Abstract
Bacterial exposure to antibiotic concentrations below the minimum inhibitory concentration (MIC) may result in a selection window allowing for the rapid evolution of resistance. These sub-MIC concentrations are commonly found in soils and water supplies in the greater environment. This study aimed to evaluate the adaptive genetic changes in Klebsiella pneumoniae 43816 after prolonged but increasing sub-MIC levels of the common antibiotic cephalothin over a fourteen-day period. Over the course of the experiment, antibiotic concentrations increased from 0.5 μg/mL to 7.5 μg/mL. At the end of this extended exposure, the final adapted bacterial culture exhibited clinical resistance to both cephalothin and tetracycline, altered cellular and colony morphology, and a highly mucoid phenotype. Cephalothin resistance exceeded 125 μg/mL without the acquisition of beta-lactamase genes. Whole genome sequencing identified a series of genetic changes that could be mapped over the fourteen-day exposure period to the onset of antibiotic resistance. Specifically, mutations in the rpoB subunit of RNA Polymerase, the tetR/acrR regulator, and the wcaJ sugar transferase each fix at specific timepoints in the exposure regimen where the MIC susceptibility dramatically increased. These mutations indicate that alterations in the secretion of colanic acid and attachment of colonic acid to LPS may contribute to the resistant phenotype. These data demonstrate that very low sub-MIC concentrations of antibiotics can have dramatic impacts on the bacterial evolution of resistance. Additionally, this study demonstrates that beta-lactam resistance can be achieved through sequential accumulation of specific mutations without the acquisition of a beta-lactamase gene.
1. Introduction
The mis- and overuse of antibiotics by the medical and agricultural industries has created a basal level of antibiotic exposure present in our environment [1,2,3,4,5,6]. Antibiotics may be introduced into the environment via three major pathways. The first is from urine and excretions of people, pets, and livestock. Between 40 and 90% of administered antibiotics are excreted through urine and feces, with the molecule still in its active form [2]. Second, antibiotics such as chlorhexidine are used in commercial agriculture and aquaculture [4]. These substances can then contaminate nearby lands via wastewater and groundwater seepage [2,7]. Finally, antibiotics can enter the local environment through improper disposal of unused or expired prescriptions [2,4]. While concentrations of antimicrobial substances in the environment will vary based on location and potential sources of contamination, it is generally agreed that the residual environmental antibiotic concentration is not high enough to eradicate native bacterial populations. However, even at low concentrations, these compounds become a source of survival stress for bacteria in the soil and water supplies. Concentrations below the bacteriostatic limit, referred to as sub-MIC levels, may result in the creation of a selection window for bacteria [8,9,10]. This window is the concentration at which the occurrence of genomic mutations is highest and can lead to the development of clinical antibiotic resistance in pathogens.
Experiments have shown that sub-MIC antibiotic treatment can alter the resistance profile, nutrient use, protein expression, gene transcription, and mutation rates among common nosocomial pathogens [5,11,12]. However, current studies of sub-MIC exposure and bacterial adaptation have a number of limitations. First, these experiments frequently utilize strains of bacteria either known to harbor antibiotic-resistance genes or are otherwise clinically resistant to one specific class of antibiotic. Second, the experimental designs include only brief exposure times to antibiotics. In most studies, the samples were exposed to the antibiotic for only 24–48 h, providing limited time for the bacteria to evolve novel genetic changes such as those seen in a clinical or environmental setting where antibiotic exposure is both consistent and long-term [5,11,12]. Finally, genetic analyses frequently used pre-determined targets or the phenotypes assumed from mutations in pre-determined targets. While the results of these studies are informative, they are not comparable to situations in which antibiotic resistance phenotypes evolve over time through prolonged sub-lethal antibiotic exposure.
In this study, we utilized Klebsiella pneumoniae 43816, which does not exhibit clinical resistance to the major antibiotic classes. K. pneumoniae was cultured using a progressive exposure method to gradually increased sub-MIC concentrations of the beta-lactam antibiotic cephalothin over a fourteen-day period. At the end of this prolonged exposure period to sub-MIC concentrations, the K. pneumoniae exhibited clinical resistance to both cephalothin and tetracycline, altered cellular and colony morphology, and a highly mucoid phenotype. Whole genome sequencing identified a series of genetic changes that could be mapped over the fourteen-day exposure period to the onset of antibiotic resistance. Specifically, mutations in the rpoB subunit of RNA Polymerase, the tetR/acrR regulator, and the wcaJ sugar transferase each fix at specific points in the exposure regimen where the MIC susceptibility dramatically increased. These mutations indicate that the secretion and export of colanic acid, and its association with lipopolysaccharides, may contribute to the resistant phenotype. These data demonstrate that very low sub-MIC concentrations of antibiotics can have dramatic impacts on the bacterial evolution of resistance. Additionally, this study demonstrates that antibiotic resistance can be achieved through the accumulation of mutations without the traditional acquisition of beta-lactamase or other drug-inactivating genes.
2. Results
2.1. Experimental Design of Progressive Sub-MIC Exposure to Cephalothin
This study aimed to evaluate the adaptive genetic changes in Klebsiella pneumoniae 43816 upon prolonged but increasing exposure to sub-MIC levels of the common antibiotic cephalothin. Klebsiella pneumoniae 43816 was utilized because Klebsiella is known for its genomic plasticity and rapid evolution of antibiotic resistance [13,14]. Additionally, the genome of this strain has been sequenced [15], which allows for genetic changes to be tracked over the course of the progressive exposure.
The experimental design of this exposure protocol is diagramed in Figure 1. Fresh cultures were grown in LB at 37 °C with 200 rpm shaking and an initial cephalothin concentration of 0.5 μg/mL. After the first 12 h of culture, a sample was transferred to a fresh LB with the same antibiotic concentration and allowed to grow for a second 12 h period. The bacteria were exposed to any one dose of antibiotics for 24 h in total. Then, the sample was transferred to a culture with an increased concentration of antibiotics. At each step, the antibiotic concentration was increased by 0.5 μg/mL to a final exposure of 7.5 μg/mL, which was well below the CLSI standard of 16 μg/mL for cephalothin resistance [16]. As a control, an untreated culture, without antibiotics, was grown and transferred in parallel to LB without any antibiotic. Frozen stocks were made of all cultures at each 12 h timepoint and saved for later analysis.

Figure 1.
Experimental design. Klebsiella pneumoniae 43816 was exposed to increasing concentrations of cephalothin in LB media over a continuous 14-day period. All cultures were grown in LB media at 37 °C with 200 rpm shaking. Each single-dose exposure lasted 24 h and included a sub-culture into fresh media with antibiotics at the 12 h mark. Frozen bacterial stocks were made every 12 h for subsequent analysis. A control culture was also grown, sub-cultured, and sampled in LB media without antibiotics, following the same schedule as the treated sample.
2.2. Progressive Antibiotic Exposure Alters K. pneumoniae Growth, Cellular Morphology, and Colony Morphology
In order to evaluate the impact of such low antibiotic concentrations, we measured the rate of growth of the culture at the 12 h transfer mark. This was determined by measuring the time it took for the culture to reach an optical density (OD600) of 1.0 after subculturing. This point was chosen to evaluate the impact of low-dose antibiotics on the ability of K. pneumoniae to exit the lag phase into true exponential growth. As seen in Table 1, the cultures exposed to antibiotics extended the time required for cultures to enter the exponential phase (time to OD 1.0), which is indicative of cellular stress. The specific growth rate of the treated culture was substantially slowed once antibiotic concentrations exceeded 5 μg/mL, while the untreated culture was not significantly affected over the entire course of the experiment.

Table 1.
Changes in specific growth rates because of low-dose antibiotic exposure. The initial impact of sub-MIC antibiotic exposure was determined by evaluating the specific growth rate of cultures upon transfer to a new antibiotic concentration. Measurement of time required for adapted and untreated K. pneumoniae 43816 sub-culture to reach an OD600 of 1.0 was measured after 12 h exposure at each time point/dose. Changes between treated and control were determined relative to the average time for all untreated cultures (142 min). Values in bold represent increases greater than one-half that of the untreated culture.
This slow entry into exponential growth was not a permanent effect, as demonstrated by the growth properties of the final adapted culture (Table 1 and Figure 2A). The final adapted culture was able to grow rapidly in 7.5 μg/mL cephalothin with both a time to OD600 1.0 and an overall growth curve highly similar to that of untreated and original K. pneumoniae cultures.
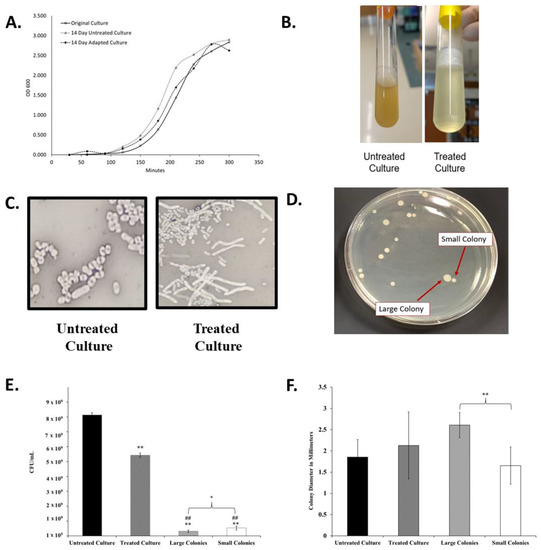
Figure 2.
