

Perchlozone Resistance in Clinical Isolates of Mycobacterium tuberculosis

 ,
, 
{kind=link}
{kind=link}
Abstract
1. Introduction
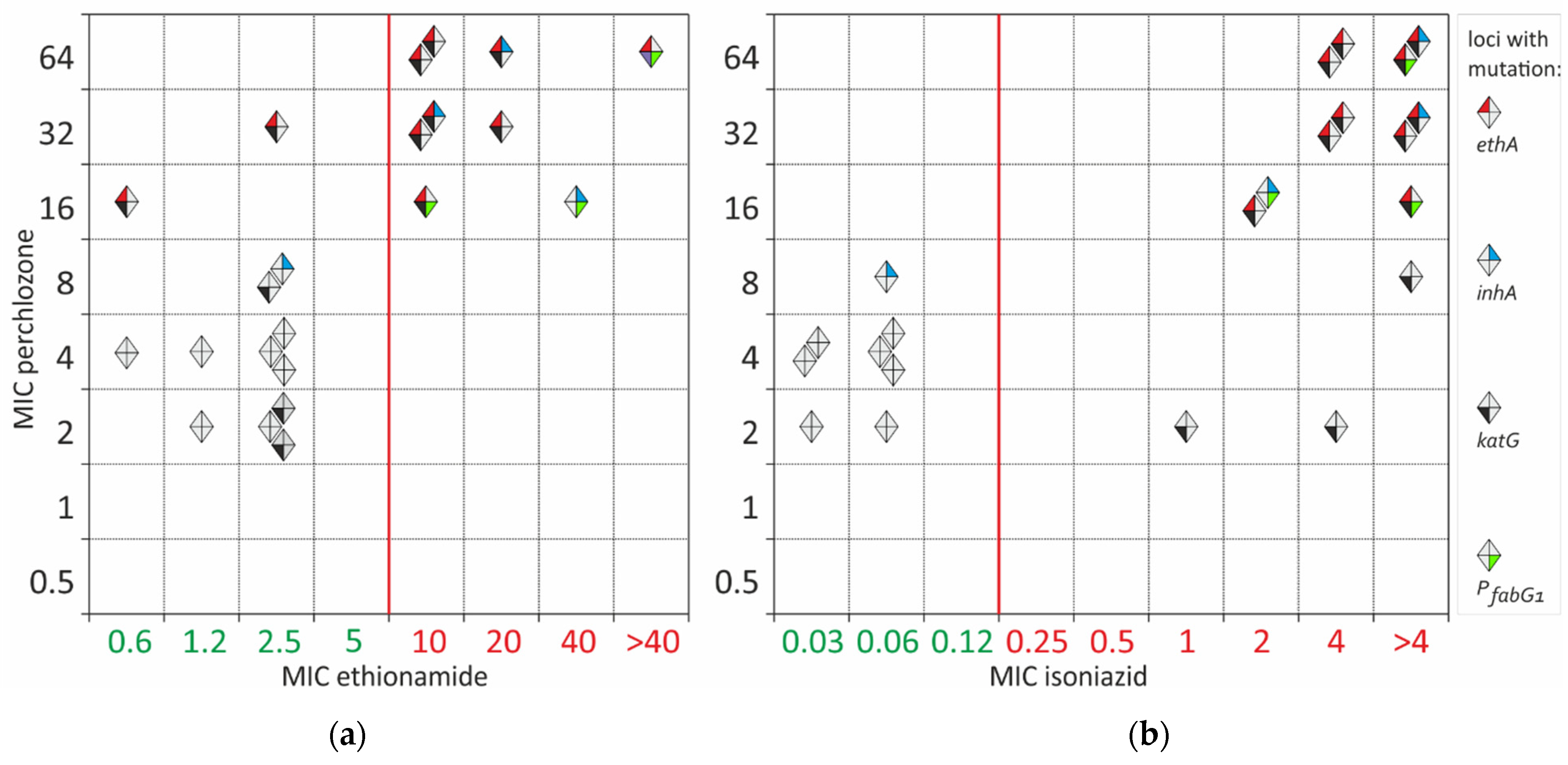
2. Results
3. Discussion
4. Materials and Methods
4.1. Mycobacterium tuberculosis Strains
4.2. DNA Isolation and Sequencing
4.3. MIRU-VNTR Typing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Domagk, G. Investigations on the Antituberculous Activity of the Thiosemicarbazones In Vitro and In Vivo. Am. Rev. Tuberc. 1950, 61, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Harland, R.D. Stevens-Johnson Syndrome with Unusual Skin Features Occurring in Two Patients Undergoing Treatment for Pulmonary Tuberculosis with Thiacetazone. Tubercle 1962, 43, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Jaju, M.; Jaju, M.; Ahuja, Y.R. Combined and Individual Effects of Isoniazid and Thiacetazone on Human Lymphocyte Chromosomes In Vitro and In Vivo. Hum. Toxicol. 1984, 3, 373–382. [Google Scholar] [CrossRef]
- Fox, H.H. Synthetic Tuberculostats. III. Isonicotinaldehyde Thiosemicarbazone And Some Related Compounds. J. Org. Chem. 1952, 17, 555–562. [Google Scholar] [CrossRef]
- Fox, H.H. The Chemical Attack on Tuberculosis. Trans. N. Y. Acad. Sci. 1953, 15, 234–242. [Google Scholar] [CrossRef]
- Stadler, E.T.; Steiner, M. Isoniazid Treatment for Tuberculosis. N. Engl. J. Med. 1971, 284, 730–731. [Google Scholar] [CrossRef]
- Parekh, M.J.; Schluger, N.W. Treatment of Latent Tuberculosis Infection. Ther. Adv. Respir. Dis. 2013, 7, 351–356. [Google Scholar] [CrossRef]
- Barry, C.E.; Slayden, R.A.; Sampson, A.E.; Lee, R.E. Use of Genomics and Combinatorial Chemistry in the Development of New Antimycobacterial Drugs. Biochem. Pharmacol. 2000, 59, 221–231. [Google Scholar] [CrossRef]
- Scardigli, A.; Caminero, J.A.; Sotgiu, G.; Centis, R.; D’Ambrosio, L.; Migliori, G.B. Efficacy and Tolerability of Ethionamide versus Prothionamide: A Systematic Review. Eur. Respir. J. 2016, 48, 946–952. [Google Scholar] [CrossRef]
- Rist, N.; Grumbach, F.; Libermann, D. Experiments on the Antituberculous Activity of Alpha-Ethylthioisonicotinamide. Am. Rev. Tuberc. Pulm. Dis. 1959, 79, 1–5. [Google Scholar] [CrossRef]
- Ministry of Health of the Russian Federation. Russian National Guidelines on Tuberculosis Treatment. Available online: https://cr.minzdrav.gov.ru/recomend/16_2 (accessed on 28 February 2023).
- Ministry of Health of the Russian Federation. Russian National Catalog of Registered Drugs. Available online: http://zdravmedinform.ru/grls/reg-lp-001899.html (accessed on 28 February 2023).
