Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation

 ,
, 

Abstract
:1. Introduction
2. Results
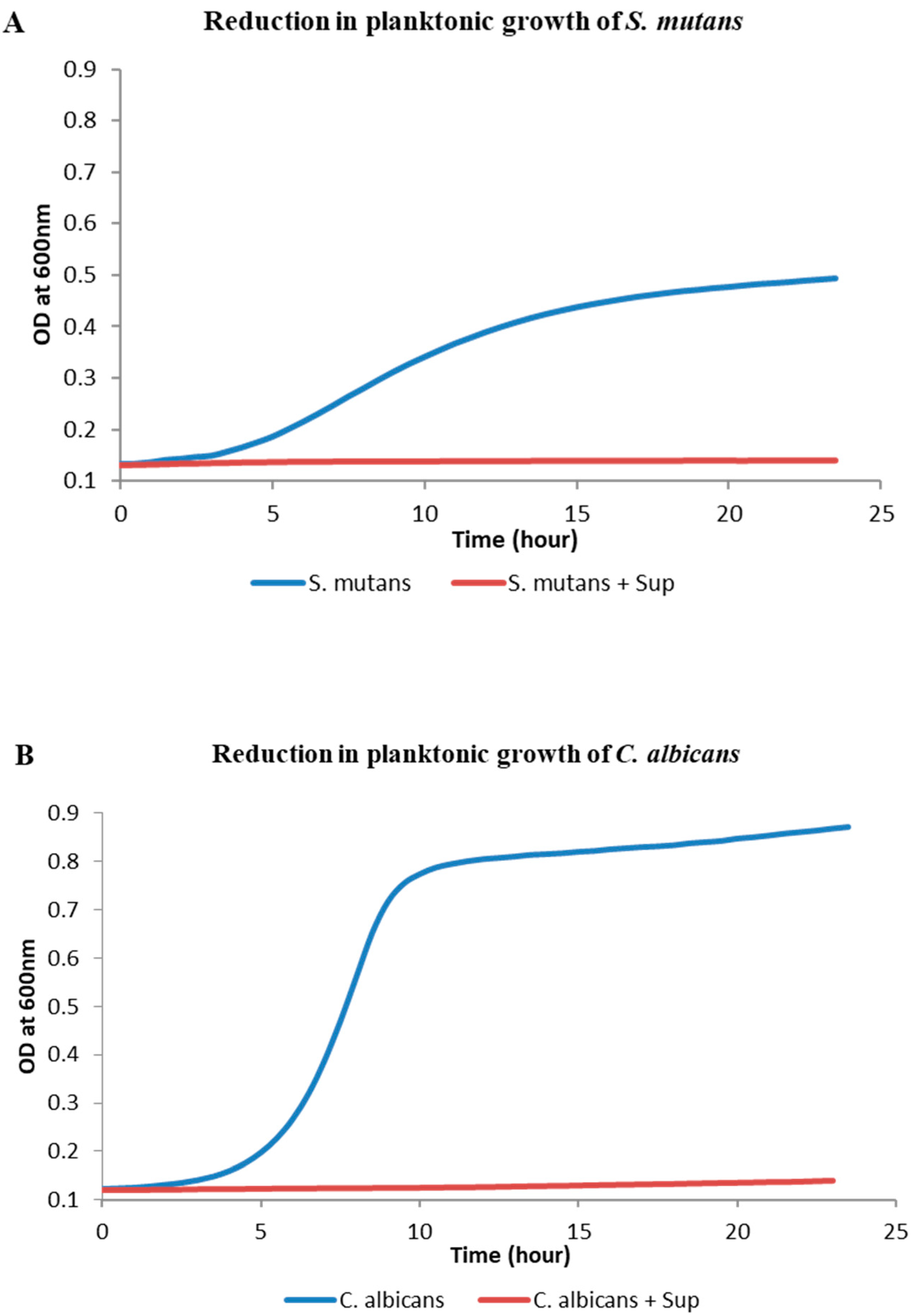
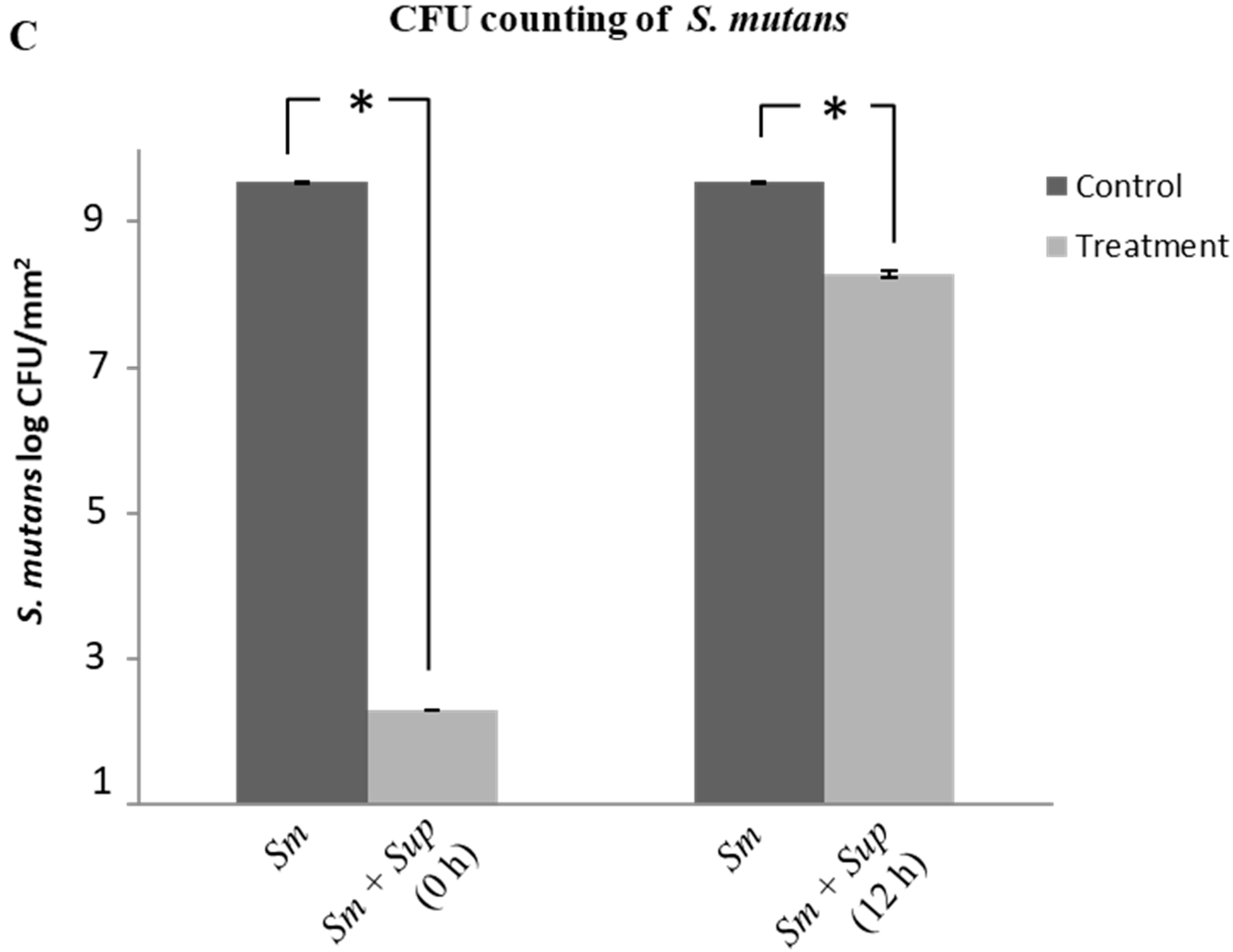
2.1. Effect of Lactobacillus plantarum 108 Supernatant on Planktonic Streptococcus mutans and Candida albicans
2.2. Effect of Lactobacillus Plantarum 108 Supernatant On Streptococcus Mutans Biofilm Formation
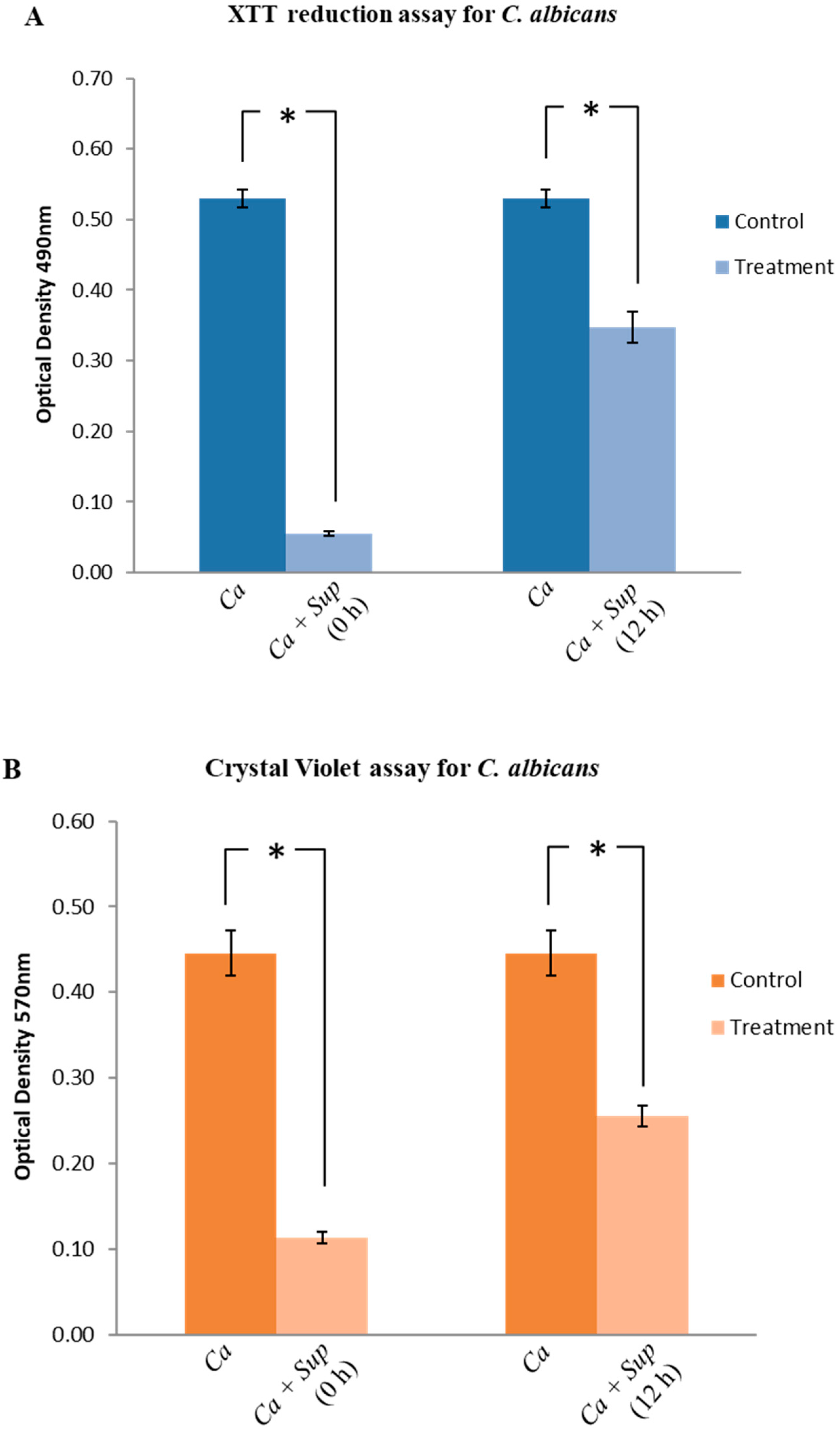
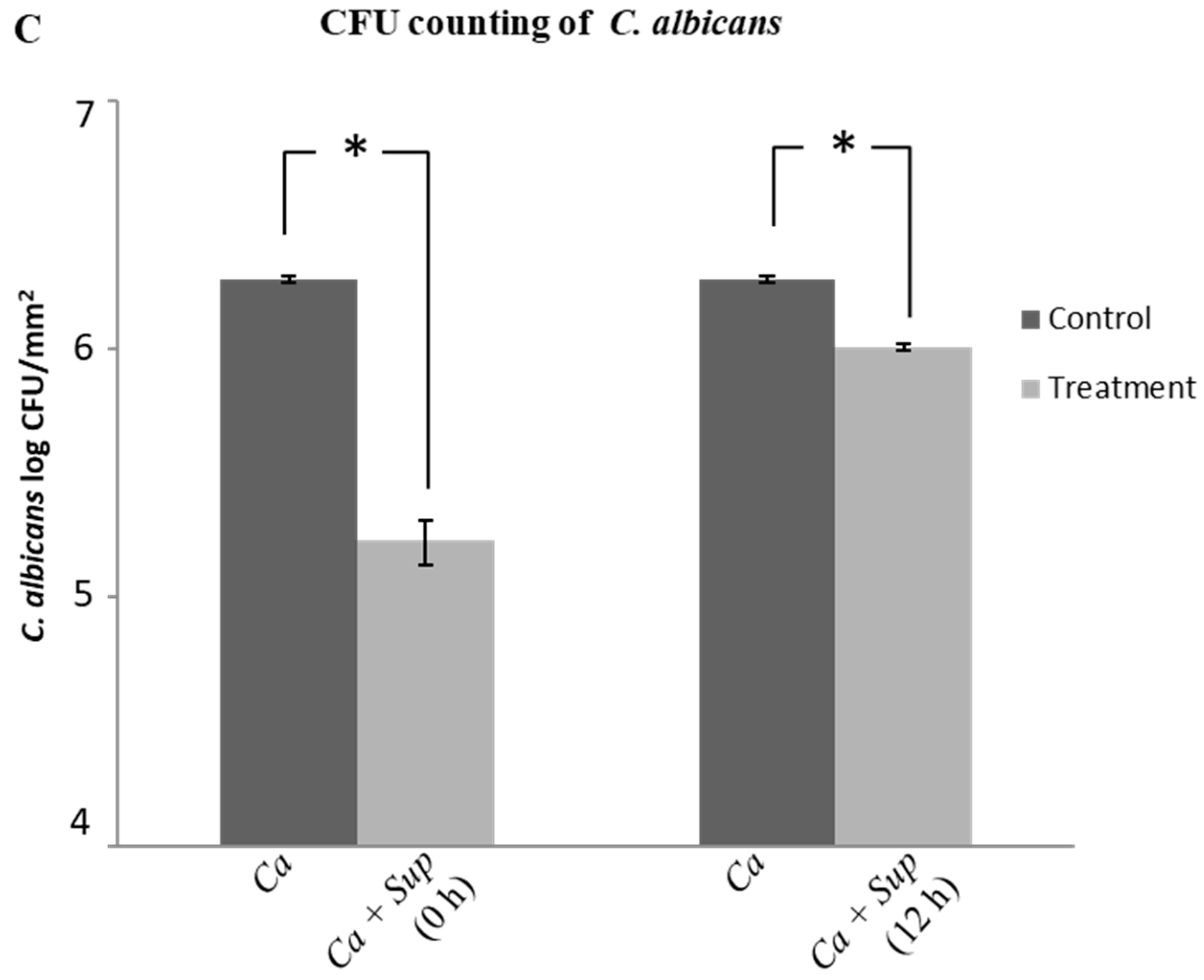
2.3. Effect of Lactobacillus Plantarum 108 Supernatant on Candida Albicans Biofilm Formation
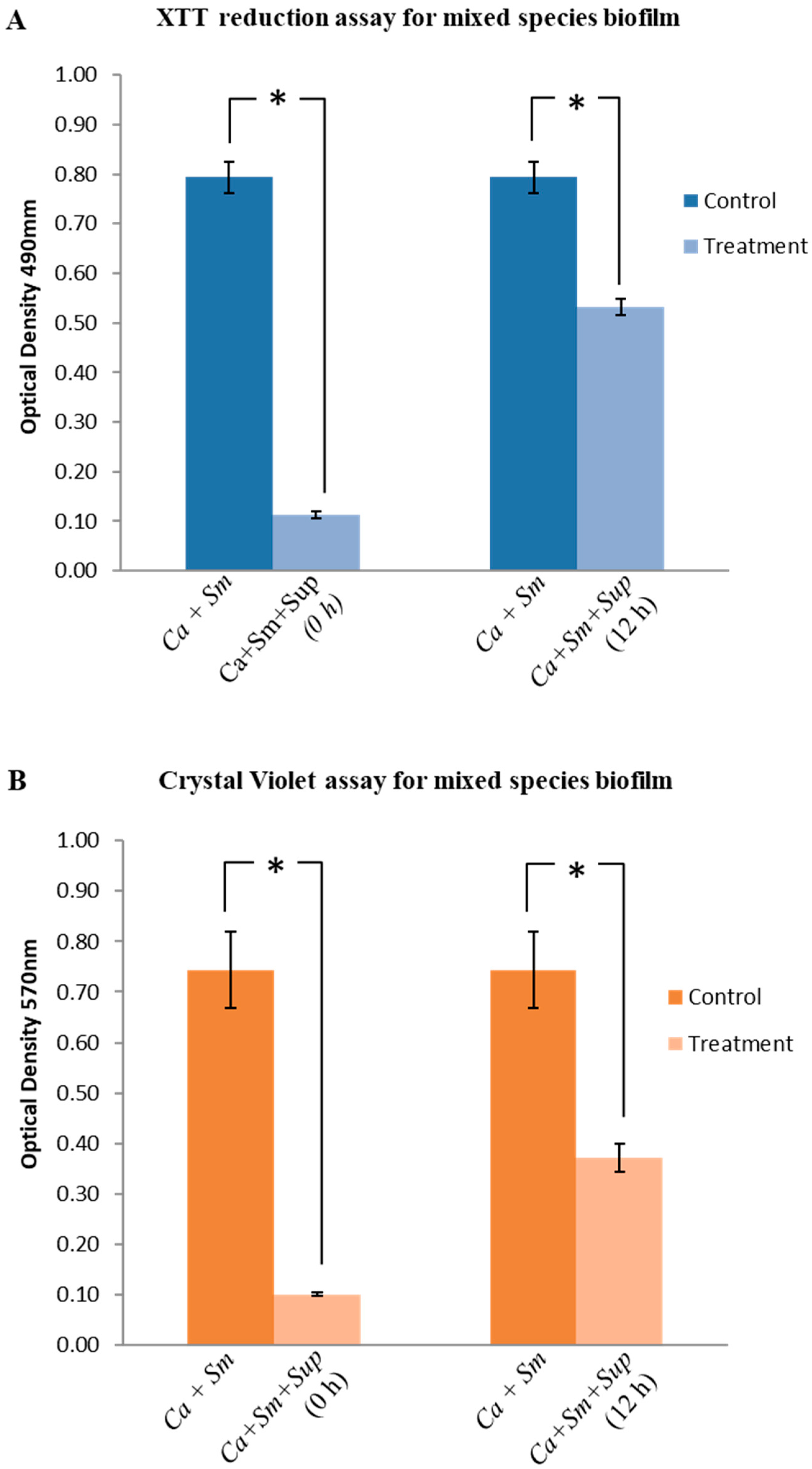
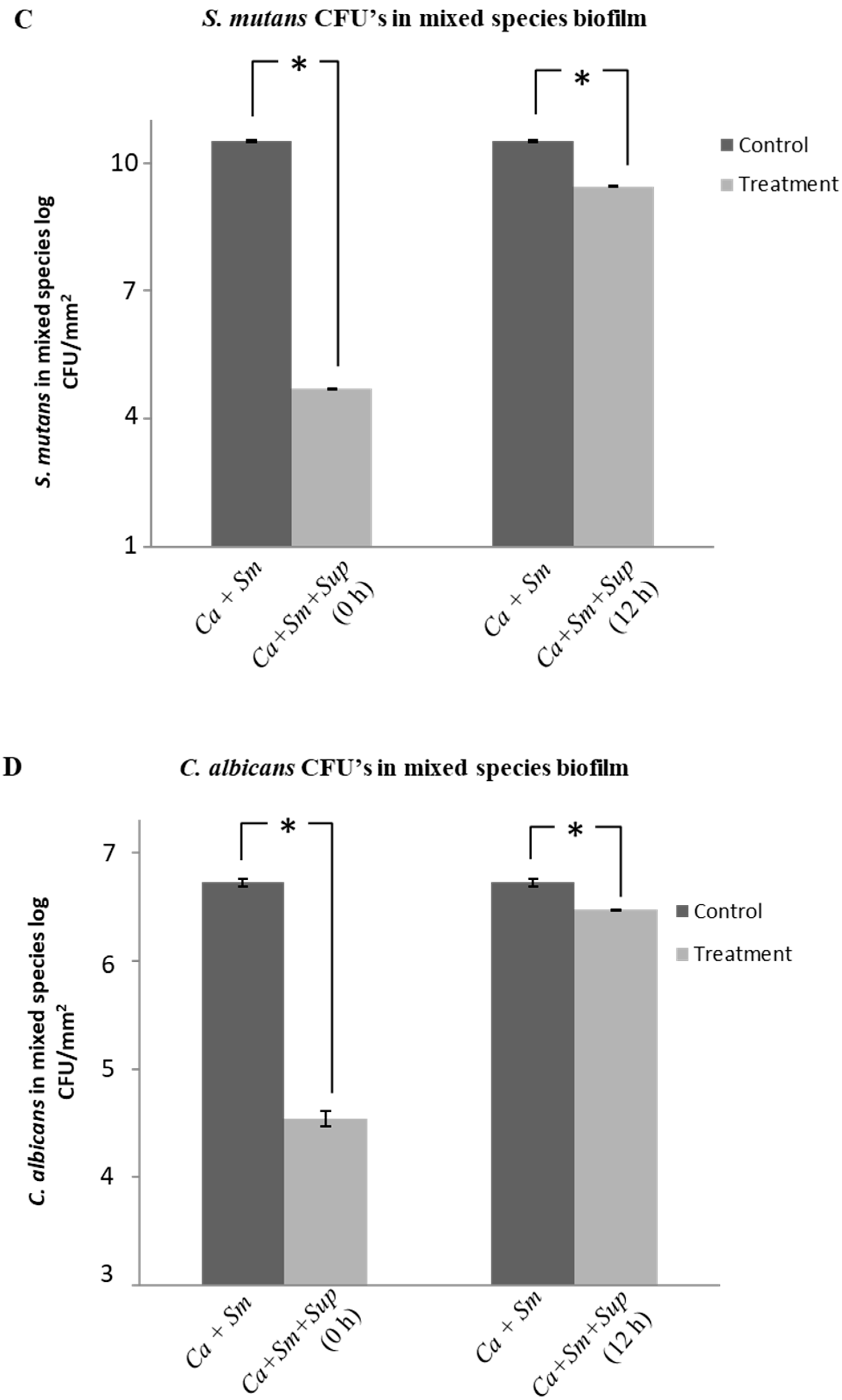
2.4. Effect of Lactobacillus plantarum 108 Supernatant on Streptococcus mutans and Candida albicans Mixed-Species Biofilms
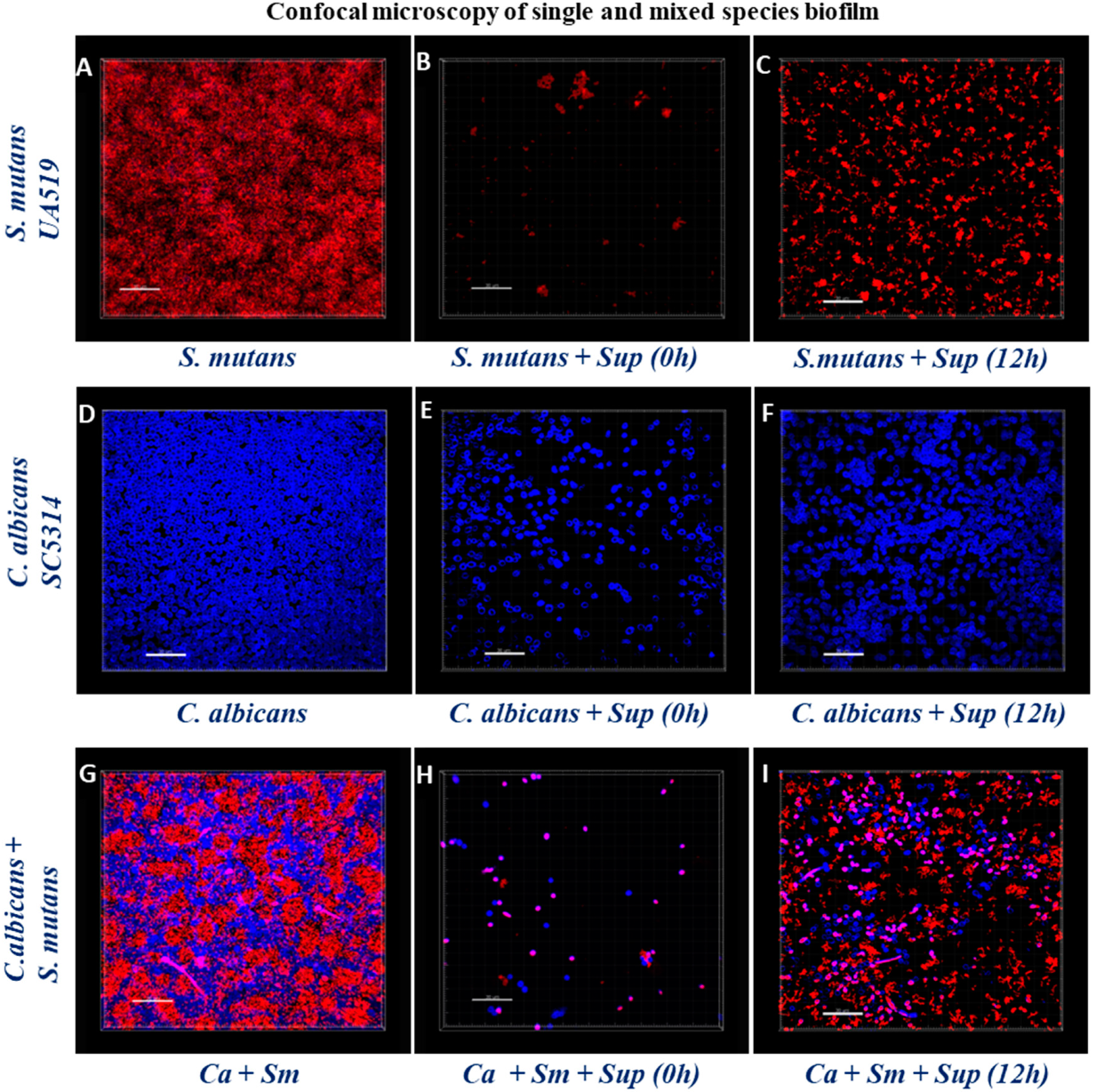
2.5. Structural Analysis of Biofilms by Confocal Laser Scanning Microscopy
2.6. Lactobacillus plantarum 108 Supernatant Down-Regulated the Gtf Gene Expression in Streptococcus mutans Single and Mixed-Species Biofilms
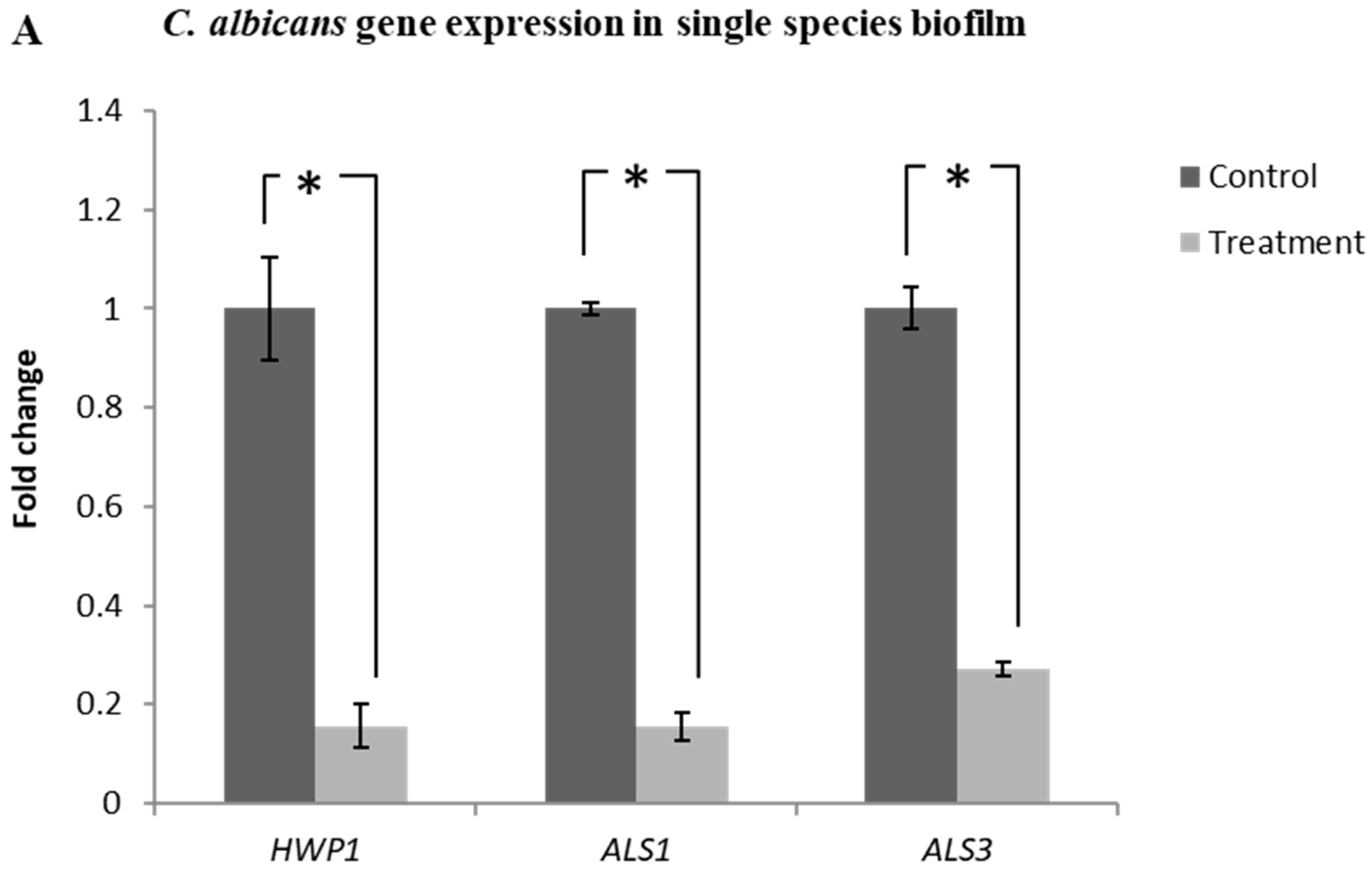
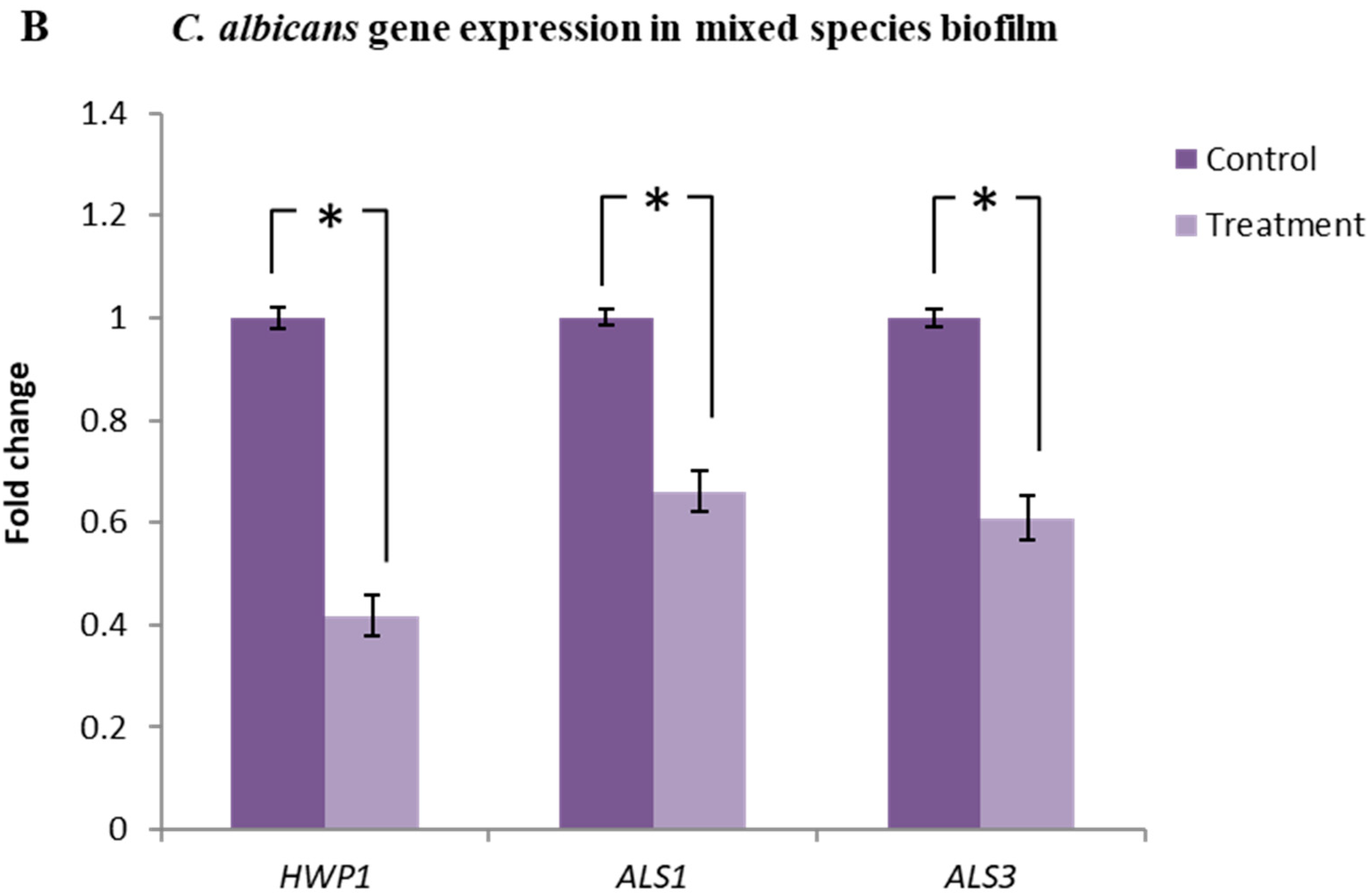
2.7. Lactobacillus plantarum 108 Supernatant Down-Regulated the Expression of HWP1, ALS1 and ALS3 Genes in Candida albicans Single and Mixed-Species Biofilms
3. Discussion
4. Materials and Methods
4.1. Microbial Strains and Culture Conditions
4.2. Preparation of Lactobacillus plantarum 108 Supernatant
4.3. Antimicrobial Activity against Planktonic Streptococcous mutans and Candida albicans
4.4. In Vitro Biofilm Formation and Treatment of Biofilms with Lactobacillus plantarum 108 Supernatant
4.5. Quantification of Biofilms
4.5.1. XTT Reduction Assay
4.5.2. Crystal Violet Assay
4.5.3. Colony-Forming Units Counting
4.6. Confocal Laser Scanning Microscopy
4.7. Gene Expression Analysis by qRT-PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rodriguez, G.; Ruiz, B.; Faleiros, S.; Vistoso, A.; Marro, M.L.; Sanchez, J.; Urzua, I.; Cabello, R. Probiotic compared with standard milk for high-caries children: A cluster randomized trial. J. Dent. Res. 2016, 95, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Hallett, K.B.; O’Rourke, P.K. Early childhood caries and infant feeding practice. Community Dent. Health 2002, 19, 237–242. [Google Scholar] [PubMed]
