Abstract
Antimicrobial resistance (AMR) is an emerging public health threat and is one of the One Health priorities for humans, animals, and environmental health. Red foxes (Vulpes vulpes) are a widespread predator species with great ecological significance, and they may serve as a sentinel of antimicrobial resistance in the general environment. The present study was carried out to detect antimicrobial resistance, antimicrobial resistance genes, and genetic diversity in faecal isolates of red foxes (Vulpes vulpes). In total, 34 Enterococcus isolates, including E. faecium (n = 17), E. faecalis (n = 12), E. durans (n = 3), and E. hirae (n = 2), were isolated. Antimicrobial resistance to 12 antimicrobial agents was detected with EUVENC panels using the minimum inhibitory concentration (MIC). The presence of antimicrobial resistance genes (ARGs) was determined using whole-genome sequencing (WGS). Resistance to tetracycline (6/34), erythromycin (3/34), ciprofloxacin (2/34), tigecycline (2/34), and daptomycin (2/34) was identified in 44% (15/34) of Enterococcus isolates, while all the isolates were found to be susceptible to ampicillin, chloramphenicol, gentamicin, linezolid, teicoplanin, and vancomycin. No multi-resistant Enterococcus spp. were detected. A total of 12 ARGs were identified in Enterococcus spp., with the presence of at least 1 ARG in every isolate. The identified ARGs encoded resistance to aminoglycosides (aac(6′)-I, ant(6)-Ia, aac(6′)-Iih and spw), tetracyclines (tet(M), tet(L) and tet(S)), and macrolide–lincosamide–streptogramin AB (lnu(B,G), lsa(A,E), and msr(C)), and their presence was associated with phenotypical resistance. Core genome multilocus sequence typing (cgMLST) revealed the high diversity of E. faecalis and E. faecium isolates, even within the same geographical area. The distribution of resistant Enterococcus spp. in wild foxes in Latvia highlights the importance of a One Health approach in tackling AMR.
1. Introduction
Enterococci are commensal bacteria of the intestines of humans and warm-blooded animals that may enter the general environment, where they may survive for a prolonged time period [1,2]. Enterococci may withstand environmental stressors and inhabit different ecological niches; hence, microorganisms have been isolated from a variety of sources, including aquatic and terrestrial vegetation, fresh and marine waters, sediment and soil, insects and arachnoids, fish, and foods [3,4,5,6,7,8]. Enterococci and E. coli are well-recognized faecal pollution indicators and have been isolated from waters contaminated with faecal waste and sewage. The detection of enterococci and E. coli has been suggested for the evaluation of drinking water [9].
In humans, enterococci are known as opportunistic pathogens, with E. faecalis and E. faecium being most frequently isolated in cases of hospital-acquired infections, and they are associated with high mortality rates in Europe [10,11]. The treatment of infections caused by Enterococcus species is challenging due to their intrinsic resistance to various antimicrobial agents, including cephalosporins, sulphonamides, and aminoglycosides [12]. The ability of Enterococcus spp. to acquire additional resistance via, e.g., mobile genetic elements or sporadic chromosomal mutations, raises concerns about the available therapeutical options for the treatment of enterococcal infections [12,13]. Vancomycin-resistant E. faecium (VRE) isolates have been recognized as high-priority pathogens by the World Health Organization, for which new antibiotics are urgently needed, and the increasing prevalence of VRE has been reported in hospitals in Europe [14,15].
The rise in antimicrobial resistance (AMR) in Enterococcus isolates of clinical, animal, and food origins has been widely recognized [16,17]. There is growing evidence that the environment has become an important reservoir of AMR due to contamination from clinical settings and agroecosystems. The One Health approach, which aims to interlink human, animal, and environmental health, recognizes that the transmission of antimicrobial agents, resistant microorganisms, and antimicrobial resistance genes (ARGs) facilitates the spread of AMR, with commensal microorganisms playing an important role [18,19]. Commensal microorganisms—E. coli and Enterococcus spp.—have been used as indicators of antimicrobial resistance due to their abundance in polluted environments, associations with the intestinal tract, and the easiness of isolation [20]. The ability of these microorganisms to develop and acquire AMR as a result of selective pressure, clonal spread, the dissemination of genetic elements (e.g., plasmids), and co-selection has made them suitable for the long-term monitoring of AMR trends [21]. The detection of AMR in E. coli and Enterococcus has been established as part of an AMR surveillance programme in humans, animals, and food of animal origin [22].
The environmental dispersion of antimicrobial agents via agri-food systems may facilitate the spread of ARGs, as suggested by the discovery of a close phylogenetic relationship between vancomycin-resistant E. faecium (VRE) swine faecal isolates and surface water isolates from Switzerland [23,24]. Despite AMR patterns in Enterococcus spp. from clinical and animal production sectors being reported to share sector-specific antibiotic-consumption-related traits, resistant enterococci may serve as vectors for the further dissemination of AMR in the human–animal–environment interface [23,24].
Antimicrobial resistance in Enterococcus spp. has been targeted, mainly in productive animals and products of animal origin, to monitor Enterococcus spp. antimicrobial resistance trends, but data on the prevalence of resistant enterococci in other animals are largely missing [25,26]. In recent years, wildlife has been reported as a source of multidrug-resistant (MDR) Enterococcus spp. [27]. It has been suggested that wildlife could serve as important sentinels of antimicrobial resistance in the environment because their habitats have not been directly exposed to antimicrobial agents, and the presence of AMR-associated indicators such as ARGs has been linked to a spillover from human and agricultural settings [28]. Hence, the occurrence of resistant and MDR strains may indicate the current AMR distribution trends in the environment [29].
