


Comprehensive Study of Components and Antimicrobial Properties of Essential Oil Extracted from Carum carvi L. Seeds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains and Media
2.3. Preparation of CEO and Component Analysis
2.4. Testing the Susceptibility of Planktonic Bacteria
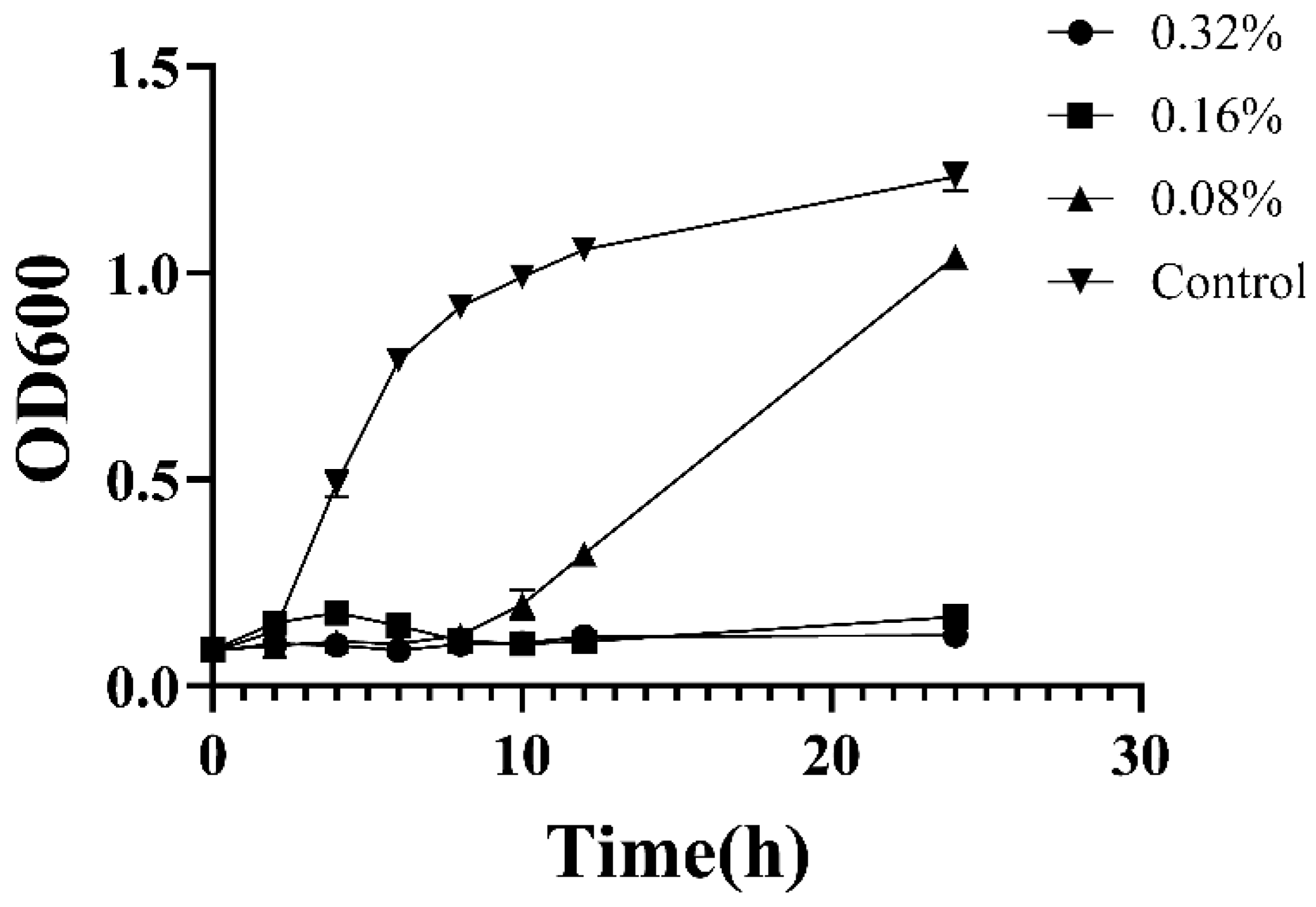
2.5. Time Kill Curve
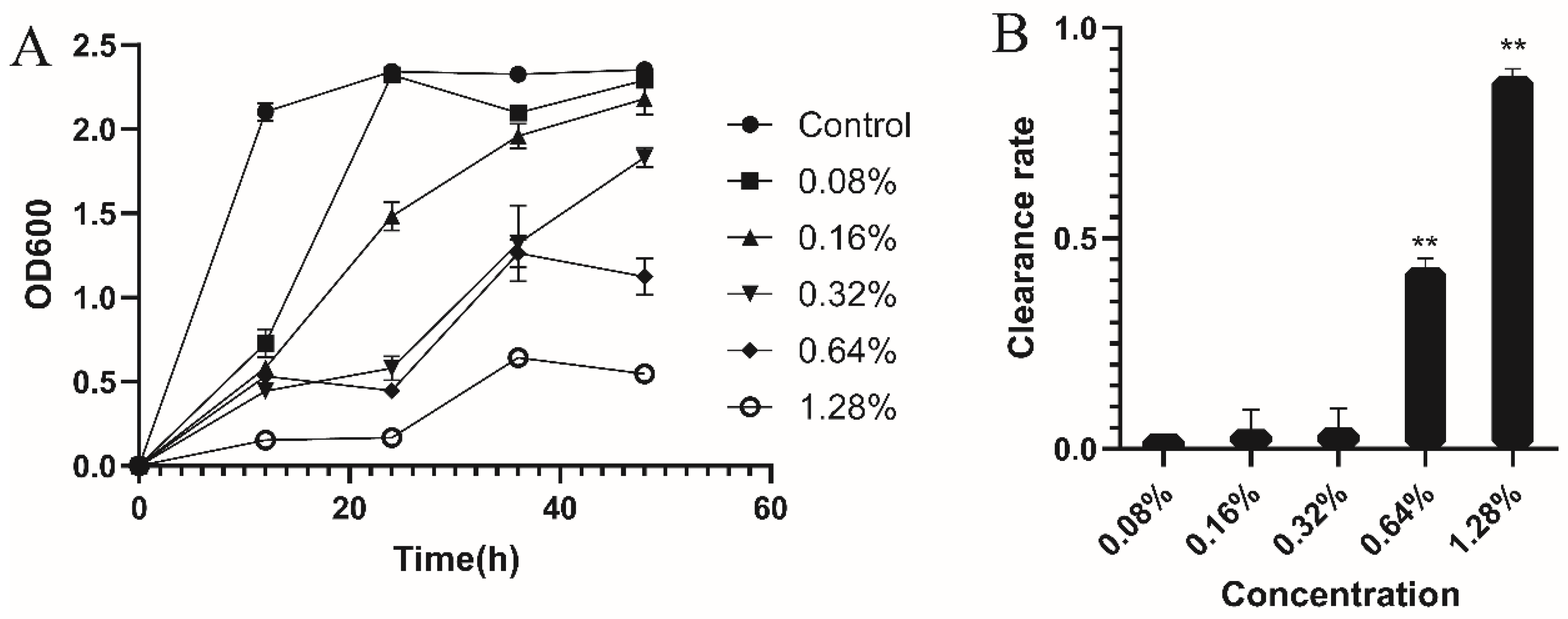
2.6. The Effect on Biofilm
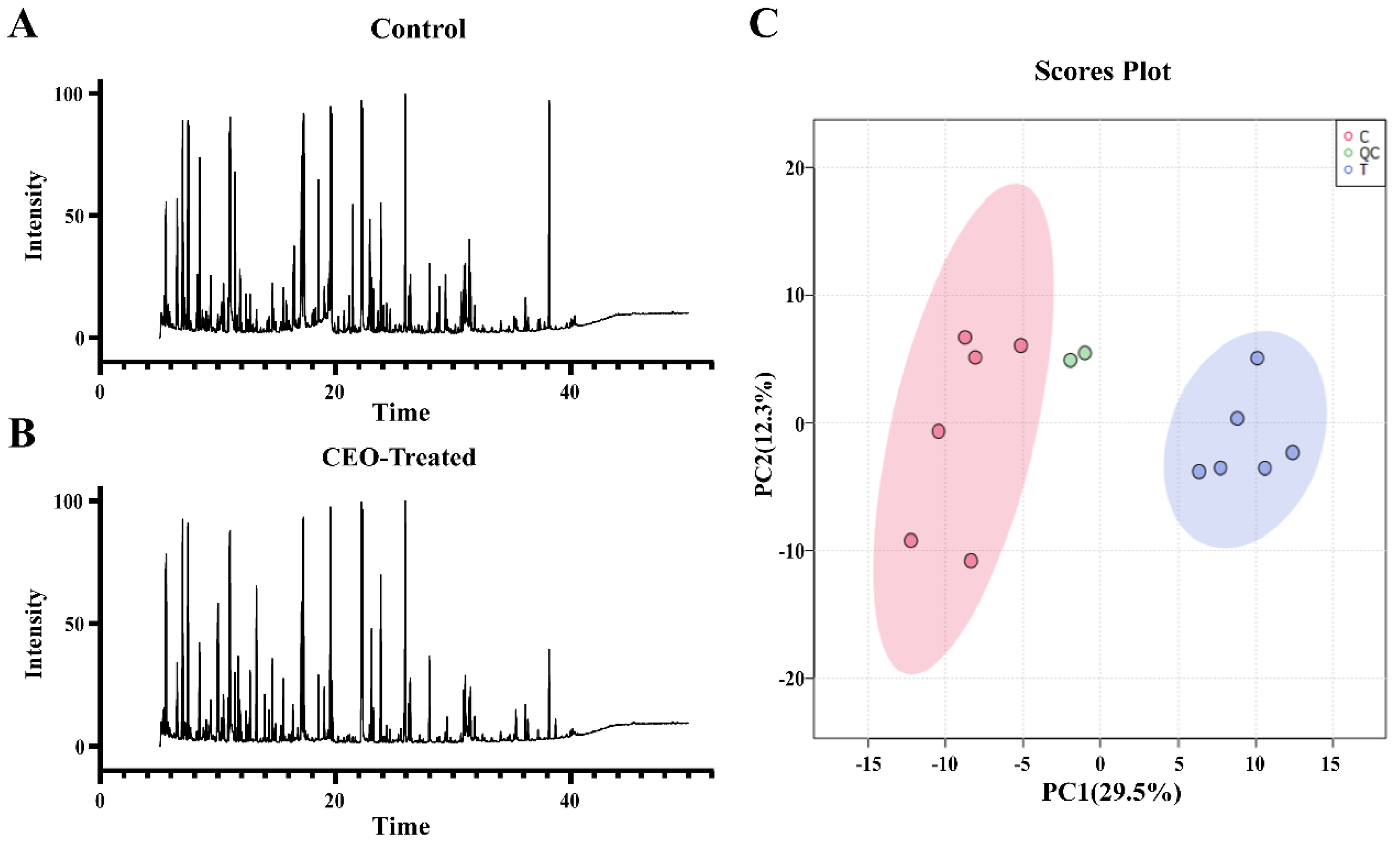
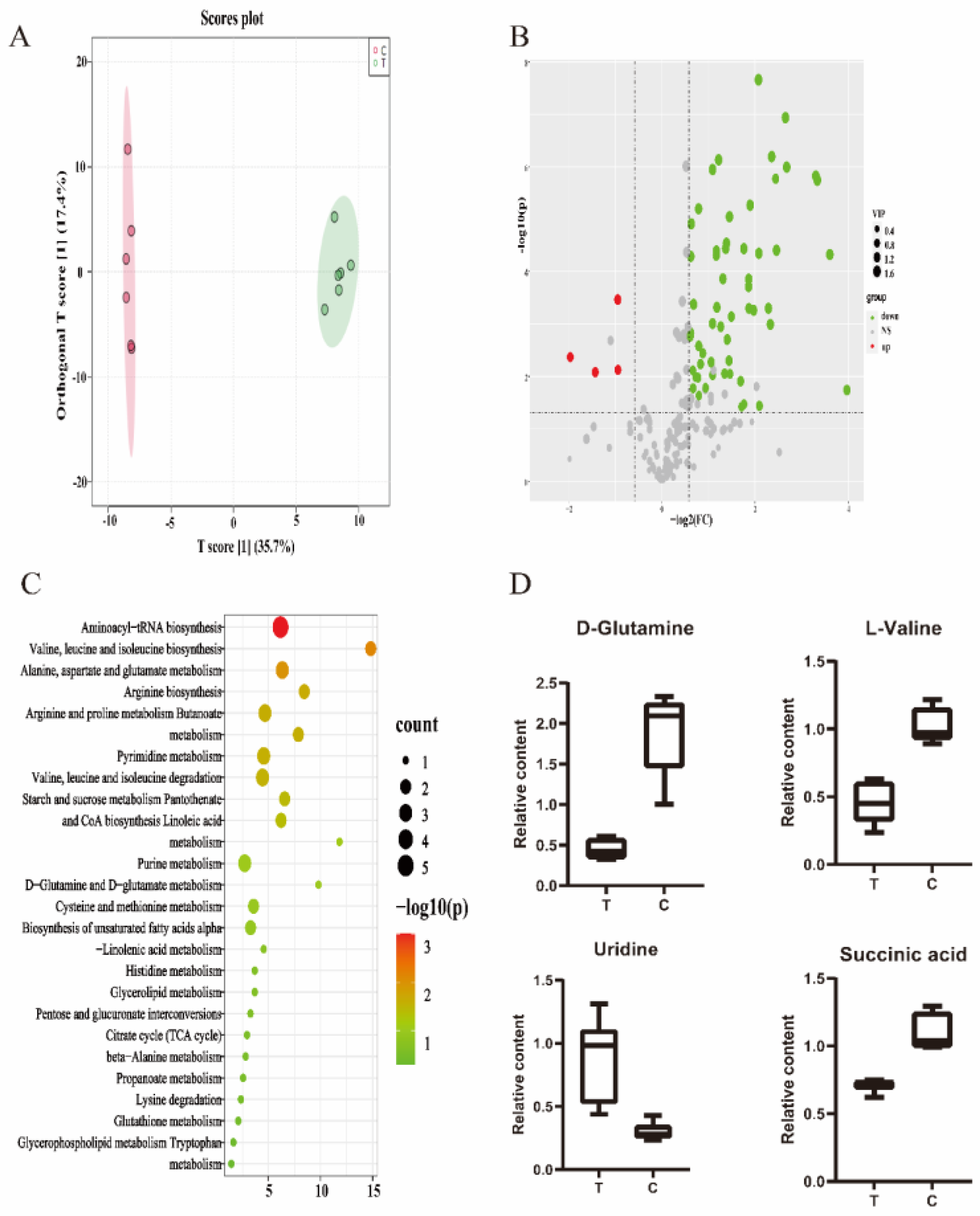
2.7. Metabolite Extraction and Analysis
2.8. Statistical Analysis
3. Results
3.1. Chemical Component of CEO
3.2. Antibacterial Activity
3.3. Antibiofilm Activity
3.4. Metabolomics
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohammed, Y.H.E.; Manukumar, H.M.; Rakesh, K.P.; Karthik, C.S.; Mallu, P.; Qin, H.L. Vision for medicine: Staphylococcus aureus biofilm war and unlocking key’s for anti-biofilm drug development. Microb. Pathog. 2018, 123, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Choo, E.J. Community-Associated Methicillin-Resistant Staphylococcus aureus in Nosocomial Infections. Infect. Chemother. 2017, 49, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Romero, L.C.; de Souza da Cunha, M.L.R. Insights into the epidemiology of community-associated methicillin-resistant Staphylococcus aureus in special populations and at the community-healthcare interface. Braz. J. Infect Dis. 2021, 25, 101636. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [Green Version]
- Dayan, G.H.; Mohamed, N.; Scully, I.L.; Cooper, D.; Begier, E.; Eiden, J.; Jansen, K.U.; Gurtman, A.; Anderson, A.S. Staphylococcus aureus: The current state of disease, pathophysiology and strategies for prevention. Expert Rev. Vaccines 2016, 15, 1373–1392. [Google Scholar] [CrossRef]
