Abstract
The emergence of Candida auris as a multidrug-resistant fungal pathogen represents a significant global health challenge, especially given the growing issue of antifungal drug resistance. This review aims to illuminate the potential of essential oils (EOs), which are volatile plant secretions containing complex mixtures of chemicals, as alternative antifungal agents to combat C. auris, thus combining traditional insights with contemporary scientific findings to address this critical health issue. A systematic literature review was conducted using the PubMed, Scopus, and Web of Science databases from 2019 to 2024, and using the Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) protocol to identify relevant studies on the antifungal efficacy of EOs or their components against C. auris. Of the 90 articles identified, 16 were selected for detailed review. The findings highlight the diverse mechanisms of action of EOs and their components, such as disrupting fungal cell membranes, inducing the production of reactive oxygen species (ROS), and impeding biofilm formation, suggesting that some of them may be as effective as, or better than, traditional antifungal drugs while potentially limiting the development of resistance. However, issues such as variability in the composition of EOs and a paucity of clinical trials have been identified as significant obstacles. In conclusion, EOs and their active ingredients are emerging as viable candidates for creating effective treatments for C. auris, underscoring their importance as alternative or complementary antifungal agents in the face of increasing drug resistance. The call for future research underscores the need for clinical trials and standardization to unlock the full antifungal potential of EOs against C. auris.
1. Introduction
In recent times, the global healthcare system has experienced an amplified burden due to the emergence of multidrug-resistant pathogens. Among these, C. auris is particularly notable due to its alarming resistance patterns to common antifungal agents used to treat these infections and its associated high mortality rates. First detected in 2009, C. auris has demonstrated rapid global dissemination and has been identified in 35 countries [1]. The yeast identification methods used by laboratories often misidentify C. auris as other yeasts, which makes the detection and control of this pathogen particularly challenging. C. auris transmission occurs in healthcare settings, even in those with rigorous infection prevention and control measures in place. The Centers for Disease Control and Prevention (CDC) categorizes this pathogen as an urgent threat in antimicrobial resistance, highlighting the critical need for innovative treatment strategies [2,3].
C. auris is distinct in its ability to efficiently spread from person to person, unlike other Candida species that typically originate from the host’s own microflora. Unlike many Candida spp., it is not among the commensal organisms of the human gastrointestinal tract, and it has a particular affinity for the skin in areas such as the axilla and groin. C. auris can quickly colonize individuals following exposure, and invasive infections may occur within days to months. Colonization can persist for several months or even indefinitely, emphasizing the importance of identifying asymptomatic carriers [4,5]. These individuals are at risk of further health complications and can also transmit the infection to others or contaminate the environment [6]. Infections resulting from C. auris exhibit a clinical presentation characterized by fungemia; skin abscess meningitis wounds; and ear, bone, or burn-wound infections [6,7].
Genomic analysis has demonstrated that a significant number of genes in C. auris are related to central metabolism, which is consistent with other pathogenic Candida spp., thereby enabling it to adapt to diverse environments. C. auris exhibits similar virulence activities to those of C. albicans, including the secretion of enzymes, iron acquisition, tissue penetration, and cell wall remodeling, although these traits may vary among strains [8]. Research has revealed that C. auris is capable of evading immune responses as it receives less targeting by neutrophils than does C. albicans [9,10]. Additionally, C. auris forms biofilms and adheres to surfaces [11]. Isolates of C. auris may aggregate, which impedes disruption and thereby promotes survival in hospitals. However, non-aggregating strains exhibit higher pathogenicity. C. auris’ thermotolerance allows it to persist in the hospital environment [8].
C. auris demonstrates a relatively low virulence compared to other Candida spp., but it poses a significant threat due to its remarkable resistance to multiple antifungal agents. This resistance is a key factor in the high mortality rate associated with C. auris [12,13]. Biofilm formation enhances resistance by trapping antifungal drugs within the extracellular matrix. Specifically, the rich mannanoglucan polysaccharide-ride content of the matrix can sequester up to 70% of triazole antifungals [12]. Some research suggests that C. auris biofilms are resistant to antifungal agents, such as fluconazole, whereas planktonic forms are less resistant to antiseptics [14,15,16]. Gene expansions linked to drug resistance and multidrug efflux pumps have been identified as factors contributing to formidable resistance in C. auris. These include mutations in specific genes responsible for resistance to azoles and echinocandins, as well as the activity of efflux pumps such as the ATP-binding cassette and major facilitator superfamily, which contribute to its remarkable azole resistance [15].
Researchers have investigated the antifungal properties of natural substances, such as EOs, which have been utilized in traditional medicine for their antimicrobial characteristics. EOs are intricate blends of volatile compounds produced by plants, and their broad-spectrum bioactivity against bacteria, fungi, and viruses has garnered considerable interest. The mechanisms through which they exert their antifungal effects are often attributed to cell membrane disruption, inhibition of cell wall formation, dysfunction of fungal mitochondria, oxidative stress induction, interaction with membrane proteins, and biofilm inhibition. These mechanisms offer a promising approach for overcoming drug resistance in pathogens [17,18].
The existing scientific literature examines the antifungal properties of EOs against a range of Candida spp., although reports on C. auris are comparatively scarce. Furthermore, the variable chemical composition of EOs due to factors such as geography, season, and plant origin presents challenges for standardization and quality control that have not yet been adequately addressed. Moreover, there is a deficiency of clinical studies assessing the safety and efficacy of EOs against C. auris, which represents a significant gap in the translation of laboratory findings into clinical practice [19].
This review offers a comprehensive summary of the current research (2019–2024) on the antifungal mechanisms of EOs against C. auris. By integrating traditional knowledge with contemporary scientific discoveries, this study sheds light on the specific mechanisms by which EOs exert their antifungal properties, including disruption of cell membranes, inhibition of cell wall formation, and inhibition of biofilms. Understanding these mechanisms is crucial for the development of targeted therapeutic interventions. Furthermore, the review identifies areas for additional research, such as the need for standardization of EO composition and the conduct of clinical trials, underscoring the importance of future research and potential clinical applications.