- Bowen, W.H.; Lawrence, R.A. Comparison of the cariogenicity of cola, honey, cow milk, human milk, and sucrose. Pediatrics 2005, 116, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Chestnutt, I.G.; Murdoch, C.; Robson, K.F. Parents and carers’ choice of drinks for infants and toddlers, in areas of social and economic disadvantage. Community Dent. Health 2003, 20, 139–145. [Google Scholar] [PubMed]
- Nigel, B.P.; Domenick, T.Z.; Phil, D.M.; Kim, E.; Jane, A.W.; Francisco, R.-G.; Junji, T.; Svante, T.; Georgios, T.; Amid, I. Dental caries. Nat. Rev. Dis. Primers 2017, 3, 17030. [Google Scholar]
- Berkowitz, R.J.; Koo, H.; McDermott, M.P.; Whelehan, M.T.; Ragusa, P.; Kopycka-Kedzierawski, D.T.; Karp, J.M.; Billings, R. Adjunctive chemotherapeutic suppression of Mutans Streptococci in the setting of severe early childhood caries: An exploratory study. J. Public Health Dent. 2009, 69, 163–167. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D. Dental biofilms: Difficult therapeutic targets. Periodontology 2000, 28, 12–55. [Google Scholar] [CrossRef]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries. Res. 2011, 45, 69–86. [Google Scholar] [CrossRef]
- Koo, H.; Falsetta, M.L.; Klein, M.I. The Exopolysaccharide matrix: A virulence determinant of cariogenic biofilm. J. Dent. Res. 2013, 92, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, F.G.; Silva, D.S.; Hebling, J.; Spolidorio, L.C.; Spolidorio, D.M. Presence of Mutans Streptococci and Candida spp. in dental plaque/dentine of carious teeth and early childhood caries. Arch. Oral. Biol. 2006, 51, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Raja, M.; Hannan, A.; Ali, K. Association of oral candidal carriage with dental caries in children. Caries Res. 2010, 44, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, S.; Xiao, J.; Silva, B.B.; Gonzalez, I.; Agidi, P.S.; Klein, M.I.; Ambatipudi, K.S.; Rosalen, P.L.; Bauserman, R.; Waugh, R.E.; et al. Role of glucosyltransferase B in interactions of Candida albicans with Streptococcus mutans and with an experimental pellicle on hydroxyapatite surfaces. Appl. Environ. Microbiol. 