
Evolution of Resistance against Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in the Environmental Opportunistic Pathogen Stenotrophomonas maltophilia


Abstract
1. Introduction
2. Results
2.1. Experimental Evolution in the Presence of Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole Leads to High Levels of Resistance in S. maltophilia
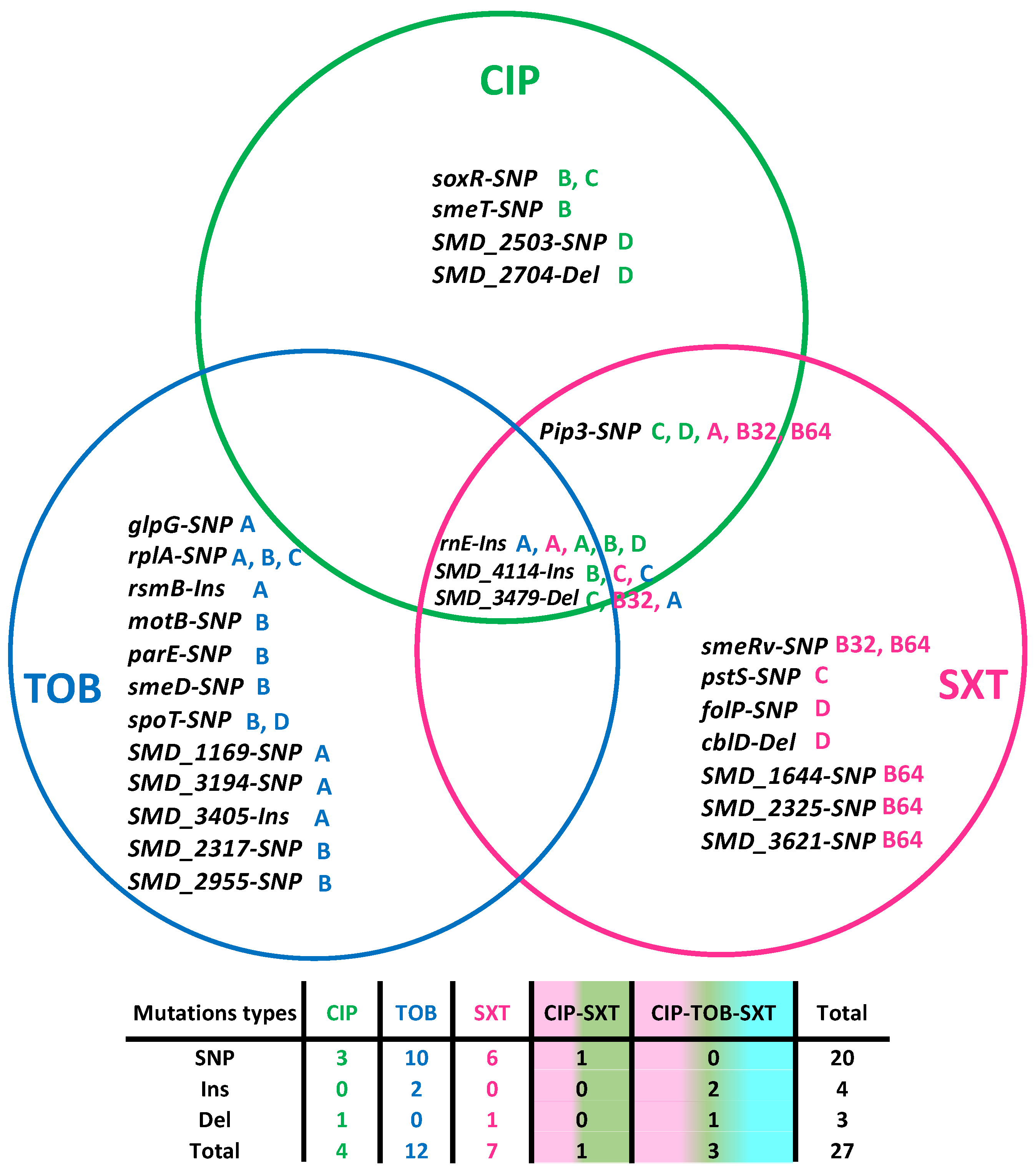
2.2. Mutations Selected in the Presence of Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in S. maltophilia D457
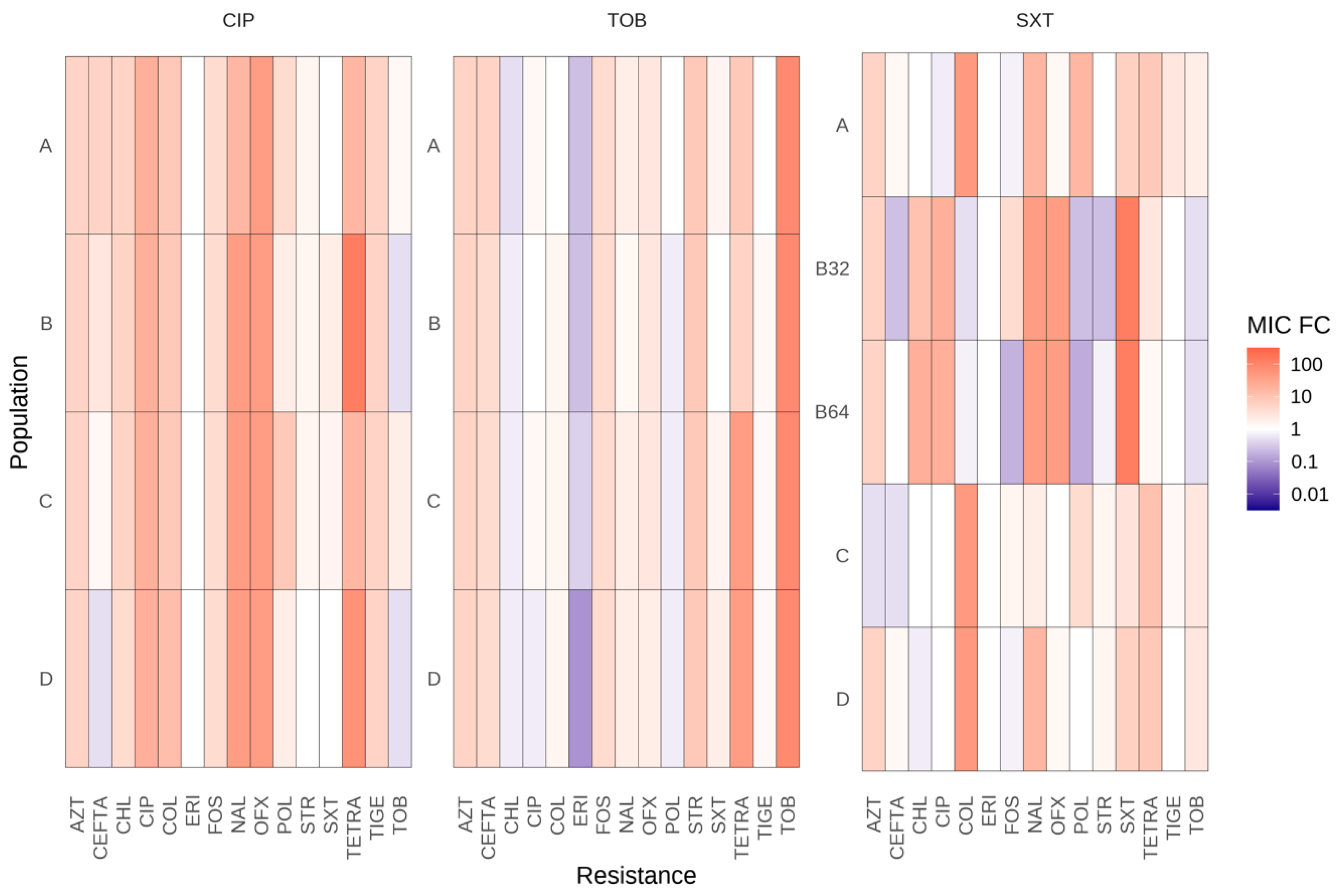
2.3. Adaptative Trajectories, Cross-Resistance, and Collateral Sensitivity of Evolved Populations
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Experimental Evolution
4.3. DNA Extraction and Whole-Genome Sequencing
4.4. Identification of Mutations
4.5. Antimicrobial Susceptibility Assays
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sanz-García, F.; Gil-Gil, T.; Laborda, P.; Ochoa-Sánchez, L.E.; Martínez, J.L.; Hernando-Amado, S. Coming from the Wild: Multidrug Resistant Opportunistic Pathogens Presenting a Primary, Not Human-Linked, Environmental Habitat. Int. J. Mol. Sci. 2021, 22, 8080. [Google Scholar] [CrossRef] [PubMed]
- Rogues, A.M.; Maugein, J.; Allery, A.; Fleureau, C.; Boulestreau, H.; Surcin, S.; Bebear, C.; Janvier, G.; Gachie, J.P. Electronic Ventilator Temperature Sensors as a Potential Source of Respiratory Tract Colonization with Stenotrophomonas maltophilia. J. Hosp. Infect. 2001, 49, 289–292. [Google Scholar] [CrossRef] [PubMed]
- De Mauri, A.; Torreggiani, M.; Chiarinotti, D.; Andreoni, S.; Molinari, G.; De Leo, M. Stenotrophomonas maltophilia: An Emerging Pathogen in Dialysis Units. J. Med. Microbiol. 2014, 63, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Stigt, J.A.; Wolfhagen, M.J.; Smulders, P.; Lammers, V. The Identification of Stenotrophomonas maltophilia Contamination in Ultrasound Endoscopes and Reproduction of Decontamination Failure by Deliberate Soiling Tests. Respiration 2015, 89, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Riquena, B.; Monte, L.d.F.V.; Lopes, A.J.; da Silva-Filho, L.V.R.F.; Damaceno, N.; Aquino, E.d.S.; Marostica, P.J.C.; Ribeiro, J.D. Microbiological Contamination of Nebulizers Used by Cystic Fibrosis Patients: An Underestimated Problem. J. Bras. Pneumol. 2019, 45, e20170351. [Google Scholar] [CrossRef] [PubMed]
- Zainulabid, U.A.; Siew, S.W.; Musa, S.M.; Soffian, S.N.; Periyasamy, P.; Ahmad, H.F. Whole-Genome Sequence of a Stenotrophomonas maltophilia Isolate from Tap Water in an Intensive Care Unit. Microbiol. Resour. Announc. 2023, 12, e00995-22. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Berg, G. Stenotrophomonas maltophilia. Trends Microbiol. 2018, 26, 637–638. [Google Scholar] [CrossRef]
- Li, X.; McLaughlin, R.W.; Grover, N.A. Characterization of Antibiotic-Resistant Stenotrophomonas Isolates from Painted Turtles Living in the Wild. Curr. Microbiol. 2023, 80, 93. [Google Scholar] [CrossRef]
- Jägevall, S.; Rabe, L.; Pedersen, K. Abundance and Diversity of Biofilms in Natural and Artificial Aquifers of the Äspö Hard Rock Laboratory, Sweden. Microb. Ecol. 2011, 61, 410–422. [Google Scholar] [CrossRef]