A comprehensive systematic literature review was conducted in accordance with the PRISMA methodology, encompassing studies published in English between 2019 and 2024. This search included PubMed, Scopus, and Web of Science, using keywords such as C. auris, EOs, and antifungal agents. The selection process comprised two stages. First, titles and abstracts were screened, followed by a full-text evaluation based on predefined inclusion and exclusion criteria. The inclusion criteria were studies published in English between 2019 and 2024. Exclusion criteria excluded studies that did not address the antifungal efficacy of EOs against human infection of C. auris, and those published before 2019 or in languages other than English. Data extraction involved collecting information from the authors, including the year of publication, study objectives, chemical composition, mechanisms of action, application areas, main findings, and conclusions. The extracted data were then synthesized and analyzed to provide a comprehensive overview. The results were categorized and presented based on the sources of EOs or compounds, mechanisms of action, and main results. No statistical analysis was performed, as this was a comprehensive literature review of scientific research. The PRISMA flow chart of the included studies is shown in Figure 1.
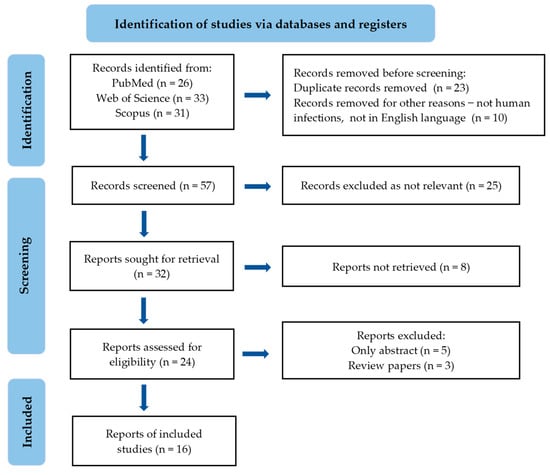
Figure 1.
PRISMA flowchart of the included studies.
2. EOs and Their Chemical Constituents with Antifungal Activity—Mechanisms of Action
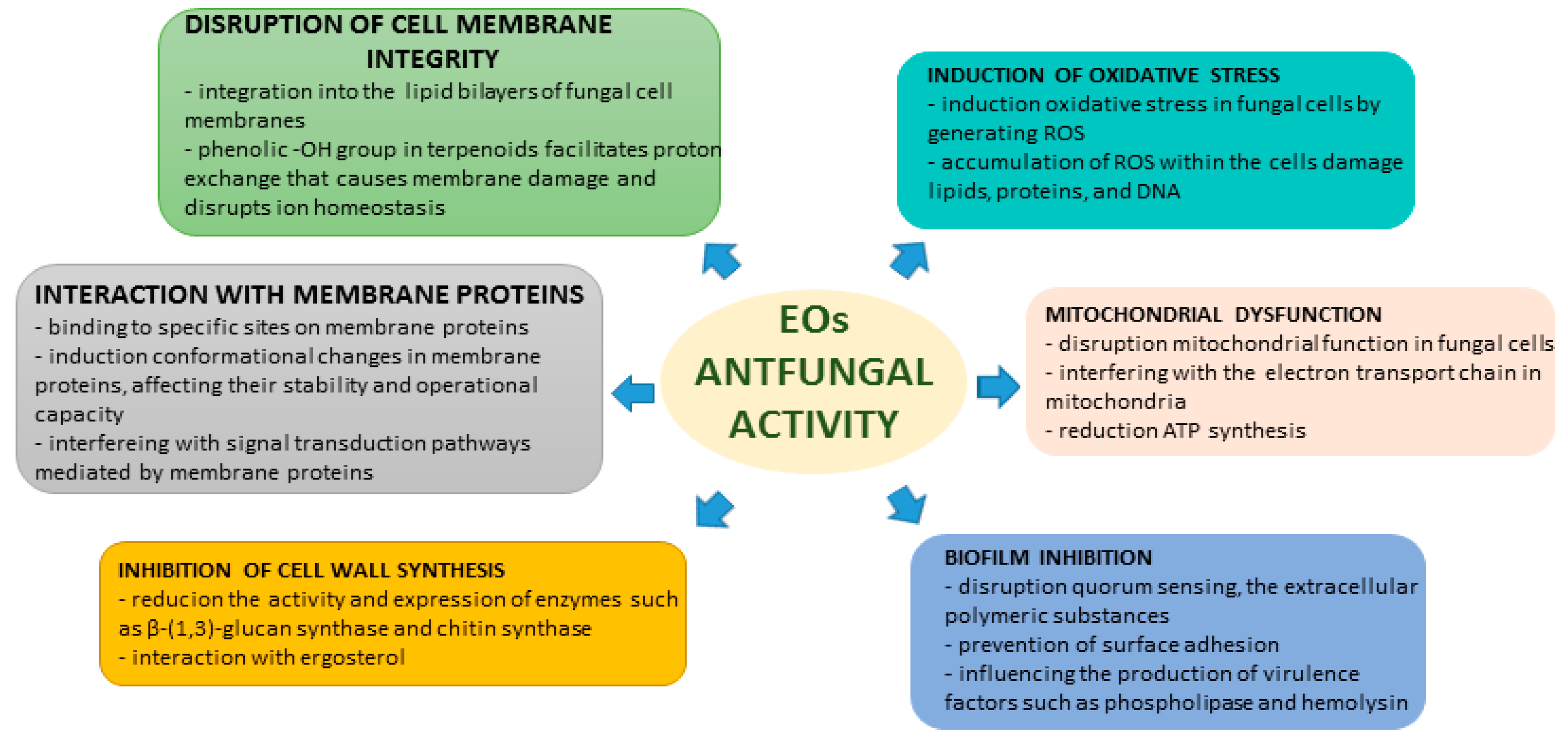
The antifungal effects of EOs and their chemical components are influenced by various mechanisms. They are composed of a diverse range of volatile compounds, such as terpenoids, aldehydes, and phenols, which exhibit extensive antimicrobial properties [17,18]. The ways in which EOs exert their antifungal effects are complex and include several primary mechanisms of action.
