

Genetic Characterization of Antibiotic-Resistant Staphylococcus spp. and Mammaliicoccus sciuri from Healthy Humans and Poultry in Nigeria
 , and
, and
Abstract
:1. Introduction
2. Results
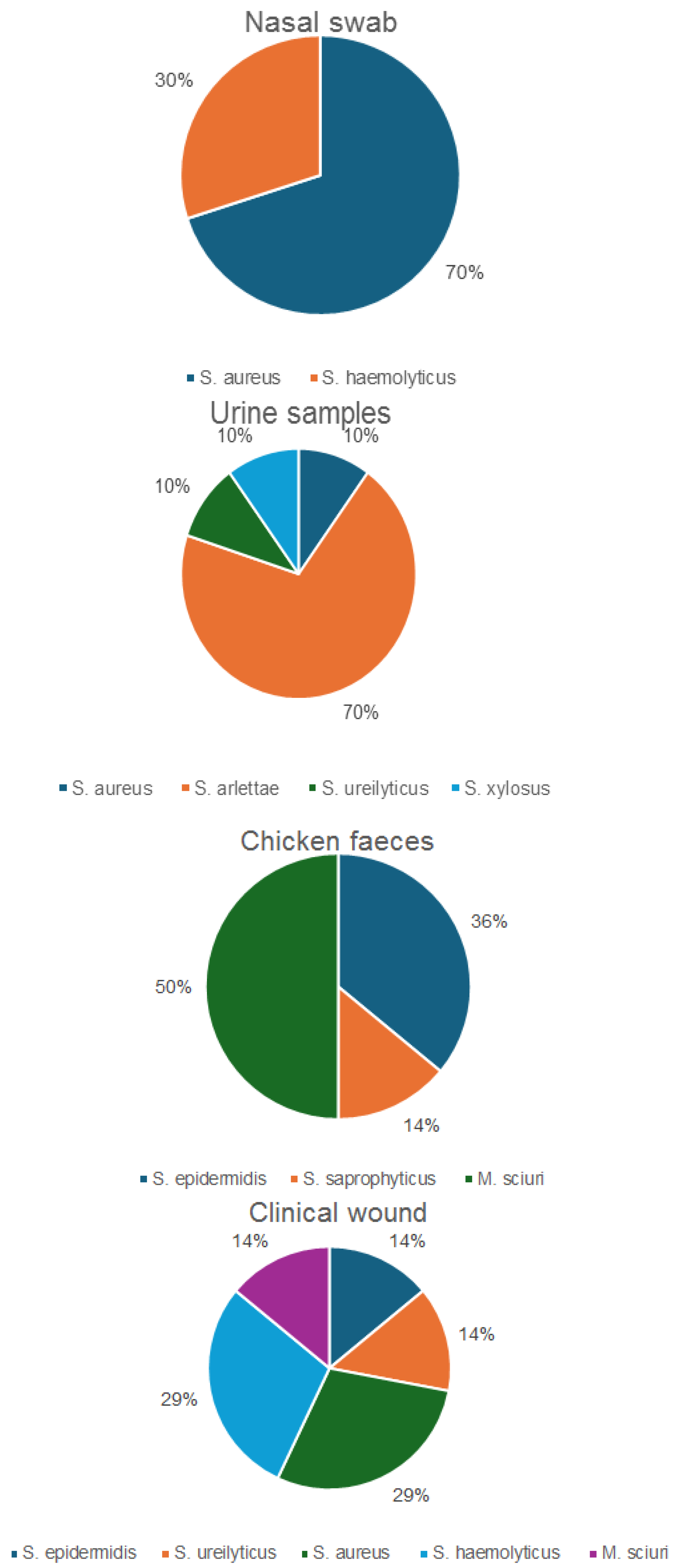
2.1. Identification of Staphylococcus spp. Including S. (Mammaliicoccus) sciuri
2.2. Antimicrobial Susceptibility
2.3. Antimicrobial Resistance Genes and SCCmec Element
2.4. Multilocus Sequence Typing of the Staphylococcus spp. Isolates
2.5. Virulence Genes Extracted from S. aureus Genomes
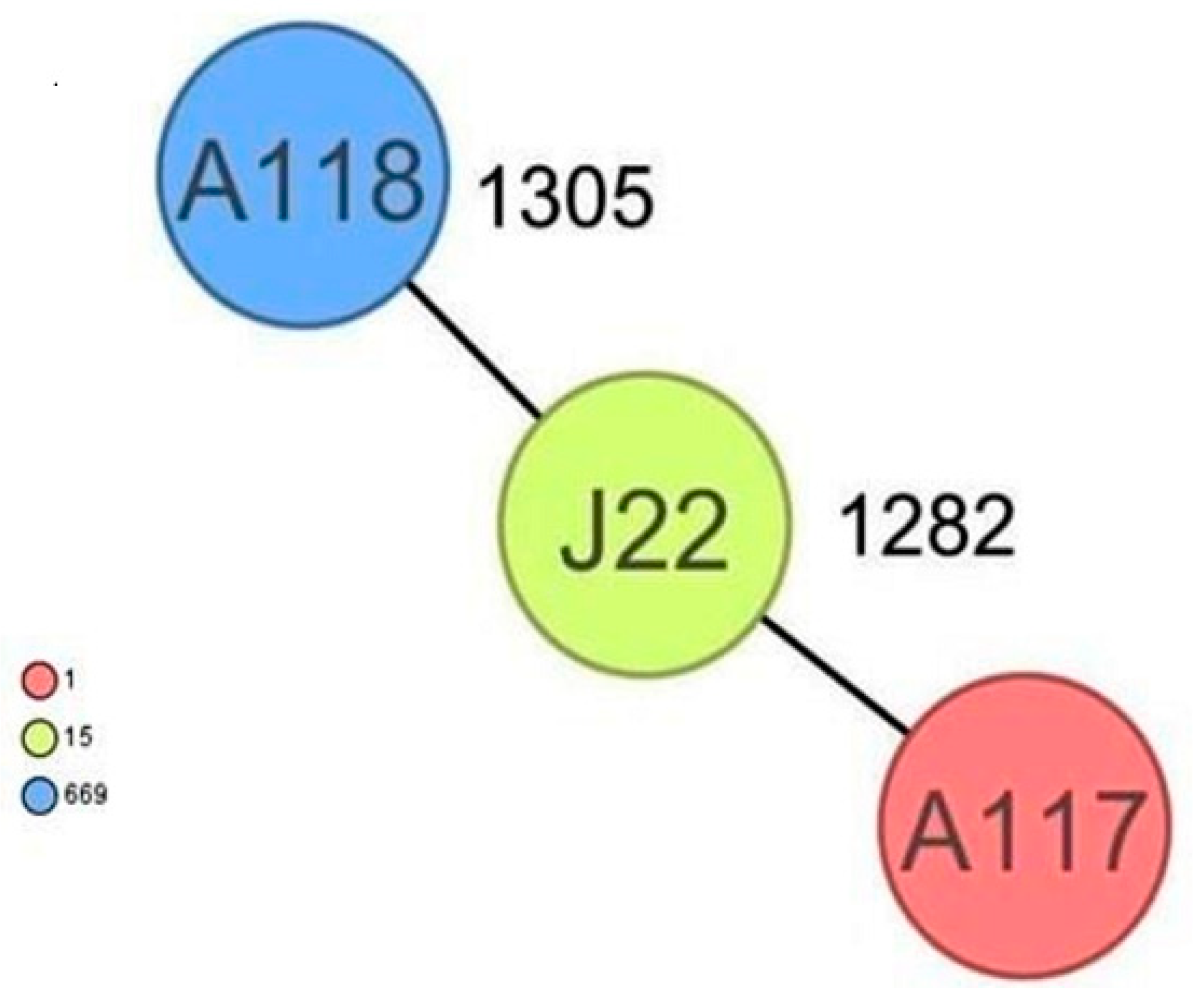
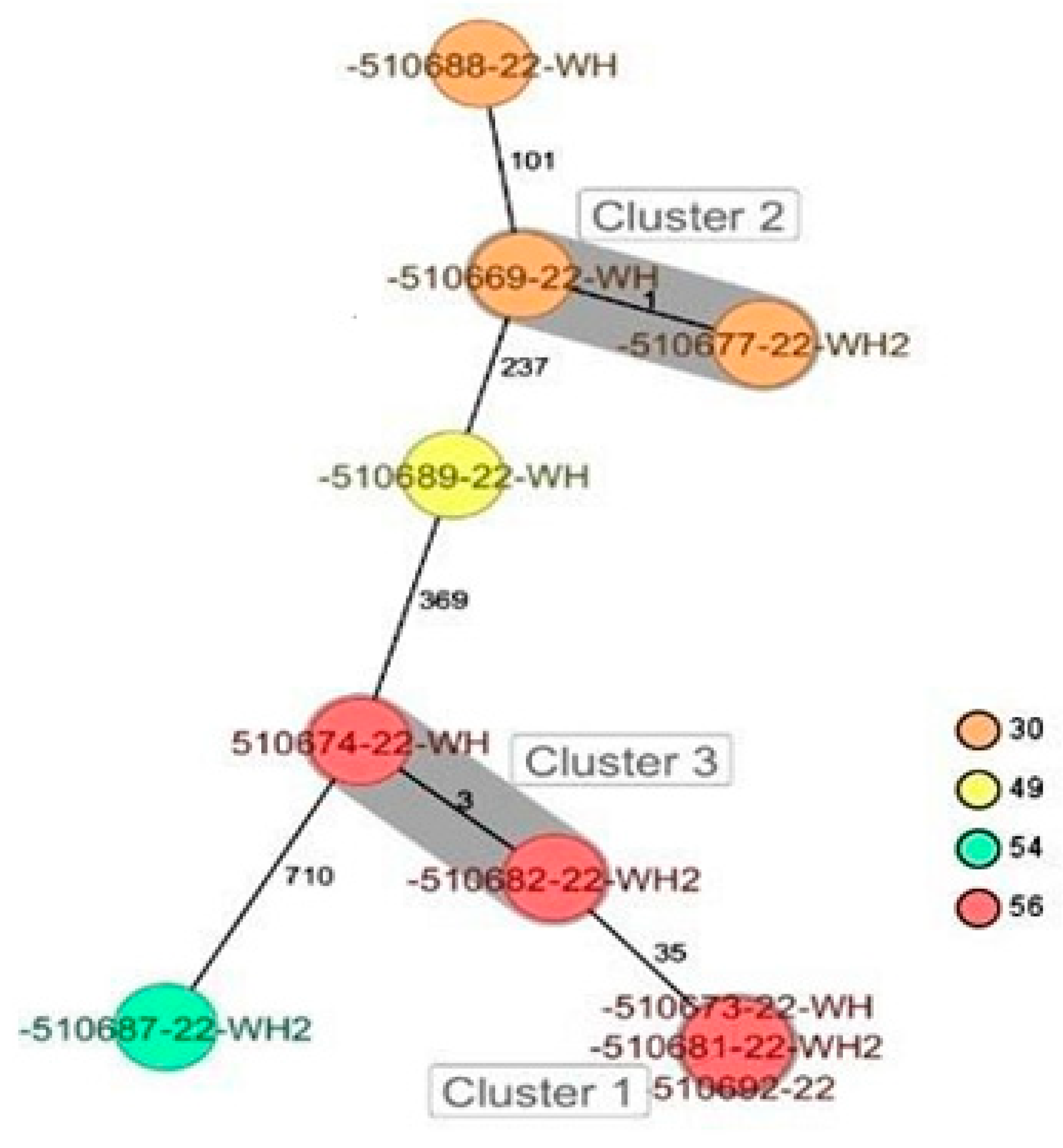
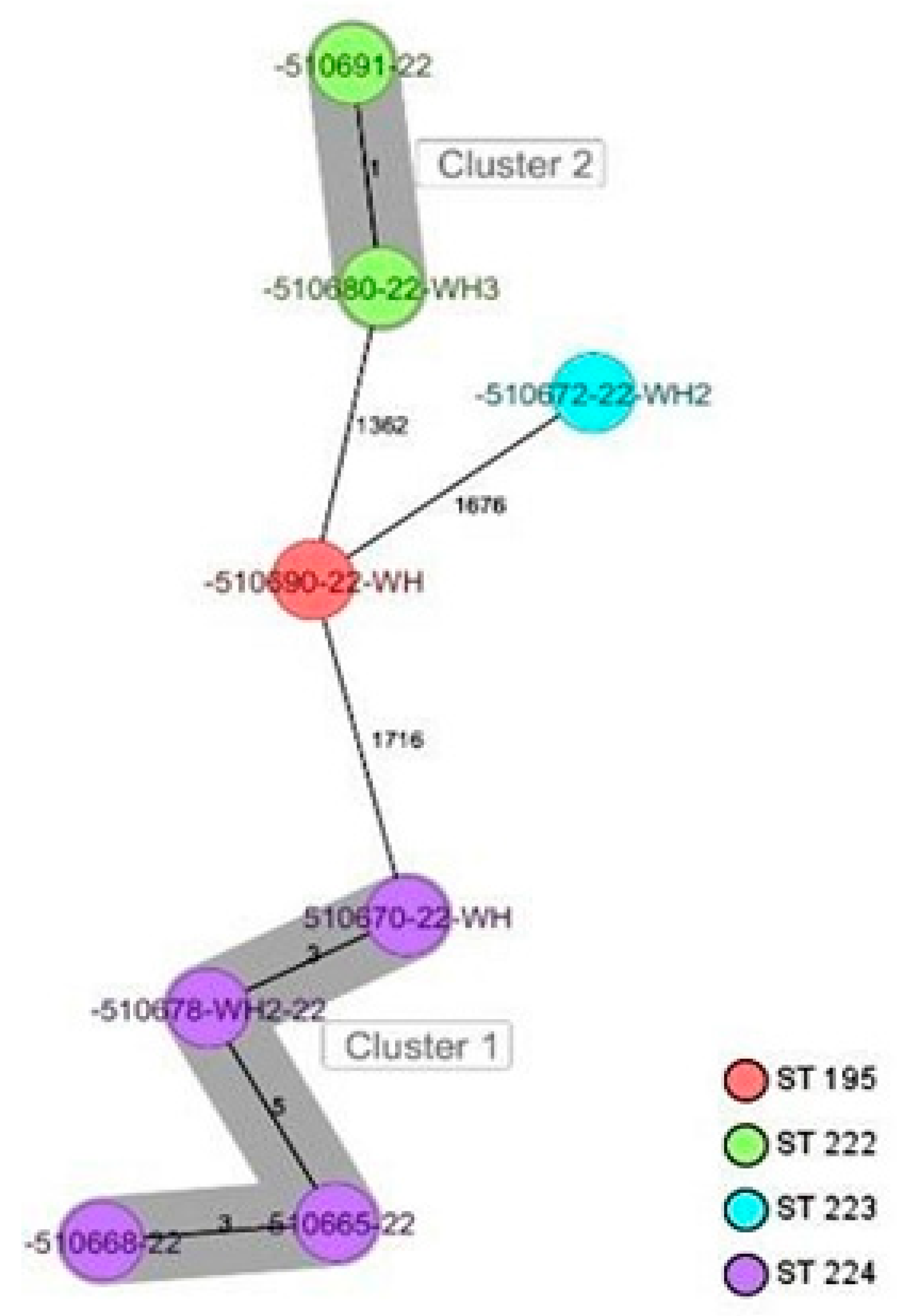
2.6. Genetic Comparison of the Staphylococcus spp. Isolates, Including M. sciuri Isolates
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. Antibiotic Susceptibility Testing
4.3. Whole-Genome Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pottumarthy, S.; Schapiro, J.M.; Prentice, J.L.; Houze, Y.B.; Swanzy, S.R.; Fang, F.C.; Cookson, B.T. Clinical isolates of Staphylococcus intermedius masquerading as methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2004, 42, 5881–5884. [Google Scholar] [CrossRef] [PubMed]
- Van Hoovels, L.; Vankeerberghen, A.; Boel, A.; Van Vaerenbergh, K.; De Beenhouwer, H. First case of Staphylococcus pseudintermedius infection in a human. J. Clin. Microbiol. 2006, 44, 4609–4612. [Google Scholar] [CrossRef] [PubMed]
- Achi, O.; Madubuike, C. Prevalence and antimicrobial resistance of Staphylococcus aureus isolated from retail ready to eat foods in Nigeria. J. Res. Microbiol. 2007, 2, 516–523. [Google Scholar]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- Chajecka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Łaniewska-Trokenheim, L. Coagulase-negative staphylococci (CoNS) isolated from ready-to-eat food of animal origin—Phenotypic and genotypic antibiotic resistance. Food Microbiol. 2015, 46, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Agersal, Y.; Ahrens, P.; Jørgensen, J.C.Ø.; Madsen, M.; Jensen, L.B. Antimicrobial susceptibility and presence of resistance genes in staphylococci from poultry. Vet. Microbiol. 2000, 74, 353–364. [Google Scholar] [CrossRef]
- Hashempour-Baltork, F.; Hosseini, H.; Shojaee-Aliabadi, S.; Torbati, M.; Alizadeh, A.M.; Alizadeh, M. Drug resistance and the prevention strategies in food borne Bacteria: An update review. Adv. Pharm. Bull. 2019, 9, 335–347. [Google Scholar] [CrossRef]
- Smith, R.; Coast, J. The true cost of antimicrobial resistance. BMJ 2013, 346, f1493. [Google Scholar] [CrossRef]