2011, 77, 6357–6367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira-Cenci, T.; Deng, D.M.; Kraneveld, E.A.; Manders, E.M.; Cury, A.A.; Cate, J.M.; Crielaard, W. The effect of Streptococcus mutans and Candida glabrata on Candida albicans biofilms formed on different surfaces. Arch. Oral. Biol. 2008, 53, 755–764. [Google Scholar] [CrossRef]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms In Vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Metwalli, K.H.; Khan, S.A.; Krom, B.P.; Jabra-Rizk, M.A. Streptococcus mutans, Candida albicans, and the human mouth: A sticky situation. PLoS Pathog. 2013, 9, e1003616. [Google Scholar] [CrossRef] [Green Version]
- Branting, C.; Sund, M.L.; Linder, L.E. The influence of Streptococcus mutans on adhesion of Candida albicans to acrylic surfaces in vitro. Archs. Oral. Biol. 1989, 34, 347–353. [Google Scholar] [CrossRef]
- Ellepola, K.; Liu, Y.; Cao, T.; Koo, H.; Seneviratne, C.J. Bacterial GtfB Augments Candida albicans Accumulation in Cross-Kingdom Biofilms. J. Dent. Res. 2017, 96, 1129–1135. [Google Scholar] [CrossRef]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics analyses reveal synergistic carbohydrate metabolism in Streptococcus mutans-Candida albicans mixed-species biofilms. Infect. Immun. 2019, 87, e00339-19. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Sengupta, A.; Niepa, T.; Lee, B.; Weljie, A.; Freitas-Blanco, V.; Murata, R.; Stebe, K.; Lee, D.; Koo, H. Candida albicans stimulates Streptococcus mutans microcolony development via cross-kingdom biofilm-derived metabolites. Sci. Rep. 2017, 7, 41332. [Google Scholar] [CrossRef]
- Seneviratne, C.J.; Jin, L.; Samaranayake, L.P. Biofilm lifestyle of Candida: A mini review. Oral. Dis. 2008, 14, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug. Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Soderling, E.M.; Marttinen, A.M.; Haukioja, A.L. Probiotic lactobacilli interfere with Streptococcus mutans biofilm formation In Vitro. Curr. Microbiol. 2011, 62, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Tomaro-Duchesneau, C.; Rodes, L.; Malhotra, M.; Tabrizian, M.; Prakash, S. Investigation of probiotic bacteria as dental caries and periodontal disease biotherapeutics. Benef. Microbes 2014, 5, 447–460. [Google Scholar] [CrossRef]