Changes in liquid culture, cell, and colony morphology because of antibiotic exposure. (A) Growth Curves of original K. pneumoniae 43816 and cultures after the 14-day regiment with antibiotics (adapted culture) and with no antibiotics (untreated culture). (B) Broth culture appearance of untreated and antibiotic-exposed culture at the endpoint of the 14-day experiment (after 12 h of growth) exhibits increased mucoid layer in antibiotic-treated culture. (C) Cell morphology of untreated culture and antibiotic-exposed culture was determined by negative staining with nigrosin at 1000× magnification. Antibiotic exposed cells exhibit an elongated cell morphology. (D) Quadrant steaking on LB of the final adapted culture revealed two colony morphologies. (E) Colony Forming units from each culture at an OD600 of 1.0. n = 3 independent replicates per culture; bars indicate standard deviation. ** p < 0.001 compared to untreated culture, ## p < 0.001 compared to treated culture, * p < 0.05 between large and small colonies. (F) The diameter of colonies from each culture as measured using Image J. n = 20 colonies for each culture; bars indicate standard deviation. ** p < 0.001 between designated colony types.
While the optical density growth properties of the adapted culture returned to a range similar to the untreated culture, the overall appearance of the antibiotic-treated culture was significantly and permanently altered after the progressive exposure experiment. The antibiotic-adapted culture exhibited a highly mucoid appearance in liquid culture when compared to the clonally related untreated group (Figure 2B). While the viscosity changed, the adapted population did not qualify as a hypermucoviscous strain, as indicated by a string test.
Viscosity may also be a result of increased cell lysis in the culture. To further investigate this, we determined the CFU/mL of each culture at an OD600 of 1.0. As seen in Figure 2E, the final treated culture achieves this optical density measurement with significantly lower cell density than the original culture.
Differences were also observed in cellular and colony morphology. The antibiotic-exposed culture resulted in cells with a greatly elongated, filamentous structure (Figure 2C). Quadrant streaking of the final antibiotic-adapted culture resulted in two consistent colony morphologies, designated small and large colonies (Figure 2D). Bacteria were isolated from either the large or small colonies and re-suspended in liquid culture. When replated separately, they only produced colonies of a statistically significantly distinct size (Figure 2F) that corresponded to the morphology of the original colony. The isolated colony morphologies also exhibited significantly lower CFU/mL counts than the adapted culture that contained both types as a mixture (Figure 2E). The persistence of these phenotypes after isolation indicated that these morphological changes were heritable.
2.3. Progressive Antibiotic Exposure Induces Clinical Resistance to Multiple Classes of Antibiotics
Changes in the minimum inhibitory concentration (MIC) of several different classes of antibiotics were determined by the broth microdilution assay (Table 2). The concentration of antibiotic that halts all growth (MIC breakpoint) was determined for tetracycline, amikacin, and five different beta-lactam antibiotics (Cephalothin, Cefoxitin, Cefotaxime, Cefepime, and Imipenem). The beta-lactam drugs were chosen as representative of different generations of cephalosporin antibiotics and a carbapenem, respectively. Clinical resistance was determined using CLSI breakpoints [16].
Table 2.
Changes in minimum inhibitory concentration (MIC) breakpoints of K. pneumoniae 43816 cultures after progressive low-dose exposures. Broth microdilution assays (n = 3) determined the minimum inhibitory concentration breakpoint in μg/mL of multiple antibiotics for endpoint cultures, the ΔwcaJ strain, and endpoint colony morphology isolates.
As seen in Table 2, the fourteen-day growth of cells without antibiotics did not greatly impact the MIC values against the antibiotic tested. The MIC values in the fourteen-day untreated cells were not as substantially elevated as the MIC values observed in the final adapted cells when compared to the day 0 control (Table 2). Overall, the untreated cells did not reach MICs above the CLSI cutoff for clinical resistance for any of the antibiotics tested, unlike the fourteen-day adapted cells. The adapted culture exhibited highly elevated MICs to cephalothin, which was used for the progressive exposure experiment. This culture exhibited clinical resistance to cephalothin and cefoxitin as determined by CLSI breakpoints. Cephalothin and cefoxitin are first- and second-generation cephalosporin antibiotics, respectively. MICs to later-generation cephalosporins and the carbapenem antibiotic were slightly elevated in both the fourteen-day untreated and adapted cells, but the MICs for each were still below the CLSI cutoff. To fully evaluate the impact of progressive sub-MIC cephalothin exposure, antibiotics with different cellular targets than the beta-lactams were also tested. Progressive exposure had little impact on the breakpoint value for the aminoglycoside antibiotic amikacin, but clinical resistance to tetracycline was achieved. These data indicate that progressive exposure to cephalothin at only half the MIC cutoff concentration [16] led to resistance to a second generation of cephalosporin in addition to an antibiotic in another class.
Additionally, MIC breakpoints were determined for the two isolated colony morphologies from the final antibiotic-adapted culture. Both the small and large morphologies exhibited clinical resistance to cephalothin and cefoxitin, with the large colonies exhibiting highly elevated MICs to cephalothin; which was well above the MIC observed for both the small and adapted final culture. Both isolates also exhibited elevated MICs to tetracycline but were more sensitive than the mixed day 14 adapted culture. Amikacin MICs were not significantly changed in either phenotype. Taken together, these data demonstrate that progressive sub-MIC concentrations of a single antibiotic provide sufficient evolutionary pressure to drive the evolution of clinical levels of resistance to multiple generations of beta-lactams as well as classes of antibiotics with different cellular targets.
2.4. Whole Genome Sequencing Reveals Distinctive Genetic Changes Because of Progressive Antibiotic Exposure
To determine the genetic changes responsible for this antibiotic-resistant mucoid phenotype, DNA was extracted from K. pneumoniae stocks on each day of the progressive antibiotic exposure and sequenced by the Microbial Genome Sequencing Center using an Illumina sequencing method and a proprietary statistical algorithm for variant calling (www.seqcenter.com, accessed on 11 April 2023) [17]. This methodology resulted in a surprisingly low number of SNPs identified, in part because it does not analyze changes to non-coding regions of the genome, such as promoters.
To confirm these results and to extend our analysis to include SNPs within promoter regions, DNA from the fourteen-day adapted, and untreated K. pneumoniae 43816 cultures were also analyzed using SMRT Cell Sequencing by the National Center for Genome Resources (www.ncgr.org, accessed on 11 April 2023). Together these analyses identified 107 mutations in the control culture and 29 mutations in the genome of the antibiotic-treated culture. Of the 29 mutations identified in the adapted strain, 15 were non-synonymous changes that occurred in protein-coding regions of the sequence. To further refine the list of potential targets, any mutations shared with the untreated population of K. pneumoniae were removed. Comparing the genomes highlighted seven protein-coding genes altered in the final antibiotic-adapted cells (Table 3). Changes to rpoB, tetR/acrR, wcaJ, and gndA were identified using both sequencing methodologies (Table 4).
Table 3.
Non-synonymous single nucleotide polymorphisms and genomic changes were identified in the final adapted culture. SMRT Cell genomic sequencing was used for whole genome sequencing. The genome sequence of K. pneumoniae 43816 was used as the reference genome.
Table 4.
SNPs and genomic changes identified in large and small colony subpopulations of 14-day adapted culture. Illumina genomic sequencing was used for the sequencing of colony subpopulations. The genome sequence of K. pneumoniae 43816 was used as the reference genome.
Notably, none of the identified mutations by either methodology were identified in penicillin-binding proteins, porins, or other cell wall modification genes highly associated with traditional beta-lactam resistance. Genes with identified mutations were searched against the CARD data base and using the amrFinder bioinformatics tool (card.mcmaster.ca, accessed on 11 April 2023) [18,19]. Neither of these analyses identified these genes as being previously identified as resistance mechanisms to beta-lactams. The tetR/acrR gene resulted in a match for the wildtype version of the gene as being associated with tetracycline resistance, while rpoB matched for rifamycin resistance [20]. This analysis indicates that the majority of these modifications and gene targets are novel or not previously characterized in terms of their impact on antibiotic resistance.
The only mutations that were not 100% fixed in the population at the conclusion of the fourteen-day progressive exposure were nucleotide substitutions in the coding region of the SGNH/GDSL hydrolase family protein and a deletion in the promoter of SGNH/GDSL (Table 3). This may be linked to the emergence of small and large colony variants. Other genetic changes that were fixed by the fourteen-day endpoint include deletions in the coding regions of the following: N-acetyltransferase, ABC transporter, ATP binding protein, and DNA recombination protein (RmuC). Additionally, a substitution in the coding region of undecaprenyl–phosphate glucose phosphotransferase was discovered. An insertion was found in a globin coding sequence, and a large insert in the tetR/acrR transcriptional regulator resulted in an early stop codon. The functions of these genes encompassed cellular processes of signal transduction, energy and metabolite use, capsule formation, and nucleic acid proofreading.