- Adenipekun, E.O.; Jackson, C.R.; Oluwadun, A.; Iwalokun, B.A.; Frye, J.G.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A. Prevalence and Antimicrobial Resistance in Escherichia coli from Food Animals in Lagos, Nigeria. Microb. Drug Resist. 2015, 21, 358–365. [Google Scholar] [CrossRef]
- Otalu, O.; Junaidu, K.; Chukwudi, O.E.; Jarlath, U.V. Multi-drug resistant coagulase positive Staphylococcus aureus from live and slaughtered chickens in Zaria, Nigeria. Int. J. Poult. Sci. 2011, 10, 871–875. [Google Scholar] [CrossRef]
- McCallum, N.; Berger-Bachi, B.; Senn, M.M. Regulation ¨of antibiotic resistance in Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 118–129. [Google Scholar] [CrossRef]
- Tiemersma, E.W.; Bronzwaer, S.L.; Lyytikäinen, O.; Degener, J.E.; Schrijnemakers, P.; Bruinsma, N.; Monen, J.; Witte, W.; Grundmann, H. Methicillin-resistant Staphylococcus aureus in Europe, 1999–2002. Emerg. Infect. Dis. 2004, 10, 1627. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard defnitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Shittu, A.O.; Taiwo, F.F.; Frobose, N.J.; Schwartbeck, B.; Niemann, S.; Mellmann, A.; Schaumburg, F. Genomic analysis of Staphylococcus aureus from the West African Dwarf (WAD) goat in Nigeria. Antimicrob. Resist. Infect. Control 2021, 10, 122. [Google Scholar] [CrossRef]
- Egyir, B.; Dsani, E.; Owusu-Nyantakyi, C.; Amuasi, G.R.; Owusu, F.A.; Allegye-Cudjoe, E.; Addo, K.K. Antimicrobial resistance and genomic analysis of staphylococci isolated from livestock and farm attendants in Northern Ghana. BMC Microbiol. 2022, 22, 180. [Google Scholar] [CrossRef]
- Pimenta, L.K.L.; Rodrigues, C.A.; Filho, A.R.G.; Coelho, C.J.; Goes, V.; Estrela, M.; Priscila de Souza Avelino, M.A.G.; Vieira, J.D.G.; Carneiro, L. Staphylococcus spp. Causatives of Infections and Carrier of blaZ, femA, and mecA Genes Associated with Resistance. Antibiotics 2023, 12, 671. [Google Scholar] [CrossRef]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host adaptation and emergence of methicillin resistance in livestock. mBio 2012, 3, e00305–e00311. [Google Scholar] [CrossRef] [PubMed]
- Richardson, E.J.; Bacigalupe, R.; Harrison, E.M.; Weinert, L.A.; Lycett, S.; Vrieling, M.; Robb, K.; Hoskisson, P.A.; Holden, M.T.G.; Feil, E.J.; et al. Gene exchange drives the ecological success of a multihost bacterial pathogen. Nat. Ecol. Evol. 2018, 2, 1468–1478. [Google Scholar] [CrossRef]