- Vilchèze, C.; Jacobs, W.R. Resistance to Isoniazid and Ethionamide in Mycobacterium Tuberculosis: Genes, Mutations, and Causalities. Microbiol. Spectr. 2014, 2, MGM2-0014-2013. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Dubnau, E.; Quemard, A.; Balasubramanian, V.; Um, K.S.; Wilson, T.; Collins, D.; de Lisle, G.; Jacobs, W.R. InhA, a Gene Encoding a Target for Isoniazid and Ethionamide in Mycobacterium Tuberculosis. Science 1994, 263, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Belardinelli, J.M.; Morbidoni, H.R. Mutations in the Essential FAS II β-Hydroxyacyl ACP Dehydratase Complex Confer Resistance to Thiacetazone in Mycobacterium Tuberculosis and Mycobacterium Kansasii. Mol. Microbiol. 2012, 86, 568–579. [Google Scholar] [CrossRef]
- Gopal, P.; Dick, T. The New Tuberculosis Drug Perchlozone® Shows Cross-Resistance with Thiacetazone. Int. J. Antimicrob. Agents 2015, 45, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Alahari, A.; Trivelli, X.; Guérardel, Y.; Dover, L.G.; Besra, G.S.; Sacchettini, J.C.; Reynolds, R.C.; Coxon, G.D.; Kremer, L. Thiacetazone, an Antitubercular Drug That Inhibits Cyclopropanation of Cell Wall Mycolic Acids in Mycobacteria. PLoS ONE 2007, 2, e1343. [Google Scholar] [CrossRef]
- Glickman, M.S. The MmaA2 Gene of Mycobacterium Tuberculosis Encodes the Distal Cyclopropane Synthase of the Alpha-Mycolic Acid. J. Biol. Chem. 2003, 278, 7844–7849. [Google Scholar] [CrossRef]
- DeBarber, A.E.; Mdluli, K.; Bosman, M.; Bekker, L.G.; Barry, C.E. Ethionamide Activation and Sensitivity in Multidrug-Resistant Mycobacterium Tuberculosis. Proc. Natl. Acad. Sci. USA 2000, 97, 9677–9682. [Google Scholar] [CrossRef]
- Baulard, A.R.; Betts, J.C.; Engohang-Ndong, J.; Quan, S.; McAdam, R.A.; Brennan, P.J.; Locht, C.; Besra, G.S. Activation of the Pro-Drug Ethionamide Is Regulated in Mycobacteria. J. Biol. Chem. 2000, 275, 28326–28331. [Google Scholar] [CrossRef]
- Vannelli, T.A.; Dykman, A.; Ortiz de Montellano, P.R. The Antituberculosis Drug Ethionamide Is Activated by a Flavoprotein Monooxygenase. J. Biol. Chem. 2002, 277, 12824–12829. [Google Scholar] [CrossRef]
- Laborde, J.; Deraeve, C.; Duhayon, C.; Pratviel, G.; Bernardes-Génisson, V. Ethionamide Biomimetic Activation and an Unprecedented Mechanism for Its Conversion into Active and Non-Active Metabolites. Org. Biomol. Chem. 2016, 14, 8848–8858. [Google Scholar] [CrossRef]
