
Anti-Staphylococcal Activities of Rosmarinus officinalis and Myrtus communis Essential Oils through ROS-Mediated Oxidative Stress

 ,
,
Abstract
1. Introduction
2. Results
2.1. Chemical Characterization of EOs
2.2. Antibacterial Activity
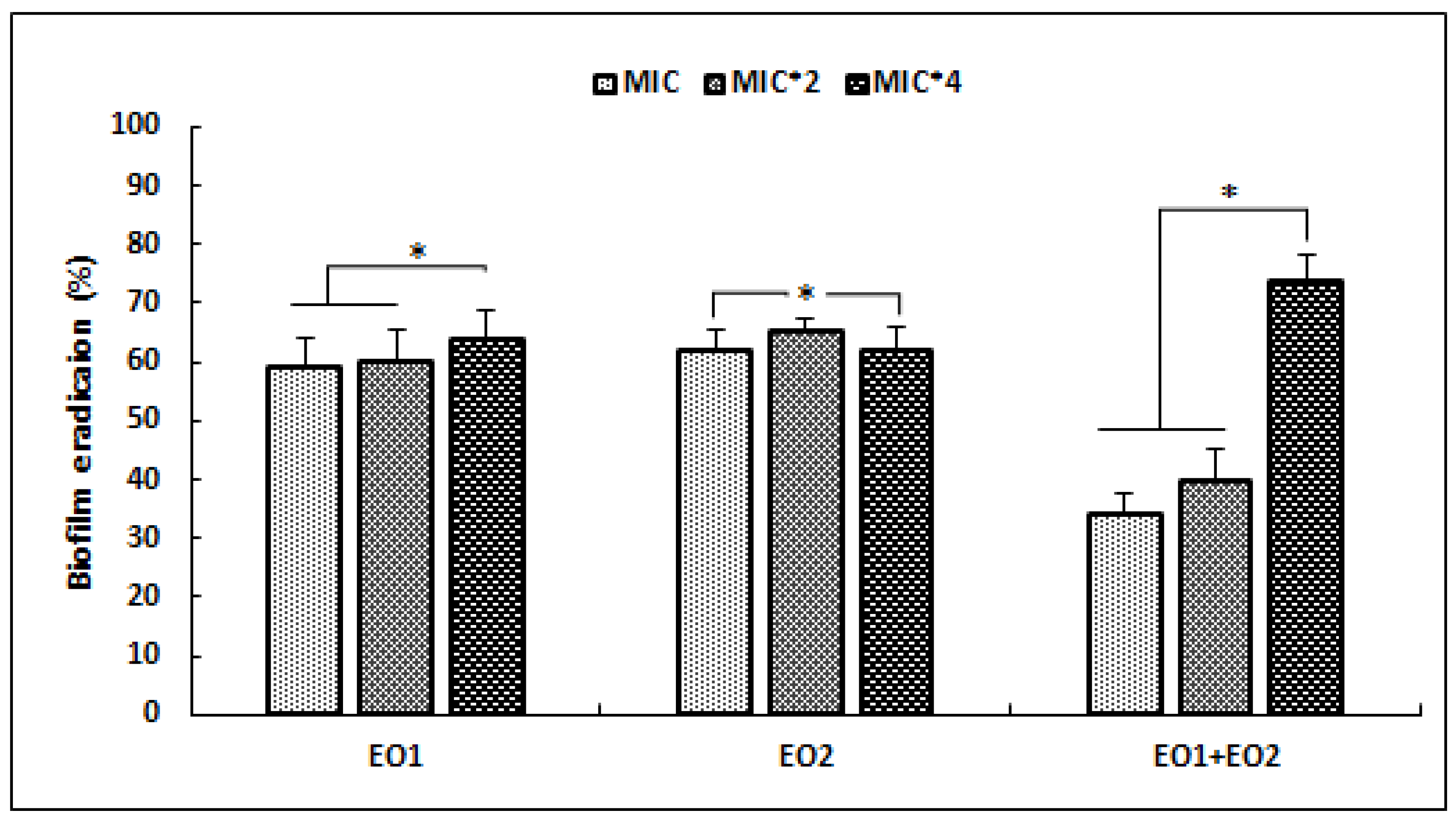
2.3. Antibiofilm Activity
2.4. Antivirulence Activities
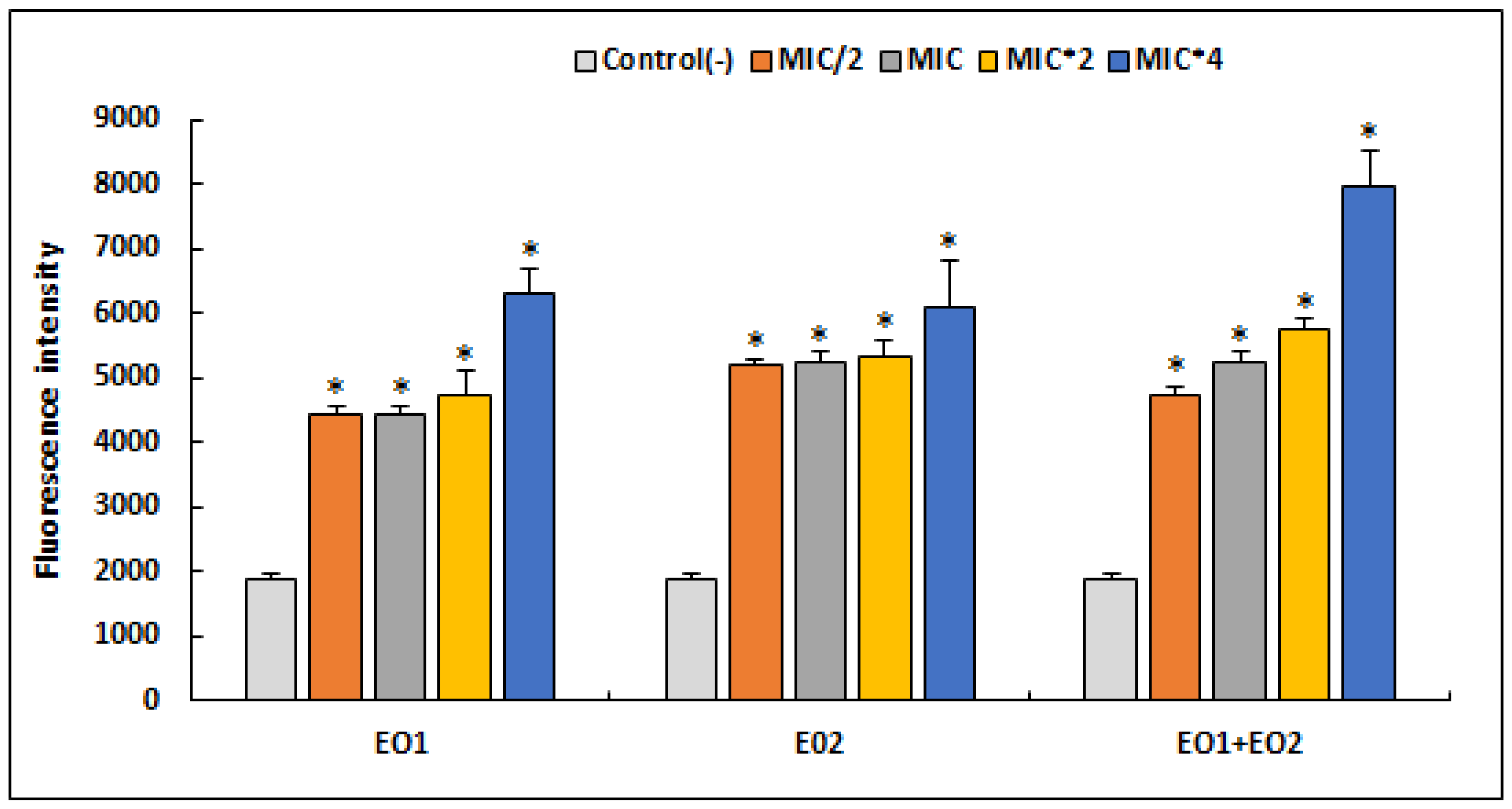
2.5. ROS Generation
2.6. Catalase Activity
3. Discussion
4. Materials and Methods
4.1. Tested Agents and Bacterial Strain
4.2. Chemical Characterization of Essential Oils
4.3. Antibacterial Activity of EOs and Their Combinations
4.4. Antibiofilm Activity
4.5. Anti-Hemolysin, Anti-DNase and Anti-Protease Activities
4.6. Reactive Oxygen Species (ROS) Generation
4.7. Antioxidant Enzyme Activity
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fridkin, S.K.; Hageman, J.C.; Morrison, M.; Sanza, L.T.; Como-Sabetti, K.; Jernigan, J.A.; Harriman, K.; Harrison, L.H.; Lynfield, R.; Farley, M.M. Active Bacterial Core Surveillance Program of the Emerging Infections Program Network. Methicillin-resistant Staphylococcus aureus disease in three communities. N. Engl. J. Med. 2005, 352, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated With Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011-2014. Infect. Control. Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef]
- Egert, M.; Späth, K.; Weik, K.; Kunzelmann, H.; Horn, C.; Kohl, M.; Blessing, F. Bacteria on smartphone touchscreens in a German university setting and evaluation of two popular cleaning methods using commercially available cleaning products. Folia Microbiol. 2015, 60, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Prévost, G. Toxins in Staphylococcus aureus pathogenesis. In Molecular and Cellular Biology, Horizon Bioscience; Proft, Ed.; Microbial Toxins: Norfolk, UK, 2005; pp. 243–284. [Google Scholar]
- Lentino, J.R. Prosthetic joint infections: Bane of orthopedists, challenge for infectious disease specialists. Clin. Infect. Dis. 2003, 36, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Akbari-Ayezloy, E.; Hosseini-Jazani, N.; Yousefi, S.; Habibi, N. Eradication of methicillin resistant S. aureus biofilm by the combined use of fosfomycin and β-chloro-L-alanine. Iran J. Microbiol. 2017, 9, 1–10. [Google Scholar]
- Cerqueira, G.M.; Peleg, A.Y. Insights into Acinetobacter baumannii pathogenicity. IUBMB Life 2011, 63, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Graber, C.J. Limitations of antibiotic options for invasive infections caused by methicillin-resistant Staphylococcus aureus: Is combination therapy the answer? J. Antimicrob. Chemother. 2010, 65, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Luo, J.; Deng, F.; Huang, Y.; Zhou, H. Antibiotic Combination Therapy: A Strategy to Overcome Bacterial Resistance to Aminoglycoside Antibiotics. Front. Pharmacol. 2022, 13, 839808. [Google Scholar] [CrossRef]