- Ochoa-Sánchez, L.E.; Vinuesa, P. Evolutionary Genetic Analysis Uncovers Multiple Species with Distinct Habitat Preferences and Antibiotic Resistance Phenotypes in the Stenotrophomonas maltophilia Complex. Front. Microbiol. 2017, 8, 1548. [Google Scholar] [CrossRef]
- Yassin, A.; Huralska, M.; Pogue, J.M.; Dixit, D.; Sawyer, R.G.; Kaye, K.S. Executive Summary: State-of-the-Art Review: State of the Management of Infections Caused by Multidrug-Resistant Gram-Negative Organisms. Clin. Infect. Dis. 2023, 77, 1223–1225. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.B. Antibiotic Resistance in the Opportunistic Pathogen Stenotrophomonas maltophilia. Front. Microbiol. 2015, 6, 140723. [Google Scholar] [CrossRef] [PubMed]
- Rello, J.; Kalwaje Eshwara, V.; Lagunes, L.; Alves, J.; Wunderink, R.G.; Conway-Morris, A.; Rojas, J.N.; Alp, E.; Zhang, Z. A Global Priority List of the TOp TEn Resistant Microorganisms (TOTEM) Study at Intensive Care: A Prioritization Exercise Based on Multi-Criteria Decision Analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Z. Role of the Acetyltransferase AAC(6′)-Iz Modifying Enzyme in Aminoglycoside Resistance in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2003, 51, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Rojas, P.; Garcia, E.; Calderón, G.M.; Ferreira, F.; Rosso, M. Successful Treatment of Stenotrophomonas maltophilia Meningitis in a Preterm Baby Boy: A Case Report. J. Med. Case Rep. 2009, 3, 7389. [Google Scholar] [CrossRef]
- Toleman, M.A.; Bennett, P.M.; Bennett, D.M.C.; Jones, R.N.; Walsh, T.R. Global Emergence of Trimethoprim/Sulfamethoxazole Resistance in Stenotrophomonas maltophilia Mediated by Acquisition of Sul Genes. Emerg. Infect. Dis. 2007, 13, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Ruiz, F.M.; Romero, A.; Martínez, J.L. The Binding of Triclosan to SmeT, the Repressor of the Multidrug Efflux Pump SmeDEF, Induces Antibiotic Resistance in Stenotrophomonas maltophilia. PLoS Pathog. 2011, 7, e1002103. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L.; Baquero, F.; Andersson, D.I. Predicting Antibiotic Resistance. Nat. Rev. Microbiol. 2007, 5, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Gil-Gil, T.; Martínez, J.L.; Blanco, P. Mechanisms of Antimicrobial Resistance in Stenotrophomonas maltophilia: A Review of Current Knowledge. Expert Rev. Anti Infect. Ther. 2020, 18, 335–347. [Google Scholar] [CrossRef]
- Li, X.-Z.; Nikaido, H. Efflux-Mediated Drug Resistance in Bacteria. Drugs 2004, 64, 159–204. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.-Z.; Poole, K. Multiple Antibiotic Resistance in Stenotrophomonas maltophilia: Involvement of a Multidrug Efflux System. Antimicrob. Agents Chemother. 2000, 44, 287–293. [Google Scholar] [CrossRef]
- Redgrave, L.S.; Sutton, S.B.; Webber, M.A.; Piddock, L.J.V. Fluoroquinolone Resistance: Mechanisms, Impact on Bacteria, and Role in Evolutionary Success. Trends Microbiol. 2014, 22, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Campoli-Richards, D.M.; Monk, J.P.; Price, A.; Benfield, P.; Todd, P.A.; Ward, A. Ciprofloxacin: A Review of Its Antibacterial Activity, Pharmacokinetic Properties and Therapeutic Use. Drugs 1988, 35, 373–447. [Google Scholar] [CrossRef] [PubMed]