- Otto, M. Community-associated MRSA: What makes them special? Int. J. Med. Microbiol. 2013, 303, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Diekema, D.J.; Pfaller, M.A.; Shortridge, D.; Zervos, M.; Jones, R.N. Twenty-Year Trends in Antimicrobial Susceptibilities among Staphylococcus aureus from the SENTRY Antimicrobial Surveillance Program. Open Forum Infect. Dis. 2019, 6, S47–S53. [Google Scholar] [CrossRef]
- Diekema, D.J.; Pfaller, M.A.; Turnidge, J.; Verhoef, J.; Bell, J.; Fluit, A.C.; Doern, G.V.; Jones, R.N.; Sentry Participants, G. Genetic relatedness of multidrug-resistant, methicillin (oxacillin)-resistant Staphylococcus aureus bloodstream isolates from SENTRY Antimicrobial Resistance Surveillance Centers worldwide, 1998. Microb. Drug Resist. 2000, 6, 213–221. [Google Scholar] [CrossRef]
- Furukawa, S.; Kuchma, S.L.; O’Toole, G.A. Keeping their options open: Acute versus persistent infections. J. Bacteriol. 2006, 188, 1211–1217. [Google Scholar] [CrossRef] [Green Version]
- Craft, K.M.; Nguyen, J.M.; Berg, L.J.; Townsend, S.D. Methicillin-resistant Staphylococcus aureus (MRSA): Antibiotic-resistance and the biofilm phenotype. Medchemcomm 2019, 10, 1231–1241. [Google Scholar] [CrossRef]
- Mah, T.F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, I.; Silva-Espinoza, B.A.; Ortega-Ramirez, L.A.; Leyva, J.M.; Siddiqui, M.W.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Ayala-Zavala, J.F. Oregano Essential Oil as an Antimicrobial and Antioxidant Additive in Food Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 1717–1727. [Google Scholar] [CrossRef]