- Singh, N.; Yeh, P.J. Suppressive drug combinations and their potential to combat antibiotic resistance. J. Antibiot. 2017, 70, 1033–1042. [Google Scholar] [CrossRef]
- Abushaheen, M.A.; Muzaheed Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; Vellappally, S.; et al. Antimicrobial resistance, mechanisms and its clinical significance. Dis. Mon. 2020, 66, 100971. [Google Scholar] [CrossRef] [PubMed]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef]
- El Amri, J.; Elbadaoui, K.; Zair, T.; Bouharb, H.; Chakir, S.; Alaoui, T.I. Étude de l’activité antibactérienne des huiles essentielles de Teucrium capitatium L et l’extrait de Siléne vulgaris sur différentes souches testées. J. Appl. Biosci. 2014, 82, 7481. [Google Scholar] [CrossRef]
- Luis, Â.; Duarte, A.; Gominho, J.; Domingues, F.; Duarte, A.P. Chemical composition, antioxidant, antibacterial and anti-quorum sensing activities of Eucalyptus globulus and Eucalyptus radiata essential oils. Ind. Crops Prod. 2016, 79, 274–282. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Sarma, B.K.; Singh, H.B. Antioxidant and anti-quorum sensing activities of green pod of Acacia nilotica L. Food Chem. Toxicol. 2009, 47, 778–786. [Google Scholar] [CrossRef]
- Al-Jabri, N.N.; Hossain, M.A. Comparative chemical composition and antimicrobial activity study of essential oils from two imported lemon fruits samples against pathogenic bacteria. Beni-Suef Univ. J. Basic Appl. Sci. 2014, 3, 247–253. [Google Scholar]
- Achour, M.; Mateos, R.; Ben Fredj, M.; Mtiraoui, A.; Bravo, L.; Saguem, S. A comprehensive characterisation of rosemary tea obtained from Rosmarinus officinalis L. collected in a sub-humid Area of Tunisia: Compounds from Rosmarinus officinalis tea. Phytochem Anal. 2018, 29, 87–100. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Carvalho-Costa, D.; Cavaleiro, C.; Costa, H.S.; Albuquerque, T.G.; Castilho, M.C.; Ramos, F.; Melo, N.R.; Sanches-Silva, A. A novel insight on an ancient aromatic plant: The rosemary (Rosmarinus officinalis L.). Trends Food Sci. Technol. 2015, 45, 355–368. [Google Scholar] [CrossRef]
- Sasikumar, B. Handbook of Herbs and Spices, 2nd ed.; Woodhead publishing series in food science, technology and nutrition; Woodhead Publishing: Cambridge, UK, 2012; Volume 1, pp. 452–468. [Google Scholar]
- Hennia, A.; Nemmiche, S.; Dandlen, S.; Miguel, M.G. Myrtus communis essential oils: Insecticidal, antioxidant and antimicrobial activities: A review. J. Essent. Oil Res. 2019, 31, 487–545. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Ochoa, S.; Chacón-Vargas, K.F.; Sánchez-Torres, L.E.; Rivera-Chavira, B.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure. Membranes 2021, 11, 405. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, S.H.; Lai, K.S. Lavender essential oil induces oxidative stress which modifies the bacterial membrane permeability of carbapenemase producing Klebsiella pneumoniae. Sci. Rep. 2020, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Long, Y.; Li, D.; Shi, A.; Deng, J.; Ma, Y.; Wen, J.; Li, X.; Zhang, Y.; Liu, S.; et al. Natural essential oils efficacious in internal organs fibrosis treatment: Mechanisms of action and application perspectives. Pharmacol. Res. 2022, 182, 106339. [Google Scholar] [CrossRef]