In addition to changes in the coding regions of those genes, six unique mutations were identified in promoter regions of five other genes that were fixed in the adapted population (Table 3). Promoter regions were defined as occurring within 150 bp of a protein-coding start site. Synonymous mutations between the untreated and adapted cultures were removed, as described above. With the exception of SGNH/GDSL, all other genetic changes in promoter regions were fully fixed in 100% of the DNA sampled. Not all genes associated with these promoters have been fully characterized in Klebsiella pneumoniae. In those cases, the data about the class of each gene and close homologues are presented. Functions of the coding regions encompassed cell metabolism, signal transduction, and mRNA proofreading.
2.5. Genome Sequencing Allows for Correlation of Genetic Changes with Increased MICs
The Illumina sequencing of the cultures from each day of the experiment allowed us to map the timepoint of 100% fixation of mutation with significant increases in MIC and changes in MIC. For this, we focused on those mutated genes (rpoB, tetR/acrR, wcaJ, and gndA) identified using both sequencing methodologies (Table 4).
As seen in Figure 3, the emergence of clinical tetracycline resistance can be matched with fully fixed alterations in rpoB, and large increases in the cephalothin MICs can be correlated with fixed changes to tetR/acrR and wcaJ. The large increase in cephalothin MIC associated with wcaJ can also be mapped to the significant slowing of the growth of all treated cultures when exposed to 5 μg/mL of the drug (Table 1).
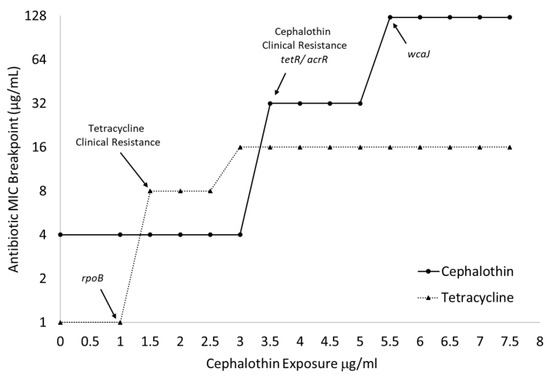
Figure 3.
MIC breakpoints map to the onset of clinical resistance and fixation of genomic changes. Broth microdilution assays (n = 3) determined MIC breakpoints to tetracycline and cephalothin of cultures over the entire 14-day progressive exposure. Arrows indicate jumps above clinical resistance breakpoint and fixation of specific genetic changes as identified by WGS.
Two separate sets of sequence changes in wcaJ and gndA were identified using the Illumina methodology (Table 4). The final adapted culture, which is composed of a mix of large and small colony morphologies, identified sequence changes with only 64% genome coverage. When the two morphologies were sequenced separately, a large deletion that impacted both genes was found in only the small colony variant.
Both the large and small colony variants had mutations in the rpoB and tetR/acrR transcriptional regulator sequences. The mutation in rpoB was 100% fixed in both variants. However, the depth of coverage of the tetR/acrR transcriptional regulator in the large colony variant was only 52%. The large colony variant also had a distinguishing marginal mutation call in a comEC family protein that was not found in the small colony variant. The small colony variant exhibited an additional SNP in a yfiR family protein that was not identified in the large colony variant. All the mutations in the small colony-forming variant were 100% fixed, indicating a more homogenous genomic identity than the large colony-forming subpopulation.
2.6. Genetic Changes in Progressive Antibiotic Adapted Cultures Are Associated with Alterations to Capsule and LPS
The final adapted culture from this progressive antibiotic exposure experiment exhibited a highly mucoid phenotype indicating that capsule production or composition may be altered. Genome sequencing then identified alterations to wcaJ, which is part of the capsule cps operon and initiates the production of colanic acid [21]. Therefore, we hypothesized that alterations to capsule production and composition might be directly related to the emergence of high-concentration cephalothin resistance.
To investigate changes in the capsule, extracellular polysaccharide was extracted and quantified by the uronic acid assay. As can be seen in Figure 4A, capsule polysaccharide increases until the fixation of the tetR/acrR mutation and the emergence of clinical cephalothin resistance. All samples after this point exhibited capsule polysaccharides at levels similar to or slightly below those produced by the untreated control culture.
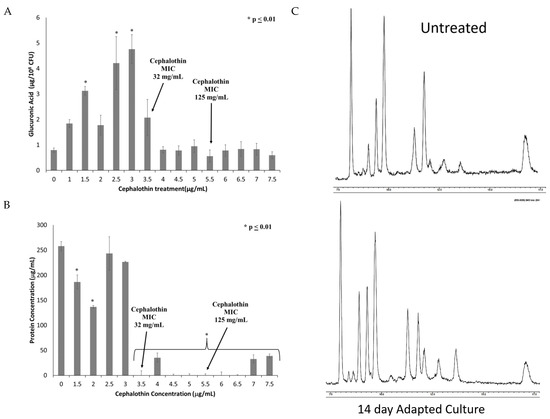
Figure 4.
Onset of clinical resistance coincides with changes in quantity and composition of capsule. Capsule polysaccharides were extracted from cultures from each treatment of the experiment. The extracted capsule was then analyzed for (A) Total capsule production as quantified by glucuronic acid; and (B) Protein content of extracted capsule quantified by BCA protein assay. n = 6 for both assays, * p ≤ 0.01 by one-way ANOVA compared to untreated control. (C) GC-MS analysis of the carbohydrate composition of capsules extracted from untreated and adapted cultures.
Given that the uronic acid assay only detects one sugar moiety within the capsule polysaccharide, other possible components of the capsule were investigated. Extracted capsules from untreated and adapted cultures were found to contain similar levels of sialic acids (2.5 ± 1.2 nmoles/108 CFU in untreated culture vs. 2.9 ± 1.8 nmoles/108 CFU in final adapted culture), which is associated with increased virulence [22]. These results indicate that sialic acid is not significantly altered by these mutations. Adapted capsule extracts also exhibited reduced total protein content as determined by BCA assay (Figure 4B). Both the reduced glucuronic acid and protein emerged concurrently with the alterations in the tetR/acrR gene. Finally, the capsule extract was analyzed by GC/MS to identify new peaks indicative of changes to the sugar composition (Figure 4C). No new peaks were identified, indicating that novel carbohydrate changes are not directly associated with the antibiotic-resistant phenotype.
To determine the relationship between alterations in capsule and antibiotic resistance, we tested a ΔwcaJ strain of Klebsiella pneumoniae 43816 for altered MICs. This deletion has been previously characterized as significantly reducing capsule uronic acid content and mucoviscosity [23]. As seen in Table 2, deletion of this gene does result in increases in MIC breakpoints to both cephalothin and cefoxitin. These increases are to the level of the breakpoint value. This data indicates that cps modification can have a significant impact on antibiotic susceptibility in Klebsiella. However, the high-level clinical resistance seen in our fully adapted strain is likely due to the combination of multiple gene alterations, including those within the cps operon.
Colanic acid, which is synthesized in part using the wcaJ gene product, is also associated with lipopolysaccharide. Therefore, we determined the LPS content of both K. pneumoniae cells and extracted the capsule using the Purpald assay. LPS content was significantly elevated in both the cell and extracted capsule of the adapted strain compared to the control (Figure 5). This elevated LPS content may indicate a high level of outer membrane turnover in the adapted culture associated with alteration in wcaJ.

Figure 5.
Lipopolysaccharide content of capsule and cells is increased after progressive sub-MIC exposure. Capsule polysaccharides were extracted from untreated and final adapted cultures. Capsule and whole cells’ LPS content was determined by the Purpald Assay. n = 6, * p < 0.001 by one-way ANOVA compared to untreated control.
3. Discussion
The goal of this study was to characterize the impact of low-concentration antibiotic exposure on the evolution of bacterial resistance. A progressive exposure model was used, in which Klebsiella pneumoniae was exposed to slowly increasing sub-MIC concentrations of the antibiotic cephalothin. The final resulting culture was exposed to a maximum antibiotic concentration of 7.5 μg/mL cephalothin. This final culture exhibited full clinical resistance to first and second-generation cephalosporins, with MIC values far exceeding the CLSI breakpoint. Additionally, the final culture had increased MICs to tetracycline, a highly mucoid appearance, and an elongated cellular morphology. Genome sequencing revealed a series of genetic alterations that could be mapped directly to the emergence of the resistant phenotype. Changes in phenotype and genetic alterations both indicate that alterations in the tetR regulator, LPS shedding, and capsule colanic acid synthesis may be directly associated with the resistance phenotype.
The three resistance-correlated mutations occurred in genes rpoB, tetR/acrR, and wcaJ. The increases in resistance occurred in a stepwise manner, similar to the increases in fitness observed in E. coli long-term evolution (LTEE) experiments [24,25,26]. LTEE studies also provided estimates for the rapidity of mutation fixation in a constant environment. The E. coli population in the LTEE experiments accumulated 20 mutations in the first 10,000 generations of growth, with a few rapid mutations that reached fixation in the population within 100 generations [25,26]. The three resistance-correlated mutations in this present study were all fixed within 36–378.9 generations using a generation time estimate from a related K. pneumoniae strain of 38–40 min [27]. Despite the use of incremental exposures to cephalothin in this study, we find that these mutations occurred rapidly in Klebsiella pneumoniae; such as what others have reported for E. coli.