2.1. Disruption of Cell Membrane
EOs possess antifungal properties primarily through disrupting the integrity of cell membranes. Due to their lipophilic nature, EOs can blend with the lipid bilayers of fungal cell membranes, altering their structural and functional characteristics. Terpenoids, such as thymol, carvacrol, and eugenol, are key components in EOs that compromise the integrity of fungal cell membranes. These terpenoids contain a phenolic-OH group, which is critical for their antifungal activity. The presence of this group, with a delocalized electron system, facilitates proton exchange, resulting in membrane damage and disruption of ion homeostasis in fungal cells. The structural attributes vital to the antioxidant activity of terpenoids include the presence of a phenolic-OH group and hydrophobicity derived from the aromatic ring structure. These features are indispensable for disrupting cell membrane integrity and inducing changes that culminate in ion leakage and cell death. Additionally, the hydrophobic structure of the aromatic ring enhances penetration and damage to yeast plasma membranes, thereby augmenting their antifungal activity. The position of the-OH group within the molecule is also critical, as the phenolic-OH group attached directly to the benzene ring (forming phenol) is more effective in damaging membranes than an -OH group attached to the benzene ring via a methylene bridge [20]. Electron microscopy and transmission electron microscopy analyses were used to demonstrate that Massoia aromatica EO caused cell shrinkage, cytoplasmic leakage, and changes in cell shape of C. albicans, also increasing membrane permeability and leading to the leakage of cytosol and eventual rupture of the cell membrane [21].
2.2. Induction of Oxidative Stress
EOs can cause oxidative stress in fungal cells by producing reactive oxygen species (ROS). The accumulation of ROS can harm cellular components, such as lipids, proteins, and DNA, resulting in cellular dysfunction and death. Studies have shown that exposure of photogene to eugenol and citral increases intracellular ROS levels in a time-dependent manner, which is correlated with exposure time and contributes to the lethality observed in C. albicans. Cells exposed to eugenol at half the minimum inhibitory concentration (MIC) and hydrogen peroxide (H2O2) at all concentrations exhibited statistically significant ROS accumulation [22,23].
2.3. Mitochondrial Dysfunction
EOs exhibit an ability to target mitochondrial function, thereby impeding fungal energy metabolism. Specifically, compounds like thymol and carvacrol, which are present in EOs derived from thyme and oregano, have been demonstrated to disrupt the electron transport chain, thereby inhibiting the production of adenosine triphosphate (ATP) that is vital for the survival and proliferation of fungi. This mechanism not only restricts the energy generation of fungi, but also triggers programmed cell death pathways [22]. EOs possess the potential to hinder crucial enzymes that are involved in mitochondrial function, including those in the electron transport chain. By disrupting the functionality of these enzymes, they can impede ATP production and disrupt cellular respiration, ultimately resulting in mitochondrial dysfunction. Additionally, they can cause the permeabilization of mitochondrial membranes, leading to the release of pro-apoptotic factors like cytochrome c. This process activates apoptotic pathways in fungal cells, ultimately leading to cell death. Moreover, EOs can also alter mitochondrial metabolism by impacting processes such as oxidative phosphorylation and ATP synthesis. These alterations can disrupt the energy balance within fungal cells and contribute to their mitochondrial dysfunction [23].
2.4. Inhibition of Cell Wall Synthesis
EOs demonstrate antifungal properties by obstructing cell wall synthesis. The fungal cell wall, which primarily comprises glucans, chitin, and glycoproteins, serves as both structural support and protection. Certain components of EOs hinder the enzymes responsible for synthesizing these essential structural elements, jeopardizing cell wall integrity and culminating in fungal cell lysis. Additionally, EOs diminish ergosterol content, a vital component of the fungal cell membrane, leading to the disruption of membrane structure and functionality and ultimately resulting in cell death [24,25]. Eucalyptus EO has been shown to hinder the activity of ergosterol, which ultimately impairs membrane fluidity and performance [26]. The use of certain EO components has been found to inhibit the activity and expression of β-(1,3)-glucan synthase and chitin synthase, which are critical to the synthesis of fungal cell walls. This reduction in activity and the subsequent decrease in glucan and chitin synthesis leads to a weakening of the cell wall structure, ultimately resulting in their failure [24,26].
2.5. Interaction with Membrane Proteins
EOs not only have detrimental effects on cell membranes but can also impair the functioning of proteins associated with these membranes, such as ion channels, enzymes, and receptors. Membrane proteins that are vital for fungal survival can be adversely affected by EOs. By binding to specific sites on these proteins, EOs inhibit their activity, resulting in cellular dysfunction. EOs may cause conformational changes in membrane proteins, affecting their stability and operational capacity. These structural changes can impact the normal functioning of membrane proteins and the viability of fungal cells. Furthermore, EOs can inhibit the enzymatic activity of membrane proteins, which function as enzymes catalyzing important metabolic pathways within fungi. EOs have the potential to interfere with signal transduction pathways mediated by membrane proteins in fungal cells. By disrupting signaling mechanisms, EOs hinder intracellular communication, impairing fungal survival. Eugenol and citral have been shown to interact with amino acids in the cell membrane of Penicillium roqueforti, leading to changes in the conformation of membrane proteins that may contribute to the disruption of membrane function and integrity [26,27].
2.6. Biofilm Inhibition
Many pathogenic fungi often form biofilms, which are intricate communities encased within a matrix that exhibit heightened resistance to antifungal agents. Various studies have demonstrated that EOs can disrupt biofilm formation and eradicate previous pre-formed biofilms. For instance, thyme EO has been shown to inhibit biofilm formation by Candida albicans and to disrupt mature biofilms, presenting a promising therapeutic option for persistent fungal infections. EOs can disrupt quorum sensing, a critical communication process utilized by microorganisms to coordinate biofilm development. By interfering with quorum sensing, EOs hinder the ability of pathogens to form biofilms. EOs can also prevent C. albicans from adhering to surfaces, which is a vital step in biofilm formation. By blocking this initial attachment, EOs reduce the potential for its development. Additionally, EOs can influence the production of virulence factors, such as phospholipase and hemolysin, which are crucial for the establishment and maintenance of biofilms. By modulating these factors, they can weaken the structural integrity and defensive capabilities of biofilms. EOs have the potential to disrupt extracellular polymeric substances that maintain the structural integrity of biofilms, leading to their destabilization and potential eradication. Additionally, EOs often exhibit antimicrobial properties that can prevent and eliminate bacteria within biofilms, prevent further growth, and facilitate biofilm eradication. Certain compounds, such as limonene and eucalyptol, have been shown to inhibit biofilm formation in Candida spp. by targeting a critical factor in their pathogenicity [25,28,29]. The EO from Massoia aromatic was found to inhibit farnesol, a quorum-sensing molecule that is necessary for biofilm formation in C. albicans. This inhibition may play a role in the anti-biofilm activity of EOs [21]. A summary of the mechanisms of antifungal actions of EOs and their chemical constituents is provided in Figure 2.