- Ohshima, T.; Kojima, Y.; Seneviratne, C.J.; Maeda, N. Therapeutic application of synbiotics, a fusion of probiotics and prebiotics, and biogenics as a new concept for oral Candida Infections: A Mini Review. Front. Microbiol. 2016, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, L.E.; Millhouse, E.; Sherry, L.; Kean, R.; Malcolm, J.; Nile, C.J.; Ramage, G. Polymicrobial Candida biofilms: Friends and foe in the oral cavity. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Bandara, H.M.; Matsubara, V.H.; Samaranayake, L.P. Future therapies targeted towards eliminating Candida biofilms and associated infections. Expert. Rev. Anti. Infect. Ther. 2017, 15, 299–318. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, T.; Kojima, Y.; Seneviratne, C.J.; Maeda, N. Synbiotics, a fusion of probiotics and prebiotics, and biogenics against oral biofilm associated diseases. In Microbial Biofilms Omics Biology, Antimicrobials and Clinical Implications; CRC Press: Boca Raton, FL, USA, 2017; pp. 237–260. [Google Scholar]
- James, K.M.; MacDonald, K.W.; Chanyi, R.M.; Cadieux, P.A.; Burton, J.P. Inhibition of Candida albicans biofilm formation and modulation of gene expression by probiotic cells and supernatant. J. Med. Microbiol. 2016, 65, 328–336. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Bandara, H.M.; Ishikawa, K.H.; Mayer, M.P.; Samaranayake, L.P. The role of probiotic bacteria in managing periodontal disease: A systematic review. Expert. Rev. Anti. Infect. Ther. 2016, 14, 643–655. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, Y.J. A comparative study of the effect of probiotics on cariogenic biofilm model for preventing dental caries. Arch. Microbiol. 2014, 196, 601–609. [Google Scholar] [CrossRef]
- Krzyściak, W.; Kościelniak, D.; Papież, M.; Vyhouskaya, P.; Zagórska-Świeży, K.; Kołodziej, I.; Bystrowska, B.; Jurczak, A. Effect of a Lactobacillus Salivarius Probiotic on a Double-Species Streptococcus Mutans and Candida Albicans Caries Biofilm. Nutrients 2017, 9, 1242. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Stamatova, I.; Kainulainen, V.; Korpela, R.; Meurman, J.H. Interactions between Lactobacillus rhamnosus GG and oral micro-organisms in an in vitro biofilm model. BMC Microbiol. 2016, 16, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, C.J.; Zeng, G.; Truong, T.; Sze, S.; Wong, W.; Samaranayake, L.; Chan, F.Y.; Wang, Y.