The speed with which these three mutations were acquired also indicates that the sub-MIC concentration of cephalothin in the growth environment provides significant selective pressure. After only 24 h of exposure, the resistance of the adapted population to tetracycline had the first jump from 1 µg/mL to 8 µg/mL, which correlated with a fixed mutation in rpoB. The rapidity of fixation and correlated increase in survivability suggest that this mutation conferred a very high fitness advantage. It is possible that a necessary step in acquiring resistance for this population was a mutation in a gene meant to monitor nucleic acid integrity, allowing for further alterations to the genome.
The selection window hypothesis holds that sub-MIC level antibiotic exposure should generally increase bacterial mutation rates [8,9] because higher mutation rates will improve the chances of generating adaptive genetic change that provides a fitness advantage in conditions of antibiotic stress. Methodologies employed by studies testing this theory have generally identified “mutants” by plating samples of bacteria exposed to an antibiotic on media inoculated with some other antibiotic substance [5,10]. Mutation rates analyzed using this methodology tend to indicate that lower antibiotic concentrations result in high mutation rates that decline as the concentration of antibiotic nears the MIC [10,28]. However, when the whole-genome analysis is incorporated along with mutation accumulation analysis, substitutions, and insertions/deletions increased in frequency as exposure to antibiotics increases in E. coli [29]. Additionally, the rate of mutation in known resistance genes tracked in P. aeruginosa isolates generated conflicting results [12].
The sub-MIC doses used in this study are relevant to the discussion of environmental exposure and the induced rate of mutations. While well above the concentration generally found in freshwater sources [30], the doses used here are within the range commonly recovered from urine. Beta-lactams are not well adsorbed, with 50–90% of the drug regimen being recovered in urine [31]. A recent study of the impact of agricultural use of antibiotics found a significant increase in cephalothin resistance bacteria in manure commonly used as a field fertilizer [32]. Given the rapid rate of genetic modification seen in our study, brief exposures of soil bacteria to urine-excreted antibiotics are likely to drive the evolution of resistant isolates.
The present study utilized whole genome sequencing to identify mutations across the genome for the antibiotic-treated and unexposed samples after 14 days of total treatment. These data showed a higher number of mutations in the untreated sample compared to the adapted cohort. As noted by Long et al., a true mutation accumulation analysis that determines the frequency of genetic polymorphism per generation would require repeatedly passing a bacteria through bottlenecks to mitigate any selective influences [29]. The fact that the adapted population in this study was maintained as a whole group when sub-cultured might affect the relative number of mutations identified by whole genome sequencing. Another factor at work is that the treatment concentration of cephalothin did surpass the MIC of 4.0 μg/mL of the original K. pneumoniae 43816 for over half of the adaptation period (Table 1 and Table 2). Both the selection window hypothesis and adaptive studies of bacterial mutation suggest that such a high concentration would decrease mutation rates compared to an unexposed cohort and explain the low number of identified polymorphisms [5,8,9,10,28].
The rpoB mutation identified by Illumina sequencing early in the exposure regimen is very close to an alteration detected in the rmuC DNA recombination protein in the final sample sequenced using the SMRT cell method. The mutations identified do not all match between both sequencing methods, nor are the calls for similar genes found in the exact same position or using the same base changes (Table 3 and Table 4). This could reflect differences in methodology, genome construction in the two methods, and/or inherent error. These results highlight the limitations of using just one sequencing methodology for this type of analysis. Recent work has suggested that both long-read methods, such as SMRT cell sequencing, and short-read methods, such as Illumina sequencing, be combined for a hybrid approach for the most accurate formation of novel sequences, especially those of the Enterobacteriaceae family [33]. The data from both rounds of sequencing found that the SMRT cell method identified more polymorphisms in the adapted population genome and could identify mutations in non-coding regions, unlike the Illumina method. Additionally, the evolution of a resistant phenotype can require cooperative mutations adding difficulty to the present study’s ability to directly trace the effects of one mutation to a quantifiable change in antibiotic survival [28].
The protein products of both rpoB and rmuC interact closely with genomic DNA. RmuC is a regulator which can prevent sequence inversion during replication [34]. RmuC has also been identified as a possible multi-drug resistance (MDR) gene in other Gram-negative bacterial species [35,36]. However, RpoB is part of the RNA polymerase protein complex and, when mutated, can inhibit the action of rifamycin in bacteria [20]. Mutations in rpoB have been linked to rifamycin resistance in E. coli [37,38] and have been identified as a resistance gene in various other classes of bacteria [39]. Additionally, an early mutation in a gene tied to nucleic acid integrity provides a possible mechanism for further evolution of resistance by increasing the occurrence of replication error mutations in progeny.
TetR regulators are global multi-target transcriptional regulators that affect multiple processes within the cell beyond just efflux pumps [40]. Members of this family of regulatory proteins have been shown to impact a variety of virulence-associated targets, including motility, biofilm formation, and osmotic tolerance [40]. One member of this family has been directly associated with ftsZ, which is known to regulate cell division and bacillus cellular morphology [41]. Therefore, it is reasonable for a mutation in a tetR-type regulator to impact cellular functions and resistance to classes of antibiotics other than tetracycline.
Other studies on the in vivo evolution of Klebsiella within a patient undergoing antibiotic therapy have further identified TetR family upstream regulators of porins as being associated with clinical resistance. Yoshino et al. tracked the evolution of a clinical isolate of Klebsiella through treatment with multiple classes of antibiotics. They identified porin loss as a result of mutations in the ramR, a TetR family transcriptional regulator of micF, an antisense RNA regulator of the ompK35 porin [42]. The ultimate result of this ramR mutation was the loss of the ompK35 porin, limiting the entry of beta-lactam antibiotics into the periplasmic space. These data indicate that antibiotic resistance can be achieved through mutations in several different genes in a given pathway. In a sub-MIC exposure to the drug, it may even be advantageous for mutations to occur in regulator proteins rather than negatively impacting the primary function of a porin or another endpoint enzyme.
None of the identified mutations were found in penicillin-binding proteins, porins, or other cell wall modification pathways highly associated with traditional beta-lactam resistance. The data presented here add to a number of other recent studies that identify the role of a broad array of genes and cellular functions as being involved in the bacterial response and survival during antibiotic exposure. In addition to regulators of porins and cell wall regulators, Lopatkin et al. have identified core metabolic mutations that mitigate antibiotic susceptibility in clinical isolates [43]. Together these findings all support the need for broader investigations of gene alterations that contribute to resistance than those traditionally classified as such. The results from this study indicate that exploring alterations in gene regulatory pathways while limiting the ability of horizontal gene transfer to acquire new genes may be a powerful technique to identify gene clusters critical for bacterial survival from antibiotic-induced stress.
It is interesting that such a large insert and formation of a stop codon would be found in tetR/acrR, a gene known for resistance to tetracyclines when exposed to beta-lactam. However, the development of cross-resistance among bacterial populations exposed to one class of antibiotics is not uncommon. Exposure to environmental chemicals such as surface antiseptic chlorhexidine or veterinary antibiotics tilmicosin and florfenicol has been shown to create cross-resistance in human pathogens to different classes of antibiotics [4,6].
The genetic alterations associated with the largest increase in beta-lactam resistance were mapped to two adjacent genomic locations. The SMRT method identified an uncharacterized undecaprenyl phosphate–glucose phosphotransferase, which is very close to the mutation found in wcaJ by Illumina sequencing (Table 3 and Table 4). The second round of sequencing specified that there was a large deletion encompassing the end of the wcaJ and the beginning of the gndA genes, which was exclusive to the small colony-forming subpopulation.
In Klebsiella pneumoniae, wcaJ is part of the capsular cps operon and initiates the production of colanic acid [21,44]. The absence of wcaJ has been linked to increased resistance to phage treatment, decreased virulence in murine models, and increased phagocytosis by macrophages [21]. Studies ablating wcaJ in Klebsiella pneumoniae result in a nonmucoid phenotype while increasing biofilm production and increasing resistance to polymyxin [21,43].
Our observation of altered cellular morphology may be related to envelope stress and remodeling involving enzymes such as WcaJ. This elongated morphology has been demonstrated in experiments using high doses of beta-lactams to be a signature event in the process of drug-induced cellular lysis [45]. However, other studies of beta-lactam exposure have documented the permanent formation of filamentous bacterial cells [46]. Additionally, Kessler et al. demonstrated altered cell shape in E. coli wcaJ knockouts that exhibited accumulation of periplasmic colanic acid precursors and activation of the Rcs osmotic stress-response pathway [47]. New studies indicate that changes in cellular shape and the surface area-to-volume ratio can be part of an overall bacterial stress response to antibiotics [48]. Our results, which only used sub-MIC concentrations of beta-lactams, indicate that these stress responses alone can result in permanent changes to cellular morphology that may play a larger role in bacterial survival.
Colanic acid polymers, synthesized using the wcaJ gene product, can be covalently linked to lipopolysaccharides [49]. We observed increased LPS content in our extracted capsule polysaccharides, indicating that the wcaJ mutation may trigger increased membrane turnover. The ability to synthesize LPS O-antigen sugars has been documented to directly impact colanic acid synthesis [50]. It is, therefore, reasonable to hypothesize that alterations in wcaJ may likewise impact LPS synthesis, modification, and turnover. Studies of more general osmotic stress indicate the possibility of a shift from O-antigen attachment to colanic acid attachment [49]. These changes may directly impact overall membrane permeability and the ability of beta-lactams to access the periplasmic space.
