
Extracellular Matrix Remodeling in the Retina and Optic Nerve of a Novel Glaucoma Mouse Model


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. RNA Isolation, cDNA Synthesis, and RT-qPCR
2.3. Immunohistochemistry and Confocal Laser Scanning Microscopy
2.4. Western Blotting
2.5. Statistics
3. Results
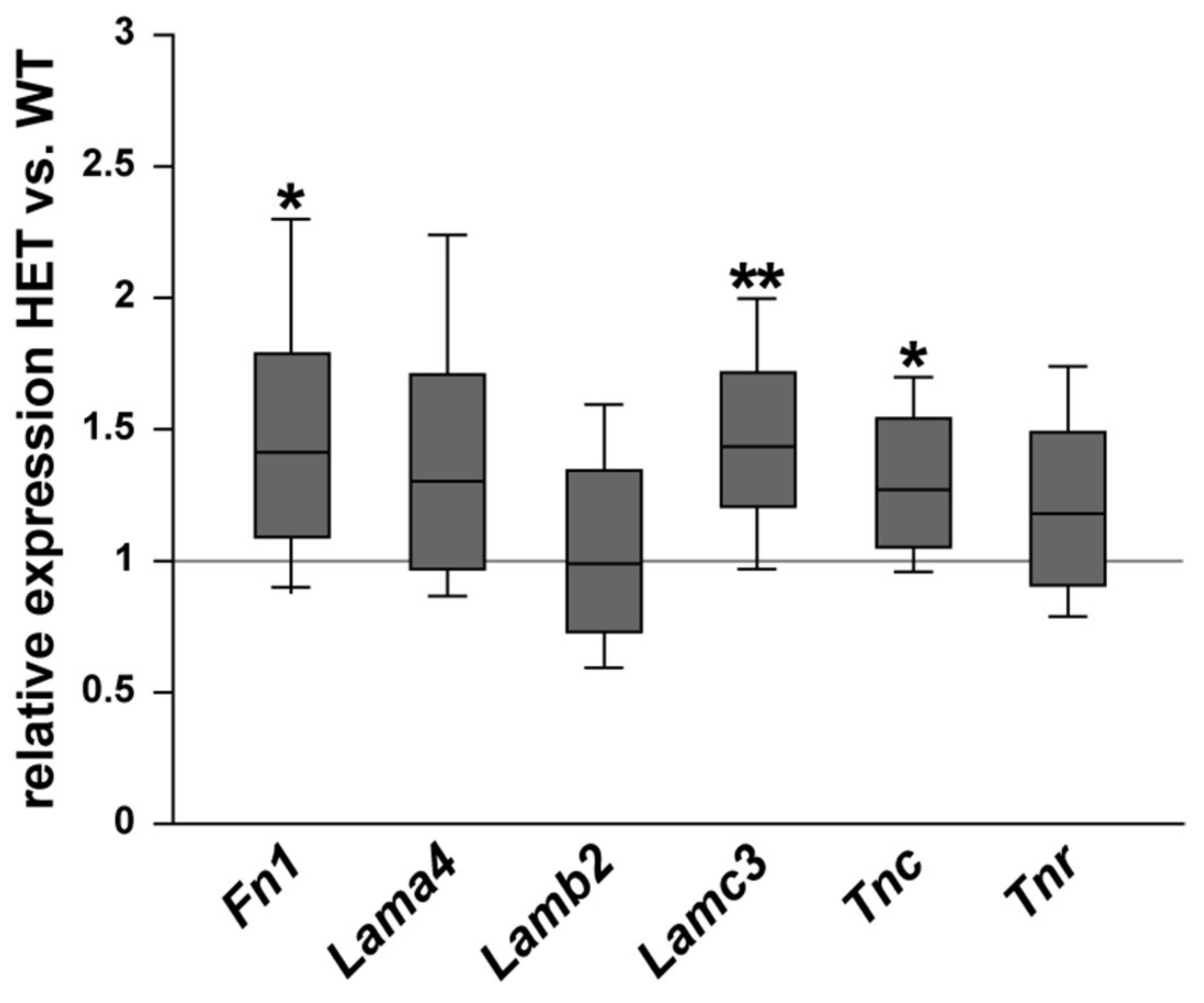
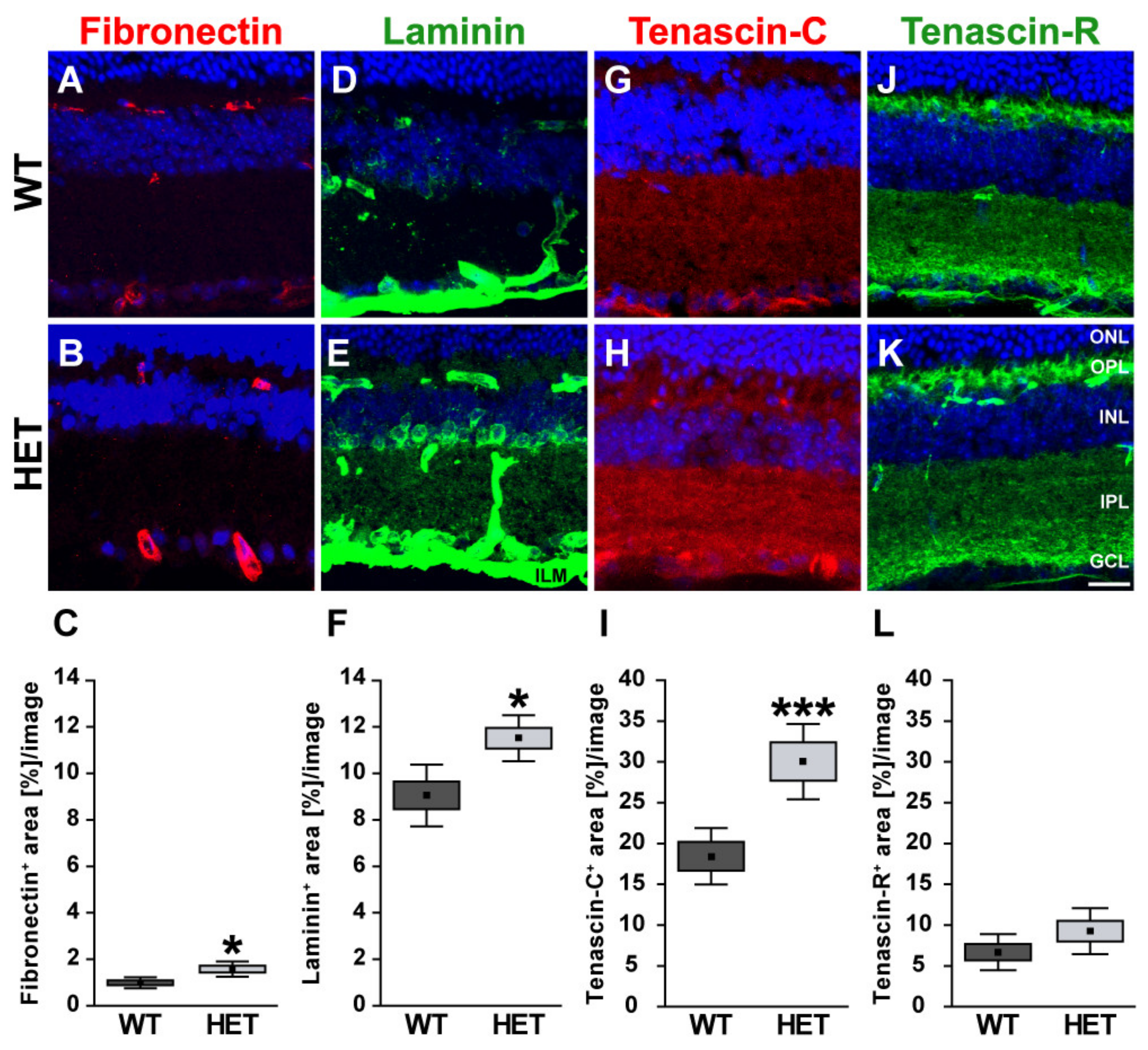
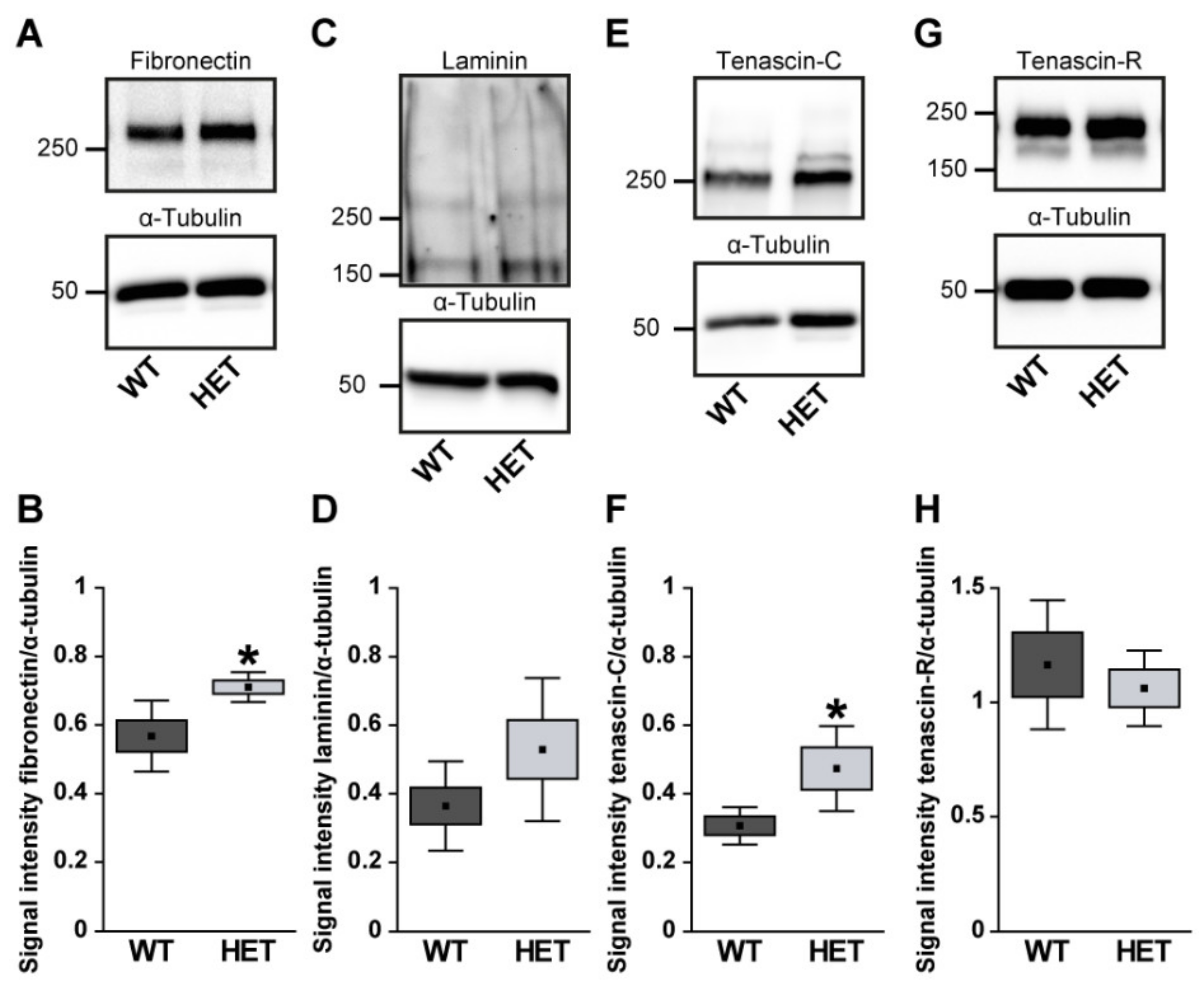
3.1. Remodeling of Glycoproteins in the Glaucomatous Retina of PTP-Meg2 HET Mice
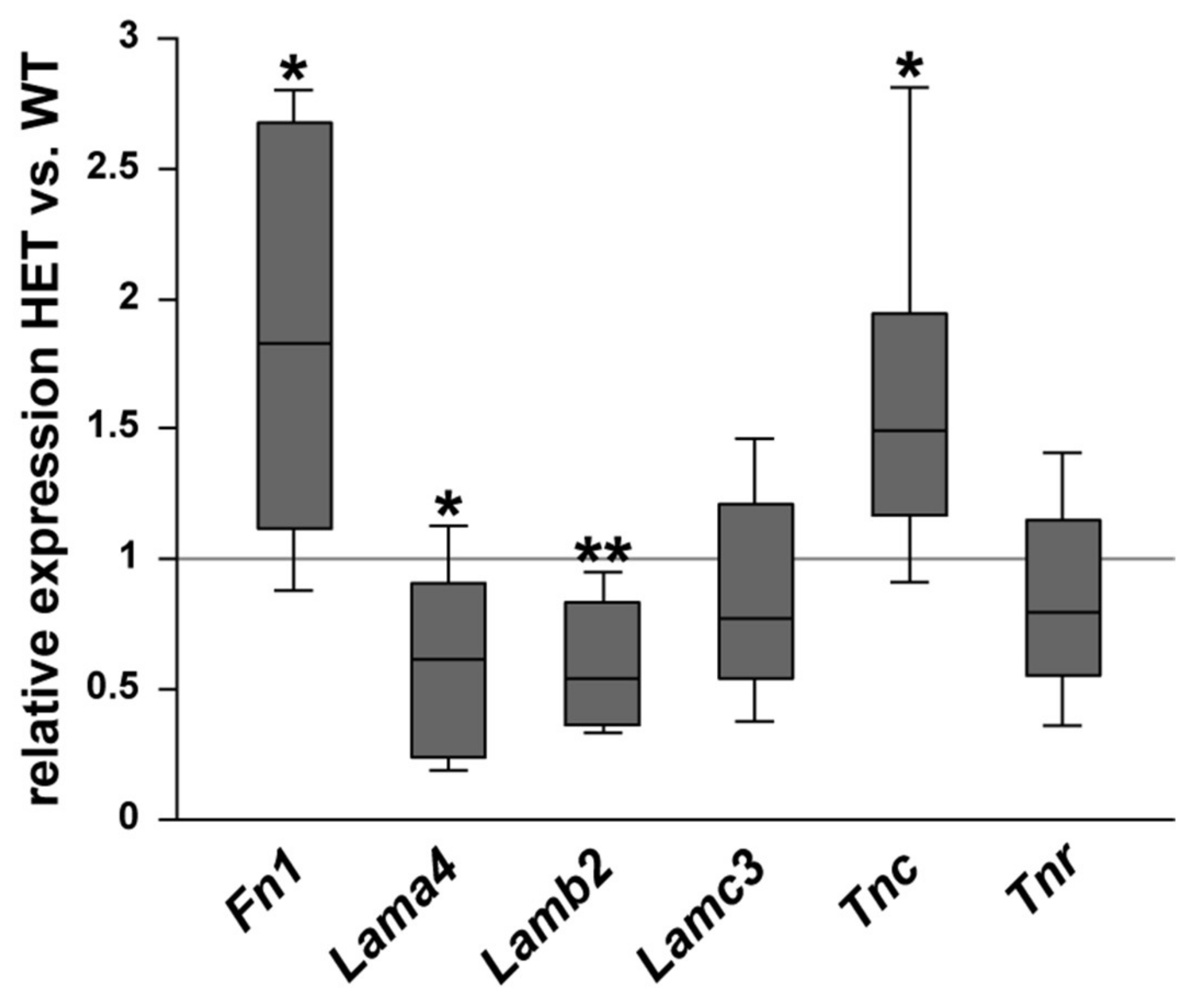
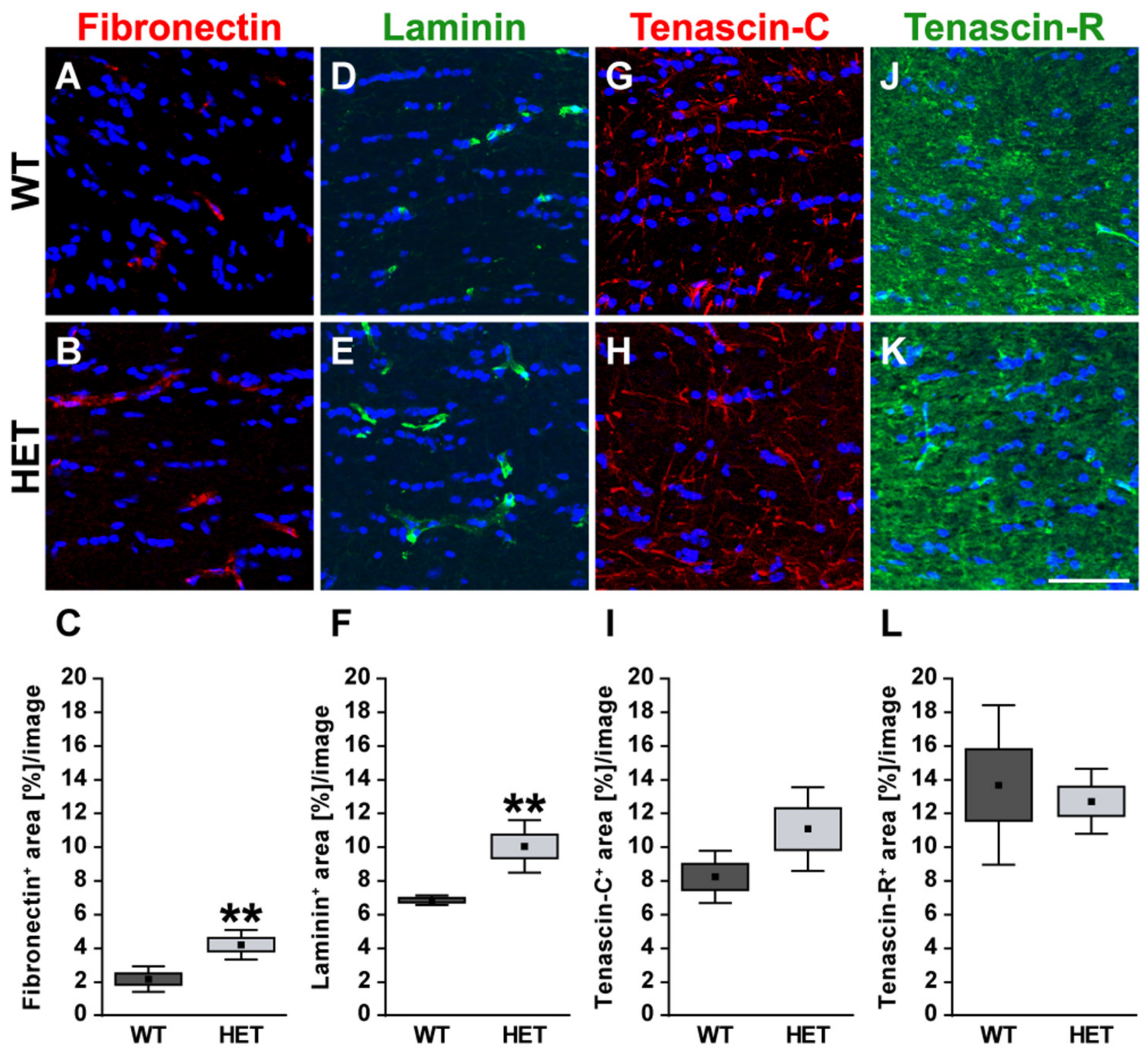
3.2. Remodeling of Glycoproteins in the Glaucomatous Optic Nerve of PTP-Meg2 HET Mice
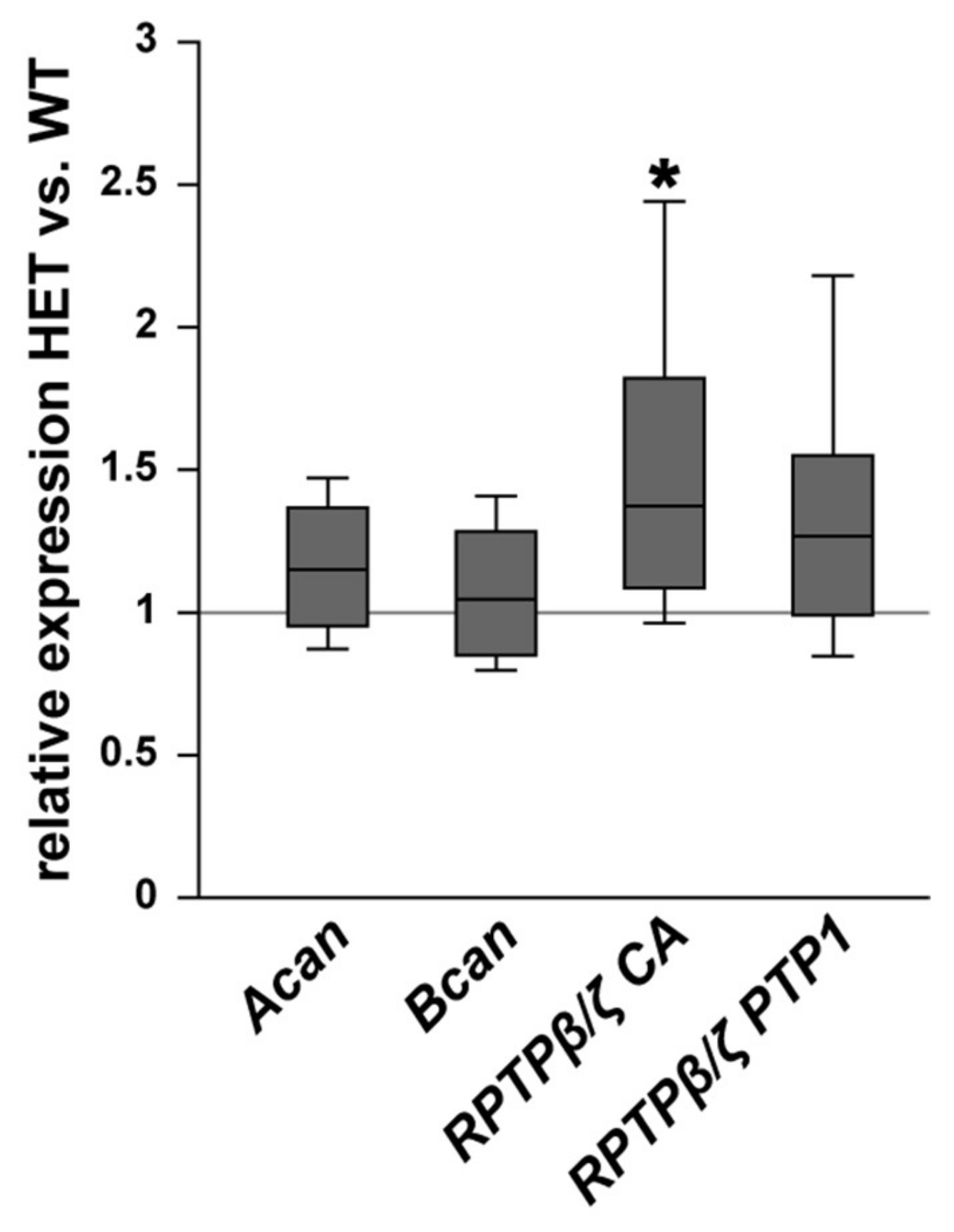
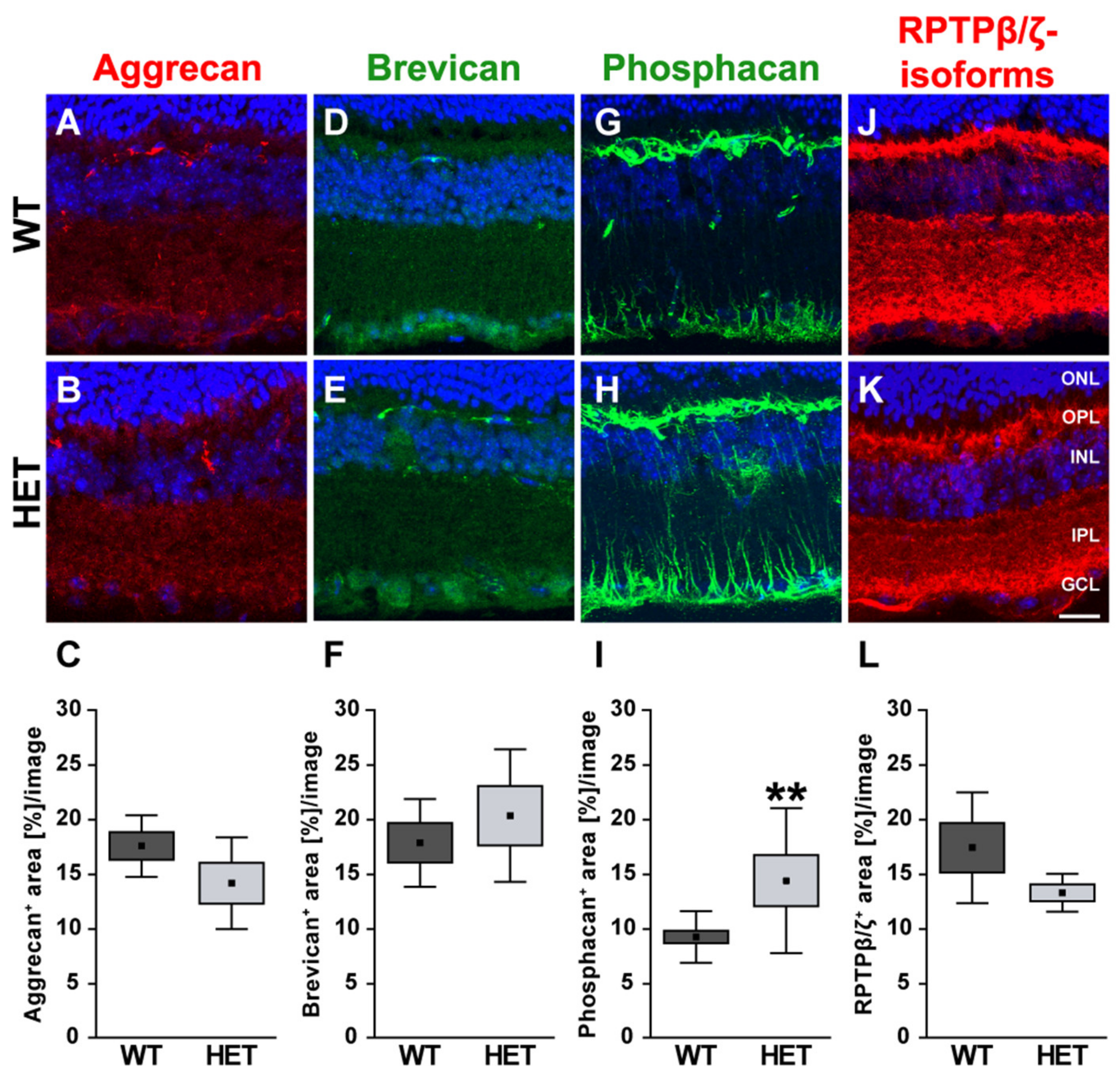
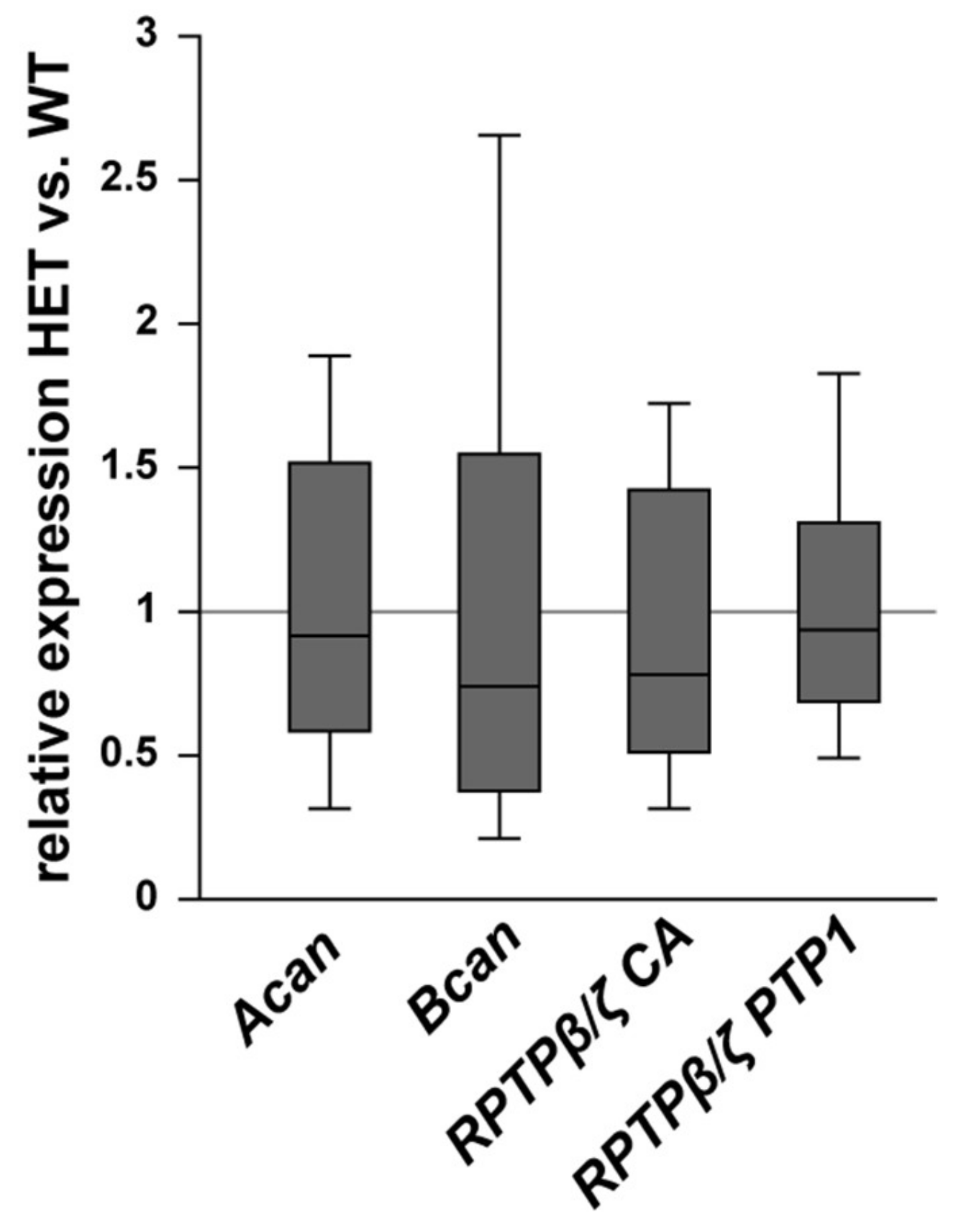
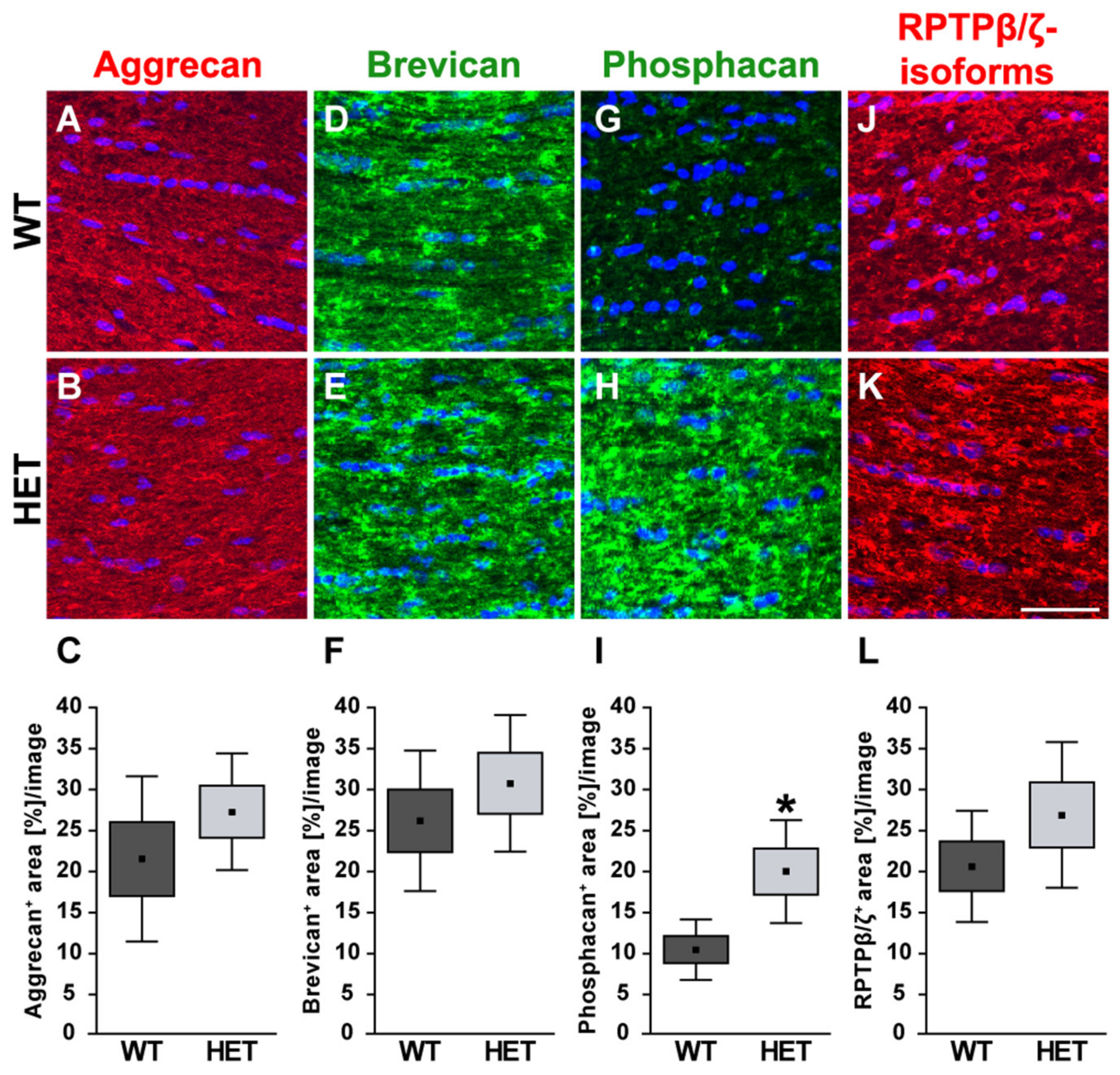
3.3. Remodeling of Proteoglycans in the Glaucomatous Retina of PTP-Meg2 HET Mice
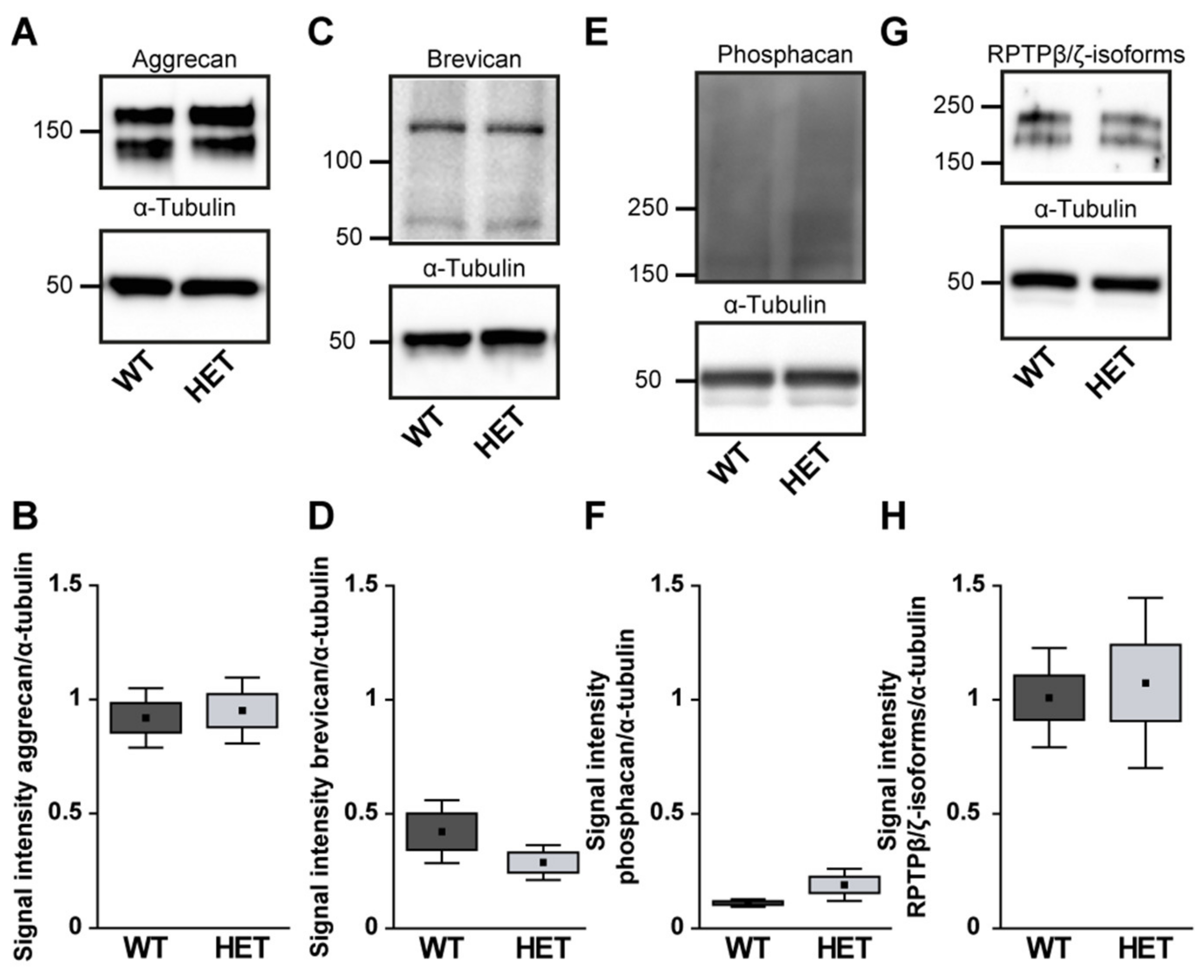
3.4. Remodeling of Proteoglycans in the Glaucomatous Optic Nerve of PTP-Meg2 HET Mice
4. Discussion
4.1. Increased Levels of the ECM Glycoprotein Fibronectin in the Glaucomatous Retina and Optic Nerve
4.2. Altered Isoform-Specific Regulation of Laminin in Glaucomatous Tissue
4.3. Upregulation of Tenascin-C in the Glaucomatous Retina and Optic Nerve
4.4. Unchanged levels of Tenascin-R
4.5. Increased Expression of the RPTPβ/ζ isoform Phosphacan in the Glaucomatous Retina
4.6. Impact of ECM Changes on Glaucomatous RGC Death
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Bp | base pairs |
| CA | carbonic anhydrase-like domain |
| CSPG | chondroitin sulfate proteoglycan |
| ECM | extracellular matrix |
| For | forward |
| GCL | ganglion cell layer |
| GFAP | glial fibrillary acidic protein |
| HET | heterozygous |
| ILM | inner limiting membrane |
| INL | inner nuclear layer |
| IOP | intraocular pressure |
| IPL | inner plexiform layer |
| ONL | outer nuclear layer |
| OPL | outer plexiform layer |
| PBS | phosphate-buffered saline |
| POAG | primary open-angle glaucoma |
| PTP1 | protein tyrosine phosphatase domain 1 |
| PTP-Meg2 | protein tyrosine phosphatase megakaryocyte 2 |
| Rev | reverse |
| RGC | retinal ganglion cell |
| RPTPβ/ζ | receptor protein tyrosine phosphatase beta/zeta |
| RT-qPCR | quantitative real-time polymerase chain reaction |
| SD | standard deviation |
| SEM | standard error of the mean |
| TLR4 | toll-like receptor 4 |
| WT | wild type |
References