-M.; Wang, H.; Gao, J.; et al. New “haploid biofilm model” unravels IRA2 as a novel regulator of Candida albicans biofilm formation. Sci. Rep. 2015, 5, 12433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, 2437. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Oh, J.S.; Lee, H.C.; Lim, H.S.; Lee, S.W.; Yang, K.H.; Choi, N.K.; Kim, S.M. Inhibitory effect of Lactobacillus reuteri on periodontopathic and cariogenic bacteria. J. Microbiol. 2011, 49, 193–199. [Google Scholar] [CrossRef]
- Kojima, Y.; Ohshima, T.; Seneviratne, C.J.; Maeda, N. Combining prebiotics and probiotics to develop novel synbiotics that suppress oral pathogens. J. Oral Biosci. 2016, 58, 27–32. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Wang, Y.; Bandara, H.M.; Mayer, M.P.; Samaranayake, L.P. Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotechnol. 2016, 100, 6415–6426. [Google Scholar] [CrossRef] [Green Version]
- Kheradmand, E.; Rafii, F.; Yazdi, M.H.; Sepahi, A.A. The antimicrobial effects of selenium nanoparticle-enriched probiotics and their fermented broth against Candida albicans. DARU 2014, 22, 48. [Google Scholar] [CrossRef] [Green Version]
- Hasslof, P.; Hedberg, M.; Twetman, S.; Stecksén-Blicks, C. Growth inhibition of oral Mutans Streptococci and Candida by commercial probiotic Lactobacilli - an in vitro study. BMC. Oral. Health. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Ashwin, D.; Ke, V.; Taranath, M.; Ramagoni, N.K.; Nara, A.; Sarpangala, M. Effect of probiotic containing ice-cream on salivary Mutans Streptococci (SMS) levels in children of 6-12 years of age: A randomized controlled double blind study with six-months follow up. J. Clin. Diagn. Res. 2015, 9, ZC06–ZC09. [Google Scholar] [CrossRef]
- Toiviainen, A.; Jalasvuori, H.; Lahti, E.; Gursoy, U.; Salminen, S.; Fontana, M.; Flannagan, S.; Eckert, G.; Kokaras, A.; Paster, B.; et al. Impact of orally administered lozenges with Lactobacillus rhamnosus GG and Bifidobacterium animalis subsp. lactis BB-12 on the number of salivary Mutans Streptococci, amount of plaque, gingival inflammation and the oral microbiome in healthy adults. Clin. Oral. Investig. 2015, 19, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraft-Bodi, E.; Jorgensen, M.R.; Keller, M.K.; Kragelund, C.; Twetman, S. Effect of probiotic bacteria on oral Candida in frail elderly. J. Dent. Res. 2015, 94, 181S–186S. [Google Scholar] [CrossRef] [PubMed]