- Ramsey, J.T.; Shropshire, B.C.; Nagy, T.R.; Chambers, K.D.; Li, Y.; Korach, K.S. Essential Oils and Health. Yale J. Biol. Med. 2020, 93, 291–305. [Google Scholar]
- Firenzuoli, F.; Jaitak, V.; Horvath, G.; Bassole, I.H.; Setzer, W.N.; Gori, L. Essential oils: New perspectives in human health and wellness. Evid. Based Complement. Altern. Med. 2014, 2014, 467363. [Google Scholar] [CrossRef]
- Yadav, E.; Kumar, S.; Mahant, S.; Khatkar, S.; Rao, R. Tea tree oil: A promising Essential Oil. J. Essent. Oil Res. 2016, 29, 201–213. [Google Scholar] [CrossRef]
- Rasooli, I.; Allameh, A. Caraway (Carum carvi L.) Essential Oils. In Essential Oils in Food Preservation, Flavor and Safety; Elsevier: Amsterdam, The Netherlands, 2016; pp. 287–293. [Google Scholar]
- Hajlaoui, H.; Arraouadi, S.; Noumi, E.; Aouadi, K.; Adnan, M.; Khan, M.A.; Kadri, A.; Snoussi, M. Antimicrobial, Antioxidant, Anti-Acetylcholinesterase, Antidiabetic, and Pharmacokinetic Properties of Carum carvi L. and Coriandrum sativum L. Essential Oils Alone and in Combination. Molecules 2021, 26, 3625. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, N.S.; Lo Cantore, P.; Capasso, F.; Senatore, F. Antibacterial activity of Cuminum cyminum L. and Carum carvi L. essential oils. J. Agric. Food Chem. 2005, 53, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, A.; Minaiyan, M.; Ghannadi, A.; Mahzouni, P. Effects of Carum carvi L. (Caraway) extract and essential oil on TNBS-induced colitis in rats. Res. Pharm. Sci. 2013, 8, 1–8. [Google Scholar]
- Klys, M.; Izdebska, A.; Malejky-Klusek, N. Repellent Effect of the Caraway Carum carvi L. on the Rice Weevil Sitophilus oryzae L. (Coleoptera, Dryophthoridae). Insects 2020, 11, 836. [Google Scholar] [CrossRef]