- Ibadin, E.E.; Enabulele, I.O.; Muinah, F. Prevalence of mecA gene among staphylococci from clinical samples of a tertiary hospital in Benin City, Nigeria. Afr. Health Sci. 2017, 17, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Karikari, A.B.; Frimpong, E.; Owusu-Ofori, A. Methicillin-resistant Staphylococcus aureus among patients in a teaching hospital in Ghana. Int. J. One Health 2017, 3, 46–49. [Google Scholar] [CrossRef]
- Sangeeta, R.; Lyndsey, L.; Roberta, M.; Linzy, J.; Doreene, R.H. Antimicrobial resistance and genetic diversity of Staphylococcus aureus collected from Livestock, poultry and Humans. One Health 2022, 15, 100407. [Google Scholar]
- Igbinosa, E.O.; Beshiru, A.; Igbinosa, I.H.; Ogofure, A.G.; Ekundayo, T.C.; Okoh, A.I. Prevalence, multiple antibiotic resistance and virulence profile of methicillin-resistant Staphylococcus aureus (MRSA) in retail poultry meat from Edo, Nigeria. Front. Cell Infect. Microbiol. 2023, 13, 1122059. [Google Scholar] [CrossRef]
- Washington, M.A.; Agee, W.A., III; Kajiura, L.; Staege, C.M.; Uyehara, C.F.; Barnhill, J.C. An Analysis of Staphylococcus aureus Infections at a Military Medical Center Using the PLEX-ID Combined Polymerase Chain Reaction-Mass Spectrometry System. Maj. Mil. Med. 2014, 179, 445. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.R.; Batistao, D.W.F.; Ribas, R.M.; Sousa, A.M.; Pereira, M.O.; Botelho, C.M. Staphylococcus aureus virulence factors and disease. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 702–710. [Google Scholar]
- Rahimi, F.; Katouli, M.; Karimi, S. Bioflm production among methicillin resistant Staphylococcus aureus strains isolated from catheterized patients with urinary tract infection. Microb. Pathog. 2016, 98, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Motallebi, M.; Jabalameli, F.; Asadollahi, K.; Taherikalani, M.; Emaneini, M. Spreading of genes encoding enterotoxins, haemolysins, adhesin and bioflm among methicillin resistant Staphylococcus aureus strains with staphylococcal cassette chromosome mec type IIIA isolated from burn patients. Microb. Pathog. 2016, 97, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Lappin-Scott, H.M.; Bass, C. Biofilm formation: Attachment, growth and detachment of microbes from surfaces. Am. J. Infect. Control 2001, 29, 250–251. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef]
- Parom, S.K.; Igwe, J.C.; Bolaji, R.O.; Adeshina, G.O.; Onaolapo, J.A. Molecular characterization of methicillin resistance gene among Staphylococcus aureus isolated from poultry farms in Kaduna, Nigeria. J. Biosci. Biotechnol. Discov. 2018, 3, 5–13. [Google Scholar] [CrossRef]