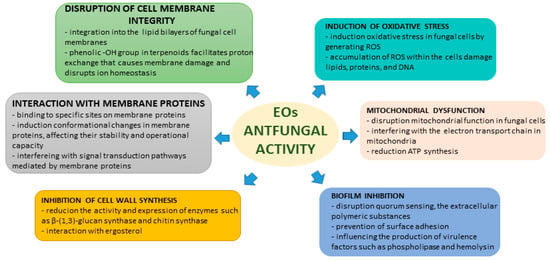
Figure 2.
EOs’ mechanisms of antifungal activity.
3. EOs and Their Compounds against C. auris
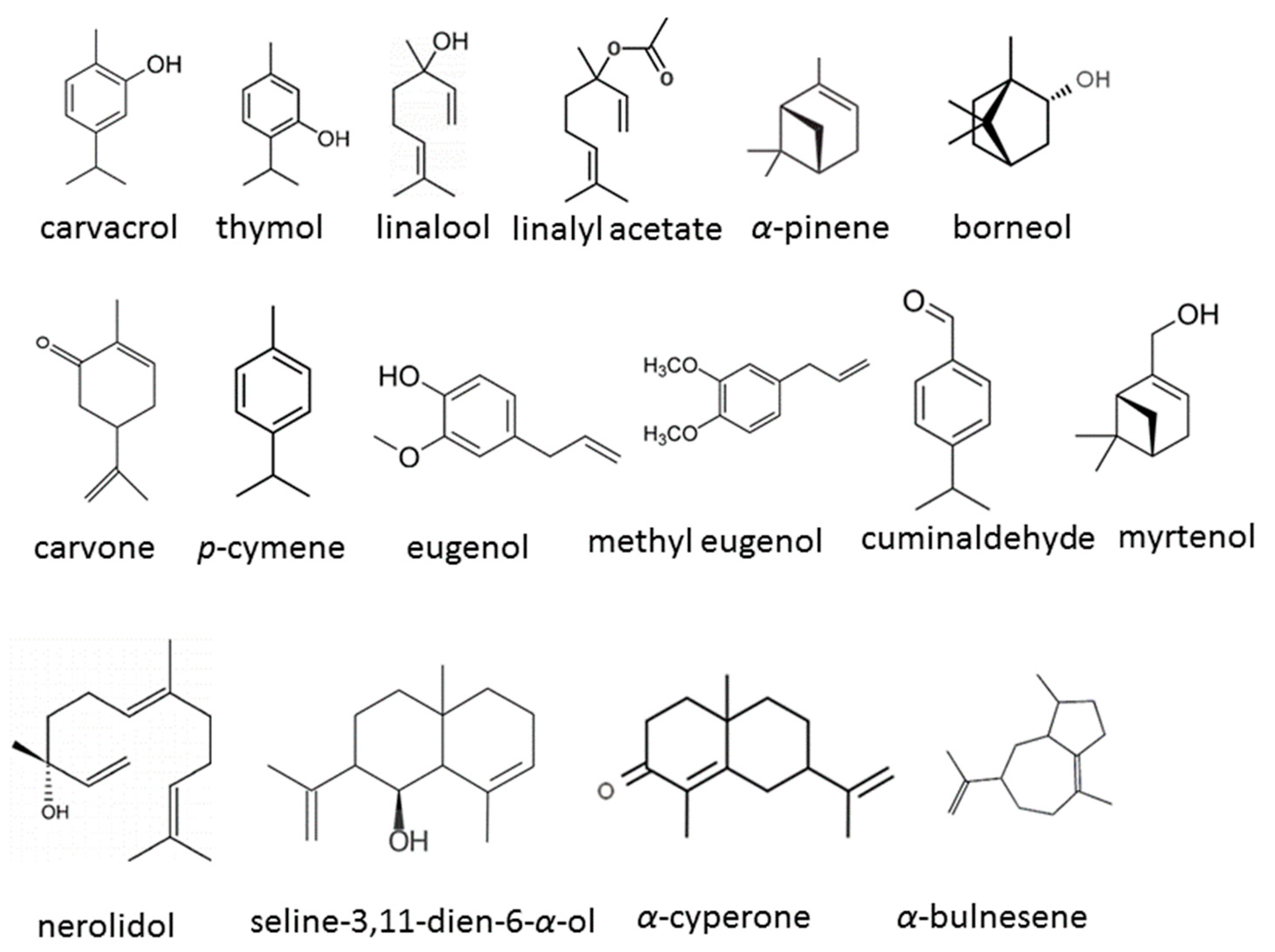
A comprehensive analysis of 16 publications from 2019 to 2024 has been carried out to elucidate the diversity, efficacy, and mechanisms of action of EOs or their components in the management of C. auris. The chemical structures of some compounds identified in these studies are depicted in Figure 3, showcasing their chemical diversity. Additionally, Table 1 presents a brief overview of the publications assessed, encapsulating the breadth and findings of the research conducted thus far.
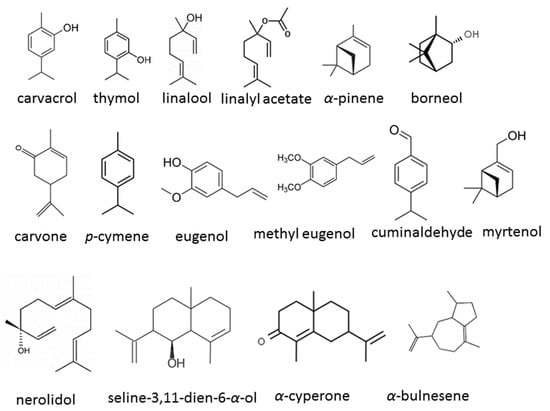
Figure 3.
Chemical structures of the bioactive compounds identified in EOs in the studies.

Table 1.
Summary of the review of the evaluated studies.