Carnivorous species have been reported as being more likely to carry resistant microorganisms [30]. The red fox (Vulpes vulpes) is a generalist predator with a wide geographic distribution in Europe, including Latvia [31]. Red foxes are characterized by broad dietary spectra and an ability to adapt to different ecological environments, even in proximity to human habitats. Despite the importance of red foxes to the control of the wildlife population, the role of these foxes in the spread of human and animal pathogens has been recorded [32]. Red foxes have been associated with carrying MDR bacteria due to their foraging behaviour, where they may acquire resistant microorganisms or ARGs from human and animal waste [33]. Since data on antimicrobial resistance in wildlife, including wild carnivores, are missing, the aim of the present study was to detect antimicrobial resistance, resistance genes, and genetic diversity among Enterococcus spp. isolates from red foxes (Vulpes vulpes).
2. Results
2.1. Antimicrobial Resistance in Enterococcus Isolates
Altogether, 34 Enterococcus isolates were obtained, which were represented by E. faecium (n = 17), E. faecalis (n = 12), E. durans (n = 3), and E. hirae (n = 2).
Antimicrobial resistance was identified in 44% (15/34) of the Enterococcus spp. using the minimum inhibitory concentration (MIC) detection method. The highest antimicrobial resistance rates were to tetracycline (18%, 6/34), erythromycin (9%, 3/34), ciprofloxacin (6%, 2/34), tigecycline (6%, 2/34), and daptomycin (6%, 2/34). No isolates were resistant to ampicillin, chloramphenicol, gentamicin, linezolid, teicoplanin, or vancomycin. E. faecium exhibited resistance to five out of the twelve antimicrobial agents tested, with 10 out the 17 isolates (59%) being resistant to at least one antimicrobial agent. The lowest rates of antimicrobial resistance were recorded for E. faecalis, with one out of twelve strains (8%) being resistant to tetracycline (Table 1).

Table 1.
Antimicrobial resistance in Enterococcus isolates (n = 34) from foxes.
2.2. Distribution of Antimicrobial Resistance Genes (ARGs) in Enterococcus Isolates
A total of 12 ARGs in 32 Enterococcus isolates were identified, and ARGs encoding resistance to aminoglycosides, tetracyclines, and macrolide–lincosamide–streptogramin AB were detected. Two out of the thirty-four Enterococcus isolates were excluded from further analysis due to low coverage (<30%, 30266 E. hirae) or a failure to meet the identification threshold (>90% of target genes, 3802 E. faecalis).
At least one ARG was present in every Enterococcus isolate. ARGs were most frequently identified in E. faecium (ten ARGs), while only one ARG was found in E. hirae. Among the ARGs, the aminoglycoside-resistant determinant aac(6′)-I (59%, 20/32) was the most abundant. tet(M) (13%, 4/32) and msr(C) (53%, 17/32) were the most frequently identified tetracycline- and macrolide-resistant determinants (Table 2). The ARGs exhibited by E. faecium were the ant(6)-Ia gene responsible for high-level streptomycin resistance (12%, 2/17); the tetracycline-resistance-encoding tet(M) (18%, 3/17); tet(L) (6%, 1/17); lsa(E) (6%, 1/17) of the efflux pump ABC superfamily, conferring resistance to macrolides, lincosamides, and streptogramins A; and lincosamide-resistant genes lnu(B) (6%, 1/17) and lnu(G) (6%, 1/17). E. durans shared only the aminoglycoside-resistant genes aac(6′)-I (6%, 1/17) and aac(6′)-Iih (6%, 1/17).
Table 2.
Distribution of resistance genes in Enterococcus isolates (n = 32) from red foxes.
All the Enterococcus isolates with at least one identified type of phenotypic resistance shared at least one virulence gene, and the number of ARGs in phenotypically resistant isolates ranged from one (three isolates) to eight (one isolate). Phenotypic resistance to quinupristin/dalfopristin was associated with the presence of the lsa(A) gene. Phenotypic resistance to tetracycline was confirmed with the presence of tet(M) in three E. faecium isolates, tet(M) and tet(L) simultaneously in one E. faecium isolate, and tet(S) in one E. faecalis isolate. High-level tetracycline resistance (MIC ≥ 128) was identified in two E. faecalis isolates. The tet genes were not identified in tetracycline-susceptible and tigecycline-resistant isolates (Table 3). Specific genes encoding daptomycin and ciprofloxacin resistance were not identified. In every phenotypically resistant isolate, with the exception of E. faecalis (429441), the presence of aac(6′)-I, encoding resistance to aminoglycosides, was found.
Table 3.
Association of Enterococcus spp. resistance phenotypes with the presence of ARGs.
2.3. Genetic Diversity in Enterococcus Isolates
Since there was a sufficient number of E. faecalis and E. faecium isolates, genetic diversity was explored using a core genome multilocus sequence typing (cgMLST) approach. For both species, no apparent clustering was observed (Figure 1 and Figure 2). Even within the same geographic region (municipality), the cgMLST genotypes were highly diverse, especially in the case of E. faecalis, where over 50% of the cgMLST genes showed different alleles between any of the isolates when compared. However, there were at least some seemingly more closely related isolates among E. faecium (isolates 2124 and 429-441 differed in 137/1423 alleles, while the largest difference in this dataset was between isolates 428-644 and 424-211/2, differing in 1251/1423 alleles). Two E. faecium isolates from the Tukuma and Ventspils municipalities, which are geographically more distant from the other regions, also appeared to be more genetically distant from the other regions’ isolates (Figure 2).
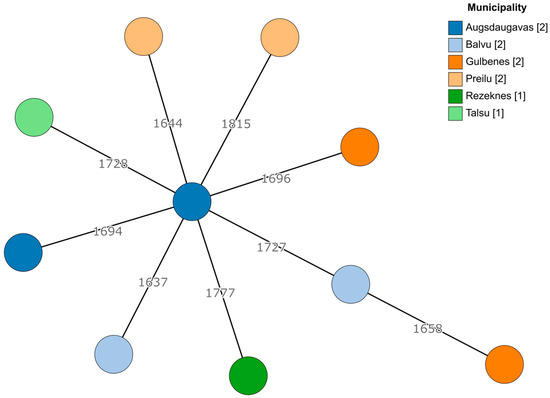
Figure 1.