- European Glaucoma Society Terminology and Guidelines for Glaucoma, 4th Edition—Chapter 2: Classification and terminology Supported by the EGS Foundation: Part 1: Foreword; Introduction; Glossary; Chapter 2 Classification and Terminology. Br. J. Ophthalmol. 2017, 101, 73–127. [CrossRef] [Green Version]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, J.; Wiemann, S.; Joachim, S.C.; Palmhof, M.; Woestmann, J.; Denecke, B.; Wang, Y.; Downey, G.P.; Faissner, A. Heterozygous Meg2 Ablation Causes Intraocular Pressure Elevation and Progressive Glaucomatous Neurodegeneration. Mol. Neurobiol. 2019, 56, 4322–4345. [Google Scholar] [CrossRef] [PubMed]
- Mecham, R.P. Overview of extracellular matrix. Curr. Protoc. Cell Biol. 2012, 57, 10.1.1–10.1.16. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Naba, A. Overview of the matrisome—An inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [Green Version]
- Faissner, A.; Reinhard, J. The extracellular matrix compartment of neural stem and glial progenitor cells. Glia 2015, 63, 1330–1349. [Google Scholar] [CrossRef]
- Krishnaswamy, V.R.; Benbenishty, A.; Blinder, P.; Sagi, I. Demystifying the extracellular matrix and its proteolytic remodeling in the brain: Structural and functional insights. Cell. Mol. Life Sci. 2019, 76, 3229–3248. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, J.; Joachim, S.C.; Faissner, A. Extracellular matrix remodeling during retinal development. Exp. Eye Res. 2015, 133, 132–140. [Google Scholar] [CrossRef]
- Gandhi, M.; Dubey, S. Evaluation of the Optic Nerve Head in Glaucoma. J. Curr. Glaucoma Pr. 2013, 7, 106–114. [Google Scholar]
- Quigley, H.A. Neuronal death in glaucoma. Prog. Retin. Eye Res. 1999, 18, 39–57. [Google Scholar] [CrossRef]
- Hernandez, M.R. Ultrastructural immunocytochemical analysis of elastin in the human lamina cribrosa. Changes in elastic fibers in primary open-angle glaucoma. Invest. Ophthalmol. Vis. Sci. 1992, 33, 2891–2903. [Google Scholar]
- Hernandez, M.R.; Andrzejewska, W.M.; Neufeld, A.H. Changes in the extracellular matrix of the human optic nerve head in primary open-angle glaucoma. Am. J. Ophthalmol. 1990, 109, 180–188. [Google Scholar] [CrossRef]
- Morrison, J.C.; Dorman-Pease, M.E.; Dunkelberger, G.R.; Quigley, H.A. Optic nerve head extracellular matrix in primary optic atrophy and experimental glaucoma. Arch. Ophthalmol. 1990, 108, 1020–1024. [Google Scholar] [CrossRef]
- Pena, J.D.; Agapova, O.; Gabelt, B.T.; Levin, L.A.; Lucarelli, M.J.; Kaufman, P.L.; Hernandez, M.R. Increased elastin expression in astrocytes of the lamina cribrosa in response to elevated intraocular pressure. Invest. Ophthalmol. Vis. Sci. 2001, 42, 2303–2314. [Google Scholar]
- Reinhard, J.; Renner, M.; Wiemann, S.; Shakoor, D.A.; Stute, G.; Dick, H.B.; Faissner, A.; Joachim, S.C. Ischemic injury leads to extracellular matrix alterations in retina and optic nerve. Sci. Rep. 2017, 7, 43470. [Google Scholar] [CrossRef]
