 |
| Cancer | Ovarian cancer | Non-coding RNA in the exosome of the epithelia ovarian cancer (expression of miRNA/lncRNA, expression of micro-RNA (miRNA) and long non-coding RNA (lncRNA) compared between high grade serous ovarian carcinoma group and control group) | O | 10/11/2018 | NCT03738319 |
| Cancer | Pancreatic cancer | Trial of ascorbic acid + triple therapy with nanoparticle paclitaxel protein bound + cisplatin + gemcitabine in patients with advanced stage IV metastatic pancreatic cancer | I | 15/12/2017 | NCT03410030 |
| Cancer | Pancreatic cancer | Circulating extracellular exosomal small RNA as potential biomarker for human pancreatic cancer | I | 01/11/2020 | NCT04636788 |
| Cancer | Pancreatic cancer | iExosomes in Treating Participants With Metastatic Pancreas Cancer With KrasG12D Mutation (Mesenchymal Stromal Cells-derived Exosomes with KRAS G12D siRNA) | I | 27/01/2021 | NCT03608631 |
| Cancer | Prostate cancer | Exosomal microRNA in predicting the aggressiveness of prostate cancer in Chinese patients by comparison of the differences in microRNA expression between non-prostate cancer subjects, pathologically insignificant and significant prostate cancer patients | O | 03/05/2018 | NCT03911999 |
| Cancer | Prostate cancer | ExoDx prostate evaluation in prior negative prostate biopsy setting | O | 15/03/2020 | NCT04357717 |
| Cancer | Rectal cancer | Exosomes in rectal cancer (characterization of plasmatic exosomal biomarker levels in patients with locally advanced rectal cancer undergoing neoadjuvant chemoradiation therapy) | O | 13/02/2018 | NCT03874559 |
| Cancer | Renal cancer | Evaluation of urinary exosomes presence from clear cell renal cell carcinoma | O | 29/01/2020 | NCT04053855 |
| Cancer | Renal cancer | A companion diagnostic study to develop circulating exosomes as predictive biomarkers for the response to immunotherapy in renal cell carcinoma (blood and urine collection) | O | 01/01/2023 | NCT05705583 |
| Cancer | Sarcoma | Study of blood exosomes in monitoring patients with sarcoma (EXOSARC) | O | 19/11/2018 | NCT03800121 |
| Cancer | Systemic autoimmune diseases | Urine miRNAs-exosomes to identify biomarkers for lupus nephritis | O | 02/08/2020 | NCT04894695 |
| Chronic disease | Diabetes type 2 | Determine the levels of circulating extracellular vesicles released by human islets of langerhans in plasma | O | 01/12/2016 | NCT03106246 |
| Chronic disease | Diabetes type 2 | Mechanisms behind severe insulin resistance during pregnancy in women with glucose metabolic disorders (SIR-MET) maternal hormonal, inflammatory and metabolic markers in the blood, as well as the level, content and bioactivity of exosomes are studied | O | 01/05/2021 | NCT04924504 |
| Chronic disease | Diabetic retinopathy | Role of the serum exosomal miRNA (sequencing) in diabetic retinopathy | O | 01/07/2018 | NCT03264976 |
| Chronic disease | Diabetic retinopathy | Study on exosome changes from plasma, atrial fluid and vitreous fluid in patients with proliferative diabetic retinopathy by proteomic analysis of proteins | O | 01/01/2024 | NCT06198543 |
| Chronic disease | Diabetic retinopathy | Proteomic study of plasma exosomes in patients with diabetic retinopathy | O | 02/01/2024 | NCT06188013 |
| Chronic disease | Hypertension | New biomarkers and difficult-to-treat hypertension; concentrations and variabilities of urinary exosomal sodium channels and plasma angiotensins in patients with difficult-to-treat arterial hypertension | O | 01/05/2016 | NCT03034265 |
| Endothelial pathology | Endothelial dysfunction | Circulating exosomes and endothelial dysfunction in patients with obstructive sleep apneas hypopneas syndrome: miRNA contained in exosomes, comparison of exosome content between obese-OSA patients with endothelial dysfunction and without | I | 14/01/2021 | NCT04459182 |
| Genetic disase | Dystrophic epidermolysis bullosa | Study to assess the effectiveness and safety of AGLE-102, an allogeneic derived MSC EVs product derived from normal donor on lesions in subjects with dystrophic epidermolysis bullosa | I | 01/01/2024 | NCT04173650 |
| Heart pathology | Cardiac remodeling | Exosome as integrative tool for prognostic stratification of adverse cardiac remodeling in stemi patients: the MIRACLE Study (verify whether the profile of circulating plasma exosomes reflects cardiovascular magnetic resonance in acute ST-segment–elevation myocardial infarction) | O | 03/05/2023 | NCT06070974 |
| Heart pathology | Atrial fibrillation | Role of exosomes derived from epicardial fat in atrial fibrillation: analysis of the differences in the quantity of exosomes derived epicardial fat biopsy in patients with and without atrial fibrillation. | I | 21/01/2018 | NCT03478410 |
| Infectious disease | COVID-19 | Intravenous infusion of CAP-1002 (cardiosphere-derived cells) in patients with COVID-19 | I | 15/11/2020 | NCT04623671 |
| Infectious disease | COVID-19 | The use of exosomes, delivered intravenously-ARDOXSO™, for the treatment of acute respiratory distress syndrome or novel coronavirus pneumonia caused by COVID-19 | I | 2023-09 | NCT04798716 |
| Infectious disease | HIV | Inflammation, NK cells, antisense protein and exosomes, and correlation with immune response during HIV infection | I | 22/04/2022 | NCT05243381 |
| Respiratory pathology | Acute respiratory distress syndrome | Screening of differential miRNAs of inflammatory exosomes in plasma and alveolar lavage fluid of patients with sepsis complicated with acute respiratory distress syndrome | O | 25/07/2022 | NCT05476029 |
| Respiratory pathology | Obstructive sleep apnea syndrome | Exosomes implication in PD1-PD-L1 activation in obstructive sleep apnea syndrome | O | 03/03/2019 | NCT03811600 |
| Respiratory pathology | Acute respiratory distress syndrome | Safety and efficacy of EXO-CD24 in preventing clinical deterioration in patients with mild–moderate acute respiratory distress syndrome | I | 04/07/2023 | NCT05947747 |
| Respiratory pathology | Respiratory dysfunction | Blood circulating microRNAs as biomarkers of respiratory dysfunction in patients with refractory epilepsy | I | 16/03/2018 | NCT03419000 |
| Systemic autoimmune diseases | Psoriasis | Safety and tolerability study of MSC exosome ointment | I | 08/03/2022 | NCT05523011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poinsot, V.; Pizzinat, N.; Ong-Meang, V. Engineered and Mimicked Extracellular Nanovesicles for Therapeutic Delivery. Nanomaterials 2024, 14, 639. https://doi.org/10.3390/nano14070639
Poinsot V, Pizzinat N, Ong-Meang V. Engineered and Mimicked Extracellular Nanovesicles for Therapeutic Delivery. Nanomaterials. 2024; 14(7):639. https://doi.org/10.3390/nano14070639
Chicago/Turabian StylePoinsot, Verena, Nathalie Pizzinat, and Varravaddheay Ong-Meang. 2024. "Engineered and Mimicked Extracellular Nanovesicles for Therapeutic Delivery" Nanomaterials 14, no. 7: 639. https://doi.org/10.3390/nano14070639