- Sutula, J.; Coulthwaite, L.A.; Thomas, L.V.; Verran, J. The effect of a commercial probiotic drink containing Lactobacillus casei strain Shirota on oral health in healthy dentate people. Microb. Ecol. Health Dis. 2013, 24, 21003. [Google Scholar]
- Mendonca, F.H.; Santos, S.S.; Faria, I.D.; Goncalves-e-Silva, C.R.; Jorge, A.O.; Leão, M.V. Effects of probiotic bacteria on Candida presence and IgA pnti-Candida in the oral oavity of elderly. Braz. Dental. J. 2012, 23, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.K.; Srivastava, S. Purification and characterization of plantaricin LR14: A novel bacteriocin produced by Lactobacillus plantarum LR/14. Appl. Microbiol. Biotechnol. 2008, 79, 759–767. [Google Scholar] [CrossRef]
- Sharma, A.; Srivastava, S. Anti-Candida activity of spent culture filtrate of Lactobacillus plantarum strain LR/14. J. Mycol. Med. 2013, 24, 25–34. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Caola, I.; Nollo, G.; Cavallo, M.; Rinaldi, M.; Fracchia, L. Inhibition of Candida albicans adhesion on medical-grade silicone by a Lactobacillus-derived biosurfactant. J. Appl. Microbiol. 2015, 118, 1116–1125. [Google Scholar] [CrossRef]
- Rodrigues, L.; Banat, I.M.; Teixeira, J.; Oliveira, R. Biosurfactants: Potential applications in medicine. J. Antimicrob. Chemother. 2006, 57, 609–618. [Google Scholar] [CrossRef]
- Saravanakumari, P.; Mani, K. Structural characterization of a novel xylolipid biosurfactant from Lactococcus lactis and analysis of antibacterial activity against multi-drug resistant pathogens. Bioresour. Technol. 2010, 101, 8851–8854. [Google Scholar] [CrossRef]
- Fracchia, L.; Cavallo, M.; Allegrone, G.; Martinotti, M.G. A Lactobacillus-derived biosurfactant inhibits biofilm formation of human pathogenic Candida albicans biofilm producers. Appl. Microbiol. Biotechnol. 2010, 2, 827–837. [Google Scholar]