- Badreddine, B.S.; Olfa, E.; Samir, D.; Hnia, C.; Lahbib, B.J. Chemical composition of Rosmarinus and Lavandula essential oils and their insecticidal effects on Orgyia trigotephras (Lepidoptera, Lymantriidae). Asian Pac. J. Trop. Med. 2015, 8, 98–103. [Google Scholar] [CrossRef]
- Moumni, S.; Elaissi, A.; Trabelsi, A.; Merghni, A.; Chraief, I.; Jelassi, B.; Chemli, R.; Ferchichi, S. Correlation between chemical composition and antibacterial activity of some Lamiaceae species essential oils from Tunisia. BMC Complement Med. Ther. 2020, 20, 103. [Google Scholar] [CrossRef]
- Bouzabata, A. Traditional Treatment of high blood pressure and diabetes in Souk Ahras District. J. Pharmacogn. Phytother. 2013, 5, 12–20. [Google Scholar]
- Cherrat, L.; Espina, L.; Bakkali, M.; García-Gonzalo, D.; Pagán, R.; Laglaoui, A. Chemical composition and antioxidant properties of Laurus nobilis L. and Myrtus communis L. essential oils from Morocco and evaluation of their antimicrobial activity acting alone or in combined processes for food preservation. J. Sci. Food Agric. 2014, 94, 1197–1204. [Google Scholar] [CrossRef]
- Merghni, A.; Marzouki, H.; Hentati, H.; Aouni, M.; Mastouri, M. Antibacterial and antibiofilm activities of Laurus nobilis L. essential oil against Staphylococcus aureus strains associated with oral infections. Pathol. Biol. 2015. Online ahead of print. [Google Scholar]
- Ait-Ouazzou, A.; Lorán, S.; Bakkali, M.; Laglaoui, A.; Rota, C.; Herrera, A.; Pagán, R.; Conchello, P. Chemical composition and antimicrobial activity of essential oils of Thymus algeriensis, Eucalyptus globulus and Rosmarinus officinalis from Morocco. J. Sci. Food Agric. 2011, 91, 2643–2651. [Google Scholar] [CrossRef]
- Djenane, D.; Yangüela, J.; Amrouche, T.; Boubrit, S.; Boussad, N.; Roncalés, P. Chemical composition and antimicrobial effects of essential oils of Eucalyptus globulus, Myrtus communis and Satureja hortensis against Escherichia coli O157, H7 and Staphylococcus aureus in minced beef. Food Sci. Technol. Int. 2011, 17, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Silou, T.; Nsikabaka, S.; Nombault Nienzy, J.P.; Baou, A.; Biassala, T.E.; Figuérédo, G.; Chalchat, J.C. Aromatic Plants from Plateau des Cataractes: Occurrence of the Citronella Chemotype of Cymbopogon flexuosus (Nees ex Steud.) W.Watson Acclimatized in Congo-Brazzaville. Chem. Biodivers. 2019, 16, e1800446. [Google Scholar] [CrossRef]