- Ogundipe, F.O.; Ojo, O.E.; Feßler, A.T.; Hanke, D.; Awoyomi, O.J.; Ojo, D.A.; Akintokun, A.K.; Schwarz, S.; Maurischat, S. Antimicrobial resistance and virulence of methicillin-resistant Staphylococcus aureus from human, chicken and environmental samples within live bird markets in three Nigerian cities. Antibiotics 2020, 9, 588. [Google Scholar] [CrossRef] [PubMed]
- Kantiyok, T.J.; Arowolo, O.A.R.; Asanga, S.A.; Gberindyer, F.A. Antibacterial Use and Level of Resistance in Poultry: A Retrospective Study in North-Central and South-West Nigeria. J. Poult. Res. 2018, 15, 13–17. [Google Scholar]
- Jesumirhewe, C.; Springer, B.; Allerberger, F.; Ruppitsch, W. Whole genome sequencing of extended-spectrum beta-lactamase genes in Enterobacteriaceae isolates from Nigeria. PLoS ONE 2020, 15, e0231146. [Google Scholar] [CrossRef] [PubMed]
- Ndahi, M.D.; Hendrikse, R.; Helwigh, B.; Card, R.M.; Fagbamila, I.O.; Abiodun-Adewusi, O.O.; Ekeng, E.; Adetunji, V.; Adebiyi, I.; Andersen, J.K. Determination of antimicrobial use in commercial poultry farms in Plateau and Oyo States, Nigeria. Antimicrob. Resist. Infect. Control 2023, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Piessens, V.; van Coillie, E.; Verbist, B.; Supré, K.; Braem, G.; Van Nuffel, A.; De Vuyst, L.; Heyndrickx, M.; De Vliegher, S. Distribution of coagulase-negative Staphylococcus species from milk and environment of dairy cows differs between herds. J. Dairy Sci. 2011, 94, 2933–2944. [Google Scholar] [CrossRef] [PubMed]
- Nemeghaire, S.; Vanderhaeghen, W.; Angeles Argudín, M.; Haesebrouck, F.; Butaye, P. Characterization of methicillin-resistant Staphylococcus sciuri isolates from industrially raised pigs, cattle and broiler chickens. J. Antimicrob. Chemother. 2014, 69, 2928–2934. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Dakić, I.; Djukić, S.; Lozuk, B.; Svabić-Vlahović, M. Surgical wound infection associated with Staphylococcus sciuri. Scand. J. Infect. Dis. 2002, 34, 685–686. [Google Scholar] [CrossRef] [PubMed]
- Dakić, I.; Morrison, D.; Vuković, D.; Savić, B.; Shittu, A.; Ježek, P.; Hauschild, T.; Stepanović, S. Isolation and molecular characterization of Staphylococcus sciuri in the hospital environment. J. Clin. Microbiol. 2005, 43, 2782–2785. [Google Scholar] [CrossRef] [PubMed]
- Rolo, J.; Worning, P.; Boye, J.; Sobral, R.; Bowden, R.; Bouchami, O.; Damborg, P.; Guardabassi, L.; Perreten, V.; Westh, H.; et al. Evidence for the evolutionary steps leading to mecA-mediated β-lactam resistance in staphylococci. PLoS Genet. 2017, 13, e1006674. [Google Scholar] [CrossRef]