- Kuyukina, M.S.; Ivshina, I.B.; Korshunova, I.O.; Stukova, G.I.; Krivoruchko, A.V. Diverse effects of a biosurfactant from Rhodococcus ruber IEGM 231 on the adhesion of resting and growing bacteria to polystyrene. AMB Express 2016, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattos-Graner, R.O.; Smith, D.J.; King, W.F.; Mayer, M. Water-insoluble glucan synthesis by Mutans Streptococcal strains correlates with caries incidence in 12- to 30-month-old children. J. Dent. Res. 2000, 79, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Salehi, R.; Savabi, O.; Kazemi, M.; Kamali, S.; Salehi, A.R.; Eslami, G.; Tahmourespour, A. Effects of Lactobacillus reuteri-derived biosurfactant on the gene expression profile of essential adhesion genes (gtfB, gtfC and ftf) of Streptococcus mutans. Adv. Biomed. Res. 2014, 3, 169. [Google Scholar] [CrossRef] [PubMed]
- Savabi, O.; Kazemi, M.; Kamali, S.; Salehi, A.R.; Eslami, G.; Tahmourespour, A.; Salehi, R. Effects of biosurfactant produced by Lactobacillus casei on gtfB, gtfC, and ftf gene expression level in S. mutans by real-time RT-PCR. Adv. Biomed. Res. 2014, 3, 231. [Google Scholar] [PubMed]
- Tahmourespour, A.; Salehi, R.; Kermanshahi, R.K.; Eslami, G. The anti-biofouling effect of Lactobacillus fermentum-derived biosurfactant against Streptococcus mutans. Biofouling 2011, 27, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Murciano, C.; Moyes, D.L.; Runglall, M.; Tobouti, P.; Islam, A.; Hoyer, L.L.; Naglik, J.R. Evaluation of the role of Candida albicans agglutinin-like sequence (Als) proteins in human oral epithelial cell interactions. PLoS ONE 2012, 7, e33362. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Nett, J.E.; Andes, D.R.; Mitchell, A.P. Function of Candida albicans adhesin Hwp1 in biofilm formation. Eukaryot. Cell. 2006, 5, 1604–1610. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Andes, D.R.; Nett, J.E.; Smith, F.J.; Yue, F.; Phan, Q.T.; Edwards, J.E.; Filler, S.G.; Mitchell, A.P. Critical role of Bcr1-dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog. 2006, 2, e63. [Google Scholar] [CrossRef]
- Bandara, H.M.; Cheung, B.P.; Watt, R.M.; Jin, L.J.; Samaranayake, L.P. Secretory products of Escherichia coli biofilm modulate Candida biofilm formation and hyphal development. J. Investig. Clin. Dent. 2013, 4, 186–199. [Google Scholar] [CrossRef]