- Laponogov, I.; Veselkov, D.A.; Crevel, I.M.-T.; Pan, X.-S.; Fisher, L.M.; Sanderson, M.R. Structure of an ‘Open’ Clamp Type II Topoisomerase-DNA Complex Provides a Mechanism for DNA Capture and Transport. Nucleic Acids Res. 2013, 41, 9911–9923. [Google Scholar] [CrossRef]
- Shariati, A.; Arshadi, M.; Khosrojerdi, M.A.; Abedinzadeh, M.; Ganjalishahi, M.; Maleki, A.; Heidary, M.; Khoshnood, S. The Resistance Mechanisms of Bacteria against Ciprofloxacin and New Approaches for Enhancing the Efficacy of This Antibiotic. Front. Public Health 2022, 10, 1025633. [Google Scholar] [CrossRef] [PubMed]
- Valdezate, S.; Vindel, A.; Saéz-Nieto, J.A.; Baquero, F.; Cantón, R. Preservation of Topoisomerase Genetic Sequences during in Vivo and in Vitro Development of High-Level Resistance to Ciprofloxacin in Isogenic Stenotrophomonas maltophilia Strains. J. Antimicrob. Chemother. 2005, 56, 220–223. [Google Scholar] [CrossRef]
- Vicens, Q.; Westhof, E. Crystal Structure of a Complex between the Aminoglycoside Tobramycin and an Oligonucleotide Containing the Ribosomal Decoding A Site. Chem. Biol. 2002, 9, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D.; Berghuis, A.M.; Mobashery, S. Aminoglycoside Antibiotics. Structures, Functions, and Resistance. Adv. Exp. Med. Biol. 1998, 456, 27–69. [Google Scholar] [PubMed]
- Davies, J.; Davis, B.D. Misreading of Ribonucleic Acid Code Words Induced by Aminoglycoside Antibiotics. The Effect of Drug Concentration. J. Biol. Chem. 1968, 243, 3312–3316. [Google Scholar] [CrossRef]
- Walsh, C. Molecular Mechanisms That Confer Antibacterial Drug Resistance. Nature 2000, 406, 775–781. [Google Scholar] [CrossRef]
- Kotra, L.P.; Haddad, J.; Mobashery, S. Aminoglycosides: Perspectives on Mechanisms of Action and Resistance and Strategies to Counter Resistance. Antimicrob. Agents Chemother. 2000, 44, 3249–3256. [Google Scholar] [CrossRef]
- Kemnic, T.R.; Coleman, M. Trimethoprim Sulfamethoxazole. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- García-León, G.; Ruiz De Alegría Puig, C.; García De La Fuente, C.; Martínez-Martínez, L.; Martínez, J.L.; Sánchez, M.B. High-Level Quinolone Resistance Is Associated with the Overexpression of smeVWX in Stenotrophomonas maltophilia Clinical Isolates. Clin. Microbiol. Infect. 2015, 21, 464–467. [Google Scholar] [CrossRef] [PubMed]
- García-León, G.; Salgado, F.; Oliveros, J.C.; Sánchez, M.B.; Martínez, J.L. Interplay between Intrinsic and Acquired Resistance to Quinolones in Stenotrophomonas maltophilia. Environ. Microbiol. 2014, 16, 1282–1296. [Google Scholar] [CrossRef] [PubMed]
- Mardle, C.E.; Shakespeare, T.J.; Butt, L.E.; Goddard, L.R.; Gowers, D.M.; Atkins, H.S.; Vincent, H.A.; Callaghan, A.J. A Structural and Biochemical Comparison of Ribonuclease E Homologues from Pathogenic Bacteria Highlights Species-Specific Properties. Sci. Rep. 2019, 9, 7952. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, F.S.L.; Schoofs, G.; Hancock, R.E.W.; De Mot, R. Influence of a Putative ECF Sigma Factor on Expression of the Major Outer Membrane Protein, OprF, in Pseudomonas aeruginosa and Pseudomonas fluorescens. J. Bacteriol. 1999, 181, 4746–4754. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, Y.J.; Lee, H.B.; Seok, Y.-J.; Lee, C.-R. Genetic Evidence for Distinct Functions of Peptidoglycan Endopeptidases in Escherichia coli. Front. Microbiol. 2020, 11, 565767. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium; Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bye-A-Jee, H.; et al. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Folcher, M.; Morris, R.P.; Dale, G.; Salah-Bey-Hocini, K.; Viollier, P.H.; Thompson, C.J. A Transcriptional Regulator of a Pristinamycin Resistance Gene in Streptomyces Coelicolor. J. Biol. Chem. 2001, 276, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Koutsolioutsou, A.; Peña-Llopis, S.; Demple, B. Constitutive soxR Mutations Contribute to Multiple-Antibiotic Resistance in Clinical Escherichia coli Isolates. Antimicrob. Agents Chemother. 2005, 49, 2746–2752. [Google Scholar] [CrossRef]
- Burrowes, E.; Abbas, A.; O’Neill, A.; Adams, C.; O’Gara, F. Characterisation of the Regulatory RNA RsmB from Pseudomonas aeruginosa PAO1. Res. Microbiol. 2005, 156, 7–16. [Google Scholar] [CrossRef]
- Morimoto, Y.V.; Namba, K.; Minamino, T. GFP Fusion to the N-Terminus of MotB Affects the Proton Channel Activity of the Bacterial Flagellar Motor in Salmonella. Biomolecules 2020, 10, 1255. [Google Scholar] [CrossRef]
- Sorlozano, A.; Gutierrez, J.; Jimenez, A.; De Dios Luna, J.; Martínez, J.L. Contribution of a New Mutation in parE to Quinolone Resistance in Extended-Spectrum-β-Lactamase-Producing Escherichia coli Isolates. J. Clin. Microbiol. 2007, 45, 2740–2742. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nathania, I.; Nainggolan, I.M.; Yasmon, A.; Nusatia, A.C.M.; Tjoa, E.; Gunardi, W.D.; Moehario, L.H. Hotspots Sequences of gyrA, gyrB, parC, and parE Genes Encoded for Fluoroquinolones Resistance from Local Salmonella Typhi Strains in Jakarta. BMC Microbiol. 2022, 22, 250. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zheng, M.; Xu, J.; Wang, M.; Jia, R.; Chen, S.; Liu, M.; Zhao, X.; Yang, Q.; Wu, Y.; et al. Prevalence of Fluoroquinolone Resistance and Mutations in the gyrA, parC and parE Genes of Riemerella anatipestifer Isolated from Ducks in China. BMC Microbiol. 2019, 19, 271. [Google Scholar] [CrossRef] [PubMed]
- Ronneau, S.; Hallez, R. Make and Break the Alarmone: Regulation of (p)ppGpp Synthetase/Hydrolase Enzymes in Bacteria. FEMS Microbiol. Rev. 2019, 43, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Martinez, J.L. Expression of Multidrug Efflux Pump SmeDEF by Clinical Isolates of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2001, 45, 1879–1881. [Google Scholar] [CrossRef]
- Buwembo, W.; Aery, S.; Rwenyonyi, C.M.; Swedberg, G.; Kironde, F. Point Mutations in the folP Gene Partly Explain Sulfonamide Resistance of Streptococcus mutans. Int. J. Microbiol. 2013, 2013, 367021. [Google Scholar] [CrossRef]
- Feirer, N.; Fuqua, C. Pterin Function in Bacteria. Pteridines 2017, 28, 23–36. [Google Scholar] [CrossRef]
- Aguena, M.; Ferreira, G.M.; Spira, B. Stability of the pstS Transcript of Escherichia coli. Arch. Microbiol. 2009, 191, 105–112. [Google Scholar] [CrossRef]
- Sajjan, U.S.; Xie, H.; Lefebre, M.D.; Valvano, M.A.; Forstner, J.F. Identification and Molecular Analysis of Cable Pilus Biosynthesis Genes in Burkholderia cepacia. Microbiology 2003, 149, 961–971. [Google Scholar] [CrossRef]
- Nejad, A.J.; Shahrokhi, N.; Nielsen, P.E. Targeting of the Essential acpP, ftsZ, and Rne Genes in Carbapenem-Resistant Acinetobacter baumannii by Antisense PNA Precision Antibacterials. Biomedicines 2021, 9, 429. [Google Scholar] [CrossRef]
