Abstract
There are real concerns that Neisseria gonorrhoeae may become untreatable in the near future due to the rapid emergence of antimicrobial resistance. Alternative therapies are thus urgently required. Bacteriophages active against N. gonorrhoeae could play an important role as an antibiotic-sparing therapy. To the best of our knowledge, no bacteriophages active against N. gonorrhoeae have ever been found. The aim of this study was to screen for bacteriophages able to lyse N. gonorrhoeae in oropharyngeal and anorectal swabs of 74 men who have sex with men attending a sexual health clinic in Antwerp, Belgium. We screened 210 swabs but were unable to identify an anti-gonococcal bacteriophage. This is the first report of a pilot screening that systematically searched for anti-gonococcal phages directly from clinical swabs. Further studies may consider screening for phages at other anatomical sites (e.g., stool samples, urine) or in environmental settings (e.g., toilet sewage water of sex clubs or sexually transmitted infection clinics) where N. gonorrhoeae can be found.
1. Introduction
Bacteriophages (or phages) are viruses that interact with and invade bacterial cells resulting in the lysis of the infected host (virulent phage) or the integration of the phage genome in the chromosome of the host cell (temperate phage). With an estimated 1031–1032 phages in total, they are the most abundant living entities on Earth and play a crucial role in the microbial balance of the planet [1]. In recent years there has been renewed interest in the use of phages to prevent and contain the emergence of antibiotic resistance [2,3,4,5].
Neisseria gonorrhoeae, the bacterial pathogen causing the sexually transmittable infection gonorrhea, has developed resistance to all previously and currently recommended antimicrobials. As such, there are valid concerns it may become untreatable with conventional antimicrobials [6,7]. Antimicrobial resistance in N. gonorrhoeae is driven by excessive antimicrobial exposure [6,8]. Phages active against N. gonorrhoeae could play an important role in preventing the emergence of untreatable gonorrhea in two ways. Firstly, as an antibiotic-sparing therapy, which could reduce the selection pressure for the emergence of resistance against conventional antimicrobials. Secondly, they could be used in combination with antimicrobial agents to treat multi-resistant N. gonorrhoeae [9,10]. Phages have a number of advantages over antimicrobial therapy. They can be extremely species- and strain-specific and could be chosen not to have a major impact on the commensal flora [11]. In particular, they will not select for antimicrobial resistance (AMR) in commensal bacteria. This property might especially be beneficial for N. gonorrhoeae since anti-gonococcal antimicrobial therapy selects for AMR in commensal Neisseria, and these resistance determinants can be taken up by the pathogenic N. gonorrhoeae via horizontal gene transfer [12,13,14,15].
The first phages reported against Neisseria were found to be active against the nonpathogenic species N. perflava [16,17,18]. Phages active against N. meningitidis were first detected as early as 1967 [19], and more recently, Aljarbou et al. reported the isolation of lytic phages targeting N. meningitidis from human tooth plaques [20,21]. Remarkably, these anti-Neisseria phages showed extremely limited host ranges, being mostly reactive against a few strains within one species [16,17,18,20].
To the best of our knowledge, no phages active against N. gonorrhoeae have ever been found. In 1985, Campbell et al. observed plaques on lawns of two different strains of N. gonorrhoeae, but no phage particles active against this pathogenic species were finally isolated and identified [22]. Bioinformatic analyses of N. gonorrhoeae strain FA1090 have revealed the presence of four integrated filamentous phage genomes (Ngoϕ6–9) [23]. A phagemid derived from N. gonorrhoeae filamentous phage Ngoϕ6 showed an ability to infect and effectively produce progeny phagemids in a broad range of Gram-negative bacteria including Escherichia coli, Haemophilus influenzae, Neisseria sicca, and Pseudomonas species [24]. In addition, five prophage encoding regions (Ngoϕ1–5) were identified in strain FA1090.
Two of them (Ngoϕ1–2) may be functional, as they appear to encode all genes necessary for lytic growth, and prophage DNA sequences were detected in culture supernatants and visualized by electron microscopy [25]. However, they were unable to propagate and produce plaques on the N. gonorrhoeae and nonpathogenic Neisseria strains tested, suggesting that each strain is expressing the appropriate lysogenic control genes.