- Finkel, J.S.; Mitchell, A.P. Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 2011, 9, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, 5439. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Theberge, S.; Semlali, A.; Alamri, A.; Leung, K.P.; Rouabhia, M.C. albicans growth, transition, biofilm formation and gene expression modulation by antimicrobial decapeptide KSL-W. BMC Microbiol. 2013, 13, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence Used for Streptococcus mutans | ||
|---|---|---|
| Gene | Primer Sequence (5’-3’) | |
| 16sRNA | F: | CCT ACG GGA GGC AGC AGT AG |
| R: | CAA CAG AGC TTT ACG ATC CGA AA | |
| gtfB | F: | AGC AAT GCA GCC AAT CTA CAA AT |
| R: | ACG AAC TTT GCC GTT ATT GTC A | |
| gtfC | F: | GGT TTA ACG TCA AAA TTA GCT GTA TTA GC |
| R: | CTC AAC CAA CCG CCA CTG TT | |
| gtfD | F: | ACA GCA GAC AGC AGC CAA GA |
| R: | ACT GGG TTT GCT GCG TTT G | |
| Primer Sequence Used for Candida albicans | ||
|---|---|---|
| Gene | Primer Sequence (5’-3’) | |
| PMA1 | F: | TTGAAGATGACCACCCAATCC |
| R: | GAAACCTCTGGAAGCAAATTCG | |
| HWP1 | F: | GCTCAACTTATTGCTATCGCTTATTACA |
| R: | GACCGTCTACCTGTGGGACAGT | |
| ALS1 | F: | GAC TAG TGA ACC AAC AAA TAC CAG A |
| R: | CCA GAA GAA ACA GCA GGT GA | |
| ALS3 | F: | AATGGTCCTTATGAATCACCATCTACTA |
| R: | GAGTTTTCATCCATACTTGATTTCACAT | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srivastava, N.; Ellepola, K.; Venkiteswaran, N.; Chai, L.Y.A.; Ohshima, T.; Seneviratne, C.J. Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation. Antibiotics 2020, 9, 478. https://doi.org/10.3390/antibiotics9080478
Srivastava N, Ellepola K, Venkiteswaran N, Chai LYA, Ohshima T, Seneviratne CJ. Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation. Antibiotics. 2020; 9(8):478. https://doi.org/10.3390/antibiotics9080478
Chicago/Turabian StyleSrivastava, Neha, Kassapa Ellepola, Nityasri Venkiteswaran, Louis Yi Ann Chai, Tomoko Ohshima, and Chaminda Jayampath Seneviratne. 2020. "Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation" Antibiotics 9, no. 8: 478. https://doi.org/10.3390/antibiotics9080478
APA StyleSrivastava, N., Ellepola, K., Venkiteswaran, N., Chai, L. Y. A., Ohshima, T., & Seneviratne, C. J. (2020). Lactobacillus Plantarum 108 Inhibits Streptococcus mutans and Candida albicans Mixed-Species Biofilm Formation. Antibiotics, 9(8), 478. https://doi.org/10.3390/antibiotics9080478