- Fadel, H.; Benayache, F.; Chalchat, J.C.; Figueredo, G.; Chalard, P.; Hazmoune, H.; Benayache, S. Essential oil constituents of Juniperus oxycedrus L. and Cupressus sempervirens L. (Cupressaceae) growing in Aures region of Algeria. Nat. Prod. Res. 2021, 35, 2616–2620. [Google Scholar] [CrossRef]
- Rosato, A.; Sblano, S.; Salvagno, L.; Carocci, A.; Clodoveo, M.L.; Corbo, F.; Fracchiolla, G. Anti-Biofilm Inhibitory Synergistic Effects of Combinations of Essential Oils and Antibiotics. Antibiotics 2020, 9, 637. [Google Scholar] [CrossRef]
- Gómez-Sequeda, N.; Cáceres, M.; Stashenko, E.E.; Hidalgo, W.; Ortiz, C. Antimicrobial and Antibiofilm Activities of Essential Oils against Escherichia coli O157, H7 and Methicillin-Resistant Staphylococcus aureus (MRSA). Antibiotics 2020, 9, 730. [Google Scholar] [CrossRef]
- Akin, M.; Aktumsek, A.; Nostro, A. Antibacterial activity and composition of the essential oils of Eucalyptus camaldulensis Dehn and Myrtus communis L. growing in Northern Cyprus. Afr. J. Biotechnol. 2010, 9, 531–535. [Google Scholar]
- Jardak, M.; Elloumi-Mseddi, J.; Aifa, S.; Mnif, S. Chemical composition, anti-biofilm activity and potential cytotoxic effect on cancer cells of Rosmarinus officinalis L. essential oil from Tunisia. Lipids Health Dis. 2017, 16, 190. [Google Scholar] [CrossRef]
- Poli, J.P.; Guinoiseau, E.; de Rocca Serra, D.; Sutour, S.; Paoli, M.; Tomi, F.; Quilichini, Y.; Berti, L.; Lorenzi, V. Anti-Quorum Sensing Activity of 12 Essential Oils on chromobacterium violaceum and Specific Action of cis-cis-p-Menthenolide from Corsican Mentha suaveolens ssp. Insularis. Molecules 2018, 23, 2125. [Google Scholar] [CrossRef]
- Passerini, D.; Fécamp, F.; Marchand, L.; Kolypczuk, L.; Bonnetot, S.; Sinquin, C.; Verrez-Bagnis, V.; Hervio-Heath, D.; Colliec-Jouault, S.; Delbarre-Ladrat, C. Characterization of Biofilm Extracts from Two Marine Bacteria. Appl. Sci. 2019, 9, 4971. [Google Scholar] [CrossRef]
- Mitra, S.; Thawrani, D.; Banerjee, P.; Gachhui, R.; Mukherjee, J. Induced biofilm cultivation enhances riboflavin production by an intertidally derived Candida famata. Appl. Biochem. Biotechnol. 2012, 166, 1991–2006. [Google Scholar] [CrossRef]
- Victor, I.U.; Kwiencien, M.; Tripathi, L.; Cobice, D.; McClean, S.; Marchant, R.; Banat, I.M. Quorum sensing as a potential target for increased production of rhamnolipid biosurfactant in Burkholderia thailandensis E264. Appl. Microbiol. Biotechnol. 2019, 103, 6505–6517. [Google Scholar] [CrossRef]
- Schmidt, S.; Blom, J.F.; Pernthaler, J.; Berg, G.; Baldwin, A.; Mahenthiralingam, E.; Eberl, L. Production of the antifungal compound pyrrolnitrin is quorum sensing-regulated in members of the Burkholderia cepacia complex. Environ. Microbiol. 2009, 11, 1422–1437. [Google Scholar] [CrossRef]