- Maurya, A.; Kumar, S.; Singh, B.K.; Chaudhari, A.K.; Dwivedy, A.K.; Prakash, B.; Dubey, N.K. Mechanistic investigations on antifungal and antiaflatoxigenic activities of chemically characterised Carum carvi L. essential oil against fungal infestation and aflatoxin contamination of herbal raw materials. Nat. Prod. Res. 2022, 36, 4569–4574. [Google Scholar] [CrossRef]
- Ghannay, S.; Aouadi, K.; Kadri, A.; Snoussi, M. GC-MS Profiling, Vibriocidal, Antioxidant, Antibiofilm, and Anti-Quorum Sensing Properties of Carum carvi L. Essential Oil: In Vitro and In Silico Approaches. Plants 2022, 11, 1072. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, C.; Zhang, R.; Ye, S.; Zhao, Z.; Huang, Y.; Xu, X.; Lan, W.; Yang, D. Metabolomics analysis to evaluate the antibacterial activity of the essential oil from the leaves of Cinnamomum camphora (Linn.) Presl. J. Ethnopharmacol. 2020, 253, 112652. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W6602009. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. MSEA: A web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res. 2010, 38, W71–W772010. [Google Scholar] [CrossRef] [Green Version]
- Laribi, B.; Kouki, K.; Bettaieb, T.; Mougou, A.; Marzouk, B. Essential oils and fatty acids composition of Tunisian, German and Egyptian caraway (Carum carvi L.) seed ecotypes: A comparative study. Ind. Crops Prod. 2013, 41, 312–318. [Google Scholar] [CrossRef]