- Coxon, G.D.; Craig, D.; Corrales, R.M.; Vialla, E.; Gannoun-Zaki, L.; Kremer, L. Synthesis, Antitubercular Activity and Mechanism of Resistance of Highly Effective Thiacetazone Analogues. PLoS ONE 2013, 8, e53162. [Google Scholar] [CrossRef]
- Alahari, A.; Alibaud, L.; Trivelli, X.; Gupta, R.; Lamichhane, G.; Reynolds, R.C.; Bishai, W.R.; Guerardel, Y.; Kremer, L. Mycolic Acid Methyltransferase, MmaA4, Is Necessary for Thiacetazone Susceptibility in Mycobacterium Tuberculosis. Mol. Microbiol. 2009, 71, 1263–1277. [Google Scholar] [CrossRef]
- Belardinelli, J.M.; Morbidoni, H.R. Recycling and Refurbishing Old Antitubercular Drugs: The Encouraging Case of Inhibitors of Mycolic Acid Biosynthesis. Expert Rev. Anti Infect. Ther. 2013, 11, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Rueda, J.; Realpe, T.; Mejia, G.I.; Zapata, E.; Rozo, J.C.; Ferro, B.E.; Robledo, J. Genotypic Analysis of Genes Associated with Independent Resistance and Cross-Resistance to Isoniazid and Ethionamide in Mycobacterium Tuberculosis Clinical Isolates. Antimicrob. Agents Chemother. 2015, 59, 7805–7810. [Google Scholar] [CrossRef] [PubMed]
- Malík, I.; Čižmárik, J.; Pecháčová, M. Focus on Perchlozone, an Anti-Tuberculosis Drug from the Russian Federation. Ceska Slov. Farm. 2020, 69, 203–210. [Google Scholar]
- Merker, M.; Blin, C.; Mona, S.; Duforet-Frebourg, N.; Lecher, S.; Willery, E.; Blum, M.G.B.; Rüsch-Gerdes, S.; Mokrousov, I.; Aleksic, E.; et al. Evolutionary History and Global Spread of the Mycobacterium Tuberculosis Beijing Lineage. Nat. Genet. 2015, 47, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Hartkoorn, R.C.; Uplekar, S.; Cole, S.T. Cross-Resistance between Clofazimine and Bedaquiline through Upregulation of MmpL5 in Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 2979–2981. [Google Scholar] [CrossRef] [PubMed]
- Poulton, N.C.; Azadian, Z.A.; DeJesus, M.A.; Rock, J.M. Mutations in Rv0678 Confer Low-Level Resistance to Benzothiazinone DprE1 Inhibitors in Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2022, 66, e0090422. [Google Scholar] [CrossRef] [PubMed]
- Yablonskiy, P.K.; Vinogradova, T.I.; Levashev, Y.N.; Pavlova, M.V.; Zilber, E.K.; Starshinova, A.A.; Sapozhnikova, N.V.; Chernokhaeva, I.V.; Archakova, L.I.; Zabolotnykh, N.V.; et al. Preclinical and Clinical Trials of the New Tuberculosis Drug Perchlozon. Ter. Arkhiv 2016, 88, 111. [Google Scholar] [CrossRef]
- WHO. Technical Report on Critical Concentrations for Drug Susceptibility Testing of Medicines Used in the Treatment of Drug-Resistant Tuberculosis; World Health Organization: Geneva, Switzerland, 2018; p. 106.