In conclusion, there is scarce evidence that these prophages are competent to produce phage particles, and we could find no evidence of any lytic phages that could be used to develop phage therapy directed towards N. gonorrhoeae. Therefore, the aim of the present study was to screen oropharyngeal and anorectal samples for the presence of phages able to infect N. gonorrhoeae.
2. Results
2.1. Isolation and Propagation of Potential Phages
A total of 194 oropharyngeal and 18 anorectal ESwabs™ from 74 MSM were screened for the presence of phages able to infect N. gonorrhoeae. Multiple clear zones were detected, ranging from confluent to opaque lysis (Figure 1).
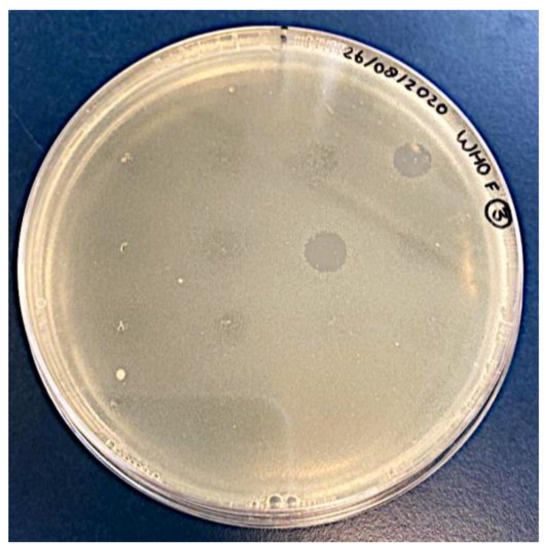
Figure 1.
Typical spot test result: semi-confluent lysis zones obtained on N. gonorrhoeae reference strain WHO-F.
None of the confluent lysis zones could be replicated through further propagation on the appropriate indicator strain. Clear zones pointed towards one pool of ESwabs™ enriched with different mixes of clinical N. gonorrhoeae strains and tested at the two different timepoints. Original individual ESwab™ filtrates comprising that pool were tested using the spot assay too. Several of these ESwabs™ inhibited the growth of some N. gonorrhoeae indicator strains, with one swab showing lysis zones on all indicator strains. None of these confluent lysis zones could be replicated through further propagation on the appropriate indicator strain.
Attempts were made to optimize the propagation methods by (i) prolonging the incubation time of the cut-out lysis zones, (ii) enriching the cut-out lysis zones with the appropriate indicator strain, and (iii) adding chloroform to assist the bacterial lysis process after incubation of the cut-out lysis zones. None of these attempts resulted in the replication of lysis zones. Enriched samples from the pool of ESwabs™ that showed clear zones were spotted on five other bacterial species (Pseudomonas aeruginosa, Staphylococcus aureus, Escherichia coli, Acinetobacter baumannii, and Enterococcus faecalis) but did not show any lytic activity.
2.2. Control Experiments
No lysis zones were observed after spotting filtrates of bacterial cultures. Pure ESwab™ medium faintly reduced growth on a limited number of bacterial overlays; however, it was not comparable with lysis zones of enriched ESwab™ samples. After centrifugation, filtration, and freezing at −80 °C up to one year period, E. coli phage GEC-3S infectivity decreased only at a minimal expected concentration of 2 log PFU/mL; in particular, from 2.14 log PFU/mL it dropped to 1.76 log PFU/mL after 3 days and to 0.94 log PFU/mL after 357 days of storage.
3. Discussion
We undertook an extensive search for lytic phages in oropharyngeal and rectal samples from individuals with a high incidence of N. gonorrhoeae by using a variety of methodologies. To the best of our knowledge, this is the first study that has systematically searched for anti-gonococcal phages directly from clinical swabs. Previous studies have detected phages active against a range of bacterial species from the oral cavity [26,27,28]. A number of studies have found phages infecting specific Neisseria species (but not N. gonorrhoeae) from dental plaque and oro- and naso-pharyngeal samples [17,18,19,20].