A minimum spanning tree based on the cgMLST scheme of 1972 target genes for E. faecalis. Branch lengths are displayed on a logarithmic scale. The number of isolates obtained from each municipality is shown in square brackets.
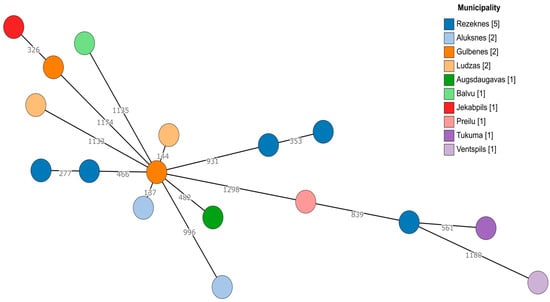
Figure 2.
A minimum spanning tree based on the cgMLST scheme of 1423 target genes for E. faecium. Branch lengths are displayed on a logarithmic scale. The number of isolates obtained from each municipality is shown in square brackets.
3. Discussion
E. faecalis, E. faecium, E. durans, and E. hirae were the main species isolated, which corresponds to previous findings on the faecal microbiota of red foxes, wild canids, and felids [34,35]. Higher isolation frequencies of E. faecium may be related to feeding behaviours, as E. faecium was more frequently carried by carnivores [36].
The percentage of identified Enterococcus spp. isolates that were resistant to at least one antimicrobial agent was lower than 73% in Portugal [37] and Italy, where 35% of all isolates were susceptible to all the tested antimicrobial agents [38]. In the present study, no MDR isolates were found that resulted in a conflicting finding with those of previous authors, who reported the prevalence of MDR in Enterococcus from 3% in red foxes in Portugal [37] to 63% in wild Pampas foxes (Lycalopex gymnocercus) in Brazil [35]. Almost every Enterococcus isolate from wildlife carnivores shares the MDR patterns identified for wildlife carnivores in Poland [29]. It has been suggested that the proximity of foxes to human habitats, rural colonization, and the urbanization of foxes sharing the same habitat as humans may promote the distribution of antimicrobial resistance and infectious agents [37,39]. In the present study, the samples mostly originated from remote areas with a low population density; thus, limited access to antimicrobial exposure may have reduced the ability of the faecal microbiota to develop antimicrobial resistance [40].
The identified high rates of antimicrobial resistance in Enterococcus spp. to tetracycline, erythromycin, and ciprofloxacin were in agreement with the findings of previous research on Carnivora [36,37,38]. Resistance to tetracyclines and ciprofloxacin in Enterococcus spp. was found in isolates from wild Pampas foxes, as well as Geoffroy’s cats in Brazil and red foxes in Portugal [34,35]. The AMR patterns of Enterococcus spp. isolates from wild animals may resemble domestic animal, food, and clinical isolates [38,41], and they were found to be in line with antimicrobial consumption data from productive animals in Latvia [42]. The antimicrobial resistance of E. faecium in calves against quinupristin/dalfopristin has been reported in Latvia [43].
Resistance to glycopeptides (vancomycin and daptomycin) and other last-resort antimicrobial agents was not identified in the present study, with the exception of tigecycline resistance in the E. hirae and E. durans isolates. Enterococcus spp. share intrinsic resistance to different classes of antibiotics, including cephalosporins, aminoglycosides, and trimethoprim–sulfamethoxazole [23]. Resistance to first-line antibiotics such as ampicillin and quinolones has been reported in clinical isolates. Acquired resistance to the glycopeptide vancomycin in E. faecalis and E. faecium significantly limits the choice of antimicrobial agents available for the treatment of vancomycin-resistant Enterococcus infections [8]. Oxazolidinones (linezolid), novel tetracyclines (tigecycline), and lipopeptides (daptomycin) may be used for the treatment of vancomycin-resistant Enterococcus infections [44]. Therefore, the monitoring of antimicrobial resistance to critically important antimicrobial agents in E. faecalis and E. faecium is important for tracking the dissemination of antimicrobial resistance in human and animal populations. Despite E. hirae and E. durans representing a low clinical significance, the identification of tigecycline-resistant Enterococcus spp. is concerning, considering the ability of Enterococcus to recruit antimicrobial resistance determinants.
Resistance to vancomycin was not identified in the present study. An increasing number of vancomycin-resistant enterococci have been identified in domestic animals and wildlife. In Portugal, 13.5% of the Enterococcus spp. isolates from red foxes exhibited phenotypical resistance to vancomycin, as confirmed by the presence of the van(A) and van(C-1) genes, while in Italy, high-level vancomycin resistance (MIC ≥ 1024 mg/mL) was not associated with the presence of genetic determinants [38,45]. Resistance as high as 41% was identified in wild birds in Italy [46]. A high prevalence of vancomycin-resistant Enterococcus spp. in the wildlife in Poland could be explained by possible interactions between different ecosystems [25,47].