- Issac Abraham, S.V.; Palani, A.; Ramaswamy, B.R.; Shunmugiah, K.P.; Arumugam, V.R. Antiquorum sensing and antibiofilm potential of Capparis spinosa. Arch. Med. Res. 2011, 42, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Okoh, S.O.; Iweriegbor, B.C.; Okoh, O.O.; Nwodo, U.U.; IOkoh, A. Bactericidal and antioxidant properties of essential oils from the fruits Dennettia tripetala G. Baker. BMC Complement Altern. Med. 2016, 16, 486. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Manrique-Moreno, M.; Klaiss-Luna, M.C.; Stashenko, E.; Zafra, G.; Ortiz, C. Effect of Essential Oils on Growth Inhibition, Biofilm Formation and Membrane Integrity of Escherichia coli and Staphylococcus aureus. Antibiotics 2021, 10, 1474. [Google Scholar] [CrossRef]
- Kim, M.K.; Zhao, A.; Wang, A.; Brown, Z.Z.; Muir, T.W.; Stone, H.A.; Bassler, B.L. Surface-attached molecules control Staphylococcus aureus quorum sensing and biofilm development. Nat. Microbiol. 2017, 2, 17080. [Google Scholar] [CrossRef] [PubMed]
- Quave, C.L.; Horswill, A.R. Identification of Staphylococcal Quorum Sensing Inhibitors by Quantification of õ-Hemolysin with High Performance Liquid Chromatography. Methods Mol. Biol. 2018, 1673, 363–370. [Google Scholar] [PubMed]
- Das, S.; Horváth, B.; Šafranko, S.; Jokić, S.; Széchenyi, A.; Kőszegi, T. Antimicrobial Activity of Chamomile Essential Oil: Effect of Different Formulations. Molecules 2019, 24, 4321. [Google Scholar] [CrossRef] [PubMed]
- Correa-Llantén, D.N.; Amenábar, M.J.; Blamey, J.M. Antioxidant capacity of novel pigments from an Antarctic bacterium. J. Microbiol. 2012, 50, 374–379. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Ong, K.S.; Cheow, Y.L.; Lee, S.M. The role of reactive oxygen species in the antimicrobial activity of pyochelin. J. Adv. Res. 2017, 8, 393–398. [Google Scholar] [CrossRef]
- Van Acker, H.; Gielis, J.; Acke, M.; Cools, F.; Cos, P.; Coenye, T. The Role of Reactive Oxygen Species in Antibiotic-Induced Cell Death in Burkholderia cepacia Complex Bacteria. PLoS ONE 2016, 11, e0159837. [Google Scholar] [CrossRef]
- Gasser, V.; Baco, E.; Cunrath, O.; August, P.S.; Perraud, Q.; Zill, N.; Schleberger, C.; Schmidt, A.; Paulen, A.; Bumann, D.; et al. Catechol siderophores repress the pyochelin pathway and activate the enterobactin pathway in Pseudomonas aeruginosa: An opportunity for siderophore-antibiotic conjugates development. Environ. Microbiol. 2016, 18, 819–832. [Google Scholar] [CrossRef]
- Micota, B.; Sadowska, B.; Podsędek, A.; Paszkiewicz, M.; Sosnowska, D.; Różalska, B. Is it true that plant-derived polyphenols are always beneficial for the human? In vitro study on Leonurus cardiaca extract properties in the context of the pathogenesis of Staphylococcus aureus infections. J. Med. Microbiol. 2016, 65, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.M.; Abdullah, N.F.; Amom, Z. Killing of Staphylococcus aureus by allylpyrocatechol is potentiated by induction of intracellular oxidative stress and inhibition of catalase activity. J. Integr. Med. 2016, 14, 456–464. [Google Scholar] [CrossRef]