- Bortolaia, V.; Espinosa-Gongora, C.; Guardabassi, L. Human health risks associated with antimicrobial-resistant enterococci and Staphylococcus aureus on poultry meat. Clin. Microbiol. Infect. 2016, 22, 130–140. [Google Scholar] [CrossRef]
- Silva, V.; Caniça, M.; Ferreira, E.; Vieira-Pinto, M.; Saraiva, C.; Pereira, J.E.; Capelo, J.L.; Igrejas, G.; Poeta, P. Multidrug-Resistant Methicillin-Resistant Coagulase-Negative Staphylococci in Healthy Poultry Slaughtered for Human Consumption. Antibiotics 2022, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Mccarthy, A.J.; Lindsay, J.A. The distribution of plasmids that carry virulence and resistance genes in Staphylococcus aureus is lineage associated. BMC Microbiol. 2012, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, Y.; Schwarz, S.; Cai, J.; Fan, R.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J. Co-location of the oxazolidinone resistance genes optrA and cfr on a multiresistance plasmid from Staphylococcus sciuri. J. Antimicrob. Chemother. 2016, 71, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Soufi, L.; Campanile, F.; Dhaouadi, F.; Sociale, M.; Lazzaro, L. Prevalence of meticillin-resistant and -susceptible coagulase-negative staphylococci with the first detection of the mecC gene among cows, humans and manure in Tunisia. Int. J. Antimicrob. Agents 2020, 55, 105826. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Coagulase-negative staphylococci as reservoirs of genes facilitating MRSA infection: Staphylococcal commensal species such as Staphylococcus epidermidis are being recognized as important sources of genes promoting MRSA colonization and virulence. Bioessays 2013, 35, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Bathavatchalam, Y.D.; Solaimalai, D.; Amladi, A.; Dwarakanathan, H.T.; Anandan, S.; Veeraraghavan, B. Vancomycin heteroresistance in Staphylococcus haemolyticus: Elusive phenotype. Future Sci. OA 2021, 7, FSO710. [Google Scholar] [CrossRef]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef]
- Sarkhoo, E.; Udo, E.E.; Boswihi, S.S.; Monecke, S.; Mueller, E.; Ehricht, R. The Dissemination and Molecular Characterization of Clonal Complex 361 (CC361) Methicillin-Resistant Staphylococcus aureus (MRSA) in Kuwait Hospitals. Front. Microbiol. 2021, 12, 658772. [Google Scholar] [CrossRef]