Production of bacterial exopolysaccharides and a structured capsule may also have a significant effect on the ability of a drug to penetrate a viscous environment and target bacterial cells at all. Increases in growth media viscosity have been demonstrated to significantly affect bacterial metabolism, CFU counts, and virulence factor production [51]. Our data demonstrate a significant change in CFU/mL in the adapted cultures. Interestingly, Walker et al. [23] were reliant on optical density measures of cell count rather than CFU in their determination of the involvement of LPS in a hypermucoviscous phenotype. Our data demonstrating the involvement of colonic acid synthesis genes in the generation of mutants with increased viscosity, lowered CFU/mL concentrations, and increased MICs indicate that this issue needs further investigation.
At low antibiotic concentrations, these modifications in membrane composition and permeability may be sufficient for bacterial survival. This idea is supported by our analysis of MIC breakpoints to cephalosporins from multiple different generations. The multiple generations of cephalosporin antibiotics are classified in large part based on molecular modifications that extend the spectrum of activity against gram-negative aerobic bacilli such as Klebsiella [52]. We observed increased MICs to first and second, but not later-generation cephalosporins in both our adapted strain and the ΔwcaJ strain. These later-generation drugs, as well as carbapenems, have been modified to enhance permeability through the Gram-negative outer membrane. These data indicate that the mutations generated by our adapted stain may have directly impacted outer membrane permeability by a mechanism other than porin loss.
Together, these data indicate that low-level beta-lactam exposure initiates a cascade of modifications to the outer envelope and capsule polysaccharides that warrants further investigation. Studies examining single knockouts of tetR did not find associated changes in beta-lactam MICs [40]. While we observed increases in the MICs in the ΔwcaJ strain of Klebsiella pneumoniae 43816, Kessler et al. did not when using a knockout of the same gene in E. coli [47]. This may be related to the more complex nature and function of the capsule in Klebsiella. Mutations in rpoB have been associated with beta-lactam resistance but not resistance to multiple classes of antibiotics [53]. It is, therefore, highly likely that a combination of multiple mutations is required to achieve the multi-drug resistant phenotype observed in this experiment.
Our final adapted culture represented a heterogeneous mixture of two colony morphologies, which was reflected in the genome sequence analysis. The deletion spanning between wcaJ and gndA only occurred in the small colony-forming population. As noted above, ablation of wcaJ in K. pneumoniae is linked to a decrease in species mucoidy and increased sensitivity to polymyxin. Similarly, gndA is a gene within the cps locus responsible for the K2 serotype and capsule formation of K. pneumoniae [54,55]. It, therefore, seems likely that the mucoid phenotype seen in the fourteen-day adapted population whole-group mixture is due to the large colony subpopulation only. This is supported by the fact that the small colony subpopulation has reduced resistance to cephalothin compared to the large colonies which have an intact wcaJ sequence.
This study also demonstrates the complexity of changes occurring within what can be characterized as a single culture study. Multiple mutations occur within different cells in this population over time, resulting in a heterogeneous, non-clonal population. As compared to studies such as the LTEE [24,25,26], we have added the directional pressure of sub-MIC antibiotic exposure and prevented the acquisition of new resistance genes from other nearby bacterial species, such as might be found with a patient. Tracking the in vivo evolution of clinical isolates has been used to demonstrate the evolution of resistance by single Klebsiella isolates to colistin [56], carbapenems [57], and beta-lactams [42,58]. However, these studies often focus primarily on the acquisition of genes by horizontal gene transfer and only recently have identified mutations in other pathways as playing a significant role [41,47] in bacterial survival within the antibiotic-treated host. This study demonstrates that limiting the opportunity for horizontal gene transfer may help to reveal gene regulatory networks critical to survival from antibiotic stress that are otherwise cloaked by the acquisition of new, more traditional resistance genes. Larger scale studies using multiple cultures provided the same antibiotic exposure in parallel and are ongoing to investigate how many different paths and mutations may be used to ensure bacterial survival.
In summary, this progressive, sub-MIC antibiotic exposure experiment resulted in a mixture of K. pneumoniae exhibiting multiple genomic changes. The multiple isolates from this experiment resulted in K. pneumoniae that were resistant rather than tolerant of the antibiotic, as they exhibited normal metabolic activity and growth in the presence of the antibiotic [59]. While both the large and small variant isolates exhibited elevated MICs to cephalothin, the mixture of these isolates demonstrated a synergistic protective effect. This mixture is reflective of what would occur in the environment, where individual cells may independently evolve, persist, or assist in the survival of nearby cells. The K. pneumoniae in this study achieved clinical resistance without the traditional acquisition of a beta-lactamase gene. These compensatory mutations, and combinations of them, warrant further investigation as they may accelerate and enhance resistance associated with traditional horizontal gene transfer.
4. Conclusions
Bacteria are constantly exposed to low levels of antibiotics in the environment. The impact of this low-level exposure on bacterial evolution is not well understood. In this work, we developed a model to expose Klebsiella pneumoniae to progressive, low doses of the antibiotic cephalothin. After a fourteen-day exposure regimen, our culture exhibited full clinical resistance to this antibiotic without the traditional acquisition of inactivating genes. This culture also exhibited resistance to tetracycline, had a highly mucoid appearance, and exhibited altered elongated cellular morphology. Whole genome sequencing identified a collection of mutations to the K. pneumoniae genome that could be mapped to the emergence of the resistant phenotype. This study demonstrates that antibiotic resistance can be achieved in response to low-level antibiotic exposure and without the traditional acquisition of resistance genes. Further, this study identifies new genes that may play a role in the evolution of antibiotic-resistant bacteria.
5. Materials and Methods
5.1. Bacterial Strains and Progressive Antibiotic Exposure
Klebsiella pneumoniae 43816 (ATCC, Manassas, VA, USA) was used as the starting culture for the progressive antibiotic exposure experiment (Figure 1). All cultures were grown in Luria-Bertani (LB) broth (BD Difco, Franklin Lakes, NJ, USA) at 37 °C with shaking at 200 rpm. All antibiotics and reagents are from Thermo Fisher Scientific (Waltham, MA, USA) unless otherwise indicated. K. pneumoniae was grown overnight from frozen stocks, and 50 μL of this culture was added to 5 mL of fresh LB with 0.5 mg/mL cephalothin added. The concentration of 0.5 μg/mL cephalothin was significantly below the MIC for this organism. Cultures were grown for 12 h, at which point 50 μL were transferred to 5 mL of fresh LB with the same concentration of cephalothin. After a total of 24 h exposure to one dose of antibiotics, 50 μL of culture was transferred to a new 5 mL of LB with a higher concentration of cephalothin. Each stepwise exposure increased the dose of cephalothin by 0.5 μg/mL to a final dose of 7.5 μg/mL cephalothin. An untreated culture of K. pneumoniae 43816 was grown in parallel without the addition of antibiotics. Frozen glycerol stocks were made of each culture at each 12 h transfer point for later analysis. Klebsiella pneumoniae 43816 ΔwcaJ (VK646) was a kind gift from Kimberly A. Walker and Virginia Miller at UNC Chapel Hill [23].
5.2. Bacterial Growth and Morphology
At the 12 h timepoint in each exposure, the bacteria were transferred to fresh LB with an antibiotic. At this point, the time required for the bacterial culture to reach an OD600 of 1.0 was determined using an Eppendorf Biophotometer (Hamburg, Germany). Decreases in the growth rate were determined by subtracting the elapsed time for the treated culture from the average time to OD600 1.0 of all untreated cultures (142 min).
Growth curves were generated for the original, untreated fourteen-day culture and antibiotic-adapted strain from frozen stocks. Frozen stocks were grown overnight in LB, and then 50 μL were transferred to fresh LB. 7.5 μg/mL of cephalothin was added to all adapted culture media. Growth was determined at OD600 every 30 min using the Eppendorf Biophotometer.
Dilution plate counts were used to determine the colony-forming units (CFU) for each culture at an OD600 of 1.0. Three independent vials of each culture were grown overnight, centrifuged at 5000 rpm for 5 min, re-suspended in sterile PBS to an OD600 of 1.0, and then serially plated on LB agar. Duplicate plates were counted for each dilution and averaged.
Changes in colony morphology were determined by quadrant streaking on LB agar plates. Colony size was measured using ImageJ to measure 20 colony diameters per plated culture. Pixel measurements were converted to millimeters based on the standard diameter of the plate (100 mm).
To determine cellular morphology, overnight bacterial cultures were centrifuged at 5000 rpm for 5 min and re-suspended in phosphate-buffered saline (PBS) (pH 8.0). Samples were then diluted in PBS to an optical density of approximately 1.0 OD600. Cells were negatively stained with 1% Nigrosin and visualized by brightfield microscopy at 1000× with an oil immersion lens.
5.3. Determination of Minimum Inhibitory Concentration Breakpoints
Broth microdilution assays were used to determine the minimum inhibitory concentration (MIC) of tetracycline, amikacin, and five different beta-lactam antibiotics (Cephalothin, Cefoxitin, Cefotaxime, Cefepime, and Imipenem). The beta-lactam drugs were chosen as representative of different generations of cephalosporin antibiotics and a carbapenem, respectively. Ninety-six well plates were seeded with LB broth containing 103 CFU/mL starting concentration of bacteria. Antibiotics were added in a two- fold serial dilution starting at 500 μg/mL. Cultures were incubated overnight, and growth was measured at OD600 using a Biotek Gen5 plate reader (Winooski, VT, USA). Wells containing only LB and bacteria were used as controls for normal growth. The MIC breakpoint was determined as the lowest antibiotic concentration at no increase in optical density over a broth; only control was detected. All samples were tested in triplicate.
