
Application of Bacteriophages on Shiga Toxin-Producing Escherichia coli (STEC) Biofilm



Abstract
:1. Introduction
2. Results
2.1. Biofilm Formation Assay
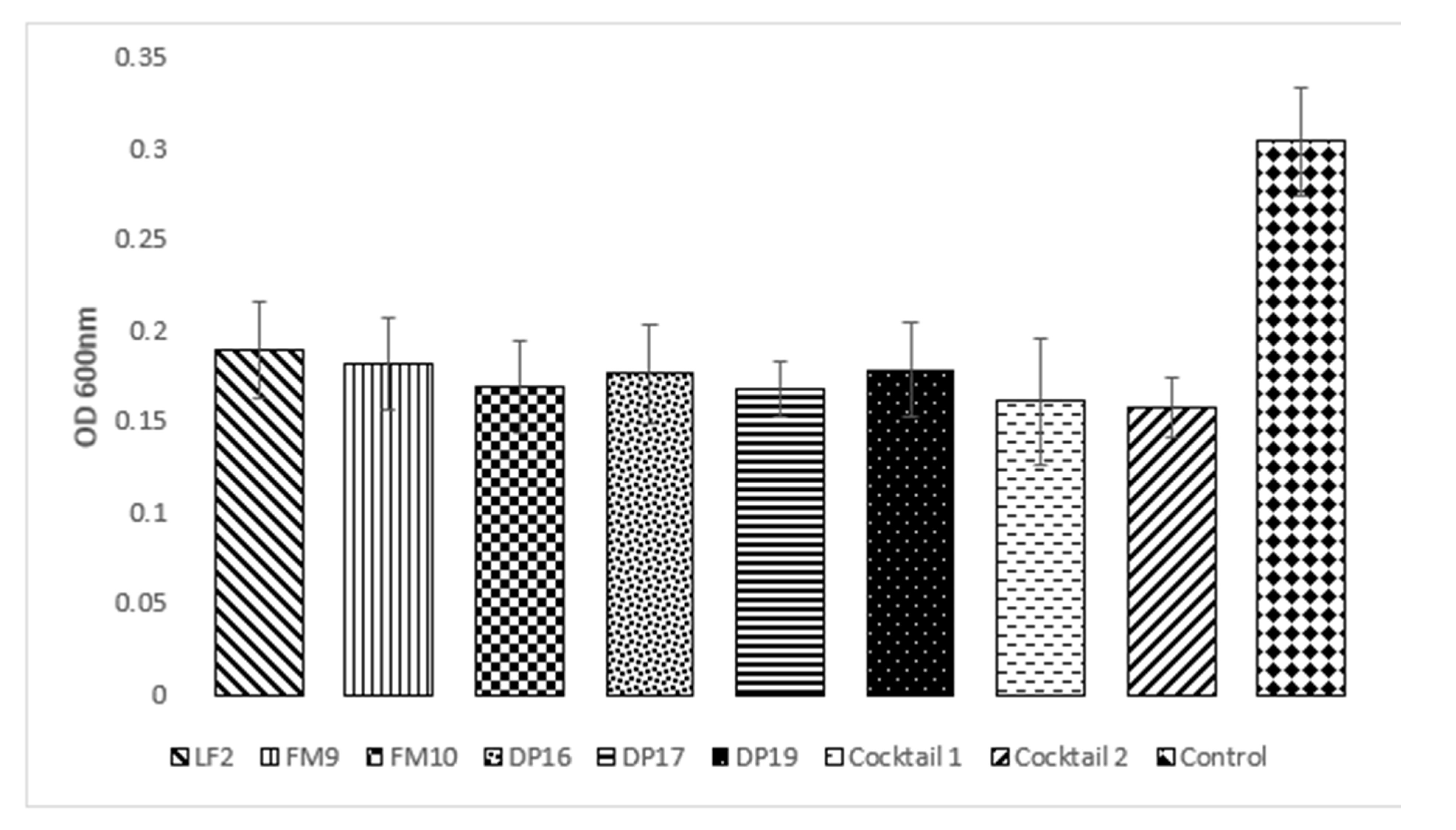
2.2. Effects of Bacteriophages on Biofilm Prevention
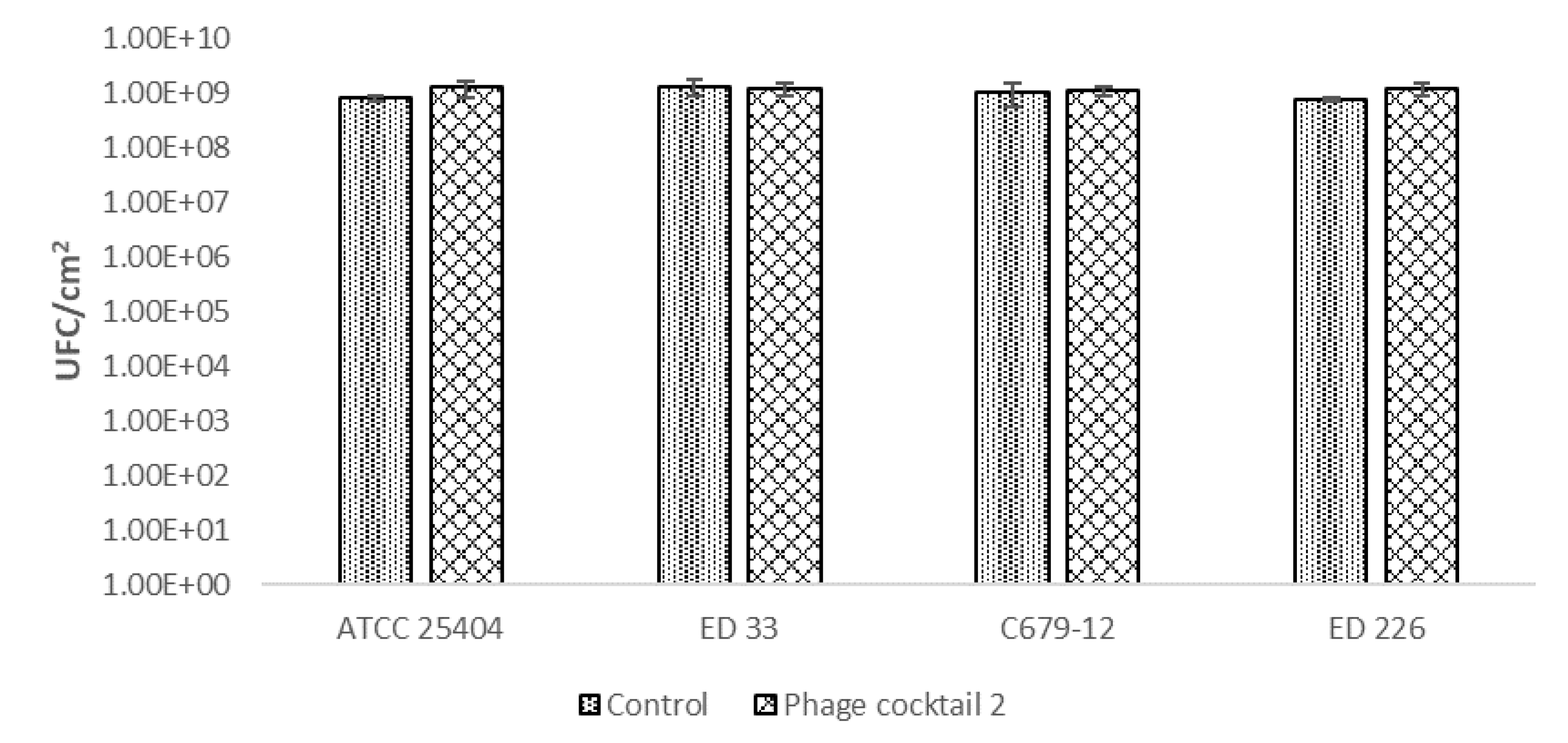
2.3. Effects of Bacteriophages on Already Formed Biofilms
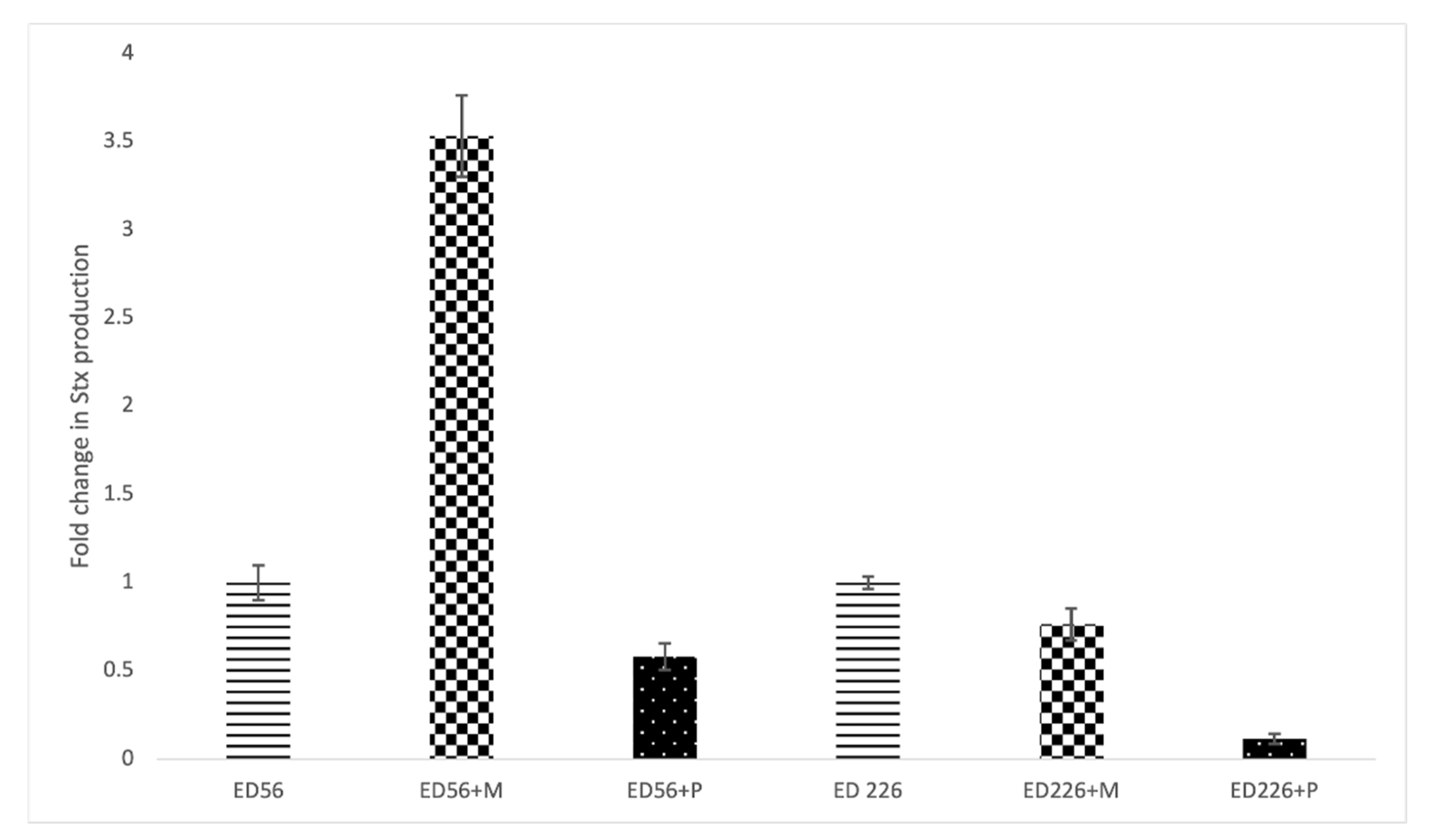
2.4. Effects of Bacteriophages on Stx Production
3. Discussion
4. Materials and Methods
4.1. Bacterial Cultures
4.2. Bacteriophage Propagation
4.3. Biofilm Formation Assay
4.4. Test for Estimating Biofilm Prevention
4.5. Removal of the Formed Biofilm
4.6. Relative Quantification of Stx
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, 1001923. [Google Scholar] [CrossRef] [Green Version]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef] [Green Version]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Pathogenicity assessment of shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, e05967. [Google Scholar] [CrossRef]
- Van Houdt, R.; Michiels, C.W. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Bono, J.L.; Kalchayanand, N.; Shackelford, S.; Harhay, D.M. Biofilm formation by shiga toxin–producing Escherichia coli O157:H7 and non-O157 strains and their tolerance to sanitizers commonly used in the food processing environment. J. Food Prot. 2012, 75, 1418–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestby, L.K.; Møretrø, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonella are correlated with persistence in fish meal- and feed factories. BMC Vet. Res. 2009, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møretrø, T.; Langsrud, S. Listeria monocytogenes: Biofilm formation and persistence in food-processing environments. Biofilms 2004, 1, 107–121. [Google Scholar] [CrossRef]
- Yang, X.; He, A.; Badoni, M.; Tran, F.; Wang, H. Mapping sources of contamination of Escherichia coli on beef in the fabrication facility of a commercial beef packing plant. Food Control 2017, 75, 153–159. [Google Scholar] [CrossRef]