3.1. Diverse Sources of EOs and Their Compounds
Researchers have carried out extensive investigation into the antifungal properties of EOs obtained from a range of plant-based sources. [30,31,32,33,34,35,36,37,38,39,40,41,42]. These EOs encompass a wide array of bioactive compounds and exhibit potent antimicrobial activity against C. auris. Nevertheless, certain researchers have redirected their attention towards individual constituents or modified formulations to enhance antifungal properties or diminish the toxicity of particular compounds [43,44,45]. The following approach is a more efficient method of evaluation, as it allows precise assessment of the performance of individual EO components, rather than evaluating the EO as a whole, which may exhibit heterogeneity due to factors such as environmental or genetic variables (different chemotypes of the same species). By studying individual components or creating new formulations, such as liposomes, encapsulated EOs in nanostructured lipid carriers, or polycaprolactone-based nano-formulations, the researchers sought to optimize the therapeutic potential of these substances while minimizing the adverse effects of specific compounds [33,38,40]. The versatility and adaptability of plant products in the field of antifungal research are demonstrated by the diverse range of sources utilized by researchers, some of whom investigate whole EOs, whereas others examine individual compounds or employ modified formulations.
3.2. Methods of Antifungal Analysis
A multitude of techniques exist for assessing antifungal efficacy against C. auris. One widely employed method is the minimum inhibitory concentration (MIC) examination, which establishes the minimal concentration of EOs or compounds necessary to obstruct the growth of C. auris. This standardized process facilitates a quantitative evaluation of activity and offers valuable insights into the potency of the test substances. It was utilized in the majority of studies with the exception of four [30,33,37,44]. Various investigations have utilized the minimum fungicidal concentration (MFC) test, which aims to determine the minimum concentration of EOs or compounds that result in fungal cell death. This assessment extends beyond mere inhibition and examines the fungicidal activity of test substances, offering valuable insights into their capacity to eradicate populations of C. auris [31,34,38,39,41,43]. The evaluation of biofilm formation was carried out using biofilm formation assays to determine the influence of EOs or their compounds on C. auris biofilms. These assays are critical for devising effective strategies to combat fungal infections, as biofilms have significant impacts on microbial virulence and resistance [33,35,36,40,41,45]. One study utilized the disk-diffusion technique in conjunction with the paper-disc method to assess the efficacy of test substances against C. auris. This was achieved by evaluating the zone of inhibition surrounding the disks saturated with the substances in question, providing a rapid and reliable assessment of their efficacy against this organism [31,44]. Lavandula angustifolia EO-derived free and liposome-enveloped EOs have been investigated for their molecular mechanisms against C. auris using gene expression analysis in one study. This method allows researchers to determine the pathways through which natural products exert their effects by examining changes in gene expression in response to treatment [33]. Furthermore, many studies have been conducted to evaluate the potential synergistic effects between EOs and antifungal drugs. Researchers have found that the combination of these two agents can enhance the antifungal activity and reduce resistance development [34,39,40,45].
3.3. Proposed Mechanisms of EO Action against C. auris
Analyzing the processes associated with the interactions between EOs and C. auris is of paramount importance for evaluating their effectiveness. EOs possess the ability to disturb the cell membranes of C. auris due to their lipophilic nature, which may result in damage to the fungal cell membrane, disrupting its integrity and functionality [31,35]. The findings also demonstrated that the EOs caused damage to the cell membranes of C. auris, as indicated by the morphological changes observed such as the shrinking of cell surfaces and the receding of the cytoplasm, ultimately resulting in cell lysis. This damage compromised the integrity of the fungal cells and contributed to their demise. The degree of membrane damage observed in Candida cells was directly proportional to the concentration of cinnamon EOs used. Higher concentrations of EOs led to more severe damage to the cell membranes. This concentration-dependent effect suggests that EOs exert their antifungal activity by disrupting the structure and integrity of the fungal cell membrane. Cinnamon EOs contain bioactive compounds, including cinnamaldehyde and eugenol, which are known for their antimicrobial properties. These compounds possess lipophilic characteristics that enable them to penetrate the lipid bilayer of fungal cell membranes. This interaction can result in the disruption of the membrane structure, affecting its permeability and integrity. Ultimately, this disruption can lead to cell death and inhibit fungal growth. This phenomenon has been documented for other fungal pathogens as well [20,24]. EOs may exhibit antifungal properties by generating ROS, which trigger oxidative stress in fungal cells, resulting in their damage and in inhibition of their growth [33,36,45]. ROS are highly reactive molecules that can adversely affect various cellular components, such as proteins, lipids, and DNA. This oxidative stress response plays a critical role in suppressing fungal growth and biofilm formation [22]. Myrtenol, a compound isolated from plants in the genus Taxus, was observed to downregulate the expression of genes associated with biofilm formation in C. auris [45]. Specifically, the expression levels of essential genes involved in biofilm development were analyzed to elucidate the inhibitory effects of myrtenol. ERG11, a gene that plays a crucial role in ergosterol biosynthesis, was found to be downregulated by myrtenol. The disruption of ergosterol production caused by this downregulation can compromise the integrity of the fungal cell membrane, ultimately inhibiting biofilm formation. FKS1, which encodes the enzyme β-1,3-glucan synthase, is crucial for the synthesis of β-glucan, which is a key component of the fungal cell wall. By suppressing FKS1 expression, myrtenol is likely to hamper β-glucan synthesis, thereby impairing the structural integrity of the fungal cell wall and ultimately preventing biofilm formation. ALS5, a gene associated with surface adhesion, is a critical factor in the initial stages of biofilm development. The downregulation of ALS5 by myrtenol suggests that the compound disrupts C. auris’s ability to adhere to surfaces, thereby inhibiting the formation of biofilms [45]. The HOG1 gene, which participates in the oxidative stress-response pathway, exhibited variable expression patterns in response to treatment with Lavandula angustifolia EO. The study revealed an increase in HOG1 gene expression when liposome-encapsulated oil was used, suggesting activation of the oxidative stress-response pathway in C. auris biofilms. This upregulation may indicate a protective mechanism against the antifungal effects of encapsulated oil. Conversely, HOG1 was downregulated when free EO was applied, thereby potentially compromising the oxidative stress response in fungal cells. Concerning the CDR1 gene, which encodes an efflux pump, its expression changes in response to L. angustifolia EO treatment were also examined. Both the application of free L. angustifolia EO and of liposome-encapsulated oil upregulated the CDR1 gene. This alteration in gene expression may suggest a potential mechanism of resistance in C. auris biofilms treated with the EO, underscoring the need for further investigation into other genes associated with resistance [33]. This mechanism of action has also been observed in relation to cell wall synthesis, providing valuable insights into the molecular mechanisms underlying EOs’ antifungal activity [24,25,26]. EOs have demonstrated the ability to hinder the formation of biofilms in C. auris through several mechanisms of action, including the induction of oxidative stress [33,36], the downregulation of genes related to biofilm formation [33,45], and the interference or disruption of the extracellular fungal matrix that makes up the biofilm matrix [35,40]. These results align with the outcomes of other studies that have investigated the inhibitory impact of EOs on the biofilms of various pathogens [13,16,17,25,28,29]. Not all studies have specified the mechanism of action, which means that the specific ways in which the test substances affected C. auris were not clearly outlined or detailed [30,32,34,37,41,42,44]. The lack of specification regarding the mechanism of action suggests that the authors did not provide clear information on how the substances interact with the pathogen at the molecular or cellular level to inhibit its growth or virulence. This absence of a specific mechanism of action in these studies may hinder the in-depth understanding and characterization of the antimicrobial properties of the substances studied. It can also be inferred that these studies primarily focused on demonstrating the general antimicrobial activity of the substances, rather than on elucidating the exact biological pathways or targets through which they exert their effects on C. auris. In scientific research and the scientific literature, determining the mechanism of action is essential to providing insight into how antimicrobials operate, understanding their interactions with pathogens, and identifying new targets for therapeutic intervention. When the mechanism of action is not clearly defined, it can lead to gaps in understanding of how substances function against pathogens and restrict the ability to optimize efficacy or develop targeted antimicrobial strategies.