In this study, antibacterial activity that resulted in the formation of zones of lysis of N. gonorrhoeae was commonly encountered but could never be replicated through further propagation. Several studies have reported similar issues. Lysis zones on N. gonorrhoeae cultures were detected when suspensions of dental plaque were plated, but none of them could be propagated, and further transmission electron microscopy failed to reveal the presence of particles resembling phages [29]. A phage infecting a specific N. perflava strain showed lysis when throat isolates were spotted on a N. gonorrhoeae strain, but plaques were never seen. Ultraviolet treatment did not reduce lytic activity, suggesting lysis was not caused by phage particles but rather by an unknown component in the bacterial lysates [18].
False positive results due to bacterial killing by media components is a well-known limitation of the spot test [11]. However, we did not observe similar lysis zones during the negative control experiments in which we used bacterial culture filtrates and pure ESwab™ medium. This observation suggests that something is released from the samples that is causing the lysis. Several microbial interactions could play a role. It was recently reported that N. gonorrhoeae is killed when it takes up DNA with a differential methylation pattern released by various commensal Neisseria species such as N. elongata [30]. In addition, N. mucosa has been reported to secrete secondary metabolites that exhibit antigonococcal activity unrelated to the previously described DNA-mediated killing, although the antimicrobial metabolites have not yet been characterized [31]. Since these commensals are prevalent in the oropharynx in a high proportion of individuals, there is a commensurately high probability that this inhibitory DNA was present in the clinical swabs we used [15,32].
Bacteriocins produced by other bacterial species are another possible source of N. gonorrhoeae lysis [33]. A bacteriocin (Viridin B) produced by Streptococcus mitis, inhabiting the human mouth, is known to be bactericidal to N. sicca [34], and caerin 1 antimicrobial peptides have been shown to inhibit N. lactamica [35]. In addition, a randomized controlled trial showed that inoculation with N. lactamica inhibits the carriage of N. meningitidis in young adults [36]. Although the exact mechanism is unknown, bacteriocins, uptake of toxic DNA, and microbial competition could be responsible for this non-phage lysis.
To test the plausibility of this hypothesis, we subjected some strains of all different commensal Neisseria species obtained from the pharyngeal samples used as a potential phage source to an adapted spot test [37]. In addition, we included ATCC strains of other bacterial species that could have been present in the oropharyngeal and rectal samples tested, including Staphylococcus aureus, Streptococcus pneumoniae, and E. coli. None of the Neisseria species tested (N. meningitidis, N. subflava, N. cinerea, N. lactamica, N. oralis, and N. elongata) showed any effect. However, S. pneumoniae, and to a lesser extent E. coli, produced significant inhibition zones on all N. gonorrhoeae target strains tested.
Other factors in addition to bacterial products may be causing the N. gonorrhoeae inhibition during our phage screening experiments. In a study that detected a novel phage targeting N. subflava, single plaques derived from dental plaque swabs were used to re-infect the same host-culture; only the first and second round of propagation resulted in plaques [21]. The exact mechanism of losing infectivity or developing host resistance is unclear but is a common observation [22].
Previously reported autoplaquing in N. gonorrhoeae [22] could be explained by autolysis or the presence of two different functionally active prophages [25,38]. However, we did not observe any lysis zones during the negative control experiments in which we used bacterial culture lysates in GBC+.
A limitation of our study is that we mainly screened oropharyngeal samples, a limited number of anorectal swabs, and no urethral, urine, dental plaque, female genital tract, or environmental samples (e.g., toilet wastewater). While commensal Neisseria species colonize the oral and nasopharyngeal cavities, the median duration of oropharyngeal N. gonorrhoeae infection among MSM has been reported to persist between 28 days and up to 16 weeks when untreated [39,40]. Future studies could aim at isolating phages infecting non-pathogenic Neisseria species and then trying to train and adapt its host range to ultimately be capable of infecting N. gonorrhoeae. Furthermore, some samples were taken after the use of a daily Listerine mouthwash (54 out of 210 samples) or an antibiotic (72 out of 210 samples) in the three months prior to sampling. These may have altered the microbiota in a way that decreased the probability of detecting relevant phages.
4. Materials and Methods
4.1. Bacterial Strains
Three mixes, each consisting of three or four clinical N. gonorrhoeae strains collected at the Institute of Tropical Medicine in Antwerp, Belgium, between 2019 and 2020, were used for phage enrichment. In addition to the 11 enrichment strains, 9 WHO N. gonorrhoeae reference strains were included for phage isolation [41,42]. We refer to this collection as our ‘indicator strains’; for further details see Supplementary Table S1.