- Ruppé, E.; Barbier, F.; Mesli, Y.; Maiga, A.; Cojocaru, R.; Benkhalfat, M.; Benchouk, S.; Hassaine, H.; Maiga, I.; Diallo, A.; et al. Diversity of staphylococcal cassette chromosome mec structures in methicillin-resistant Staphylococcus epidermidis and Staphylococcus haemolyticus strains among outpatients from four countries. Antimicrob. Agents Chemother. 2009, 53, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Bouchami, O.; Hassen, A.; Lencastre, H.; Miragaia, M. High prevalence of mec complex C and ccrC is independent of SCCmec type V in Staphylococcus haemolyticus. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 605–614. [Google Scholar] [CrossRef]
- Szczuka, E.; Krajewska, M.; Lijewska, D.; Bosacka, K.; Kaznowski, A. Diversity of staphylococcal cassette chromosome mec elements in nosocomial multiresistant Staphylococcus haemolyticus isolates. J. Appl. Genet. 2016, 57, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim-Saraie, H.S.; Motamedifar, M.; Sarvari, J.; Alfatemi, S.M.H. Emergence of SCCmec Type I Obtained from Clinical Samples in Shiraz Teaching Hospitals, South-West of Iran. Jundishapur. J. Microbiol. 2015, 8, e16998. [Google Scholar] [CrossRef] [PubMed]
- Cheesebrough, M. District Laboratory Practice in Tropical Countries, 2nd ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 178–187. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters; Version 12.0; European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2022. [Google Scholar]
- Jesumirhewe, C.; Springer, B.; Allerberger, F.; Ruppitsch, W. Genetic characterization of antibiotic resistant Enterobacteriaceae isolates from bovine animals and the environment in Nigeria. Front. Microbiol. 2022, 13, 793541. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Leopold, S.R.; Goering, R.V.; Witten, A.; Harmsen, D.; Mellmann, A. Bacterial whole-genome sequencing revisited: Portable, scalable, and standardized analysis for typing and detection of virulence and antibiotic resistance genes. J. Clin. Microbiol. 2014, 52, 2365–2370. [Google Scholar] [CrossRef] [PubMed]
- Ruppitsch, W.; Pietzka, A.; Prior, K.; Bletz, S.; Fernandez, H.; Allerberger, F.; Harmsen, D.; Mellmann, A. Defining and evaluating a core genome MLST scheme for whole genome sequence-based typing of Listeria monocytogenes. J. Clin. Microbiol. 2015, 53, 2869–2876. [Google Scholar] [CrossRef]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- Strauß, L.; Rufng, U.; Abdulla, S.; Alabi, A.; Akulenko, R.; Garrine, M.; Germann, A.; Grobusch, M.P.; Helms, V.; Herrmann, M.; et al. Detecting Staphylococcus aureus virulence and resistance genes: A comparison of whole-genome sequencing and DNA microarray technology. J. Clin. Microbiol. 2016, 54, 1008–1016. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017, expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- International Working Group on the Classifcation of Staphylococcal Cassette Chromosome. Classifcation of Staphylococcal Cassette Chromosome mec (SCCmec): Guidelines for Reporting Novel SCCmec Elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistant Isolates | VA 30 | TEC 30 | LZD 10 | FD 10 | RD 5 | W 5 | CN 10 | AK 30 | MXF 5 | MH 30 | E 15 | DA 2 | P 1 | FOX 30 | AMC 30 | CIP 5 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus (n = 3) | A | A | - | - | 33 (n = 1) | 100 (n = 3) | 100 (n = 3) | - | 33 (n = 1) | - | 33 (n = 1) | 100 (n = 3) | 100 (n = 3) | 33 (n = 1) | B | 33 (n = 1) |
| M. sciuri (n = 8) | A | A | - | 87.5 (n = 7) | 25 (n = 2) | 37.5 (n = 3) | 87.5 (n = 7) | - | 100 (n = 8) | 25 (n = 2) | - | 87.5 (n = 7) | 87.5 (n = 7) | 37.5 (n = 3) | B | 50 (n = 4) |
| S. haemolyticus (n = 10) | A | A | - | 20 (n = 2) | 20 (n = 2) | 90 (n = 9) | 80 (n = 8) | - | 90 (n = 9) | 10 (n = 1) | 40 (n = 4) | 30 (n = 3) | 100 (n = 10) | 80 (n = 8) | B | 90 (n = 9) |
| S. arlettae (n = 7) | A | A | - | 100 (n = 7) | 57 (n = 4) | 100 (n = 7) | - | - | - | - | 100 (n = 7) | 100 (n = 7) | 100 (n = 7) | - | B | - |
| S. saprophyticus (n = 1) | A | A | - | 100 (n = 1) | - | 100 (n = 1) | 100 (n = 1) | - | 100 (n = 1) | - | 100 (n = 1) | 100 (n = 1) | 100 (n = 1) | - | B | 100 (n = 1) |
| S. ureilyticus (n = 2) | A | A | - | 100 (n = 2) | 50 (n = 1) | 50 (n = 1) | - | - | - | - | 100 (n = 2) | 50 (n = 1) | 100 (n = 2) | - | B | - |
| S. xylosus (n = 1) | A | A | - | 100 (n = 1) | 100 (n = 1) | - | - | - | 100 (n = 1) | - | 100 (n = 1) | 100 (n = 1) | 100 (n = 1) | - | B | 100 (n = 1) |
| S. epidermidis (n = 2) | A | A | - | - | - | 100 (n = 2) | 100 (n = 2) | - | - | - | - | - | 100 (n = 2) | - | B | - |
| Isolate ID | Isolate | Resistance Profile |
|---|---|---|
| J6 | S. haemolyticus | W-CN-MXF-P-FOX-CIP |
| J9 | S. haemolyticus | W-CN-MXF-DA-P-FOX-CIP |
| J24 | S. haemolyticus | W-CN-MXF-P-FOX-CIP |
| J5 | S. haemolyticus | RD-W-MXF-E-DA-P-FOX-CIP |
| J31 | S. haemolyticus | W-CN-MXF-P-FOX-CIP |
| T77 | S. haemolyticus | FD-MXF-P |
| A128 | S. haemolyticus | FD-RD-W-CN-MXF-MH-E-DA-P-FOX-CIP |
| A119 | S. haemolyticus | W-CN-MXF-E-P-FOX-CIP |
| J8 | S. haemolyticus | W-CN-E-P-CIP |
| J12 | S. haemolyticus | W-CN-MXF-P-FOX-CIP |
| T66 | S. saprophyticus | FD-W-CN-MXF-E-DA-P-CIP |
| T76 | M. sciuri | FD-MXF-DA-P |
| J27 | M. sciuri | W-CN-MXF-P-FOX-CIP |
| T51 | M. sciuri | W-CN-MXF-DA-P |
| T66-2 | M. sciuri | FD-RD-W-CN-MXF-MH-DA-P-FOX-CIP |