3.4. Findings
The studies conducted thus far have consistently indicated substantial antifungal effects against C. auris of the substances under investigation, with MIC and MFC values exhibiting diversity, which may be attributed to the distinct chemical positions of the analyzed EOs. Moreover, some studies have revealed the potential of the tested substances to affect C. auris biofilm formation [35,45]. The results showed a reduction in biofilm mass and interference with biofilm structures, highlighting the potential of EOs to interfere with virulence factors associated with pathogen biofilms. Synergistic interactions between the test substances and conventional antimicrobial agents were also studied [34,39,43,45]. These results indicated synergistic effects when the substances were combined with antifungal drugs, potentially increasing their effectiveness in inhibiting C. auris biofilms and overcoming drug resistance. The effects of these substances on gene expression in C. auris were investigated, revealing downregulation of genes related to fungal cell wall synthesis and upregulation of stress-response genes, providing insights into the molecular mechanisms underlying antimicrobial activity [33,45]. Some studies demonstrated an increase in ROS production in C. auris cells exposed to the test substances [33,45]. This oxidative stress response played a key role in the inhibition of fungal growth and biofilm formation, highlighting a key mechanism of action against pathogens. The effects of these substances on virulence factors of C. auris were also evaluated, which demonstrated inhibition of hemolytic activity and adhesion capacity, thus suggesting a potential role in reducing virulence and pathogenicity of the pathogen [31,43]. Considering the diverse mechanisms through which EOs and compounds exert their antimicrobial activity against C. auris, these findings underscore the potential of these natural products as promising agents for combating fungal infections.
4. Limitations, Challenges, and Future Directions
Although the antifungal properties of EOs have been documented against C. auris, there are still significant limitations to current findings. These studies were conducted in vitro, under conditions that do not always accurately mimic the complex biological environments found in vivo. The chemical complexity of EOs, which can contain hundreds of bioactive compounds, complicates the identification of the active ingredients and their mechanisms of action. This variability makes it difficult to standardize treatments and accurately predict therapeutic outcomes. Translating the in vitro antifungal efficacy of EOs into clinical applications faces several obstacles. First, their safety profiles must be carefully evaluated, as some compounds can be toxic or cause allergic reactions. The volatility and instability of EOs present additional challenges for their formulation and delivery in the clinical setting. Achieving effective concentrations without harming the host is a major challenge. As with all antimicrobial agents, the development of resistance to EOs poses a serious threat. Although they are thought to have a lower risk of inducing resistance owing to the complex mixtures of compounds that act on multiple microbial targets, there may still be cases of reduced susceptibility.
To overcome these challenges and unlock the potential of EOs for antifungal applications, a number of future research avenues can be proposed. The creation of standardized EO formulations calls for the use of advanced analytical techniques to identify and characterize their bioactive components. By doing so, stable, bioavailable, and effective products can be formulated. Cutting-edge technologies such as nanoparticles, liposomes, and emulsions can be employed to demonstrate the delivery and targeting of EOs, thus enhancing their efficacy and safety. Combining EOs with conventional antifungal drugs or other antimicrobial agents can improve their effectiveness and reduce the likelihood of resistance. High-throughput screening of EO components using existing antifungals can be employed to identify synergistic combinations. Undertaking rigorous clinical trials is necessary to confirm the efficacy and safety of EO-based therapies. Conducting comparative studies on the use of EOs and conventional antifungal therapies is crucial to establishing the therapeutic potential of these treatments.
5. Conclusions
This comprehensive review presents recent studies that have explored the impact of EOs and their constituents on C. auris, a multidrug-resistant fungal pathogen. The results of these studies indicate that the EOs and their compounds that were tested exhibited potent antifungal activity against C. auris, as evidenced by the various MIC values recorded. These compounds were found to inhibit biofilm formation and disrupt biofilm structure, which is crucial for the pathogen’s virulence and persistence. Moreover, their combination with antifungal drugs resulted in synergistic effects that enhanced their efficacy in inhibiting C. auris and potentially overcoming drug resistance. The compounds also modulated gene expression in C. auris, leading to the downregulation of biofilm-related genes and the upregulation of stress-response genes. The induction of ROS in C. auris cells by the EOs is a key factor in inhibiting the growth of this fungus. Additionally, they showed inhibitory effects on C. auris virulence factors such as hemolytic activity and adhesion capacity. The results of these studies suggest the promising potential of EOs and compounds against C. auris. However, significant challenges must be addressed in order to realize their full potential in clinical settings. These challenges include variability in chemical composition, concerns about toxicity and allergenicity, stability issues, and the lack of standardized dosing regimens. Furthermore, the mechanisms of action must be clearly defined to facilitate the development of more effective and safer therapeutic forms. Future research endeavors should concentrate on overcoming the challenges associated with EOs by expanding their chemical characterization and improving delivery systems to enhance bioavailability and reduce toxicity. Additionally, controlled clinical trials should be conducted to establish efficacy and safety profiles. Research should also focus on understanding the mechanisms underlying potential resistance to EOs and developing strategies to mitigate this risk. EOs and their constituents possess significant potential for the treatment of fungal infections, and their therapeutic potential can be expanded through multicenter research and interdisciplinary collaboration. In addition to scientific and technological advancements, a regulatory framework is necessary to support the safe, effective, and sustainable use of EOs in clinical settings.