- Mokrousov, I.; Vyazovaya, A.; Akhmedova, G.; Solovieva, N.; Turkin, E.; Zhuravlev, V. Genetic Variation Putatively Associated with Mycobacterium Tuberculosis Resistance to Perchlozone, a New Thiosemicarbazone: Clues from Whole Genome Sequencing and Implications for Treatment of Multidrug-Resistant Tuberculosis. Antibiotics 2020, 9, 669. [Google Scholar] [CrossRef]
- Morlock, G.P.; Metchock, B.; Sikes, D.; Crawford, J.T.; Cooksey, R.C. EthA, InhA, and KatG Loci of Ethionamide-Resistant Clinical Mycobacterium Tuberculosis Isolates. Antimicrob. Agents Chemother. 2003, 47, 3799–3805. [Google Scholar] [CrossRef] [PubMed]
- Ushtanit, A.; Kulagina, E.; Mikhailova, Y.; Makarova, M.; Safonova, S.; Zimenkov, D. Molecular Determinants of Ethionamide Resistance in Clinical Isolates of Mycobacterium Tuberculosis. Antibiotics 2022, 11, 133. [Google Scholar] [CrossRef]
- Sharma, M.; Thibert, L.; Chedore, P.; Shandro, C.; Jamieson, F.; Tyrrell, G.; Christianson, S.; Soualhine, H.; Wolfe, J. Canadian Multicenter Laboratory Study for Standardized Second-Line Antimicrobial Susceptibility Testing of Mycobacterium Tuberculosis. J. Clin. Microbiol. 2011, 49, 4112–4116. [Google Scholar] [CrossRef]
- Barrera, L.; Cooreman, E.; de Dieu Iragena, J.; Drobniewski, F.; Duda, P.; Havelkova, M.; Hoffner, S.; Kam, K.M.; Kim, S.J.; Labelle, S.; et al. Policy Guidance on Drug-Susceptibility Testing (DST) of Second-Line Antituberculosis Drugs; WHO Guidelines Approved by the Guidelines Review Committee; World Health Organization: Geneva, Switzerland, 2008.
- Nosova, E.Y.; Zimenkov, D.V.; Khakhalina, A.A.; Isakova, A.I.; Krylova, L.Y.; Makarova, M.V.; Galkina, K.Y.; Krasnova, M.A.; Safonova, S.G.; Litvinov, V.I.; et al. A Comparison of the Sensititre MycoTB Plate, the Bactec MGIT 960, and a Microarray-Based Molecular Assay for the Detection of Drug Resistance in Clinical Mycobacterium Tuberculosis Isolates in Moscow, Russia. PLoS ONE 2016, 11, e0167093. [Google Scholar] [CrossRef] [PubMed]
- Supply, P.; Allix, C.; Lesjean, S.; Cardoso-Oelemann, M.; Rüsch-Gerdes, S.; Willery, E.; Savine, E.; de Haas, P.; van Deutekom, H.; Roring, S.; et al. Proposal for Standardization of Optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat Typing of Mycobacterium Tuberculosis. J. Clin. Microbiol. 2006, 44, 4498–4510. [Google Scholar] [CrossRef] [PubMed]
- Allix-Béguec, C.; Harmsen, D.; Weniger, T.; Supply, P.; Niemann, S. Evaluation and Strategy for Use of MIRU-VNTRplus, a Multifunctional Database for Online Analysis of Genotyping Data and Phylogenetic Identification of Mycobacterium Tuberculosis Complex Isolates. J. Clin. Microbiol. 2008, 46, 2692–2699. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ushtanit, A.; Mikhailova, Y.; Krylova, L.; Grigorash, D.; Makarova, M.; Safonova, S.; Zimenkov, D. Perchlozone Resistance in Clinical Isolates of Mycobacterium tuberculosis. Antibiotics 2023, 12, 590. https://doi.org/10.3390/antibiotics12030590
Ushtanit A, Mikhailova Y, Krylova L, Grigorash D, Makarova M, Safonova S, Zimenkov D. Perchlozone Resistance in Clinical Isolates of Mycobacterium tuberculosis. Antibiotics. 2023; 12(3):590. https://doi.org/10.3390/antibiotics12030590
Chicago/Turabian StyleUshtanit, Anastasia, Yulia Mikhailova, Ludmila Krylova, Dmitry Grigorash, Marina Makarova, Svetlana Safonova, and Danila Zimenkov. 2023. "Perchlozone Resistance in Clinical Isolates of Mycobacterium tuberculosis" Antibiotics 12, no. 3: 590. https://doi.org/10.3390/antibiotics12030590
APA StyleUshtanit, A., Mikhailova, Y., Krylova, L., Grigorash, D., Makarova, M., Safonova, S., & Zimenkov, D. (2023). Perchlozone Resistance in Clinical Isolates of Mycobacterium tuberculosis. Antibiotics, 12(3), 590. https://doi.org/10.3390/antibiotics12030590