- Georgellis, D.; Arvidson, S.; Von Gabain, A. Decay of ompA mRNA and Processing of 9S RNA Are Immediately Affected by Shifts in Growth Rate, but in Opposite Manners. J. Bacteriol. 1992, 174, 5382–5390. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cullen, P.A.; Haake, D.A.; Adler, B. Outer Membrane Proteins of Pathogenic Spirochetes. FEMS Microbiol. Rev. 2004, 28, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Rozner, M.; Nukarinen, E.; Wolfinger, M.T.; Amman, F.; Weckwerth, W.; Bläsi, U.; Sonnleitner, E. Rewiring of Gene Expression in Pseudomonas aeruginosa During Diauxic Growth Reveals an Indirect Regulation of the MexGHI-OpmD Efflux Pump by Hfq. Front. Microbiol. 2022, 13, 919539. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.H.; Greenberg, J.T.; Demple, B. Posttranscriptional Repression of Escherichia coli OmpF Protein in Response to Redox Stress: Positive Control of the micF Antisense RNA by the soxRS Locus. J. Bacteriol. 1993, 175, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.F.; Sulavik, M.C. Overlaps and Parallels in the Regulation of Intrinsic Multiple-Antibiotic Resistance in Escherichia coli. Mol. Microbiol. 1996, 21, 441–448. [Google Scholar] [CrossRef] [PubMed]
- White, D.G.; Goldman, J.D.; Demple, B.; Levy, S.B. Role of the acrAB Locus in Organic Solvent Tolerance Mediated by Expression of marA, soxS, or robA in Escherichia coli. J. Bacteriol. 1997, 179, 6122–6126. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Q.; Wang, R.; Zhang, Y.; Wang, X.; Wang, H. Global Regulator SoxR Is a Negative Regulator of Efflux Pump Gene Expression and Affects Antibiotic Resistance and Fitness in Acinetobacter baumannii. Medicine 2017, 96, e7188. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Davenet, S.; Marcon, E.; Leflon-Guibout, V.; Lavigne, J.-P.; Bert, F.; Moreau, R.; Nicolas-Chanoine, M.-H. In Vitro Selection of ramR and soxR Mutants Overexpressing Efflux Systems by Fluoroquinolones as Well as Cefoxitin in Klebsiella Pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Martínez, J.L. Cloning and Characterization of SmeDEF, a Novel Multidrug Efflux Pump from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2000, 44, 3079–3086. [Google Scholar] [CrossRef]
- Calvopiña, K.; Dulyayangkul, P.; Avison, M.B. Mutations in Ribosomal Protein RplA or Treatment with Ribosomal Acting Antibiotics Activates Production of Aminoglycoside Efflux Pump SmeYZ in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2020, 64, e01524-19. [Google Scholar] [CrossRef]
- Potrykus, K.; Cashel, M. (P)ppGpp: Still Magical? Annu. Rev. Microbiol. 2008, 62, 35–51. [Google Scholar] [CrossRef]
- Sy, J. Reversibility of the Pyrophosphoryl Transfer from ATP to GTP by Escherichia coli Stringent Factor. Proc. Natl. Acad. Sci. USA 1974, 71, 3470–3473. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, L.F.; Liu, L.; Kant, S.; Kim, J.-S.; Till, J.K.; Jones-Carson, J.; Porwollik, S.; McClelland, M.; Vazquez-Torres, A. SpoT Induces Intracellular Salmonella Virulence Programs in the Phagosome. mBio 2020, 11, e03397-19. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Bhadra, R.K. (P)ppGpp Metabolism and Antimicrobial Resistance in Bacterial Pathogens. Front. Microbiol. 2020, 11, 563944. [Google Scholar] [CrossRef]