- Homayounpour, P.; Shariatifar, N.; Alizadeh-Sani, M. Development of nanochitosan-based active packaging films containing free and nanoliposome caraway (Carum carvi. L) seed extract. Food Sci. Nutr. 2021, 9, 553–563. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Fekry, M.; Yahya, G.; Osman, A.; Al-Rabia, M.W.; Mostafa, I.; Abbas, H.A. GC-MS Analysis and Microbiological Evaluation of Caraway Essential Oil as a Virulence Attenuating Agent against Pseudomonas aeruginosa. Molecules 2022, 27, 8523. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Feng, M.; Yang, K.; Cao, Y.; Zhang, J.; Xu, J.; Hernandez, S.H.; Wei, X.; Fan, M. Transcriptomic and metabolomic analyses reveal antibacterial mechanism of astringent persimmon tannin against Methicillin-resistant Staphylococcus aureus isolated from pork. Food Chem. 2020, 309, 125692. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, J.; Zhu, L.; Qian, C.; Tian, H.; Zhao, Z.; Jin, L.; Yang, D. Antibacterial Activity of the Essential Oil from Litsea cubeba against Cutibacterium acnes and the Investigations of Its Potential Mechanism by Gas Chromatography-Mass Spectrometry Metabolomics. Front. Microbiol. 2022, 13, 823845. [Google Scholar] [CrossRef]
- Kaiser, J.C.; King, A.N.; Grigg, J.C.; Sheldon, J.R.; Edgell, D.R.; Murphy, M.E.P.; Brinsmade, S.R.; Heinrichs, D.E. Repression of branched-chain amino acid synthesis in Staphylococcus aureus is mediated by isoleucine via CodY, and by a leucine-rich attenuator peptide. PLoS Genet. 2018, 14, e1007159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Sirobhushanam, S.; Ring, R.P.; Singh, S.; Gatto, C.; Wilkinson, B.J. Roles of pyruvate dehydrogenase and branched-chain alpha-keto acid dehydrogenase in branched-chain membrane fatty acid levels and associated functions in Staphylococcus aureus. J. Med. Microbiol. 