Almost all of the Enterococcus isolates shared ARGs that confer resistance to aminoglycosides. Enterococcus spp. intrinsically exbibit low-level resistance to aminoglycosides, with aac(6′)-I enzymes being important for amikacin resistance [48]. Our findings on the widespread occurrence of the aac(6′) gene in E. faecium genomes are in agreement with those of Zaheer et al. [23]. aac(6′)-Ii-like genes specific to E. hirae and E. durans, which confer resistance to the synergy of the association of penicillin and tobramycin, were identified in the present study [49]. The genes conferring resistance to gentamicin include aph(2′)-Ib, aph(2′)-Ic, aph(2″)-Id, and the high-level gentamicin-resistance (HLGR) gene aac(6′)-Ie-aph(2’’)-Ia which inactivates all aminoglycosides with the exception of streptomycin. Genes with high-level resistance to streptomycin and kanamycin have not been identified, in contrast to previous studies indicating an overall low spread of ARGs in the environment and wildlife in Latvia [29,34,50,51].
tet(M) was the predominant gene in our study among tetracycline-resistant isolates, and was among the most frequently identified genes conferring resistance to tetracyclines [52]. tet(M) was among the most abundant ARGs in red foxes in Spain and other carnivores—otters (Lutra lutra) in Portugal, racoon dogs (Nyctereutes procyonoides Gray, 1834), American minks (Neovison vison Schreber, 1777), and beech martens (Martes foina) in Poland [29,37,50]. Two E. faecalis isolates showed high-level resistance to tetracyclines, with a MIC ≥ 128, and one isolate with resistance to tetracycline was associated with the presence of the high-level streptomycin resistance gene ant(6)-Ia and the tet(M) and tet(L) genes. The tet(M) gene confers antimicrobial resistance via ribosomal protection, while tet(L) mediates the efflux of tetracycline from cells [53]. The tet(M) gene was associated with conjugative transposons of the Tn916/Tn1545 family which facilitate the conjugation process via horizontal ARG transfer. Tn916/Tn1545 transposon can promote the transfer of tetM, ermB, aph(3″)-IIIa, or other transposons or plasmids to ensure horizontal gene transfer [54]. Thus, the presence of high-level Enterococcus resistance isolates in red foxes may indicate the opportunity for ARG dissemination in wildlife via AMR transfer from highly populated areas to remote locations, with further spread in wild animals and transfer to human habitats. This demonstrates the need to develop a comprehensive understanding of AMR dissemination in wildlife in order to implement a One Health approach for tackling AMR.
While traditional tetracycline determinants such as tet(M) and tet(L) have been associated with the increased transcription and expansion of gene copy numbers in tigecycline-resistant phenotypes [55,56], the determinants of resistance to tetracycline in tigecycline-resistant E. hirae and E. durans isolates were not identified in our study.
msr(C) was most abundant in E. faecium isolates (94%), and it is thought to be species-specific. The msrA,B,C genes were found in both erythromycin-resistant and -susceptible isolates [38]. The mrs(C) gene of E. faecium may encode the efflux pump of macrolides [57], and it was identified in all the erythromycin-resistant isolates in the present study. The lsa(E) and msr(C) genes are reported to be important for lincosamine, streptogramin A, and macrolide resistance, with the lsa genes involved in the development of resistance to clindamycin, lincosamides, and dalfopristin [58]. The lnu(B) gene confers resistance to lincosamides only, and Nowakiewicz et al. have suggested its circulation between farm animals and wildlife [59].
The investigated isolates of E. faecalis and E. faecium were highly diverse, as assessed using cgMLST typing, and the high diversity of Enterococcus isolates from environmental and animal sources has been reported previously [60]. This indicates the absence of the clonal distribution of Enterococcus strains among the fox population in the observed geographical region. The genotypes between and within municipalities were highly variable in most cases. The only case indicating the possibility of a common genetic background involved E. faecium isolates 428-644 and 426-804, both coming from the Rezekne municipality and displaying resistance against ciprofloxacin. Still, they were separated by a significant 353-allele difference based on their cgMLST genotypes. However, continued monitoring with more dense sampling would be beneficial for assessing the distribution of genetic diversity among resistant Enterococcus species in wild carnivore populations.
The present study reveals the distribution of resistant Enterococcus isolates in areas largely untouched by human activities and agricultural practices, which may have an impact on Enterococcus antimicrobial resistance rates. This study was limited by the availability of animals from a single geographic area, which is the subject of a national rabies vaccination monitoring program. Further studies including other carnivorous species are intended for the purpose of obtaining a comprehensive overview of the dissemination of antimicrobial resistance in the environment.
4. Materials and Methods
4.1. Samples and Sampling
Altogether, 32 dead red foxes (Vulpes vulpes) were delivered to the reference laboratory to monitor the efficiency of a peroral rabies vaccination program in wildlife [61,62]. Red foxes were collected from 25 parishes, with one to three foxes per parish (Table S1). The caecum of each fox was aseptically removed during a necropsy. The external surface of the caecum was disinfected with 70% ethanol and left for 5 min. Then, the caecum was aseptically incised and 1 g of the content of the caecum was collected.
4.2. Microbiological Testing of Samples
An amount of 1 g of the caecal content was transferred into 9 mL of the azide dextrose broth (Biolife, Milan, Italy) and incubated for 24 h at 37 °C. A loop of the enriched suspension was streaked onto Slanetz and Bartley agar (Biolife, Italy) and incubated for 44 ± 4 h at 44 ± 1 °C. After incubation, colonies with a typical red, maroon, or pink colour were subcultured on blood agar (Biolife, Italy) with the addition of 5% defibrinated horse blood (TCS Biosciences Ltd., Buckingham, UK) for 24 h at 37 °C. Enterococcus spp. were confirmed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOFF, Bruker, Bremen, Germany).
4.3. Detection of Antimicrobial Resistance in Enterococcus spp. Isolates
Antimicrobial resistance was detected with the microbroth dilution method using EUVENC test panels (ThermoFisher Scientific, East Grinstead, West Sussex, UK), which are used by the European Union (EU) for the surveillance of AMR in Enterococcus spp. The investigated isolates were suspended in saline (0.5 McFarland) and transferred into 11 mL of cation-adjusted Mueller–Hinton (MH) broth (Thermo Fisher Scientific, Landsmeer, Netherlands). The test panels were inoculated with the bacterial suspension in MH broth and incubated at 37 °C for 24 h. In the EUVSEC test panel, the following antimicrobial agents were included (in μg/mL): ampicillin (0.5–64), chloramphenicol (4–128), ciprofloxacin (0.12–16), daptomycin (0.25–32), erythromycin (1–128), gentamicin (8–1024), linezolid (0.5–64), quinupristin/dalfopristin (0.5–64), teicoplanin (0.5–64), tetracycline (1–128), tigecycline (0.03–4), and vancomycin (1–128). The resistance thresholds were determined in accordance with the EUCAST [63].