5.4. Genomic DNA Isolation
Genomic DNA was extracted using a protocol modified by Wright et al. [60]. Briefly, 50 mL of overnight culture was pelleted for 10 min at 10,000 rpm at 4 °C. The pellet was washed twice with TE25S buffer (25 mM Tris-HCl, 25 mM EDTA, 0.3 M sucrose, pH 8.0) and re-suspended in TE25S with 10 μg/mL lysozyme and RNAse. The mixture was incubated for two hours at 37 °C with shaking at 150 rpm. Proteinase K and 10% SDS were added and incorporated by inversion and incubated for 1–2 h at 50–55 °C with periodic inversions. 5 M NaCl was added, followed by 3.25 mL of CTAB (Cetyl Trimethyl Ammonium Bromide)/NaCl. The solution was mixed by inversion and incubated at 55 °C for 10 min. A 24:1 chloroform/isoamyl alcohol solution was added and incubated at room temperature with shaking at 100 rpm for 20 min. After incubation, the solution was centrifuged at 10,000 rpm and 4 °C for 15 min, and the upper aqueous layers were transferred into fresh tubes. This chloroform treatment was repeated a second time. The upper aqueous layers of both samples were combined with an 0.6 volume of isopropyl alcohol, and the mixture was gently inverted. After five minutes, the purified DNA was spooled from the tube onto a sterile Pasteur pipette. The spooled DNA was washed with approximately 5 mL of 70% ethanol and dried before being suspended in 300 µL of EB buffer (QIAGEN, Hilden, Germany). Purified DNA was quantified by Qubit (Thermo Fisher, Waltham, MA, USA).
5.5. Genome Sequencing
Whole genome sequencing was performed on the following K. pneumoniae samples: days 1–14 of the adaptation experiment, the small colony-forming variant, the large colony-forming variant, and the untreated sample after 14 days of culture. These DNA samples were analyzed by the Microbial Genome Sequencing Center (https://www.seqcenter.com, accessed on 11 April 2023), which utilized an Illumina sequencing technique similar to that used by Baym et al. [61]. Samples were compared against the published reference sequence NZ_CP009208.1 for K. pneumoniae 43816 [15]. Any variations were analyzed using a proprietary breseq variant calling algorithm [62].
To confirm these results and to extend our analysis to include SNPs within promoter regions, endpoint samples of DNA from K. pneumoniae 43816, the fourteen-day antibiotic-adapted culture, and the fourteen-day untreated population were also analyzed using SMRT Cell Sequencing by the National Center for Genome Resources (www.ncgr.org, accessed on 11 April 2023). Samples were again compared against the published reference sequence NZ_CP009208.1 for K. pneumoniae 43816 [15]. Genetic variation between genomes was calculated using a modified form of FST or analysis of variance referred to as θ [63]. FST values are evaluated against the null hypothesis that the populations are not genetically unique [63]. Pacific Biosciences calculated allele frequencies and utilized a proprietary Quiver Algorithm to maximize accuracy in sequence reads using variation between the published genome and prior records.
5.6. Data Availability
The complete genome sequences are available via the NCBI BioProject data base. The project ID is PRJNA854906.
5.7. Capsule Extraction and Characterization
Capsular polysaccharide (CPS) was extracted using the protocol outlined by Domineco et al. [64]. Briefly, 500 mL of an overnight culture was mixed with 100 µL of 1% Zwittergent 3–14 in 100 mM citric acid, pH 2.0. The mixture was vortexed vigorously, incubated at 50 °C for 20 min, and centrifuged for 5 min at 14,000 rpm. The supernatant was then transferred to a fresh tube, mixed with 1.2 mL of absolute ethanol, and incubated for 90 min at 4 °C. Precipitate was collected after centrifugation at 14,000 rpm for 10 min and dissolved in 200 µL DI water.
CPS d-glucuronic acid was quantified following a previously established protocol by Lin et al. [65]. Purified CPS was vortexed vigorously with 1.2 mL of 12.5 mM sodium tetraborate in concentrated sulfuric acid and heated for 5 min at 95 °C. The samples were cooled before the addition of 20 µL of 0.15% m-hydroxydiphenyl, and the absorbance was measured at 540 nm. A standard curve was generated using d-glucuronic acid to determine the concentration of glucuronic acid in the CPS samples. To ensure quantification of CPS from the same number of bacteria, strains were normalized to 108 CFUs/mL. Each assay was performed in triplicate from six individual cultures.
The extracted capsule samples were also quantified for protein content and sialic acid content. Protein content was determined using a BCA Protein Assay kit (Thermo Scientific), according to the manufacturer’s directions. Sialic acid was quantified using the Sialic Acid Quantitation Kit (Sigma, St. Louis, MO, USA), according to the manufacturer’s directions.
The carbohydrate composition of the CPS was characterized by GC-MS, as previously detailed in Brunson et al. [66]. Briefly, CPS was purified from LPS using sodium deoxycholate at a final concentration of 6 mM, as described by Kachlany et al. [67]. The CPS was pelleted with cold ethanol, then freeze-dried before being hydrolyzed in 0.5 M HCl at 85 °C for 18 h. The hydrolyzed carbohydrates were modified with the Tri-Sil HTP reagent (Thermo Scientific, Wltham, MA, USA), as described by York et al. [68]. The modified carbohydrates were dried and re-suspended in 1 mL hexane. The carbohydrate suspension in hexane was centrifuged at 1000× g for 5 min, and the supernatant was collected for GC-MS analysis.
GC-MS analyses were conducted with a CP-3800 GC (Varian, Palo Alto, CA, USA) using a Supelco SPB-608 30-m fused silica capillary column containing a bonded stationary phase (0.25 μm film thickness). The TMS (tri-methyl silyl) conjugated glycans were analyzed using the electron ionization mode with a Saturn 2200 GC/MS (Varian, Palo Alto, CA, USA). The initial oven temperature was 80 °C, held for 2 min. Then, the temperature was raised by 20 °C/min and held at 160 °C for 12 min. Finally, the oven temperature was raised by 20 °C/min and held at 260 °C for 7 min.
5.8. Quantification of Lipopolysaccharide
Whole-cell lipopolysaccharide levels were quantified using the Purpald assay [69,70]. Briefly stated, cultures were centrifuged at 5000 rpm for 5 min and re-suspended in PBS. 50 µL of this cell solution was treated with 32 mM sodium periodate solution in a 96-well plate and incubated at room temperature for 25 min. After this incubation period, a 136 mM Purpald solution in 2 N NaOH was added, and the plate was incubated for an additional 20 min. Following the completion of this step, 64 mM sodium periodate solution was added, and the plate was incubated for 20 min. Absorbance was immediately determined at 540 nm and compared to a standard curve of pure lipopolysaccharide isolated from K. pneumoniae (Sigma, St. Louis, MO, USA). This assay was also used for capsule extraction, as described above for the glucuronic acid assay. Results are presented with LPS concentrations normalized to 108 CFUs.
5.9. Statistical Analysis
All statistical significance was determined using a one-way ANOVA and Tukey’s post hoc test using XLSTAT software. The significance of the d-glucuronic acid and protein quantification was compared against results from the time zero, untreated sample.
Author Contributions
Funding acquisition, T.N.E.; investigation, J.R.A., T.T., N.B.L., J.L.J., S.M.D., E.M., A.V., M.R.C. and T.N.E.; resources, T.N.E.; supervision, T.N.E.; visualization, T.N.E.; writing—original draft, J.R.A. and T.N.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by funding from UNF to T.N.E., including a Dean’s Council Fellowship Award, a Research Enhancement Plan Award, and the Transformational Learning Opportunity Program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The complete genome sequences are available via the NCBI BioProject data base. The project ID is PRJNA854906. All other data available upon request.
Acknowledgments
We thank Frank Smith for assistance with bioinformatics software and all members of the Ellis lab for thoughtful input and comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman, C.U.; Mohan, D. Pharmaceuticals of Emerging Concern in Aquatic Systems: Chemistry, Occurrence, Effects, and Removal Methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [PubMed]
- Polianciuc, S.I.; Gurzău, A.E.; Kiss, B.; Georgia Ștefan, M.; Loghin, F. Antibiotics in the Environment: Causes and Consequences. Med. Pharm. Reports 2020, 93, 231–240. [Google Scholar] [CrossRef] [PubMed]
- National Action Plan to Combat Antibiotic-Resistant Bacteria. 2015. Available online: https://www.cdc.gov/drugresistance/us-activities/national-action-plan.html (accessed on 11 April 2023).