| T40 | M. sciuri | FD-CN-MXF-DA |
| T50 | M. sciuri | FD-RD-W-CN-MXF-MH-DA-P-FOX-CIP |
| T37 | M. sciuri | FD-CN-MXF-DA-P |
| A125 | M. sciuri | FD-CN-MXF-DA-P-CIP |
| J22 | S. aureus | W-CN-P |
| A118 | S. aureus | RD-W-CN-MXF-E-P-FOX-CIP |
| A117 | S. aureus | W-CN-P |
| A146 | S. ureilyticus | FD-E-DA-P |
| H96 | S. ureilyticus | FD-RD-W-E-P |
| H97 | S. arlettae | FD-W-E-DA-P |
| H93 | S. arlettae | FD-RD-W-E-DA-P |
| H87-2 | S. arlettae | FD-W-E-DA-P |
| H99-2 | S. arlettae | FD-W-E-DA-P |
| H101 | S. arlettae | FD-RD-W-E-DA-P |
| H111 | S. arlettae | FD-RD-W-E-DA-P |
| H98-1 | S. arlettae | FD-RD-W-E-DA-P |
| H99-1 | S. xylosus | FD-RD-MXF-E-DA-P-CIP |
| A116 | S. epidermidis | W-CN-DA |
| J19-2 | S. epidermidis | W-CN-DA |
| Resistance Phenotypes | Number of Isolates |
|---|---|
| W-CN-MXF-P-FOX-CIP | 5 |
| FD-RD-W-E-DA-P | 4 |
| FD-W-E-DA-P | 3 |
| W-CN-P | 2 |
| F-CN-MXF-DA | 1 |
| W-CN-MXF-DA-P-FOX-CIP | 1 |
| FD-CN-MXF-DA-P | 1 |
| FD-CN-MXF-DA-P-CIP | 1 |
| RD-W-CN-MXF-E-P-FOX-CIP | 1 |
| FD-RD-W-CN-MXF-MH-DA-P-FOX-CIP | 2 |
| FD-RD-MXF-E-DA-P-CIP | 1 |
| W-CN-DA | 2 |
| FD-RD-W-E-P | 1 |
| FD-E-DA-P | 1 |
| W-CN-E-P-CIP | 1 |
| FD-W-CN-MXF-E-DA-P-CIP | 1 |
| FD-MXF-DA-P | 1 |
| W-CN-MXF-E-P-FOX-CIP | 1 |
| FD-RD-W-CN-MXF-MH-E-DA-P-FOX-CIP | 1 |
| FD-MXF-P | 1 |
| RD-W-MXF-E-DA-P-FOX-CIP | 1 |
| W-CN-MXF-DA-P | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesumirhewe, C.; Odufuye, T.O.; Ariri, J.U.; Adebiyi, A.A.; Sanusi, A.T.; Stöger, A.; Daza-Prieto, B.; Allerberger, F.; Cabal-Rosel, A.; Ruppitsch, W. Genetic Characterization of Antibiotic-Resistant Staphylococcus spp. and Mammaliicoccus sciuri from Healthy Humans and Poultry in Nigeria. Antibiotics 2024, 13, 733. https://doi.org/10.3390/antibiotics13080733
Jesumirhewe C, Odufuye TO, Ariri JU, Adebiyi AA, Sanusi AT, Stöger A, Daza-Prieto B, Allerberger F, Cabal-Rosel A, Ruppitsch W. Genetic Characterization of Antibiotic-Resistant Staphylococcus spp. and Mammaliicoccus sciuri from Healthy Humans and Poultry in Nigeria. Antibiotics. 2024; 13(8):733. https://doi.org/10.3390/antibiotics13080733
Chicago/Turabian StyleJesumirhewe, Christiana, Tolulope Oluwadamilola Odufuye, Juliana Ukinebo Ariri, Amdallat Arike Adebiyi, Amina Tanko Sanusi, Anna Stöger, Beatriz Daza-Prieto, Franz Allerberger, Adriana Cabal-Rosel, and Werner Ruppitsch. 2024. "Genetic Characterization of Antibiotic-Resistant Staphylococcus spp. and Mammaliicoccus sciuri from Healthy Humans and Poultry in Nigeria" Antibiotics 13, no. 8: 733. https://doi.org/10.3390/antibiotics13080733