- Cai, J.Y.; Li, J.; Hou, Y.N.; Ma, K.; Yao, G.D.; Liu, W.W.; Hayashi, T.; Itoh, K.; Tashiro, S.I.; Onodera, S.; et al. Concentration-dependent dual effects of silibinin on kanamycin-induced cells death in Staphylococcus aureus. Biomed Pharmacother. 2018, 102, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Sinsinwar, S.; Vadivel, V. Catechin isolated from cashew nut shell exhibits antibacterial activity against clinical isolates of MRSA through ROS-mediated oxidative stress. Appl. Microbiol. Biotechnol. 2020, 104, 8279–8297. [Google Scholar] [CrossRef]
- Golenia, A.; Leśkiewicz, M.; Regulska, M.; Budziszewska, B.; Szczęsny, E.; Jagiełła, J.; Wnuk, M.; Ostrowska, M.; Lasoń, W.; Basta-Kaim, A.; et al. Catalase activity in blood fractions of patients with sporadic ALS. Pharm. Rep. 2014, 66, 704–707. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Qureshi, K.A.; Ali, H.M.; Al-Omar, M.S.; Khan, O.; Mohammed, S.A.A. Bio-Evaluation of the Wound Healing Activity of Artemisia judaica L. as Part of the Plant’s Use in Traditional Medicine; Phytochemical, Antioxidant, Anti-Inflammatory, and Antibiofilm Properties of the Plant’s Essential Oils. Antioxidants 2022, 11, 332. [Google Scholar] [CrossRef] [PubMed]
- Monsalves, M.T.; Ollivet-Besson, G.P.; Amenabar, M.J.; Blamey, J.M. Isolation of a Psychrotolerant and UV-C-Resistant Bacterium from Elephant Island, Antarctica with a Highly Thermoactive and Thermostable Catalase. Microorganisms 2020, 8, 95. [Google Scholar] [CrossRef]
- Barros, D.; Pradhan, A.; Mendes, V.M.; Manadas, B.; Santos, P.M.; Pascoal, C.; Cássio, F. Proteomics and antioxidant enzymes reveal different mechanisms of toxicity induced by ionic and nanoparticulate silver in bacteria. Environ. Sci. Nano 2019, 6, 1207–1218. [Google Scholar] [CrossRef]
- Hamida, R.S.; Ali, M.A.; Goda, D.A.; Khalil, M.I.; Al-Zaban, M.I. Novel Biogenic Silver Nanoparticle-Induced Reactive Oxygen Species Inhibit the Biofilm Formation and Virulence Activities of Methicillin-Resistant Staphylococcus aureus (MRSA) Strain. Front. Bioeng. Biotechnol. 2020, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Statti, G.A.; Tundis, R.; Menichini, F.; Houghton, P. Antioxidant activity of methanolic extract of Hypericum triquetrifolium Turra aerial part. Fitoterapia 2002, 73, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Magina, M.D.; Dalmarco, E.M.; Wisniewski AJr Simionatto, E.L.; Dalmarco, J.B.; Pizzolatti, M.G.; Brighente, I.M.C. Chemical composition and antibacterial activity of essen-tial oils of Eugenia species. J. Nat. Med. 2009, 63, 345–350. [Google Scholar] [CrossRef]
- Quinn, P.J.; Carter, M.E.; Markey, B.K.; Carter, G.R. The streptococci and related cocci. In Clinical Veterinary Microbiology; Quinn, P.J., Carter, M.E., Markey, B., Carter, G.R., Eds.; Mosby Ltd.: London, UK, 1994; p. 127e36. [Google Scholar]