2018, 67, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and electron microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.G.; Hu, F.; Wei, Z.J. Antibacterial Activity and Mechanism of Ginger Essential Oil against Escherichia coli and Staphylococcus aureus. Molecules 2020, 25, 3955. [Google Scholar] [CrossRef]
- Bautista-Silva, J.P.; Seibert, J.B.; Amparo, T.R.; Rodrigues, I.V.; Teixeira, L.F.M.; Souza, G.H.B.; Dos Santos, O.D.H. Melaleuca leucadendra Essential Oil Promotes Loss of Cell Membrane and Wall Integrity and Inhibits Bacterial Growth: An In Silico and In Vitro Approach. Curr. Microbiol. 2020, 77, 2181–2191. [Google Scholar] [CrossRef]
- Karaolis, D.K.; Rashid, M.H.; Chythanya, R.; Luo, W.; Hyodo, M.; Hayakawa, Y. c-di-GMP (3′-5′-cyclic diguanylic acid) inhibits Staphylococcus aureus cell-cell interactions and biofilm formation. Antimicrob. Agents Chemother. 2005, 49, 1029–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garavaglia, M.; Rossi, E.; Landini, P. The pyrimidine nucleotide biosynthetic pathway modulates production of biofilm determinants in Escherichia coli. PLoS ONE 2012, 7, e31252. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Mo, Y.; Chen, K.; Shang, X.; Yang, Z.; Hao, B.; Shang, R.; Liang, J.; Liu, Y. Integration of metabolomics and transcriptomics indicates changes in MRSA exposed to terpinen-4-ol. BMC Microbiol. 2021, 21, 305. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Component | RT | RI | Percentage (%) | Structure |
|---|---|---|---|---|---|
| 1 | β-Pinene | 6.652 | 990 | 0.11 |  |
| 2 | Limonene | 7.989 | 1027 | 28.55 |  |
| 3 | γ-Terpinene | 9.146 | 1057 | 0.15 |  |
| 4 | Dihydrocarvone | 14.319 | 1194 | 0.13 |  |