- Kampf, G. Acquired Resistance to Chlorhexidine—Is It Time to Establish an ‘Antiseptic Stewardship’ Initiative? J. Hosp. Infect. 2016, 94, 213–227. [Google Scholar] [CrossRef]
- Nair, C.G.; Chao, C.; Ryall, B.; Williams, H.D. Sub-Lethal Concentrations of Antibiotics Increase Mutation Frequency in the Cystic Fibrosis Pathogen Pseudomonas Aeruginosa. Lett. Appl. Microbiol. 2013, 56, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Bhunia, A.K. Animal-Use Antibiotics Induce Cross-Resistance in Bacterial Pathogens to Human Therapeutic Antibiotics. Curr. Microbiol. 2019, 76, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of Antimicrobial Resistance in the Environment and Its Relevance to Environmental Regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef]
- Drlica, K. The Mutant Selection Window and Antimicrobial Resistance. J. Antimicrob. Chemother. 2003, 52, 11–17. [Google Scholar] [CrossRef]
- Drlica, K.; Zhao, X. Mutant Selection Window Hypothesis Updated. Clin. Infect. Dis. 2007, 44, 681–688. [Google Scholar] [CrossRef]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of Resistant Bacteria at Very Low Antibiotic Concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef]
- Low, Y.M.; Chong, C.W.; Yap, I.K.S.; Chai, L.C.; Clarke, S.C.; Ponnampalavanar, S.; Abdul Jabar, K.; Md Yusof, M.Y.; Teh, C.S.J. Elucidating the Survival and Response of Carbapenem Resistant Klebsiella Pneumoniae after Exposure to Imipenem at Sub-Lethal Concentrations. Pathog. Glob. Health 2018, 112, 378–386. [Google Scholar] [CrossRef]
- Migliorini, L.B.; Brüggemann, H.; De Sales, R.O.; Koga, P.C.M.; De Souza, A.V.; Martino, M.D.V.; Galhardo, R.S.; Severino, P. Mutagenesis Induced by Sub-Lethal Doses of Ciprofloxacin: Genotypic and Phenotypic Differences between the Pseudomonas Aeruginosa Strain PA14 and Clinical Isolates. Front. Microbiol. 2019, 10, 1553. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella Pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Simmonds, A.; Uhlemann, A.C. Clinical Implications of Genomic Adaptation and Evolution of Carbapenem-Resistant Klebsiella Pneumoniae. J. Infect. Dis. 2017, 215, S18–S27. [Google Scholar] [CrossRef] [PubMed]
- Broberg, C.A.; Wu, W.; Cavalcoli, J.D.; Miller, V.L.; Bachman, M.A. Complete Genome Sequence of Klebsiella Pneumoniae Strain ATCC 43816 KPPR1, a Rifampin-Resistant Mutant Commonly Used in Animal, Genetic, and Molecular Biology Studies. Genome Announc. 2014, 2, e00924-14. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Barrick, J.E. Breseq Manual—Breseq 0.35.2rc1 Documentation. Available online: https://barricklab.org/twiki/pub/Lab/ToolsBacterialGenomeResequencing/documentation/index.html (accessed on 11 April 2023).
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Bortolaia, V.; Tate, H.; Tyson, G.H.; Aarestrup, F.M.; McDermott, P.F. Using Genomics to Track Global Antimicrobial Resistance. Front. Public Health 2019, 7, 242. [Google Scholar] [CrossRef]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural Mechanism for Rifampicin Inhibition of Bacterial RNA Polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Cai, R.; Wang, G.; Le, S.; Wu, M.; Cheng, M.; Guo, Z.; Ji, Y.; Xi, H.; Zhao, C.; Wang, X.; et al. Three Capsular Polysaccharide Synthesis-Related Glucosyltransferases, GT-1, GT-2 and WcaJ, Are Associated with Virulence and Phage Sensitivity of Klebsiella Pneumoniae. Front. Microbiol. 2019, 10, 1189. [Google Scholar] [CrossRef]
- Lee, C.H.; Chang, C.C.; Liu, J.W.; Chen, R.F.; Yang, K.D. Sialic Acid Involved in Hypermucoviscosity Phenotype of Klebsiella Pneumoniae and Associated with Resistance to Neutrophil Phagocytosis. Virulence 2014, 5, 673–679. [Google Scholar] [CrossRef]
- Walker, K.A.; Treat, L.P.; Sepúlveda, V.E.; Miller, V.L. The Small Protein RmpD Drives Hypermucoviscosity in Klebsiella Pneumoniae. MBio 2020, 11, e01750-20. [Google Scholar] [CrossRef]
- Lenski, R.E. Experimental Evolution and the Dynamics of Adaptation and Genome Evolution in Microbial Populations. ISME J. 2017, 11, 2181–2194. [Google Scholar] [CrossRef] [PubMed]
- Good, B.H.; McDonald, M.J.; Barrick, J.E.; Lenski, R.E.; Desai, M.M. The Dynamics of Molecular Evolution over 60,000 Generations. Nature 2017, 551, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, A. Fixation and Adaptation in the Lenski E. Coli Long Term Evolution Experiment. Biomed. J. Sci. Tech. Res. 2019, 20, 14754–14760. [Google Scholar] [CrossRef][Green Version]
- Regué, M.; Hita, B.; Piqué, N.; Izquierdo, L.; Merino, S.; Fresno, S.; Benedí, V.J.; Tomás, J.M. A Gene, Uge, Is Essential for Klebsiella Pneumoniae Virulence. Infect. Immun. 2004, 72, 54–61. [Google Scholar] [CrossRef]
- Martinez, J.L.; Baquero, F. Mutation Frequencies and Antibiotic Resistance. Antimicrob. Agents Chemother. 2000, 44, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Miller, S.F.; Strauss, C.; Zhao, C.; Cheng, L.; Ye, Z.; Griffin, K.; Te, R.; Lee, H.; Chen, C.C.; et al. Antibiotic Treatment Enhances the Genome-Wide Mutation Rate of Target Cells. Proc. Natl. Acad. Sci. USA 2016, 113, E2498–E2505. [Google Scholar] [CrossRef]
- Sta Ana, K.M.; Madriaga, J.; Espino, M.P. β-Lactam Antibiotics and Antibiotic Resistance in Asian Lakes and Rivers: An Overview of Contamination, Sources and Detection Methods. Environ. Pollut. 2021, 275, 116624. [Google Scholar] [CrossRef]
- Brumfitt, W.; Kosmidis, J.; Hamilton-Miller, J.M.; Gilchrist, J.N. Cefoxitin and Cephalothin: Antimicrobial Activity, Human Pharmacokinetics, and Toxicology. Antimicrob. Agents Chemother. 1974, 6, 290–299. [Google Scholar] [CrossRef]
- Udikovic-Kolic, N.; Wichmann, F.; Broderick, N.A.; Handelsman, J. Bloom of Resident Antibiotic-Resistant Bacteria in Soil Following Manure Fertilization. Proc. Natl. Acad. Sci. USA 2014, 111, 15202–15207. [Google Scholar] [CrossRef]
- De Maio, N.; Shaw, L.P.; Hubbard, A.; George, S.; Sanderson, N.D.; Swann, J.; Wick, R.; Oun, M.A.; Stubberfield, E.; Hoosdally, S.J.; et al. Comparison of Long-Read Sequencing Technologies in the Hybrid Assembly of Complex Bacterial Genomes. Microb. Genom. 2019, 5, e000294. [Google Scholar] [CrossRef]
- Slupska, M.M.; Chiang, J.H.; Luther, W.M.; Stewart, J.L.; Amii, L.; Conrad, A.; Miller, J.H. Genes Involved in the Determination of the Rate of Inversions at Short Inverted Repeats. Genes Cells 2000, 5, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, S.M.; Ernst, R.K.; Miller, S.I. PmrAB, a Two-Component Regulatory System of Pseudomonas Aeruginosa That Modulates Resistance to Cationic Antimicrobial Peptides and Addition of Aminoarabinose to Lipid A. J. Bacteriol. 2004, 186, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Van Dyk, T.K.; DeRose, E.J.; Gonye, G.E. LuxArray, a High-Density, Genomewide Transcription Analysis of Escherichia Coli Using Bioluminescent Reporter Strains. J. Bacteriol. 2001, 183, 5496–5505. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.J.; Gross, C.A. Mapping and Sequencing of Mutations in the Escherichia ColirpoB Gene That Lead to Rifampicin Resistance. J. Mol. Biol. 1988, 202, 45–58. [Google Scholar] [CrossRef]
- Severinov, K.; Soushko, M.; Goldfarb, A.; Nikiforov, V. RifR Mutations in the Beginning of the Escherichia Coli RpoB Gene. MGG Mol. Gen. Genet. 1994, 244, 120–126. [Google Scholar] [CrossRef]
- Alifano, P.; Palumbo, C.; Pasanisi, D.; Talà, A. Rifampicin-Resistance, RpoB Polymorphism and RNA Polymerase Genetic Engineering. J. Biotechnol. 2015, 202, 60–77. [Google Scholar] [CrossRef]
- Colclough, A.L.; Scadden, J.; Blair, J.M.A. TetR-Family Transcription Factors in Gram-Negative Bacteria: Conservation, Variation and Implications for Efflux-Mediated Antimicrobial Resistance. BMC Genom. 2019, 20, 731. [Google Scholar] [CrossRef]