- Solheim, H.T.; Sekse, C.; Urdahl, A.M.; Wasteson, Y.; Nesse, L.L. Biofilm as an environment for dissemination of stx genes by transduction. Appl. Environ. Microbiol. 2013, 79, 896–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogeleer, P.; Tremblay, Y.D.N.; Mafu, A.A.; Jacques, M.; Harel, J. Life on the outside: Role of biofilms in environmental persistence of Shiga-toxin producing Escherichia coli. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahid, I.K.; Ha, S.-D. A review of microbial biofilms of produce: Future challenge to food safety. Food Sci. Biotechnol. 2012, 21, 299–316. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Yan, J.; Mao, J.; Xie, J. Bacteriophage polysaccharide depolymerases and biomedical applications. BioDrugs 2014, 28, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [Green Version]
- Kazi, M.; Annapure, U.S. Bacteriophage biocontrol of foodborne pathogens. J. Food Sci. Technol. 2016, 53, 1355–1362. [Google Scholar] [CrossRef] [Green Version]
- Ferriol-González, C.; Domingo-Calap, P. Phages for biofilm removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Örmälä, A.-M.; Jalasvuori, M. Phage therapy: Should bacterial resistance to phages be a concern, even in the long run? Bacteriophage 2013, 3, e24219. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, P.D.; Buckling, A.; Hall, A.R. Experimental evolution and bacterial resistance: (Co)evolutionary costs and trade-offs as opportunities in phage therapy research. Bacteriophage 2015, 5, e1050153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Naves, P.; Del Prado, G.; Huelves, L.; Gracia, M.; Ruiz, V.; Blanco, J.; Rodríguez-Cerrato, V.; Ponte, M.C.; Soriano, F. Measurement of biofilm formation by clinical isolates of Escherichia coli is method-dependent. J. Appl. Microbiol. 2008, 105, 585–590. [Google Scholar] [CrossRef]
- Picozzi, C.; Antoniani, D.; Vigentini, I.; Foschino, R. Genotypic characterization and biofilm formation of shiga-toxin producing Escherichia coli. FEMS Microbiol. Lett. 2016, 364, fnw291. [Google Scholar] [CrossRef]