4.2. Potential Phage Containing Samples
Oropharyngeal and anorectal ESwabs™ (COPAN Diagnostics Inc., Brescia, Italy) were collected from men who have sex with men (MSM) as part of two clinical studies conducted at the Institute of Tropical Medicine (ITM) in Antwerp, Belgium.
The first study, Preventing Resistance in Gonorrhoea (PReGO), was a placebo-controlled crossover randomized clinical trial assessing whether the regular use of an essential oil-based mouthwash could reduce the incidence of sexually transmitted infections in 343 MSM taking HIV preexposure prophylaxis at ITM [43] (registered at clinicaltrials.gov as NCT03881007). The second study, the Resistogenicity study, was an observational study that evaluated the effect of ceftriaxone and azithromycin on the antimicrobial susceptibilities of commensal Neisseria in 10 MSM attending ITM [32]. For further details see Table 1.

Table 1.
Oropharyngeal and anorectal ESwabs™ collected from men who have sex with men from two clinical studies at ITM Antwerp, Belgium, used for N. gonorrhoeae phage screening.
All oropharyngeal samples were taken by rubbing both tonsillar pillars and the posterior oropharyngeal wall by study physicians. Anorectal swabs were generally taken by the study participants themselves. After collection, the ESwabs™ were inoculated on Colombia Agar with 5% Sheep Blood and GC-LectTM agar plates (Becton Dickinson) to isolate Neisseria species as part of the two above-mentioned clinical studies. The following Neisseria species were present in samples from the PReGO study: N. sublfava/flavescens, N. macacae/mucosa, N. meningitidis, and N. gonorrhoeae (unpublished data).
In addition to the above species, N. oralis was found in samples from the Resistogenicity study [32]. The remaining ESwab™ transport media were stored at −80 °C after transfer into cryotubes. Frozen ESwab™ transport media were thawed and centrifuged at 5000× g for 15 min in 0.22 µm filter tubes (Costar™, Spin-X™ Centrifuge Tube Filters CA Membrane) to filter out bacteria, cells, and debris present in the sample. The supernatant was stored at 4 °C and used, within 24 h, for phage enrichment.
4.3. Phage Enrichment Culture
After filtration, 450 µL media of each ESwab™ was used to generate pools of approximately 20 samples. Each pool was divided into 3 parts (circa 150 µL media from each ESwab™). Each fraction was incubated in 10 mL gonococcal broth (double-distilled water (ddH2O) supplemented with 15 g/L Bacto proteose peptone no. 3, 1 g/L soluble starch, 4 g/L K2HPO4, 1 g/L KH2PO4, 5 g/L NaCl, and 10 mL/L IsoVitaleX; further referred to as “GCB+”) and with a different mix of enrichment strains at 37 °C in 5% CO2 (using 10 µL of 107 CFU/mL suspension of each strain) (Table 1). The turbidity of the cultures was measured, and a 1 mL sample was taken in the mid-log phase (after 6–7 h). Six milliliters of fresh GCB+ was added to supply sufficient nutrition for overnight incubation, whereafter a second sample was taken (18–21 h). Samples were centrifuged at 5000× g for 15 min in 0.22 µm filter tubes (Costar™, Spin-X™ Centrifuge Tube Filters CA Membrane) to remove bacteria, and samples were stored at 4 °C until spot testing was carried out.
4.4. Spot Test
The spot test method was used as an initial indicator test to screen enriched samples for the presence of phages by measuring lytic activity. An amount of 3 µL of GCB+ containing 0.6% agar and 100 µL of a 108 CFU/mL N. gonorrhoeae indicator strain culture was poured onto a gonococcal base (BD DifcoTM) supplemented with 1% IsoVitalex. The agar mixture was swirled to produce a uniform top layer. Once solidified, 10 µL of potential phage suspensions, obtained after enrichment, centrifugation, and filtering, was spotted onto the overlay. Plates were incubated at 37 °C in 5% CO2 for 24 to 48 h before inspection of cleared zones.
4.5. Phage Isolation and Propagation
Confluent lysis zones were cut with a sterile inoculating loop and inoculated into 4 mL GCB+ in 14 mL sterile tubes and incubated at 37 °C in 5% CO2 for 2–3 h. Subsequently, 500 µL of the sample was centrifuged at 5000× g for 15 min in 0.22 µm filter tubes (Costar™, Spin-X™ Centrifuge Tube Filters CA Membrane) to remove bacteria and debris and stored at 4 °C until a new spot test was carried out.