Funding
This research was funded by Wrocław Medical University, Poland (grant number SUBZ.D110.24.085).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Sabino, R.; Veríssimo, C.; Pereira, Á.A.; Antunes, F. Candida auris, an agent of hospital-associated outbreaks: Which challenging issues do we need to have in mind? Microorganisms 2020, 8, 181. [Google Scholar] [CrossRef]
- CDC’s Antibiotic Resistance Threats in the United States. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 8 May 2024).
- Rhodes, J.; Fisher, M.C. Global epidemiology of emerging Candida auris. Curr. Opin. Microbiol. 2019, 52, 84–89. [Google Scholar] [CrossRef]
- Spivak, E.S.; Hanson, K.E. Candida auris: An emerging fungal pathogen. J. Clin. Microbiol. 2018, 56, e01588-17. [Google Scholar] [CrossRef]
- Day, A.M.; McNiff, M.M.; da Silva Dantas, A.; Gow, N.A.R.; Quinna, J. Hog1 Regulates stress tolerance and virulence in the emerging fungal pathogen Candida auris. MBP 2018, 3, e00506-18. [Google Scholar]
- Forsberg, K.; Woodworth, K.; Walters, M.; Berkow, E.L.; Jackson, B.; Chiller, T.; Vallabhaneni, S. Candida auris: The recent emergence of a multidrug-resistant fungal pathogen. Med. Mycol. 2019, 57, 1–12. [Google Scholar] [CrossRef]
- Biswal, M.; Rudramurthy, S.M.; Jain, N.; Shamanth, A.S.; Sharma, D.; Jain, K.; Yaddanapudi, L.N.; Chakrabarti, A. Controlling a possible outbreak of Candida auris infection: Lessons learnt from multiple interventions. J. Hosp. Infect. 2017, 97, 363–370. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef]
- Johnson, J.C.; Davis, M.; Huttenlocher, A.; Kernien, A.F.; Nett, J.E. Emerging fungal pathogen Candida auris evades neutrophil attack. mBIO 2018, 9, e01403-18. [Google Scholar]
- Navarro-Arias, M.J.; Hernández-Chávez, M.J.; García-Carnero, L.C.; Amezcua-Hernández, D.G.; Lozoya-Pérez, N.E.; Estrada-Mata, E.; Martínez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef]
- Dominguez, E.G.; Zarnowski, R.; Choy, H.L.; Zhao, M.; Sanchez, H.; Nett, J.E.; Andes, D.R. Conserved role for biofilm matrix polysaccharides in Candida auris drug resistance. mSphere 2019, 4, 2–8. [Google Scholar] [CrossRef]
- Romera, D.; Aguilera-Correa, J.J.; Gadea, I.; Viñuela-Sandoval, L.; García-Rodríguez, J.; Esteban, J. Candida auris: A comparison between planktonic and biofilm susceptibility to antifungal drugs. J. Med. Microbiol. 2019, 68, 1353–1358. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009-17) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef]
- Kean, R.; McKloud, E.; Townsend, E.M.; Sherry, L.; Delaney, C.; Jones, B.L.; Williams, C.; Ramage, G. The comparative efficacy of antiseptics against Candida auris biofilms. Int. J. Antimicrob. Agents 2018, 52, 673–677. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef]
- Córdoba, S.; Vivot, W.; Szusz, W.; Albo, G. Antifungal activity of essential oils against Candida species isolated from clinical samples. Mycopathologia 2019, 184, 615–623. [Google Scholar] [CrossRef]
- Rashed, A.A.; Rathi, D.N.G.; Nasir, N.A.H.A.; Rahman, A.Z.A. Antifungal properties of essential oils and their compounds for application in skin fungal infections: Conventional and nonconventional approaches. Molecules 2021, 26, 1093. [Google Scholar] [CrossRef]
- Konuk, H.B.; Ergüden, B. Phenolic –OH group is crucial for the antifungal activity of terpenoids via disruption of cell membrane integrity. Folia Microbiol. 2020, 65, 775–783. [Google Scholar] [CrossRef]
- Hertiani, T.; Burhan, F.; Bafadal, M.; Pratiwi, U.S.T. Membrane cell disruption of Candida albicans by Masoyi bark essential oil. IJRPS 2020, 11, 2598–2602. [Google Scholar] [CrossRef]
- Shahina, Z.; Ndlovu, E.; Persaud, O.; Sultana, T. Candida albicans reactive oxygen species (ROS)-dependent. Microbiol. Spectr. 2022, 10, e03183-22. [Google Scholar] [CrossRef]
- Mani-López, E.; Cortés-Zavaleta, O.; López-Malo, A. A Review of the methods used to determine the target site or the mechanism of action of essential oils and their components against fungi. SN Appl. Sci. 2021, 3, 44. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Q.; Wu, H.; Huang, J.; Ju, J. Synergistic antifungal mechanism of effective components from essential oil against Penicillium roqueforti. Eng. Microbiol. 2023, 3, 100057. [Google Scholar] [CrossRef]
- Saracino, I.M.; Foschi, C.; Pavoni, M.; Spigarelli, R.; Valerii, M.C.; Spisni, E. Antifungal activity of natural compounds vs. Candida spp.: A mixture of cinnamaldehyde and eugenol shows promising in vitro results. Antibiotics 2022, 11, 73. [Google Scholar]
- Andrade-Ochoa, S.; Chacón-Vargas, K.F.; Sánchez-Torres, L.E.; Rivera-Chavira, B.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Differential antimicrobial effect of essential oils and their main components: Insights based on the cell membrane and external structure. Membranes 2021, 11, 405. [Google Scholar] [CrossRef]