- Orskov, I.; Orskov, F.; Jann, B.; Jann, K. Serology, chemistry, and genetics of O and K antigens of Escherichia coli. Bacteriol. Rev. 1977, 41, 667–710. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial biofilms in the food industry—A comprehensive review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef]
- Ahiwale, S.; Prakash, D.; Gajbhiye, M.; Jagdale, S.; Patil, N.; Kapadnis, B. BVPaP-3, a T7-like lytic phage of pseudomonas aeruginosa: Its isolation and characterisation. Curr. Microbiol. 2012, 64, 305–311. [Google Scholar] [CrossRef]
- Pires, D.; Sillankorva, S.; Faustino, A.; Azeredo, J. Use of newly isolated phages for control of Pseudomonas aeruginosa PAO1 and ATCC 10145 biofilms. Res. Microbiol. 2011, 162, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanlon, G.W.; Denyer, S.P.; Olliff, C.J.; Ibrahim, L.J. Reduction in exopolysaccharide viscosity as an aid to bacteriophage penetration through Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2001, 67, 2746–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knezevic, P.; Kostanjsek, R.; Obreht, D.; Petrovic, O. Isolation of Pseudomonas aeruginosa specific phages with broad activity spectra. Curr. Microbiol. 2009, 59, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, P.; Obreht, D.; Curcin, S.; Petrusic, M.; Aleksic, V.; Kostanjsek, R.; Petrovic, O. Phages of Pseudomonas aeruginosa: Response to environmental factors and in vitro ability to inhibit bacterial growth and biofilm formation. J. Appl. Microbiol. 2011, 111, 245–254. [Google Scholar] [CrossRef]
- Bedi, M.S.; Verma, V.; Chhibber, S. Amoxicillin and specific bacteriophage can be used together for eradication of biofilm of Klebsiella pneumoniae B5055. World J. Microbiol. Biotechnol. 2009, 25, 1145–1151. [Google Scholar] [CrossRef]
- Chibeu, A.; Lingohr, E.J.; Masson, L.; Manges, A.; Harel, J.; Ackermann, H.W.; Kropinski, A.M.; Boerlin, P. Bacteriophages with the ability to degrade uropathogenic Escherichia Coli biofilms. Viruses 2012, 4, 471–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bumunang, E.W.; Ateba, C.N.; Stanford, K.; Niu, Y.D.; Wang, Y.; McAllister, T.A. Activity of bacteriophage and complex tannins against biofilm-forming shiga toxin-producing Escherichia coli from Canada and South Africa. Antibiotics 2020, 9, 257. [Google Scholar] [CrossRef]