- Miller, C.G.; Budoff, G.; Prenner, J.L.; Schwarzbauer, J.E. Minireview: Fibronectin in retinal disease. Exp. Biol. Med. (Maywood) 2017, 242, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Sato, T.; Paryani, G.; Kao, R. Downregulation of fibronectin overexpression reduces basement membrane thickening and vascular lesions in retinas of galactose-fed rats. Diabetes 2003, 52, 1229–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasetti, R.B.; Maddineni, P.; Millar, J.C.; Clark, A.F.; Zode, G.S. Increased synthesis and deposition of extracellular matrix proteins leads to endoplasmic reticulum stress in the trabecular meshwork. Sci. Rep. 2017, 7, 14951. [Google Scholar] [CrossRef] [Green Version]
- Faralli, J.A.; Filla, M.S.; Peters, D.M. Role of Fibronectin in Primary Open Angle Glaucoma. Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.L.; Mavlyutov, T.A.; Perlmutter, T.E.; Curry, S.M.; Harris, S.L.; Chauhan, A.K.; McDowell, C.M. Fibronectin extra domain A (FN-EDA) elevates intraocular pressure through Toll-like receptor 4 signaling. Sci. Rep. 2020, 10, 9815. [Google Scholar] [CrossRef] [PubMed]
- Libby, R.T.; Champliaud, M.F.; Claudepierre, T.; Xu, Y.; Gibbons, E.P.; Koch, M.; Burgeson, R.E.; Hunter, D.D.; Brunken, W.J. Laminin expression in adult and developing retinae: Evidence of two novel CNS laminins. J. Neurosci. 2000, 20, 6517–6528. [Google Scholar] [CrossRef]
- Edwards, M.M.; Lefebvre, O. Laminins and retinal vascular development. Cell Adhes. Migr. 2013, 7, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Thyboll, J.; Kortesmaa, J.; Cao, R.; Soininen, R.; Wang, L.; Iivanainen, A.; Sorokin, L.; Risling, M.; Cao, Y.; Tryggvason, K. Deletion of the laminin alpha4 chain leads to impaired microvessel maturation. Mol. Cell. Biol. 2002, 22, 1194–1202. [Google Scholar] [CrossRef] [Green Version]
- Yousif, L.F.; Di Russo, J.; Sorokin, L. Laminin isoforms in endothelial and perivascular basement membranes. Cell Adhes. Migr. 2013, 7, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Pinzon-Duarte, G.; Daly, G.; Li, Y.N.; Koch, M.; Brunken, W.J. Defective formation of the inner limiting membrane in laminin beta2- and gamma3-null mice produces retinal dysplasia. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1773–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnanaguru, G.; Bachay, G.; Biswas, S.; Pinzon-Duarte, G.; Hunter, D.D.; Brunken, W.J. Laminins containing the beta2 and gamma3 chains regulate astrocyte migration and angiogenesis in the retina. Development 2013, 140, 2050–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, J.; Roll, L.; Faissner, A. Tenascins in Retinal and Optic Nerve Neurodegeneration. Front. Integr. Neurosci. 2017, 11, 30. [Google Scholar] [CrossRef]