- Mandras, N.; Roana, J.; Scalas, D.; Del Re, S.; Cavallo, L.; Ghisetti, V.; Tullio, V. The inhibition of non-albicans Candida species and uncommon yeast pathogens by selected essential oils and their major compounds. Molecules 2021, 26, 4937. [Google Scholar] [CrossRef]
- El-Baz, A.M.; Mosbah, R.A.; Goda, R.M.; Mansour, B.; Sultana, T.; Dahms, T.E.S.; El-Ganiny, A.M. Back to nature: Combating Candida albicans biofilm, phospholipase and hemolysin using plant essential oils. Antibiotics 2021, 10, 81. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Ożarowski, M.; Seremak-Mrozikiewicz, A.; Wolski, H. Anti-Candida and antibiofilm activity of selected lamiaceae essential oils. Front. Biosci.-Landmark 2023, 28, 28. [Google Scholar] [CrossRef]
- McGregor, R.C.; Parker, K.A.; Hornby, J.M.; Latta, L.C. Microbial population dynamics under microdoses of the essential oil arborvitae. BMC Complement. Altern. Med. 2019, 19, 247. [Google Scholar] [CrossRef]
- Tran, H.N.H.; Graham, L.; Adukwu, E.C. In vitro antifungal activity of cinnamomum zeylanicum bark and leaf essential oils against Candida albicans and Candida auris. Appl. Microbiol. Biotechnol. 2020, 104, 8911–8924. [Google Scholar] [CrossRef]
- Hamdy, R.; Fayed, B.; Hamoda, A.M.; Rawas-Qalaji, M.; Haider, M.; Soliman, S.S.M. Essential oil-based design and development of novel anti- Candida azoles formulation. Molecules 2020, 25, 1463. [Google Scholar] [CrossRef]
- De Alteriis, E.; Maione, A.; Falanga, A.; Bellavita, R.; Galdiero, S.; Albarano, L.; Salvatore, M.M.; Galdiero, E.; Guida, M. Activity of free and liposome-encapsulated essential oil from Lavandula angustifolia against persister-derived biofilm of Candida auris. Antibiotics 2022, 11, 26. [Google Scholar] [CrossRef]
- Parker, R.A.; Gabriel, K.T.; Graham, K.D.; Butts, B.K.; Cornelison, C.T. Antifungal activity of select essential oils against Candida auris and their interactions with antifungal drugs. Pathogens 2022, 11, 821. [Google Scholar] [CrossRef]
- Fernandes, L.; Ribeiro, R.; Costa, R.; Henriques, M.; Rodrigues, M.E. Essential oils as a good weapon against drug-resistant Candida auris. Antibiotics 2022, 11, 977. [Google Scholar] [CrossRef]
- Spengler, G.; Gajdács, M.; Donadu, M.G.; Usai, M.; Marchetti, M.; Ferrari, M.; Mazzarello, V.; Zanetti, S.; Nagy, F.; Kovács, R. Evaluation of the antimicrobial and antivirulent potential of essential oils isolated from Juniperus oxycedrus L. ssp. macrocarpa aerial parts. Microorganisms 2022, 10, 758. [Google Scholar] [CrossRef]
- Ribeiro, R.; Fernandes, L.; Costa, R.; Cavaleiro, C.; Salgueiro, L.; Henriques, M.; Rodrigues, M.E. Comparing the effect of Thymus spp. essential oils on Candida auris. Ind. Crops Prod. 2022, 178, 114667. [Google Scholar] [CrossRef]
- Baldim, I.; Paziani, M.H.; Grizante Barião, P.H.; von Zeska Kress, M.R.; Oliveira, W.P. Nanostructured lipid carriers loaded with lippia sidoides essential oil as a strategy to combat the multidrug-resistant Candida auris. Pharmaceutics 2022, 14, 180. [Google Scholar] [CrossRef]
- Di Vito, M.; Garzoli, S.; Rosato, R.; Mariotti, M.; Gervasoni, J.; Santucci, L.; Ovidi, E.; Cacaci, M.; Lombarini, G.; Torelli, R.; et al. A new potential resource in the fight against Candida auris: The Cinnamomum zeylanicum essential oil in synergy with antifungal drug. Microbiol. Spectr. 2023, 11, e0438522. [Google Scholar] [CrossRef]
- Rosato, R.; Napoli, E.; Granata, G.; Di Vito, M.; Garzoli, S.; Geraci, C.; Rizzo, S.; Torelli, R.; Sanguinetti, M.; Bugli, F. Study of the chemical profile and anti-fungal activity against Candida auris of Cinnamomum aassia essential oil and of its nano-formulations based on polycaprolactone. Plants 2023, 12, 358. [Google Scholar] [CrossRef]
- Ruiz-Duran, J.; Torres, R.; Stashenko, E.E.; Ortiz, C. Antifungal and antibiofilm activity of colombian essential oils against different Candida strains. Antibiotics 2023, 12, 668. [Google Scholar] [CrossRef]
- Zapata-Zapata, C.; Loaiza-Oliva, M.; Martínez-Pabón, M.C.; Stashenko, E.E.; Mesa-Arango, A.C. In vitro activity of essential oils distilled from colombian plants against Candida auris and other Candida species with different antifungal susceptibility profiles. Molecules 2022, 27, 6837. [Google Scholar] [CrossRef]
- Shaban, S.; Patel, M.; Ahmad, A. Improved efficacy of antifungal drugs in combination with monoterpene phenols against Candida auris. Sci. Rep. 2020, 10, 1162. [Google Scholar] [CrossRef]
- Horn, C.; Vediyappan, G. Anticapsular and antifungal activity of α-cyperone. Antibiotics 2021, 10, 51. [Google Scholar] [CrossRef]
- Maione, A.; La Pietra, A.; de Alteriis, E.; Mileo, A.; De Falco, M.; Guida, M.; Galdiero, E. Effect of myrtenol and its synergistic interactions with antimicrobial drugs in the inhibition of single and mixed biofilms of Candida auris and Klebsiella pneumoniae. Microorganisms 2022, 10, 1773. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).