- Du, S.; Lutkenhaus, J. SlmA Antagonism of FtsZ Assembly Employs a Two-Pronged Mechanism like MinCD. PLoS Genet. 2014, 10, e1004460. [Google Scholar] [CrossRef]
- Yoshino, M.; Aihara, M.; Gotoh, Y.; Akimoto, M.; Tatsuhara, W.; Kiyosuke, M.; Matsushima, Y.; Uchiumi, T.; Hayashi, T.; Kang, D. Stepwise Evolution of a Klebsiella Pneumoniae Clone within a Host Leading to Increased Multidrug Resistance. mSphere 2021, 6, e00734-21. [Google Scholar] [CrossRef]
- Lopatkin, A.J.; Bening, S.C.; Manson, A.L.; Stokes, J.M.; Kohanski, M.A.; Badran, A.H.; Earl, A.M.; Cheney, N.J.; Yang, J.H.; Collins, J.J. Clinically Relevant Mutations in Core Metabolic Genes Confer Antibiotic Resistance. Science 2021, 371, eaba0862. [Google Scholar] [CrossRef]
- Pal, S.; Verma, J.; Mallick, S.; Rastogi, S.K.; Kumar, A.; Ghosh, A.S. Absence of the Glycosyltransferase Wcaj in Klebsiella Pneumoniae Atcc13883 Affects Biofilm Formation, Increases Polymyxin Resistance and Reduces Murine Macrophage Activation. Microbiology 2019, 165, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Kahne, D.; Kishony, R. Distinct Single-Cell Morphological Dynamics under Beta-Lactam Antibiotics. Mol. Cell 2012, 48, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Inui, T.; Endo, T.; Matsushita, T. Morphological Changes and Lysis Induced by Beta-Lactams Associated with the Characteristic Profiles of Affinities of Penicillin-Binding Proteins in Actinobacillus Pleuropneumoniae. Antimicrob. Agents Chemother. 2000, 44, 1518–1523. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kessler, N.G.; Caraballo Delgado, D.M.; Shah, N.K.; Dickinson, J.A.; Moorea, S.D. Exopolysaccharide Anchoring Creates an Extreme Resistance to Sedimentation. J. Bacteriol. 2021, 203, e00023-21. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, N.; Serbanescu, D.; Banerjee, S. Antibiotic Resistance via Bacterial Cell Shape-Shifting. MBio 2022, 13, e00659-22. [Google Scholar] [CrossRef]
- Meredith, T.C.; Mamat, U.; Kaczynski, Z.; Lindner, B.; Holst, O.; Woodard, R.W. Modification of Lipopolysaccharide with Colanic Acid (M-Antigen) Repeats in Escherichia Coli. J. Biol. Chem. 2007, 282, 7790–7798. [Google Scholar] [CrossRef]
- Ren, G.; Wang, Z.; Li, Y.; Hu, X.; Wang, X. Effects of Lipopolysaccharide Core Sugar Deficiency on Colanic Acid Biosynthesis in Escherichia Coli. J. Bacteriol. 2016, 198, 1576–1584. [Google Scholar] [CrossRef]
- Borić, M.; Danevčič, T.; Stopar, D. Viscosity Dictates Metabolic Activity of Vibrio Ruber. Front. Microbiol. 2012, 3, 255. [Google Scholar] [CrossRef]
- Cephalosporins—Infectious Diseases—Merck Manuals Professional Edition. Available online: https://www.merckmanuals.com/professional/infectious-diseases/bacteria-and-antibacterial-drugs/cephalosporins (accessed on 26 November 2022).
- Palace, S.G.; Wang, Y.; Rubin, D.H.F.; Welsh, M.A.; Mortimer, T.D.; Cole, K.; Eyre, D.W.; Walker, S.; Grad, Y.H. Rna Polymerase Mutations Cause Cephalosporin Resistance in Clinical Neisseria Gonorrhoeae Isolates. Elife 2020, 9, e51407. [Google Scholar] [CrossRef]
- Nelson, K.; Selander, R.K. Intergeneric Transfer and Recombination of the 6-Phosphogluconate Dehydrogenase Gene (Gnd) in Enteric Bacteria. Proc. Natl. Acad. Sci. USA 1994, 91, 10227–10231. [Google Scholar] [CrossRef]
- Arakawa, Y.; Wacharotayankun, R.; Nagatsuka, T.; Ito, H.; Kato, N.; Ohta, M. Genomic Organization of the Klebsiella Pneumoniae Cps Region Responsible for Serotype K2 Capsular Polysaccharide Synthesis in the Virulent Strain Chedid. J. Bacteriol. 1995, 177, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Jousset, A.B.; Bonnin, R.A.; Rosinski-Chupin, I.; Girlich, D.; Cuzon, G.; Cabanel, N.; Frech, H.; Farfour, E.; Dortet, L.; Glaser, P.; et al. A 4.5-Year Within-Patient Evolution of a Colistin-Resistant Klebsiella Pneumoniae Carbapenemase-Producing K. Pneumoniae Sequence Type 258. Clin. Infect. Dis. 2018, 67, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, Q.; Perlaza-Jiménez, L.; Zheng, X.; Zhao, Y.; Dhanasekaran, V.; Fang, R.; Li, J.; Wang, C.; Liu, H.; et al. First Description of Antimicrobial Resistance in Carbapenem-Susceptible Klebsiella Pneumoniae after Imipenem Treatment, Driven by Outer Membrane Remodeling. BMC Microbiol. 2020, 20, 218. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Nishida, R.; Akimoto, M.; Gotoh, Y.; Kiyosuke, M.; Uchiumi, T.; Nishioka, M.; Matsushima, Y.; Hayashi, T.; Kang, D. Within-Host Evolution of a Klebsiella Pneumoniae Clone: Selected Mutations Associated with the Alteration of Outer Membrane Protein Expression Conferred Multidrug Resistance. J. Antimicrob. Chemother. 2021, 76, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Chebotar, I.V.; Emelyanova, M.A.; Bocharova, J.A.; Mayansky, N.A.; Kopantseva, E.E.; Mikhailovich, V.M. The Classification of Bacterial Survival Strategies in the Presence of Antimicrobials. Microb. Pathog. 2021, 155, 104901. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Adelskov, J.; Greene, A.C. Bacterial DNA Extraction Using Individual Enzymes and Phenol/Chloroform Separation. J. Microbiol. Biol. Educ. 2017, 18, 18–22. [Google Scholar] [CrossRef]
- Baym, M.; Kryazhimskiy, S.; Lieberman, T.D.; Chung, H.; Desai, M.M.; Kishony, R.K. Inexpensive Multiplexed Library Preparation for Megabase-Sized Genomes. PLoS ONE 2015, 10, e0128036. [Google Scholar] [CrossRef]
- Deatherage, D.E.; Barrick, J.E. Identification of Mutations in Laboratory-Evolved Microbes from next-Generation Sequencing Data Using Breseq. Methods Mol. Biol. 2014, 1151, 165–188. [Google Scholar] [CrossRef]
- Martin, A.P. Phylogenetic Approaches for Describing and Comparing the Diversity of Microbial Communities. Appl. Environ. Microbiol. 2002, 68, 3673–3682. [Google Scholar] [CrossRef]
- Domenico, P.; Schwartz, S.; Cunha, B.A. Reduction of Capsular Polysaccharide Production in Klebsiella Pneumoniae by Sodium Salicylate. Infect. Immun. 1989, 57, 3778–3782. [Google Scholar] [CrossRef]
- Lin, T.L.; Yang, F.L.; Yang, A.S.; Peng, H.P.; Li, T.L.; Tsai, M.D.; Wu, S.H.; Wang, J.T. Amino Acid Substitutions of MagA in Klebsiella Pneumoniae Affect the Biosynthesis of the Capsular Polysaccharide. PLoS ONE 2012, 7, e46783. [Google Scholar] [CrossRef] [PubMed]
- Brunson, D.N.; Maldosevic, E.; Velez, A.; Figgins, E.; Ellis, T.N. Porin Loss in Klebsiella Pneumoniae Clinical Isolates Impacts Production of Virulence Factors and Survival within Macrophages. Int. J. Med. Microbiol. 2019, 309, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Kachlany, S.C.; Levery, S.B.; Kim, J.S.; Reuhs, B.L.; Lion, L.W.; Ghiorse, W.C. Structure and Carbohydrate Analysis of the Exopolysaccharide Capsule of Pseudomonas Putida G7. Environ. Microbiol. 2001, 3, 774–784. [Google Scholar] [CrossRef] [PubMed]
- York, W.S.; Darvill, A.G.; McNeil, M.; Stevenson, T.T.; Albersheim, P. Isolation and Characterization of Plant Cell Walls and Cell Wall Components. Methods Enzymol. 1986, 118, 3–40. [Google Scholar] [CrossRef]
- Turner, K.L.; Cahill, B.K.B.K.; Dilello, S.K.S.K.; Gutel, D.; Brunson, D.N.D.N.; Albertí, S.; Ellis, T.N.T.N. Porin Loss Impacts the Host Inflammatory Response to Outer Membrane Vesicles of Klebsiella Pneumoniae. Antimicrob. Agents Chemother. 2015, 60, 1360–1369. [Google Scholar] [CrossRef]
- Velkov, T.; Soon, R.L.; Chong, P.L.; Huang, J.X.; Cooper, M.A.; Azad, M.A.K.; Baker, M.A.; Thompson, P.E.; Roberts, K.; Nation, R.L.; et al. Molecular Basis for the Increased Polymyxin Susceptibility of Klebsiella Pneumoniae Strains with Under-Acylated Lipid A. Innate Immun. 2013, 19, 265–277. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).