- Wang, C.; Hang, H.; Zhou, S.; Niu, Y.D.; Du, H.; Stanford, K.; McAllister, T.A. Bacteriophage biocontrol of shiga toxigenic Escherichia coli (STEC) O145 biofilms on stainless steel reduces the contamination of beef. Food Microbiol. 2020, 92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shigemura, K.; Duc, H.M.; Shen, C.; Huang, H.H.; Sato, J.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Effects of bacteriophage on inhibition and removal of mixed biofilm of enterohemorrhagic Escherichia coli O157: H7 and O91: H. LWT 2020, 134, 109945. [Google Scholar] [CrossRef]
- Montso, P.K.; Mlambo, V.; Ateba, C.N. Efficacy of novel phages for control of multi-drug resistant Escherichia coli O177 on artificially contaminated beef and their potential to disrupt biofilm formation. Food Microbiol. 2021, 94, 103647. [Google Scholar] [CrossRef]
- Curtin, J.J.; Donlan, R.M. Using bacteriophages to reduce formation of catheter-associated biofilms by Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2006, 50, 1268–1275. [Google Scholar] [CrossRef] [Green Version]
- Lacqua, A.; Wanner, O.; Colangelo, T.; Martinotti, M.G.; Landini, P. Emergence of biofilm-forming subpopulations upon exposure of Escherichia coli to environmental bacteriophages. Appl. Environ. Microbiol. 2006, 72, 956–959. [Google Scholar] [CrossRef] [Green Version]
- Gödeke, J.; Paul, K.; Lassak, J.; Thormann, K.M. Phage-induced lysis enhances biofilm formation in Shewanella oneidensis MR-1. ISME J. 2011, 5, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, S.; Sneppen, K.; Krishna, S. Coexistence of phage and bacteria on the boundary of self-organized refuges. Proc. Natl. Acad. Sci. USA 2012, 109, 12828–12833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jassim, S.A.A.; Abdulamir, A.S.; Bakar, F.A. Novel phage-based bio-processing of pathogenic Escherichia coli and its biofilms. World J. Microbiol. Biotechnol. 2012, 28, 47–60. [Google Scholar] [CrossRef]