- D’Alessandri, L.; Ranscht, B.; Winterhalter, K.H.; Vaughan, L. Contactin/F11 and tenascin-C co-expression in the chick retina correlates with formation of the synaptic plexiform layers. Curr. Eye Res. 1995, 14, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, S.; Reinhard, J.; Faissner, A. Immunomodulatory role of the extracellular matrix protein tenascin-C in neuroinflammation. Biochem. Soc. Trans. 2019, 47, 1651–1660. [Google Scholar] [CrossRef]
- Jakovcevski, I.; Miljkovic, D.; Schachner, M.; Andjus, P.R. Tenascins and inflammation in disorders of the nervous system. Amino Acids 2013, 44, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Dzyubenko, E.; Manrique-Castano, D.; Kleinschnitz, C.; Faissner, A.; Hermann, D.M. Role of immune responses for extracellular matrix remodeling in the ischemic brain. Adv. Neurol. Disord. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Reinhard, J.; Wiemann, S.; Stute, G.; Kuehn, S.; Woestmann, J.; Dick, H.B.; Faissner, A.; Joachim, S.C. Early remodelling of the extracellular matrix proteins tenascin-C and phosphacan in retina and optic nerve of an experimental autoimmune glaucoma model. J. Cell. Mol. Med. 2016, 20, 2122–2137. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.C.; Jia, L.; Cepurna, W.O.; Doser, T.A.; Morrison, J.C. Global changes in optic nerve head gene expression after exposure to elevated intraocular pressure in a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3161–3177. [Google Scholar] [CrossRef] [PubMed]
- Pesheva, P.; Gloor, S.; Schachner, M.; Probstmeier, R. Tenascin-R is an intrinsic autocrine factor for oligodendrocyte differentiation and promotes cell adhesion by a sulfatide-mediated mechanism. J. Neurosci. 1997, 17, 4642–4651. [Google Scholar] [CrossRef] [Green Version]
- Pesheva, P.; Probstmeier, R.; Skubitz, A.P.; McCarthy, J.B.; Furcht, L.T.; Schachner, M. Tenascin-R (J1 160/180 inhibits fibronectin-mediated cell adhesion--functional relatedness to tenascin-C. J. Cell Sci. 1994, 107, 2323–2333. [Google Scholar]
- Eskici, N.F.; Erdem-Ozdamar, S.; Dayangac-Erden, D. The altered expression of perineuronal net elements during neural differentiation. Cell. Mol. Biol. Lett. 2018, 23, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartsch, U.; Pesheva, P.; Raff, M.; Schachner, M. Expression of janusin (J1-160/180) in the retina and optic nerve of the developing and adult mouse. Glia 1993, 9, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Wintergerst, E.S.; Fuss, B.; Bartsch, U. Localization of janusin mRNA in the central nervous system of the developing and adult mouse. Eur. J. Neurosci. 1993, 5, 299–310. [Google Scholar] [CrossRef]