When another confluent lysis zone was obtained, a double agar overlay assay was carried out to determine the concentration of infectious phage particles.
4.6. Control Experiments
To exclude the interference of spontaneous induction of prophages from the bacterial hosts, bacterial strains used for enrichment were cultured in GCB+ for 24 h at 37 °C in 5% CO2. Obtained cultures were either centrifuged at 5000× g for 15 min in 0.22 µL m filter tubes (Costar™, Spin-X™ Centrifuge Tube Filters CA Membrane) or treated with chloroform to lyse bacteria and tested for lysis zones using the spot test. In addition, pure ESwab™ medium was directly tested for lysis using the spot test to exclude bacterial killing by media components.
E. coli phage GEC-3S (The Eliava IBMV, Tbilisi, Georgia) [44] was used to test the efficiency of our phage isolation procedure and to assess phage stability in ESwab™ medium.
Hundred-fold dilutions of phage GEC-3S with initial titer of 10.1 log PFU/mL were made in ESwab™ medium, centrifuged at 5000× g for 15 min through a 0.22 µm filter, and stored at −80 °C for up to a one-year period. After storage, phage-ESwab™ samples were thawed and titered using the double-agar overlay assay. Minimal concentration of phage GEC-3S tested was 2 log PFU/mL. In brief, 1 mL of phage dilutions in ESwab™ medium was added to 100 µL of E. coli strain DSM 613 grown overnight on Blood Agar at 37 °C. The phage–bacteria mixture was vortexed briefly and added to 4 mL preheated GCB+ top agar (0.6% agar) and overlaid on GCB agar plates. Plates were overnight incubated at 37 °C until plaques could be detected and counted.
4.7. Ethics Approval
Ethics approval for this study was obtained from ITM’s Institutional Review Board (1276/18) and the Ethics Committee of the University of Antwerp (19/06/058).
5. Conclusions
This is the first report of a pilot screening of clinical samples for the presence of N. gonorrhoeae phages. A strictly virulent N. gonorrhoeae phage would be of considerable utility in the fight against extensively resistant N. gonorrhoeae infections.
Further studies may consider screening for phages at other anatomical sites (e.g., stool samples, urine) or in environmental settings (e.g., toilet sewage water of sex clubs or sexually transmitted infection clinics) where N. gonorrhoeae can be found. It may also be possible to engineer a related phage to target N. gonorrhoeae [45,46]. The phages that target N. meningitidis would be a logical place to start [19,21].
Supplementary Materials
The following are available online at www.mdpi.com/article/10.3390/antibiotics11020268/s1, Table S1: N. gonorrhoeae strains used for phage enrichment and isolation.
Author Contributions
Conceptualization, J.G.E.L., J.-P.P., M.M. and C.K.; methodology, J.G.E.L., J.-P.P. and M.M.; validation, J.G.E.L., S.A. and M.M.; formal analysis, J.G.E.L., S.A. and M.M.; investigation, J.G.E.L., S.A. and M.M.; writing—original draft preparation, J.G.E.L. and C.K.; writing—review and editing, J.G.E.L., S.A., S.S.M.-B., C.V.D., D.V.d.B., I.D.B., T.d.B., S.M.-K., P.S., J.-P.P., C.K. and M.M.; supervision, M.M., J.-P.P. and C.K.; project administration, D.V.d.B. and I.D.B.; funding acquisition, C.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and the study was approved by the Ethics Committee of Antwerp University Hospital and University of Antwerp (protocol code 19/06/058, 25 February 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kutter, E.; Sulakvelidze, A. Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2004; ISBN 0203491750. [Google Scholar]