- Clemmer, K.M.; Sturgill, G.M.; Veenstra, A.; Rather, P.N. Functional Characterization of Escherichia coli GlpG and Additional Rhomboid Proteins Using an aarA Mutant of Providencia Stuartii. J. Bacteriol. 2006, 188, 3415–3419. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Linares, J.F.; López, J.A.; Camafeita, E.; Albar, J.P.; Rojo, F.; Martínez, J.L. Overexpression of the Multidrug Efflux Pumps MexCD-OprJ and MexEF-OprN Is Associated with a Reduction of Type III Secretion in Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, H.; O’Callaghan, J.; O’Grady, E.P.; Adams, C.; O’Gara, F. The Posttranscriptional Regulator RsmA Plays a Role in the Interaction between Pseudomonas aeruginosa and Human Airway Epithelial Cells by Positively Regulating the Type III Secretion System. Infect. Immun. 2006, 74, 3012–3015. [Google Scholar] [CrossRef]
- Blanco, P.; Corona, F.; Martínez, J.L. Involvement of the RND Efflux Pump Transporter SmeH in the Acquisition of Resistance to Ceftazidime in Stenotrophomonas maltophilia. Sci. Rep. 2019, 9, 4917. [Google Scholar] [CrossRef]
- Lamarche, M.G.; Dozois, C.M.; Daigle, F.; Caza, M.; Curtiss, R.; Dubreuil, J.D.; Harel, J. Inactivation of the Pst System Reduces the Virulence of an Avian Pathogenic Escherichia coli O78 Strain. Infect. Immun. 2005, 73, 4138–4145. [Google Scholar] [CrossRef]
- Gil-Marqués, M.L.; Labrador Herrera, G.; Miró Canturri, A.; Pachón, J.; Smani, Y.; Pachón-Ibáñez, M.E. Role of PstS in the Pathogenesis of Acinetobacter baumannii under Microaerobiosis and Normoxia. J. Infect. Dis. 2020, 222, 1204–1212. [Google Scholar] [CrossRef]
- Kheirandish, F.; Bandehpour, M.; Davoudi, N.; Mosaffa, N.; Dawood, S.; Kazemi, B.; Haghighi, A.; Khamesipour, A.; Masjedi, H.; Mohebali, M.; et al. Gene Regulation of Pteridine Reductase 1 in Leishmania promastigotes and Amastigotes Using a Full-Length Antisense Construct. Iran. J. Parasitol. 2013, 8, 190–196. [Google Scholar] [PubMed]
- Hernando-Amado, S.; Laborda, P.; Valverde, J.R.; Martínez, J.L. Mutational Background Influences P. aeruginosa Ciprofloxacin Resistance Evolution but Preserves Collateral Sensitivity Robustness. Proc. Natl. Acad. Sci. USA 2022, 119, e2109370119. [Google Scholar] [CrossRef] [PubMed]
- Lira, F.; Hernández, A.; Belda, E.; Sánchez, M.B.; Moya, A.; Silva, F.J.; Martínez, J.L. Whole-Genome Sequence of Stenotrophomonas maltophilia D457, a Clinical Isolate and a Model Strain. J. Bacteriol. 2012, 194, 3563–3564. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Terminology Relating to Methods for the Determination of Susceptibility of Bacteria to Antimicrobial Agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.A.; Peltzer, A.; Fillinger, S.; Patel, H.; Alneberg, J.; Wilm, A.; Garcia, M.U.; Di Tommaso, P.; Nahnsen, S. The Nf-Core Framework for Community-Curated Bioinformatics Pipelines. Nat. Biotechnol. 2020, 38, 276–278. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Population | Day 0 | Day 3 | Day 6 | Day 9 | Day 12 | Day 15 | Day 18 | Day 21 | |
|---|---|---|---|---|---|---|---|---|---|
| Ciprofloxacin | CIP-A | 0.75 | 6 | 8 | >32 | >32 | >32 | >32 | >32 |
| CIP-B | 0.75 | 8 | 8 | >32 | >32 | >32 | >32 | >32 | |
| CIP-C | 0.75 | 8 | 8 | >32 | >32 | >32 | >32 | >32 | |