- Roll, L.; Faissner, A. Tenascins in CNS lesions. Semin. Cell Dev. Biol. 2019, 89, 118–124. [Google Scholar] [CrossRef]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Dissecting spinal cord regeneration. Nature 2018, 557, 343–350. [Google Scholar] [CrossRef]
- Wang, H.; Song, G.; Chuang, H.; Chiu, C.; Abdelmaksoud, A.; Ye, Y.; Zhao, L. Portrait of glial scar in neurological diseases. Int. J. Immunopathol. Pharm. 2018, 31. [Google Scholar] [CrossRef] [Green Version]
- Pearson, C.S.; Solano, A.G.; Tilve, S.M.; Mencio, C.P.; Martin, K.R.; Geller, H.M. Spatiotemporal distribution of chondroitin sulfate proteoglycans after optic nerve injury in rodents. Exp. Eye Res. 2020, 190, 107859. [Google Scholar] [CrossRef]
- Klausmeyer, A.; Garwood, J.; Faissner, A. Differential expression of phosphacan/RPTPbeta isoforms in the developing mouse visual system. J. Comp. Neurol. 2007, 504, 659–679. [Google Scholar] [CrossRef]
- Seidenbecher, C.I.; Richter, K.; Rauch, U.; Fassler, R.; Garner, C.C.; Gundelfinger, E.D. Brevican, a chondroitin sulfate proteoglycan of rat brain, occurs as secreted and cell surface glycosylphosphatidylinositol-anchored isoforms. J. Biol. Chem. 1995, 270, 27206–27212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czopka, T.; Von Holst, A.; Schmidt, G.; Ffrench-Constant, C.; Faissner, A. Tenascin C and tenascin R similarly prevent the formation of myelin membranes in a RhoA-dependent manner, but antagonistically regulate the expression of myelin basic protein via a separate pathway. Glia 2009, 57, 1790–1801. [Google Scholar] [CrossRef] [PubMed]
- Faissner, A.; Kruse, J. J1/tenascin is a repulsive substrate for central nervous system neurons. Neuron 1990, 5, 627–637. [Google Scholar] [CrossRef]
- Rathjen, F.G.; Wolff, J.M.; Chiquet-Ehrismann, R. Restrictin: A chick neural extracellular matrix protein involved in cell attachment co-purifies with the cell recognition molecule F11. Development 1991, 113, 151–164. [Google Scholar]
- Faissner, A.; Clement, A.; Lochter, A.; Streit, A.; Mandl, C.; Schachner, M. Isolation of a neural chondroitin sulfate proteoglycan with neurite outgrowth promoting properties. J. Cell Biol. 1994, 126, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, D.; Franco, C.A.; Estrach, S.; Mettouchi, A.; Sauvaget, D.; Rosewell, I.; Schertel, A.; Armer, H.; Domogatskaya, A.; Rodin, S.; et al. Endothelial basement membrane limits tip cell formation by inducing Dll4/Notch signalling in vivo. EMBO Rep. 2011, 12, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecino, E.; Heller, J.P.; Veiga-Crespo, P.; Martin, K.R.; Fawcett, J.W. Influence of extracellular matrix components on the expression of integrins and regeneration of adult retinal ganglion cells. PLoS ONE 2015, 10, e0125250. [Google Scholar] [CrossRef] [Green Version]