- Chanishvili, N.; Chanishvili, T.; Tediashvili, M.; Barrow, P.A. Phages and their application against drug-resistant bacteria. J. Chem. Technol. Biotechnol. 2001, 76, 689–699. [Google Scholar] [CrossRef]
- Vos, D.D.; Pirnay, J.-P. Phage therapy: Could viruses help resolve the worldwide antibiotic crisis? Altern. Antibiot. 2015, 110–114. [Google Scholar]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Rashel, M.; Uchiyama, J.; Sakurai, S.; Ujihara, T.; Kuroda, M.; Ikeuchi, M.; Tani, T.; Fujieda, M.; Wakiguchi, H.; et al. Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11, 211–219. [Google Scholar] [CrossRef]
- Unemo, M.; Shafer, W.M. Antimicrobial resistance in Neisseria gonorrhoeae in the 21st Century: Past, evolution, and future. Clin. Microbiol. Rev. 2014, 27, 587–613. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xue, J.; Chen, Y.; Chen, S.; Wang, Q.; Zhang, C.; Wu, S.; Lv, H.; Yu, Y.; Van Der Veen, S. Increasing prevalence of Neisseria gonorrhoeae with decreased susceptibility to ceftriaxone and resistance to azithromycin in Hangzhou, China (2015–2017). J. Antimicrob. Chemother. 2019, 74, 29–37. [Google Scholar] [CrossRef]
- Kenyon, C.; Buyze, J.; Spiteri, G.; Cole, M.J.; Unemo, M. Population-Level Antimicrobial Consumption Is Associated with Decreased Antimicrobial Susceptibility in Neisseria gonorrhoeae in 24 European Countries: An Ecological Analysis. J. Infect. Dis. 2020, 221, 1107–1116. [Google Scholar] [CrossRef]
- Gu Liu, C.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-Antibiotic Synergy is Driven by a Unique Combination of Antibacterial Mechanism of Action and Stoichiometry. MBio 2020, 11, e01462-20. [Google Scholar] [CrossRef]
- Morrisette, T.; Lev, K.L.; Kebriaei, R.; Abdul-Mutakabbir, J.C.; Stamper, K.C.; Morales, S.; Lehman, S.M.; Canfield, G.S.; Duerkop, B.A.; Arias, C.A. Bacteriophage-antibiotic combinations for Enterococcus faecium with varying bacteriophage and daptomycin susceptibilities. Antimicrob. Agents Chemother. 2020, 64, e00993-20. [Google Scholar] [CrossRef]
- Hyman, P. Phages for phage therapy: Isolation, characterization, and host range breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, C.B.; Arnold, B.J.; Sater, M.R.A.; Grad, Y.H. Azithromycin resistance through interspecific acquisition of an epistasis-dependent efflux pump component and transcriptional regulator in Neisseria gonorrhoeae. MBio 2018, 9, e01419-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marri, P.R.; Paniscus, M.; Weyand, N.J.; Rendón, M.A.; Calton, C.M.; Hernández, D.R.; Higashi, D.L.; Sodergren, E.; Weinstock, G.M.; Rounsley, S.D.; et al. Genome sequencing reveals widespread virulence gene exchange among human Neisseria species. PLoS ONE 2010, 5, e11835. [Google Scholar] [CrossRef] [PubMed]
- Spratt, B.G.; Bowler, L.D.; Zhang, Q.Y.; Zhou, J.; Smith, J.M. Role of interspecies transfer of chromosomal genes in the evolution of penicillin resistance in pathogenic and commensal Neisseria species. J. Mol. Evol. 1992, 34, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.V.; Pham, L.Q.; Nguyen, H.T.; Nguyen, M.X.B.; Nguyen, T.V.; May, F.; Le, G.M.; Klausner, J.D. Decreased Cephalosporin Susceptibility of Oropharyngeal Neisseria Species in Antibiotic-using Men Who Have Sex with Men in Hanoi, Vietnam. Clin. Infect. Dis. 2020, 70, 1169–1175. [Google Scholar] [CrossRef]
- Stone, R.L.; Culbertson, C.G.; Powell, H.M. Studies of a bacteriophage active against a chromogenic Neisseria. J. Bacteriol. 1956, 71, 516–520. [Google Scholar] [CrossRef] [Green Version]