| CIP-D | 0.75 | 8 | 8 | >32 | >32 | >32 | >32 | >32 | |
| Tobramycin | TOB-A | 4 | 16 | >256 | >256 | >256 | >256 | >256 | >256 |
| TOB-B | 4 | 16 | >256 | >256 | >256 | >256 | >256 | >256 | |
| TOB-C | 4 | >256 | >256 | >256 | >256 | >256 | >256 | >256 | |
| TOB-D | 4 | 48 | >256 | >256 | >256 | >256 | >256 | >256 | |
| Sulfamethoxazole-trimethoprim (SXT) | SXT-A | 0.5 | 0.5 | 0.75 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 |
| SXT-C | 0.5 | 0.5 | 0.75 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | |
| SXT-D | 0.5 | 0.5 | 0.75 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | |
| SXT-B32 | 0.5 | 0.5 | 0.75 | 1.5 | >32 | >32 | >32 | >32 | |
| SXT-B64 | 0.25 | 0.5 | 0.75 | 1.5 | >32 | >32 | >32 | >32 |
| L | Gene | Product | Localization | Type | Nucleotide Change | Amino Acid Change | Frequency (%) | Domain | |
|---|---|---|---|---|---|---|---|---|---|
| Ciprofloxacin | A | rnE | Rne/Rng family ribonuclease | 3139248 | Ins | A⟶ACCGAGCTGGGTG | N486Fs | 98 | Ribonuclease E |
| B | soxR | Redox-sensitive transcriptional activator | 1129982 | SNP | C⟶T | R45W | 100 | Helix-Turn-Helix DNA binding | |
| smeT | Efflux transporter SmeDEF transcriptional repressor | 4099641 | SNP | T⟶A | L166Q | 100 | PRK10668 DNA binding | ||
| rnE | Rne/Rng family ribonuclease | 3139248 | Ins | A⟶ACCGAGCTGGGTG | N486Fs | 99 | Ribonuclease E | ||
| SMD_4114 | S9 family peptidase | 4619837 | Ins | A⟶AGTG | H773Fs | 99 | DAP2 peptidase | ||
| C | pip3 | Prolyl aminopeptidase | 830195 | SNP | C⟶A | A46N | 57 | ||
| soxR | Redox-sensitive transcriptional activator | 1129982 | SNP | C⟶T | R45W | 90 | Helix-Turn-Helix DNA binding | ||
| SMD_3479 | YiiG family protein | 3881832 | Del | GGGA⟶G | P195Fs | 99 | DUF3829 | ||
| D | pip3 | Prolyl aminopeptidase | 830195 | SNP | C⟶A | A46N | 47 | ||
| SMD_2503 | ESPR-type extended signal peptide-containing protein | 2791884 | SNP | A⟶C | G893G | 96 | |||
| SMD_2704 | Hypothetical protein | 3006994 | Del | CAAACA⟶C | Q276Fs | 99 | |||
| rnE | Rne/Rng family ribonuclease | 3139248 | Ins | A⟶ACCGAGCTGGGTG | N486Fs | 80 | Ribonuclease E | ||
| Tobramycin | A | glpG | Rhomboid family intramembrane serine protease | 412958 | SNP | T⟶C | T17A | 100 | Membrane-associated serine protease |
| rplA | 50S ribosomal protein L1 | 884707 | SNP | T⟶G | F22C | 100 | Ribosomal L1 bact | ||
| SMD_1169 | GNAT family N-acetyltransferase | 1297246 | SNP | A⟶G | Q28P | 95 | C0G3818 acetyltransferase | ||
| SMD_3194 | ATP-binding protein | 3550846 | SNP | C⟶G | R1017P | 9 | |||
| rnE | Rne/Rng family ribonuclease | 3139248 | Ins | A⟶ACCGAGCTGGGTG | N486Fs | 90 | Ribonuclease E | ||
| SMD_3405 | DUF2339 domain-containing protein | 3798031 | Ins | C⟶CTCTGGCGGCCGG | A47Fs | 99 | DUF2339 | ||
| SMD_3479 | YiiG family protein | 3881834 | Del | GAAT⟶G | T194Fs | 100 | DUF3829 | ||
| rsmB | 16S rRNA (cytosine(967)-C(5))-methyltransferase | 4243528 | Ins | G⟶GC | R191Fs | 90 | PRK10901 | ||
| B | rplA | 50S ribosomal protein L1 | 885157 | SNP | A⟶G | H172R | 93 | Ribosomal L1 bact | |
| motB | Flagellar motor protein | 1081733 | SNP | A⟶T | L197Q | 95 | |||
| parE | DNA topoisomerase IV subunit B | 1823237 | SNP | G⟶T | R291Q | 94 | PRK05559 | ||
| SMD_2317 | ABC transporter six-transmembrane domain-containing protein | 2575602 | SNP | G⟶A | R287Q | 92 | |||