- Kanemitsu, K.; Nishino, T.; Kunishima, H.; Okamura, N.; Takemura, H.; Yamamoto, H.; Kaku, M. Quantitative determination of gelatinase activity among enterococci. J. Microbiol. Methods 2001, 47, 11–16. [Google Scholar] [CrossRef]
- Arakha, M.; Pal, S.; Samantarrai, D.; Panigrahi, T.K.; Mallick, B.C.; Pramanik, K. Antimicrobial activity of iron oxide nanoparticle upon modulation of nanoparticle-bacteria interface. Sci. Rep. 2015, 5, 14813. [Google Scholar] [CrossRef]
- Han, L.; Patil, S.; Boehm, D.; Milosavljevic, V.; Cullen, P.J.; Bourke, P. Mechanisms of inactivation by high-voltage atmospheric cold plasma differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 450–458. [Google Scholar] [CrossRef]
- Martins, D.; McKay, G.; Sampathkumar, G.; Khakimova, M.; English, A.M.; Nguyen, D. Superoxide dismutase activity confers (p)ppGppmediated antibiotic tolerance to stationary-phase Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2018, 115, 9797–9802. [Google Scholar] [CrossRef]
- Acuña, L.G.; Calderón, I.L.; Elías, A.O.; Castro, M.E.; Vásquez, C.C. Expression of the yggE gene protects Escherichia coli from potassium tellurite-generated oxidative stress. Arch. Microbiol. 2009, 191, 473–476. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N ° | KI * | Component | R. officinalis | M. communis |
|---|---|---|---|---|
| 1 | 923 | Tricyclene | 0.24 | 0.00 |
| 2 | 927 | α-Thujene | 0.11 | 0.51 |
| 3 | 934 | α-Pinene | 3.39 | 45.3 |
| 4 | 949 | Camphene | 6.52 | 0.04 |
| 5 | 974 | Sabinene | 0.05 | 4.42 |
| 6 | 978 | β-Pinene | 3.59 | 0.00 |
| 7 | 992 | β-Myrcene | 0.05 | 0.30 |
| 8 | 1002 | α-Phellandrene | 1.84 | 0.00 |
| 9 | 1004 | δ-2-Carene | 0.00 | 0.72 |
| 10 | 1006 | δ-3-Carene | 0.24 | 0.67 |
| 11 | 1012 | α-Terpinene | 0.40 | 0.45 |
| 12 | 1025 | p-Cymene | 0.95 | 1.64 |
| 13 | 1029 | Limonene | 2.52 | 8.45 |
| 14 | 1038 | (Z)-β-Ocimene | 0.08 | 0.06 |
| 15 | 1048 | (E)-β -Ocimene | 0.07 | 0.23 |
| 16 | 1058 | ϒ-Terpinene | 0.47 | 0.39 |
| 17 | 1090 | α-Terpinolene | 0.35 | 0.42 |
| 18 | 1032 | 1.8-Cineole | 37.56 | 22.02 |
| 19 | 1101 | Linalool | 0.51 | 2.63 |
| 20 | 1109 | α-Thujone | 0.07 | 0.00 |
| 21 | 1119 | β-Thujone | 0.05 | 0.00 |
| 22 | 1127 | allo-Ocimene | 0.00 | 0.00 |
| 23 | 1127 | α-Campholenal | 0.04 | 0.01 |
| 24 | 1139 | (E)-pinocarveol | 0.00 | 0.13 |
| 25 | 1146 | Camphor | 7.10 | 0.23 |
| 26 | 1158 | Isoborneol | 0.01 | 0.00 |
| 27 | 1166 | β-Fenchyl alcohol | 0.00 | 0.06 |
| 28 | 1167 | Borneol | 4.37 | 0.00 |
| 29 | 1178 | Terpinen-4-ol | 0.53 | 0.85 |
| 30 | 1186 | (Z)-Pinocarveol | 0.00 | 0.07 |
| 31 | 1186 | p-Cymen-8-ol | 0.04 | 0.00 |
| 32 | 1192 | α-Terpineol | 1.57 | 1.60 |
| 33 | 1207 | Verbenone | 0.02 | 0.00 |
| 34 | 1258 | Linalyl acetate | 0.00 | 0.95 |
| 35 | 1287 | Bornyl acetate | 0.24 | 0.17 |
| 36 | 1294 | Thymol | 0.03 | 0.00 |
| 37 | 1304 | Carvacrol | 0.00 | 0.05 |
| 38 | 1325 | Myrtenyl acetate | 0.00 | 0.06 |
| 39 | 1350 | α-Terpinyl-acetate | 0.00 | 0.08 |
| 40 | 1373 | α-Ylangene | 0.11 | 0.00 |