- Phelps, L.N. Isolation and Characterization of Bacteriophages for Neisseria. J. Gen. Virol. 1967, 1, 529–536. [Google Scholar] [CrossRef]
- Steinberg, V.I.; Hart, E.J.; Handley, J.; Goldberg, I.D. Isolation and characterization of a bacteriophage specific for Neisseria perflava. J. Clin. Microbiol. 1976, 4, 87–91. [Google Scholar] [CrossRef]
- Cary, S.G.; Hunter, D.H. Isolation of Bacteriophages Active against Neisseria meningitidis. J. Virol. 1967, 1, 538–542. [Google Scholar] [CrossRef] [Green Version]
- Aljarbou, A.N.; De Luca, A.A.M. Isolation of a New Neisseria Phage from the Oral Cavity of Healthy Humans. Open Access Sci. Reports 2012, 17, 18. [Google Scholar] [CrossRef]
- Aljarbou, A.N.; Aljofan, M. Genotyping, morphology and molecular characteristics of a lytic phage of Neisseria strain obtained from infected human dental plaque. J. Microbiol. 2014, 52, 609–618. [Google Scholar] [CrossRef]
- Campbell, L.A.; Short, H.B.; Young, F.E.; Clark, V.L. Autoplaquing in Neisseria gonorrhoeae. J. Bacteriol. 1985, 164, 461–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicja Niewiadomska, J.K.; Niewiadomska, A.; Klama, J. Analysis of the Filamentous Bacteriophage Genomes Integrated into Neisseria gonorrhoeae FA1090 Chromosome. Polish J. Microbiol. 2005, 54, 43–48. [Google Scholar]
- Piekarowicz, A.; Klyz, A.; Majchrzak, M.; Szczesna, E.; Piechucki, M.; Kwiatek, A.; Maugel, T.K.; Stein, D.C. Neisseria gonorrhoeae Filamentous Phage Ngo 6 Is Capable of Infecting a Variety of Gram-Negative Bacteria. J. Virol. 2014, 88, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piekarowicz, A.; Kłyz, A.; Majchrzak, M.; Adamczyk-Popławska, M.; Maugel, T.K.; Stein, D.C. Characterization of the dsDNA prophage sequences in the genome of Neisseria gonorrhoeae and visualization of productive bacteriophage. BMC Microbiol. 2007, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachrach, G.; Leizerovici-Zigmond, M.; Zlotkin, A.; Naor, R.; Steinberg, D. Bacteriophage isolation from human saliva. Lett. Appl. Microbiol. 2003, 36, 50–53. [Google Scholar] [CrossRef]
- Tylenda, C.A.; Calvert, C.; Kolenbrander, P.E.; Tylenda, A. Isolation of Actinomyces bacteriophage from human dental plaque. Infect. Immun. 1985, 49, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Machuca, P.; Daille, L.; Vinés, E.; Berrocal, L.; Bittner, M. Isolation of a novel bacteriophage specific for the periodontal pathogen Fusobacterium nucleatum. Appl. Environ. Microbiol. 2010, 76, 7243–7250. [Google Scholar] [CrossRef] [Green Version]
- Hitch, G.; Pratten, J.; Taylor, P.W. Isolation of bacteriophages from the oral cavity. Lett. Appl. Microbiol. 2004, 39, 215–219. [Google Scholar] [CrossRef]
- Kim, W.J.; Higashi, D.; Goytia, M.; Rendón, M.A.; Pilligua-Lucas, M.; Bronnimann, M.; McLean, J.A.; Duncan, J.; Trees, D.; Jerse, A.E.; et al. Commensal Neisseria Kill Neisseria gonorrhoeae through a DNA-Dependent Mechanism. Cell Host Microbe 2019, 26, 228–239. [Google Scholar] [CrossRef]
- Aho, E.L.; Ogle, J.M.; Finck, A.M. The Human Microbiome as a Focus of Antibiotic Discovery: Neisseria mucosa Displays Activity Against Neisseria gonorrhoeae. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Laumen, J.G.E.; Van Dijck, C.; Abdellati, S.; Manoharan-Basil, S.S.; Baetselier De, I.; Martiny, D.; Crucitti, T.; Kenyon, C. Markedly Reduced Azithromycin and Ceftriaxone Susceptibility in Commensal Neisseria Species in Clinical Samples from Belgian Men Who Have Sex with Men. Clin. Infect. Dis. 2020, 72, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Law, D.J.; Dajani, A.S. Interactions between Neisseria sicca and viridin B, a bacteriocin produced by Streptococcus mitis. Antimicrob. Agents Chemother. 1978, 13, 473–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollins-Smith, L.A.; Smith, P.B.; Ledeczi, A.M.; Rowe, J.M.; Reinert, L.K. Caerin 1 antimicrobial peptides that inhibit HIV and neisseria may spare protective lactobacilli. Antibiotics 2020, 9, 661. [Google Scholar] [CrossRef]