- Kelly, D.; Mcauliffe, O.; Ross, R.P.; Coffey, A. Prevention of Staphylococcus aureus biofilm formation and reduction in established biofilm density using a combination of phage K and modified derivatives. Lett. Appl. Microbiol. 2012, 54, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Simões, L.C.; Vieira, M.J. A review of current and emergent biofilm control strategies. LWT Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Mangieri, N.; Picozzi, C.; Cocuzzi, R.; Foschino, R. Evaluation of a potential bacteriophage cocktail for the control of shiga-toxin producing Escherichia coli in food. Front. Microbiol. 2020, 11, 1801. [Google Scholar] [CrossRef]
- Carey-Smith, G.V.; Billington, C.; Cornelius, A.J.; Hudson, J.A.; Heinemann, J.A. Isolation and characterization of bacteriophages infecting Salmonella spp. FEMS Microbiol. Lett. 2006, 258, 182–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 28 September 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Serogroup | TSB 30 °C | TSB 37 °C | M9 30 °C | M9 37 °C |
|---|---|---|---|---|---|
| ATCC35150 a | O157 | 1.59 | 1.12 | 2.09 | 1.10 |
| 393 b | O26 | 2.37 | 2.25 | 1.26 | 1.06 |
| 15R b | O76 | 1.97 | 1.10 | 1.30 | 1.26 |
| 33C b | O23 | 3.91 | 3.95 | 1.32 | 1.44 |
| 380USA b | O157 | 2.58 | 3.65 | 1.36 | 0.96 |
| 62 19/L b | O157 | 2.19 | 2.38 | 3.02 | 1.23 |
| 214CH c | O157 | 1.62 | 1.26 | 1.16 | 0.94 |
| 228GS c | O145 | 2.90 | 1.38 | 1.22 | 1.08 |
| 229RACH c | O111 | 1.12 | 1.22 | 1.32 | 1.25 |
| 239RA c | O26 | 1.48 | 1.06 | 1.27 | 1.06 |
| 243RACH c | O26 | 1.68 | 1.01 | 1.36 | 1.08 |
| 243ROI-A c | O26 | 2.60 | 1.70 | 1.56 | 1.14 |
| F1-1 c | O26 | 1.37 | 0.93 | 1.43 | 1.09 |
| F10-4 c | O26 | 1.96 | 1.29 | 1.51 | 1.63 |
| F11-4 c | O26 | 2.17 | 1.20 | 1.64 | 1.07 |
| F95 c | O26 | 6.77 | 4.83 | 9.87 | 7.15 |
| F95-3 c | O26 | 1.44 | 1.37 | 1.41 | 1.10 |
| PO128 c | O128 | 1.36 | 1.02 | 1.25 | 1.32 |
| 6182-50 d | O113 | 2.40 | 2.31 | 2.50 | 1.62 |
| C679-12 e | O104 | 1.80 | 6.06 | 1.29 | 0.87 |