| SMD_2955 | PepSY-associated TM helix domain-containing protein | 3271344 | SNP | A⟶C | A466A | 91 | |||
| smeD | Multidrug efflux RND transporter periplasmic adaptor subunit | 4098218 | SNP | G⟶A | Q235K | 92 | PRK15030 | ||
| spoT | Bifunctional (p)ppGpp synthetase/guanosine-3’,5’-bis(diphosphate) 3’-pyrophosphohydrolase | 3846537 | Ins | A⟶ACAGGCGGCG | T712Fs | 99 | SpoT superfamily | ||
| C | rplA | 50S ribosomal protein L1 | 884715 | SNP | G⟶A | A25T | 73 | Ribosomal L1 bact | |
| SMD_4114 | S9 family peptidase | 4619837 | Ins | A⟶AGTC | H773Fs | 99 | DAP2 peptidase | ||
| D | spoT | Bifunctional (p)ppGpp synthetase/guanosine-3’,5’-bis(diphosphate) 3’-pyrophosphohydrolase | 3847032 | SNP | C⟶T | G547S | 100 | SpoT superfamily | |
| SXT | A | pip3 | Prolyl aminopeptidase | 830195 | SNP | C⟶A | A46N | 70 | |
| rnE | Rne/Rng family ribonuclease | 3139248 | Ins | A⟶ACCGAGCTGGGTG | N486Fs | 99 | Ribonuclease E | ||
| B32 | pip3 | Prolyl aminopeptidase | 830195 | SNP | C⟶A | A46N | 55 | ||
| smeRv | LysR family transcriptional regulator | 1936539 | SNP | C⟶T | G266D | 100 | C-terminal domain of LysR | ||
| SMD_3479 | YiiG family protein | 3881831 | SNP | G⟶T | P195H | 100 | DUF3829 | ||
| B64 | pip3 | Prolyl aminopeptidase | 830195 | SNP | C⟶A | A46N | 53 | ||
| SMD_1644 | DUF47 family protein | 1818431 | SNP | T⟶G | L205W | 100 | YkaA | ||
| smeRv | LysR family transcriptional regulator | 1936735 | SNP | T⟶A | N201K | 100 | C-terminal domain of LysR | ||
| SMD_2325 | Hypothetical protein | 2584531 | SNP | T⟶C | N50E | 100 | |||
| SMD_3621 | Pteridine reductase | 4056328 | SNP | T⟶G | V179G | 96 | PRK09135 | ||
| C | pstS | Phosphate ABC transporter substrate-binding protein | 1551867 | SNP | C⟶T | W215R | 92 | Periplasmic Binding Protein Type 2 | |
| SMD_4114 | S9 family peptidase | 4619837 | Ins | A⟶AGTG | H773Fs | 99 | DAP2 peptidase | ||
| D | folP | Dihydropteroate synthase | 1841316 | SNP | G⟶C | G151A | 87 | DHPS | |
| cblD | CfaE/CblD family pilus tip adhesin | 3834921 | Del | ATGTACTT⟶A | Q206Fs | 99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochoa-Sánchez, L.E.; Martínez, J.L.; Gil-Gil, T. Evolution of Resistance against Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in the Environmental Opportunistic Pathogen Stenotrophomonas maltophilia. Antibiotics 2024, 13, 330. https://doi.org/10.3390/antibiotics13040330
Ochoa-Sánchez LE, Martínez JL, Gil-Gil T. Evolution of Resistance against Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in the Environmental Opportunistic Pathogen Stenotrophomonas maltophilia. Antibiotics. 2024; 13(4):330. https://doi.org/10.3390/antibiotics13040330
Chicago/Turabian StyleOchoa-Sánchez, Luz Edith, José Luis Martínez, and Teresa Gil-Gil. 2024. "Evolution of Resistance against Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in the Environmental Opportunistic Pathogen Stenotrophomonas maltophilia" Antibiotics 13, no. 4: 330. https://doi.org/10.3390/antibiotics13040330
APA StyleOchoa-Sánchez, L. E., Martínez, J. L., & Gil-Gil, T. (2024). Evolution of Resistance against Ciprofloxacin, Tobramycin, and Trimethoprim/Sulfamethoxazole in the Environmental Opportunistic Pathogen Stenotrophomonas maltophilia. Antibiotics, 13(4), 330. https://doi.org/10.3390/antibiotics13040330