- Deasy, A.M.; Guccione, E.; Dale, A.P.; Andrews, N.; Evans, C.M.; Bennett, J.S.; Bratcher, H.B.; Maiden, M.C.J.; Gorringe, A.R.; Read, R.C. Nasal inoculation of the commensal neisseria lactamica inhibits carriage of neisseria meningitidis by young adults: A controlled human infection study. Clin. Infect. Dis. 2015, 60, 1512–1520. [Google Scholar] [CrossRef]
- Abdellati, S.; Laumen, J.; Gonzalez, N.; Basil, S.; Van Dijck, C.; De Block, T.; De Baetselier, I.; Martiny, D.; Kenyon, C. Circulating Isolates of Neisseria Mucosa do not Inhibit the Growth of Neisseria Gonorrhoeae. Preprints 2021, in press. [Google Scholar] [CrossRef]
- Breyen, S.A.; Dworkin, M. Autoplaquing in Myxococcus strains. J. Bacteriol. 1984, 158, 1163–1164. [Google Scholar] [CrossRef] [Green Version]
- Chow, E.P.F.; Vodstrcil, L.A.; Williamson, D.A.; Maddaford, K.; Hocking, J.S.; Ashcroft, M.; De Petra, V.; Bradshaw, C.S.; Fairley, C.K. Incidence and duration of incident oropharyngeal gonorrhoea and chlamydia infections among men who have sex with men: Prospective cohort study. Sex. Transm. Infect. 2020, 97, 452–457. [Google Scholar] [CrossRef]
- Barbee, L.A.; Soge, O.O.; Khosropour, C.M.; Haglund, M.; Yeung, W.; Hughes, J.; Golden, M.R. The Duration of Pharyngeal Gonorrhea: A Natural History Study. Clin. Infect. Dis. 2021, 73, 575–582. [Google Scholar] [CrossRef]
- Unemo, M.; Golparian, D.; Sánchez-Busó, L.; Grad, Y.; Jacobsson, S.; Ohnishi, M.; Lahra, M.M.; Limnios, A.; Sikora, A.E.; Wi, T.; et al. The novel 2016 WHO Neisseria gonorrhoeae reference strains for global quality assurance of laboratory investigations: Phenotypic, genetic and reference genome characterization. J. Antimicrob. Chemother. 2016, 71, 3096–3108. [Google Scholar] [CrossRef] [Green Version]
- Unemo, M.; Fasth, O.; Fredlund, H.; Limnios, A.; Tapsall, J. Phenotypic and genetic characterization of the 2008 WHO Neisseria gonorrhoeae reference strain panel intended for global quality assurance and quality control of gonococcal antimicrobial resistance surveillance for public health purposes. J. Antimicrob. Chemother. 2009, 63, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Van Dijck, C.; Tsoumanis, A.; Rotsaert, A.; Vuylsteke, B.; Van den Bossche, D.; Paeleman, E.; Florence, E.; De Baetselier, I.; Brosius, I.; Laumen, J.; et al. Does an antibacterial mouthwash prevent sexually transmitted infections in men who have sex with men taking HIV pre-exposure prophylaxis? A randomised, placebo controlled, crossover trial. Lancet Infect. Dis. 2021, 21, 657–667. [Google Scholar] [CrossRef]
- Merabishvili, M.; de Vos, D.; Verbeken, G.; Kropinski, A.M.; Vandenheuvel, D.; Lavigne, R.; Wattiau, P.; Mast, J.; Ragimbeau, C.; Mossong, J.; et al. Selection and Characterization of a Candidate Therapeutic Bacteriophage That Lyses the Escherichia coli O104:H4 Strain from the 2011 Outbreak in Germany. PLoS ONE 2012, 7, e52709. [Google Scholar] [CrossRef] [Green Version]
- Kilcher, S.; Loessner, M.J. Engineering Bacteriophages as Versatile Biologics. Trends Microbiol. 2019, 27, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic engineering of bacteriophages against infectious diseases. Front. Microbiol. 2019, 10, 954. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).