- Mooradian, D.L.; Lucas, R.C.; Weatherbee, J.A.; Furcht, L.T. Transforming growth factor-beta 1 binds to immobilized fibronectin. J. Cell. Biochem. 1989, 41, 189–200. [Google Scholar] [CrossRef]
- Hoshijima, M.; Hattori, T.; Inoue, M.; Araki, D.; Hanagata, H.; Miyauchi, A.; Takigawa, M. CT domain of CCN2/CTGF directly interacts with fibronectin and enhances cell adhesion of chondrocytes through integrin alpha5beta1. FEBS Lett. 2006, 580, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.M.; Mitsi, M.; Nugent, M.A.; Symes, K. PDGF-A interactions with fibronectin reveal a critical role for heparan sulfate in directed cell migration during Xenopus gastrulation. Proc. Natl. Acad. Sci. USA 2009, 106, 21683–21688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, M.M.; Hubbell, J.A. The 12th-14th type III repeats of fibronectin function as a highly promiscuous growth factor-binding domain. FASEB J. 2010, 24, 4711–4721. [Google Scholar]
- Simha, A.; Braganza, A.; Abraham, L.; Samuel, P.; Lindsley, K. Anti-vascular endothelial growth factor for neovascular glaucoma. Cochrane Database Syst. Rev. 2013, CD007920. [Google Scholar] [CrossRef] [Green Version]
- SooHoo, J.R.; Seibold, L.K.; Kahook, M.Y. Recent advances in the management of neovascular glaucoma. Semin. Ophthalmol. 2013, 28, 165–172. [Google Scholar] [CrossRef]
- Hohenester, E.; Yurchenco, P.D. Laminins in basement membrane assembly. Cell Adhes. Migr. 2013, 7, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.N.; Radner, S.; French, M.M.; Pinzon-Duarte, G.; Daly, G.H.; Burgeson, R.E.; Koch, M.; Brunken, W.J. The gamma3 chain of laminin is widely but differentially expressed in murine basement membranes: Expression and functional studies. Matrix Biol. 2012, 31, 120–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J.; Johnson, A.R. Differential effects of laminin and merosin on neurite outgrowth by developing retinal ganglion cells. J. Cell Sci. Suppl. 1991, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chintala, S.K.; Zhang, X.; Austin, J.S.; Fini, M.E. Deficiency in matrix metalloproteinase gelatinase B (MMP-9) protects against retinal ganglion cell death after optic nerve ligation. J. Biol. Chem. 2002, 277, 47461–47468. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Moss, S.E.; Alexander, R.A.; Ali, R.R.; Fitzke, F.W.; Cordeiro, M.F. Retinal ganglion cell apoptosis in glaucoma is related to intraocular pressure and IOP-induced effects on extracellular matrix. Investig. Ophthalmol. Vis. Sci. 2005, 46, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Pena, J.D.; Varela, H.J.; Ricard, C.S.; Hernandez, M.R. Enhanced tenascin expression associated with reactive astrocytes in human optic nerve heads with primary open angle glaucoma. Exp. Eye Res. 1999, 68, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Soto, I.; Howell, G.R. The complex role of neuroinflammation in glaucoma. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef]
- Goh, F.G.; Piccinini, A.M.; Krausgruber, T.; Udalova, I.A.; Midwood, K.S. Transcriptional regulation of the endogenous danger signal tenascin-C: A novel autocrine loop in inflammation. J. Immunol. 2010, 184, 2655–2662. [Google Scholar] [CrossRef] [Green Version]
- Midwood, K.; Sacre, S.; Piccinini, A.M.; Inglis, J.; Trebaul, A.; Chan, E.; Drexler, S.; Sofat, N.; Kashiwagi, M.; Orend, G.; et al. Tenascin-C is an endogenous activator of Toll-like receptor 4 that is essential for maintaining inflammation in arthritic joint disease. Nat. Med. 2009, 15, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Haage, V.; Elmadany, N.; Roll, L.; Faissner, A.; Gutmann, D.H.; Semtner, M.; Kettenmann, H. Tenascin C regulates multiple microglial functions involving TLR4 signaling and HDAC1. Brain Behav. Immun. 2019, 81, 470–483. [Google Scholar] [CrossRef]
- Wiemann, S.; Reinhard, J.; Reinehr, S.; Cibir, Z.; Joachim, S.C.; Faissner, A. Loss of the extracellular matrix molecule tenascin-C leads to absence of reactive gliosis and promotes anti-inflammatory cytokine expression in an autoimmune glaucoma mouse model. Front. Immun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Bu, W.Y.; Wang, T.H.; Ahmed, S.; Xiao, Z.C. Tenascin-R plays a role in neuroprotection via its distinct domains that coordinate to modulate the microglia function. J. Biol. Chem. 2005, 280, 8316–8323. [Google Scholar] [CrossRef] [Green Version]
- Becker, T.; Anliker, B.; Becker, C.G.; Taylor, J.; Schachner, M.; Meyer, R.L.; Bartsch, U. Tenascin-R inhibits regrowth of optic fibers in vitro and persists in the optic nerve of mice after injury. Glia 2000, 29, 330–346. [Google Scholar] [CrossRef]
- Susarla, B.T.; Laing, E.D.; Yu, P.; Katagiri, Y.; Geller, H.M.; Symes, A.J. Smad proteins differentially regulate transforming growth factor-beta-mediated induction of chondroitin sulfate proteoglycans. J. Neurochem. 2011, 119, 868–878. [Google Scholar] [CrossRef] [Green Version]