| 5 | Dihydrocarvone | 14.584 | 1201 | 0.10 |  |
| 6 | Carveol | 15.161 | 1217 | 0.07 |  |
| 7 | Ethanol, 2-(3,3-dimethylcyclohexylidene) | 15.479 | 1226 | 0.11 |  |
| 8 | Carvone | 16.039 | 1242 | 69.78 |  |
| 9 | Perilla aldehyde | 17.042 | 1271 | 0.14 |  |
| 10 | Others | 0.85 | |||
| Total | 100.00 |
| Bacteria | Strains | CEO (v/v) | Chlorhexidine (μg/mL) | |
|---|---|---|---|---|
| MIC | MBC | MIC | ||
| MRSA | ATCC 43300 | 0.16% | 0.64% | 6.4 |
| S. aureus | ATCC 6538 | 0.16% | 0.32% | 6.4 |
| E. coli | ATCC 43895 | 0.16% | 0.32% | 6.4 |
| E. coli | ATCC 25932 | 0.16% | 0.32% | 6.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Cheng, F.; Aisa, H.A.; Maiwulanjiang, M. Comprehensive Study of Components and Antimicrobial Properties of Essential Oil Extracted from Carum carvi L. Seeds. Antibiotics 2023, 12, 591. https://doi.org/10.3390/antibiotics12030591
Liu C, Cheng F, Aisa HA, Maiwulanjiang M. Comprehensive Study of Components and Antimicrobial Properties of Essential Oil Extracted from Carum carvi L. Seeds. Antibiotics. 2023; 12(3):591. https://doi.org/10.3390/antibiotics12030591
Chicago/Turabian StyleLiu, Changhai, Feng Cheng, Haji Akber Aisa, and Maitinuer Maiwulanjiang. 2023. "Comprehensive Study of Components and Antimicrobial Properties of Essential Oil Extracted from Carum carvi L. Seeds" Antibiotics 12, no. 3: 591. https://doi.org/10.3390/antibiotics12030591
APA StyleLiu, C., Cheng, F., Aisa, H. A., & Maiwulanjiang, M. (2023). Comprehensive Study of Components and Antimicrobial Properties of Essential Oil Extracted from Carum carvi L. Seeds. Antibiotics, 12(3), 591. https://doi.org/10.3390/antibiotics12030591