4.4. DNA Extraction and Whole-Genome Sequencing (WGS)
Prior to DNA extraction, the bacterial cultures were grown on blood agar (Biolife Italiana, Monza, Italy). For the DNA extraction, the commercially available kit “NucleoSpin Tissue” (Macherey-Nagel, Düren, Germany) was used. Since enterococci are Gram-positive bacteria, a protocol for hard-to-lyse bacteria was used. The protocol included an additional lysozyme step to hydrolyse the peptidoglycan glycosidic bonds in order to obtain better-quality DNA; after that, the standard protocol nr.5 was followed. The purity of the samples was assessed using a NanoDrop spectrophotometer and the quantity was measured using a Qubit fluorometer (both ThermoFisher Scientific, Landsmeer, The Netherlands).
To prepare the libraries for the whole-genome sequencing, an Illumina DNA prep (Illumina, San Diego, CA, USA) library preparation kit was used by following the provided instructions. For a sample quality assessment before pooling, the gel capillary electrophoresis (QIAxcel Advanced Instrument, QIAGEN, Venlo, Limburg, The Netherlands) was used with a high-resolution cartridge and a 50–1500 bp size marker.
The library was sequenced using the Illumina MiSeq next-generation sequencing system (Illumina, San Diego, CA, USA).
4.5. Genome Assembly, Detection of Antimicrobial Resistance Genes (ARGs) and Core Genome MLST (cgMLST)
The raw reads were assembled and antimicrobial resistance was identified using an in-house pipeline written in Snakemake (v7.32.3) [64]. First, the quality of the raw reads (and later, the trimmed reads) was assessed with FastQC (v0.12.1) [65]. Then, the raw reads were trimmed using Trimmomatic (v0.39) [66] with the following settings: 2:30:9 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:33. The assemblies were created using SPAdes (v3.15.5), with the option of using careful and paired reads as the input [67]. Afterwards, QUAST (v5.2.0) was used to assess the quality of the assemblies [68]. Then, the assemblies were indexed as references and the raw reads were aligned to them using bwa-mem2 (v2.2.1) [69]. The resulting bam files were sorted and indexed with SAMtools (v1.179) [70]. QualiMap (v2.2.2-dev) was used to assess the coverage [71]. Due to the low coverage (<30), two samples were excluded from further analyses (30266 and 427166). The contamination or misidentification of Enterococcus species was detected using Kraken 2 (v2.1.3) with the Standard-16 database (v3/14/2023) on the raw reads [72]. MultiQC (v1.14) was used to compile a report on these different statistics to make it easier to assess the different aspects of quality [73]. Finally, on the assemblies, we also ran AMRFinderPlus (3.11.2) [74] with the database version 2023-08-08.2 to detect ARGs (Supplementary Material Table S2). All of the aforementioned computations were performed on the high-performance computing centre cluster at Riga Technical University.
Afterwards, to allow the cgMLST to be determined and to create a minimum spanning tree (MST), the assembled genomes were imported into Ridom SeqSphere+ (v9.0.10) (Ridom, Muenster, Germany) [75]. We determined the cgMLST and created an MST based on the previously existing schemes for E. faecium and E. faecalis (available at https://cgmlst.org, accessed on 13 November 2023). One sample (3802) did not reach the threshold of >90% target genes, and therefore it was excluded from the MST. The MST was created in GrapeTree (v1.5.0) using the MSTreeV2 algorithm (https://genome.cshlp.org/content/28/9/1395, accessed on 10 December 2023).
5. Conclusions
The present study shows the presence of resistant Enterococcus species in red foxes in sparsely populated areas without direct anthropogenic effects. The identification of antimicrobial resistance and antimicrobial resistance determinants in Enterococcus species isolated from wild carnivores may be the result of environmental contamination and indirect contact with humans and agricultural animals, since wild predators are at the top of the food chain. The presence of resistant enterococci in red foxes highlights the importance of a One Health approach in tackling antimicrobial resistance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics13020114/s1, Table S1: Isolates included in the present study; Table S2: ARGs targeted according to AMRFinderPlus (3.11.2), with the database version 2023-08-08.2.
Author Contributions
Conceptualisation, M.T. and A.B.; methodology, J.A., J.Ķ. and A.C.; software, J.Ķ. and A.C.; validation, M.T., J.Ķ., J.A., A.C. and L.L; formal analysis, M.T., J.A., J.Ķ. and A.C.; investigation, M.T., J.Ķ., J.A., A.C. and L.L.; resources, J.A., A.B., and J.Ķ.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, J.A., J.Ķ., A.C., L.L. and A.B.; visualization, J.Ķ. and A.C.; supervision, A.B.; project administration, A.B.; funding acquisition, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the State Research Programme of Latvia project No. VPP-EM-BIOMEDICĪNA-2022/1-0001: “State research project in the field of biomedicine, medical technologies and pharmacy”.