| ED13 f | O157 | 1.14 | 0.92 | 1.49 | 0.81 |
| ED142 f | O111 | 1,71 | 2.45 | 1.70 | 1.16 |
| ED161 f | O86 | 1.98 | 2.14 | 1.29 | 1.20 |
| ED172 f | O103 | 3.83 | 1.87 | 1.62 | 1.29 |
| ED173 f | O145 | 2.90 | 1.98 | 1.33 | 1.33 |
| ED226 f | O113 | 4.30 | 3.60 | 1.44 | 1.68 |
| ED33 f | O139 | 2.62 | 1.54 | 1.95 | 1.66 |
| ED56 f | O26 | 2.37 | 2.28 | 1.28 | 1.16 |
| ED76 f | O91 | 4.51 | 3.29 | 1.21 | 1.22 |
| ED82 f | O111 | 1.27 | 0.77 | 1.29 | 0.95 |
| ED238 f | O121 | 1.96 | 3.44 | 1.24 | 1.13 |
| Phage/Cocktail | MOI 1 | MOI 2 | MOI 10 | MOI 100 | Average |
|---|---|---|---|---|---|
| LF2 | 0.187 | 0.226 | 0.181 | 0.163 | 0.189 ± 0.03 |
| FM9 | 0.179 | 0.215 | 0.179 | 0.154 | 0.182 ± 0.03 |
| FM10 | 0.159 | 0.206 | 0.164 | 0.148 | 0.169 ± 0.03 |
| DP16 | 0.159 | 0.205 | 0.193 | 0.147 | 0.176 ± 0.03 |
| DP17 | 0.167 | 0.189 | 0.164 | 0.153 | 0.168 ± 0.02 |
| DP19 | 0.166 | 0.214 | 0.179 | 0.154 | 0.178 ± 0.03 |
| Cocktail 1 | 0.150 | 0.215 | 0.135 | 0.145 | 0.161 ± 0.04 |
| Cocktail 2 | 0.159 | 0.180 | 0.142 | 0.147 | 0.157 ± 0.02 |
| Average | 0.166 cd | 0.206 b | 0.167 c | 0.151d | * 0.304 a |
| ±0.012 | ±0.015 | ±0.02 | ±0.06 | ±0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangieri, N.; Foschino, R.; Picozzi, C. Application of Bacteriophages on Shiga Toxin-Producing Escherichia coli (STEC) Biofilm. Antibiotics 2021, 10, 1423. https://doi.org/10.3390/antibiotics10111423
Mangieri N, Foschino R, Picozzi C. Application of Bacteriophages on Shiga Toxin-Producing Escherichia coli (STEC) Biofilm. Antibiotics. 2021; 10(11):1423. https://doi.org/10.3390/antibiotics10111423
Chicago/Turabian StyleMangieri, Nicola, Roberto Foschino, and Claudia Picozzi. 2021. "Application of Bacteriophages on Shiga Toxin-Producing Escherichia coli (STEC) Biofilm" Antibiotics 10, no. 11: 1423. https://doi.org/10.3390/antibiotics10111423
APA StyleMangieri, N., Foschino, R., & Picozzi, C. (2021). Application of Bacteriophages on Shiga Toxin-Producing Escherichia coli (STEC) Biofilm. Antibiotics, 10(11), 1423. https://doi.org/10.3390/antibiotics10111423