| 41 | 1377 | (E)-Methyl cinnamate | 0.00 | 0.07 |
| 42 | 1385 | Geranyl acetate | 0.36 | 0.00 |
| 43 | 1420 | (E)-Caryophyllene | 5.04 | 0.67 |
| 44 | 1429 | Aromadendrene | 0.05 | 0.00 |
| 45 | 1438 | α-Guaiene | 0.00 | 0.11 |
| 46 | 1455 | α-Humulene | 0.75 | 0.15 |
| 47 | 1462 | allo-Aromadendrene | 0.00 | 0.00 |
| 48 | 1496 | α-Muurolene | 0.27 | 0.00 |
| 49 | 1498 | (E)-β-Guaiene | 0.00 | 0.06 |
| 50 | 1500 | α-Amorphene | 0.12 | 0.00 |
| 51 | 1510 | δ-Amorphene | 0.00 | 0.06 |
| 52 | 1523 | (Z)-Calamenene | 0.00 | 0.13 |
| 53 | 1525 | δ-Cadinene | 0.65 | 0.00 |
| 54 | 1584 | Caryophyllene oxide | 0.06 | 0.12 |
| 55 | 1593 | Vidiflorol | 0.03 | 0.00 |
| 56 | 1656 | α-Cadinol | 0.00 | 0.03 |
| Total (%) | 96.39 | 96.57 |
| EOs | Concentration (mg/mL) | |
|---|---|---|
| EO1 | MIC | 11.25 |
| MBC | 360 | |
| MBC/MIC | 32 | |
| EO2 | MIC | 0.7 |
| MBC | 22.5 | |
| MBC/MIC | 32.14 | |
| EO1 + EO2 | MIC | 5.63 |
| MBC | 180 | |
| MBC/MIC | 31.97 | |
| EO1 | EO2 | EO1 + EO2 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A-H | A-D | A-P | A-H | A-D | A-P | A-H | A-D | A-P | |
| Control | 20 ± 0.0 | 20 ± 0.0 | 18.4 ± 0.5 | 20 ± 0.0 | 20 ± 0.0 | 18.4 ± 0.5 | 20 ± 0.0 | 20 ± 0.0 | 18.4 ± 0.5 |
| MIC/2 | 15.5 ± 0.5 * | 18 ± 0.0 | 16 ± 0.47 | 14.6 ± 0.5 * | 17 ± 0.8 | 14.6 ± 0.4 | 13 ± 0.8 * | 17 ± 1.4 | 14.6 ± 0.5 |
| MIC | 15.3 ± 0.5 * | 17 ± 0.0 | 16 ± 1.25 | 14 ± 0.8 * | 16.3 ± 1.2 | 14.5 ± 0.4 | 12.5 ± 0.5 * | 17 ± 0.8 | 14.3 ± 0.5 |
| MIC × 2 | 13.6 ± 1.2 * | 17 ± 0.0 | 16 ± 0.47 | 13.6 ± 0.9 * | 16 ± 1.0 | 13.3 ± 0.9 | 11 ± 0.8 * | 17 ± 0.8 | 14 ± 0.0 |
| MIC × 4 | 13 ± 00 * | 15.5 ± 0.5 * | 16 ± 0 | 12.6 ± 0.5 * | 15.3 ± 0.5 * | 13 ± 0.4 * | 9 ± 0.8 * | 16 ± 0.0 | 11.6 ± 0.5 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowbe, K.H.; Salah, K.B.H.; Moumni, S.; Ashkan, M.F.; Merghni, A. Anti-Staphylococcal Activities of Rosmarinus officinalis and Myrtus communis Essential Oils through ROS-Mediated Oxidative Stress. Antibiotics 2023, 12, 266. https://doi.org/10.3390/antibiotics12020266
Bowbe KH, Salah KBH, Moumni S, Ashkan MF, Merghni A. Anti-Staphylococcal Activities of Rosmarinus officinalis and Myrtus communis Essential Oils through ROS-Mediated Oxidative Stress. Antibiotics. 2023; 12(2):266. https://doi.org/10.3390/antibiotics12020266
Chicago/Turabian StyleBowbe, Khadijetou Hamoud, Karima Bel Hadj Salah, Sarra Moumni, Mada F. Ashkan, and Abderrahmen Merghni. 2023. "Anti-Staphylococcal Activities of Rosmarinus officinalis and Myrtus communis Essential Oils through ROS-Mediated Oxidative Stress" Antibiotics 12, no. 2: 266. https://doi.org/10.3390/antibiotics12020266
APA StyleBowbe, K. H., Salah, K. B. H., Moumni, S., Ashkan, M. F., & Merghni, A. (2023). Anti-Staphylococcal Activities of Rosmarinus officinalis and Myrtus communis Essential Oils through ROS-Mediated Oxidative Stress. Antibiotics, 12(2), 266. https://doi.org/10.3390/antibiotics12020266