Institutional Review Board Statement
The present experiment did not require an institutional review board statement because the dead animals were collected by the competent authority according to the rabies eradication program of the Republic of Latvia and delivered to the reference laboratory (the Research Institute of Food Safety, Animal Health, and Environment “BIOR) for necropsies.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw sequence reads from the bacterial isolates included in this study have been deposited in the European Nucleotide Archive under the study accession number PRJEB71100.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Aarestrup, F.M.; Butaye, P.; Witte, W. Non-human reservoirs of enterococci. In The Enterococci: Pathogenesis, Molecular Biology, and Antibiotic Resistance; Gilmore, M.S., Clewell, D.B., Courvalin, P., Dunny, G.M., Murray, B.E., Rice, L.B., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 55–100. [Google Scholar]
- Gilmore, M.S.; Lebreton, F.; van Schaik, W. Genomic transition of enterococci from gut commensals to leading causes of multidrug-resistant hospital infection in the antibiotic era. Curr. Opin. Microbiol. 2013, 16, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Whitman, R.L.; Shively, D.A.; Pawlik, H.; Nevers, M.B.; Byappanahalli, M.N. Occurrence of Escherichia coli and enterococci in Cladophora (Chlorophyta) in nearshore water and beach sand of Lake Michigan. Appl. Environ. Microbiol. 2003, 69, 4717–4719. [Google Scholar] [CrossRef] [PubMed]
- Staley, C.; Dunny, G.M.; Sadowsky, M.J. Environmental and animal-associated Enterococci. Adv. Appl. Microbiol. 2014, 87, 147–186. [Google Scholar] [CrossRef]
- Novais, C.; Campos, J.; Freitas, A.R.; Barros, M.; Silveira, E.; Coque, T.M.; Antunes, P.; Peixe, L. Water supply and feed as sources of antimicrobial-resistant Enterococcus spp. in aquacultures of rainbow trout (Oncorhyncus mykiss), Portugal. Sci. Total Environ. 2018, 625, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zarzecka, U.; Zadernowska, A. Enterococci isolated from plant-derived food—Analysis of antibiotic resistance and the occurrence of resistance genes. LWT 2021, 139, 110549. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Kowalczewski, P.Ł.; Babošová, M.; Porhajašová, J.I.; Hikal, W.M.; Fedoriak, M. Bacteriota and Antibiotic Resistance in Spiders. Insects 2022, 13, 680. [Google Scholar] [CrossRef] [PubMed]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Recommendations incorporating the first and second addenda. In Guidelines for Drinking-Water Quality, 3rd ed.; WHO: Geneve, Switzerland, 2008; Volume 1, p. 688. Available online: https://www.who.int/publications/i/item/9789241547611 (accessed on 10 December 2023).
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef]
- Brinkwirth, S.; Ayobami, O.; Eckmanns, T.; Markwart, R. Hospital-acquired infections caused by enterococci: A systematic review and meta-analysis, WHO European Region, 1 January 2010 to 4 February 2020. Euro Surveill. 2021, 26, 2001628. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef]
- Cimen, C.; Berends, M.S.; Bathoorn, E.; Lokate, M.; Voss, A.; Friedrich, A.W.; Glasner, C.; Hamprecht, A. Vancomycin-resistant enterococci (VRE) in hospital settings across European borders: A scoping review comparing the epidemiology in the Netherlands and Germany. Antimicrob. Resist. Infect. Control 2023, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial Resistance in the EU/EEA (EARS-Net)-Annual Epidemiological Report 2019; ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- Markwart, R.; Willrich, N.; Haller, S.; Noll, I.; Koppe, U.; Werner, G.; Eckmanns, T. The rise in vancomycin-resistant Enterococcus faecium in Germany: Data from the German Antimicrobial Resistance Surveillance (ARS). Antimicrob. Resist. Infect. Control 2019, 8, 147. [Google Scholar] [CrossRef]
- Paschoalini, B.R.; Nuñez, K.V.M.; Maffei, J.T.; Langoni, H.; Guimarães, F.F.; Gebara, C.; Freitas, N.E.; dos Santos, M.V.; Fidelis, C.E.; Kappes, R.; et al. The emergence of antimicrobial resistance and virulence characteristics in Enterococcus species isolated from bovine milk. Antibiotics 2023, 12, 1243. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, S.P.; Stokes, H.W.; Roy Chowdhury, P. Mobile elements, zoonotic pathogens and commensal bacteria: Conduits for the delivery of resistance genes into humans, production animals and soil microbiota. Front. Microbiol. 2023, 4, 86. [Google Scholar] [CrossRef] [PubMed]
- Lawpidet, P.; Tengjaroenkul, B.; Saksangawong, C.; Sukon, P. Global prevalence of vancomycin-resistant enterococci in food of animal origin: A meta-analysis. Foodborne Pathog. Dis. 2021, 18, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Dungan, R.S.; Bjorneber, D.L. Antimicrobial resistance in Escherichia coli and enterococcal isolates from irrigation return flows in a high-desert watershed. Front. Microbiol. 2021, 12, 660697. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar] [CrossRef]
- Anjum, M.F.; Schmitt, H.B.; Börjesson, S.; Berendonk, T.U.; on behalf of the WAWES network. The potential of using E. coli as an indicator for the surveillance of antimicrobial resistance (AMR) in the environment. Curr. Opin. Microbiol. 2021, 64, 152–158. [Google Scholar] [CrossRef]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef]
- Biggel, M.; Nüesch-Inderbinen, M.; Raschle, S.; Stevens, M.J.A.; Stephan, R. Spread of vancomycin-resistant Enterococcus faecium ST133 in the aquatic environment in Switzerland. J. Glob. Antimicrob. Resist. 2021, 27, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Decision (EU) 2020/1729 of 17 November 2020 on the Monitoring and Reporting of Antimicrobial Resistance in Zoonotic and Commensal Bacteria and Repealing Implementing Decision 2013/652/EU. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32020D1729 (accessed on 1 December 2023).
- Malik, F.; Nawaz, M.; Anjum, A.A.; Firyal, S.; Shahid, M.A.; Irfan, S.; Ahmed, F.; Bhatti, A.A. Molecular characterization of antibiotic resistance in poultry gut origin Enterococci and horizontal gene transfer of antibiotic resistance to Staphylococcus aureus. Pak. Vet. J. 2022, 42, 383–389. [Google Scholar] [CrossRef]
- Grassotti, T.T.; de Angelis Zvoboda, D.; da Fontoura Xavier Costa, L.; de Araújo, A.J.G.; Pereira, R.I.; Soares, R.O.; Wagner, P.G.C.; Frazzon, J.; Frazzon, A.P.G. Antimicrobial resistance profiles in Enterococcus spp. isolates from fecal samples of wild and captive black capuchin monkeys (Sapajus nigritus) in South Brazil. Front. Microbiol. 2018, 9, 2366. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as sentinels of antimicrobial resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [CrossRef] [PubMed]
- Nowakiewicz, A.; Zięba, P.; Gnat, S.; Trościeńczyk, A.; Osińska, M.; Łagowski, D.; Kosior-Korzecka, U.; Puzio, I. A significant number of multi-drug resistant Enterococcus faecalis in wildlife animals; Long-term consequences and new or known reservoirs of resistance. Sci. Total Environ. 2020, 705, 135830. [Google Scholar] [CrossRef] [PubMed]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthier-Clerc, M.; et al. Antimicrobial Resistance in Wildlife. J. Appl. Ecol. 2016, 53, 519–529. [Google Scholar] [CrossRef]
- Díaz-Ruiz, F.; Delibes-Mateos, M.; García-Moreno, J.L.; López-Martín, J.M.; Ferreira, C.; Ferrers, P. Biogeographical patterns in the diet of an opportunistic predator: The red fox Vulpes vulpes in the Iberian Peninsula. Mammal Rev. 2013, 43, 59–70. [Google Scholar] [CrossRef]
- Lindsø, L.K.; Dupont, P.; Rød-Eriksen, L.; Andersskog, I.P.Ø.; Ulvund, K.R.; Flagstad, Ø.; Bischof, R.; Eide, N.E. Estimating red fox density using non-invasive genetic sampling and spatial capture–recapture modelling. Oecologia 2022, 198, 139–151. [Google Scholar] [CrossRef]
- Mo, S.S.; Urdahl, A.M.; Madslien, K.; Sunde, M.; Nesse, L.L.; Slettemeås, J.S.; Norströn, M. What does the fox say? Monitoring antimicrobial resistance in the environment using wild red foxes as an indicator. PLoS ONE 2018, 13, e0198019. [Google Scholar] [CrossRef]
- Radhouani, H.; Igrejas, G.; Conçalves, A.; Pacheco, R.; Monteiro, R.; Sargo, R.; Brito, F.; Torres, C.; Poeta, P. Antimicrobial resistance and virulence genes in Escherichia coli and enterococci from red foxes (Vulpes vulpes). Anaerobe 2013, 23, 82–86. [Google Scholar] [CrossRef]
- Oliveira de Araujo, G.; Huff, R.; Favarini, M.O.; Mann, M.B.; Peters, F.B.; Frazzon, J.; Guedes Frazzon, A.P. Multidrug resistance in Enterococci isolated from wild Pampas foxes (Lycalopex gymnocercus) and Geoffroy’s cats (Leopardus geoffroyi) in the Brazilian Pampa Biome. Front. Vet. Sci. 2020, 7, 606377. [Google Scholar] [CrossRef] [PubMed]
- García, L.A.; Torres, C.; López, A.R.; Rodríguez, C.O.; Valencia, C.S. Antimicrobial resistance of Enterococcus species isolated from wild mammals in Aragón, Spain. J. Vet. Res. 2022, 66, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.; Hipólito, D.; Figueiredo, A.; Fonseca, C.; Caetano, T.; Mendo, S. Unravelling the diversity and abundance of the red fox (Vulpes vulpes) faecal resistome and their phenotypic antibiotic susceptibility of indicator bacteria. Animals 2022, 12, 2572. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Stępień-Pyśniak, D.; Gnat, S.; Fratini, F.; Urban-Chmiel, R.; Cerri, R.; Winiarczyk, S.; Turchi, B. Antibiotic susceptibility and virulence genes in Enterococcus isolates from wild mammals living in Tuscany, Italy. Microb. Drug Resist. 2020, 26, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Víchová, B.; Bona, M.; Miterpáková, M.; Kralijk, J.; Čabanová, V.; Nemčíková, G.; Hurníková, Z.; Oravec, M. Fleas and ticks of red foxes as vectors of canine bacterial and parasitic pathogens, in Slovakia, Central Europe. Vector Borne Zoonotic Dis. 2018, 18, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Central Statistical Bureau, Republic of Latvia. Usually Resident Population Density in Regions, Cities and Towns, Municipalities, and Rural Territories, at the Beginning of Year 2022–2023. Available online: https://data.stat.gov.lv/pxweb/en/OSP_PUB/START__POP__IR__IRD/IRD062/ (accessed on 1 December 2023).
- Busani, L.; Del Grosso, M.; Paladini, C.; Graziani, C.; Pantosti, A.; Biavasco, F.; Caprioli, A. Antimicrobial susceptibility of vancomycin-susceptible and -resistant enterococci isolated in Italy from raw meat products, farm animals, and human infections. Int. J. Food Microbiol. 2004, 97, 17–22. [Google Scholar] [CrossRef] [PubMed]
- EMA/299538/2023; Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2022. European Medicines Agency, European Surveillance of Veterinary Antimicrobial Consumption: Amsterdam, The Netherlands, 2022.
- Terentjeva, M.; Streikiša, M.; Avsejenko, J.; Trofimova, J.; Kovaļenko, K.; Elferts, D.; Bērziņš, A. Prevalence and antimicrobial resistance of Escherichia coli, Enterococcus spp. and the major foodborne pathogens in calves in Latvia. Foodborne Pathog. Dis. 2019, 16, 35–41. [Google Scholar] [CrossRef]
- Zhou, X.; Willems, R.J.L.; Friedrich, A.W.; Rossen, J.W.A.; Bathoorn, E. Enterococcus faecium: From microbiological insights to practical recommendations for infection control and diagnostics. Antimicrob. Resist. Infect. Control 2020, 9, 130. [Google Scholar] [CrossRef]
- Radhouani, H.; Igrejas, G.; Carvalho, C.; Pinto, L.; Gonçalves, A.; Lopez, M.; Sargo, R.; Cardoso, L.; Martinho, A.; Rego, V.; et al. Clonal lineages, antibiotic resistance and virulence factors in vancomycin-resistant Enterococci isolated from fecal samples of red foxes (Vulpes Vulpes). J. Wildl. Dis. 2011, 47, 769–773. [Google Scholar] [CrossRef]
- Cagnoli, G.; Bertelloni, F.; Interrante, P.; Ceccherelli, R.; Marzoni, M.; Ebani, V.V. Antimicrobial-resistant Enterococcus spp. in wild avifauna from central Italy. Antibiotics 2022, 11, 852. [Google Scholar] [CrossRef]
- Nowakiewicz, A.; Ziołkowska, G.; Zieba, P.; Kostruba, A. Undomesticated animals as a reservoir of multidrug resistant Enterococcus in eastern Poland. J. Wildl. Dis. 2014, 50, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef] [PubMed]
- del Campo, R.; Galán, J.C.; Tenorio, C.; Ruiz-Garbajosa, P.; Zarazaga, M.; Torres, C.; Baquero, F. New aac(6′)-I genes in Enterococcus hirae and Enterococcus durans: Effect on β-lactam/aminoglycoside synergy. J. Antimicrob. Chemother. 2005, 55, 1053–1055. [Google Scholar] [CrossRef] [PubMed]
- Semedo-Lamsaddek, T.; Nóbrega, C.S.; Ribeiro, T.; Pedroso, N.M.; Sales-Luís, T.; Lemsaddek, A.; Tenreiro, R.; Tavares, L.; Vilela, C.L.; Oliveira, M. Virulence traits and antibiotic resistance among enterococci isolated from Eurasian otter (Lutra lutra). Vet. Microbiol. 2013, 163, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Shete, V.; Grover, N.; Kumar, M. Analysis of aminoglycoside modifying enzyme genes responsible for high-level aminoglycoside resistance among Enterococcal isolates. J. Pathog. 2017, 64, 3256952. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Agerso, Y.; Smidt, P.G.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Huys, G.; D’Haene, K.; Collard, J.M.; Swings, J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl. Environ. Microbiol. 2004, 70, 1555–1562. [Google Scholar] [CrossRef]
- Roberts, A.P.; Mullany, P. A modular master on the move: The Tn916 family of mobile genetic elements. Trends Microbiol. 2009, 17, 251–258. [Google Scholar] [CrossRef]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid encoded tetracycline determinants tet(L) and tet(M). J. Antimicrob. Chemother. 2016, 71, 871–881. [Google Scholar] [CrossRef]
- Miller, W.R.; Murray, B.E.; Rice, L.B.; Arias, C.A. Resistance in vancomycin-resistant enterococci. Infect. Dis. Clin. N. Am. 2020, 34, 751–771. [Google Scholar] [CrossRef]
- Singh, K.V.; Malathum, K.; Murray, B.E. Disruption of an Enterococcus faecium species-specific gene, a homologue of acquired macrolide resistance genes of staphylococci, is associated with an increase in macrolide susceptibility. Antimicrob. Agents Chemother. 2001, 45, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.N.; Elleboudy, N.S.; El-Housseuny, G.S.; Yassien, M.A. Cloning and sequencing of lsaE efflux pump gene from MDR enterococci and its role in erythromycin resistance. Infect. Genet. Evol. 2021, 94, 105010. [Google Scholar] [CrossRef] [PubMed]
- Nowakiewicz, A.; Ziółkowska, G.; Zięba, P.; Gnat, S.; Trościańczyk, A.; Adaszek, Ł. Characterization of multidrug resistant E. faecalis strains from pigs of local origin by ADSRRS-fingerprinting and MALDI -TOF MS; evaluation of the compatibility of methods employed for multidrug resistance analysis. PLoS ONE 2017, 12, e0171160. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.S.; Lee, S.M.; Gan, H.M.; Dykes, G.A.; Rahman, S. Genetic diversity of Enterococcus faecalis isolated from environmental, animal and clinical sources in Malaysia. J. Infect. Public Health 2017, 10, 617–623. [Google Scholar] [CrossRef]
- Cabinets of Ministers. Cabinet Regulations No.178. Procedures for the Prevention and Combating Rabies. Available online: https://likumi.lv/ta/en/en/id/205795 (accessed on 1 December 2023).
- Food and Veterinary Service. State Program for Surveillance and Control of Rabies. 2021. Available online: https://www.pvd.gov.lv/lv/media/1109/download (accessed on 1 December 2023).
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical Breakpoints. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 1 December 2023).
- Mölder, F.; Jablonski, K.P.; Letcher, B.; Hall, M.B.; Tomkins-Tinch, C.H.; Sochat, V.; Forster, J.; Lee, S.; Twardziok, S.O.; Kanitz, A.; et al. Sustainable Data Analysis with Snakemake. F1000Res 2021, 10, 33. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 December 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Vasimuddin, M.; Misra, S.; Li, H.; Aluru, S. Efficient architecture-aware acceleration of BWA-MEM for multicore systems. In Proceedings of the 2019 IEEE International Parallel and Distributed Processing Symposium (IPDPS), Rio de Janeiro, Brazil, 20–24 May 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 314–324. [Google Scholar]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the reference gene catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; Von Haeseler, A.; Stoye, J.; et al. Updating Benchtop Sequencing Performance Comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).