- Galtrey, C.M.; Fawcett, J.W. The role of chondroitin sulfate proteoglycans in regeneration and plasticity in the central nervous system. Brain Res. Rev. 2007, 54, 1–18. [Google Scholar] [CrossRef]
- Chen, L.F.; FitzGibbon, T.; He, J.R.; Yin, Z.Q. Localization and developmental expression patterns of CSPG-cs56 (aggrecan) in normal and dystrophic retinas in two rat strains. Exp. Neurol. 2012, 234, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Milev, P.; Fischer, D.; Haring, M.; Schulthess, T.; Margolis, R.K.; Chiquet-Ehrismann, R.; Margolis, R.U. The fibrinogen-like globe of tenascin-C mediates its interactions with neurocan and phosphacan/protein-tyrosine phosphatase-zeta/beta. J. Biol. Chem. 1997, 272, 15501–15509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnea, G.; Grumet, M.; Milev, P.; Silvennoinen, O.; Levy, J.B.; Sap, J.; Schlessinger, J. Receptor tyrosine phosphatase beta is expressed in the form of proteoglycan and binds to the extracellular matrix protein tenascin. J. Biol. Chem. 1994, 269, 14349–14352. [Google Scholar] [CrossRef]
- Zhang, X.; Cheng, M.; Chintala, S.K. Kainic acid-mediated upregulation of matrix metalloproteinase-9 promotes retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2374–2383. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Sainio, A.; Jarvelainen, H. Extracellular matrix-cell interactions: Focus on therapeutic applications. Cell. Signal. 2020, 66, 109487. [Google Scholar] [CrossRef]
- Ren, T.; van der Merwe, Y.; Steketee, M.B. Developing Extracellular Matrix Technology to Treat Retinal or Optic Nerve Injury (1,2,3). eNeuro 2015, 2. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene Name | Primer Sequence | GenBank Accession Number | Product Size (bp) |
|---|---|---|---|---|
| β-actin_for | Actb | ctaaggccaaccgtgaaaag | NM_007393.5 | 104 |
| β-actin_rev | accagaggcatacagggaca | |||
| Aggrecan_for | Acan | ccagcctacaccccagtg | NM_007424.3 | 66 |
| Aggrecan_rev | gagggtgggaagccatgt | |||
| Brevican_for | Bcan | tcaatgtctactgcttccgaga | NM_007529.2 | 72 |
| Brevican_rev | catctgaggctgggctagag | |||
| Cyclophilin D_for | Ppid | aaggatggcaaggattgaaa | NM_026352.4 | 105 |
| Cyclophilin D_rev | ctttaagcaattctgcctgga | |||
| Fibronectin_for | Fn1 | gccaccggagtctttactacc | NM_010233.2 | 61 |
| Fibronectin_rev | cctcggtgttgtaaggtgga | |||
| Laminin α4_for | Lama4 | tgtttgttggaggtgttcca | NM_010681.4 | 130 |
| Laminin α4_rev | ctgaccagggcagctttact | |||
| Lamininβ2_for | Lamb2 | tttcatttcacccacctcatt | NM_008483.3 | 77 |
| Lamininβ2_rev | aagtctgcagaacgctccac | |||
| Laminin γ3_for | Lamc3 | gctgacctcagaagcacaca | NM_011836.4 | 70 |
| Laminin γ3_rev | cacagtgctcagcccaga | |||
| RPTPβ/ζ CA_for | RPTPβ/ζ | gaatcctgcagagcttcctc | NM_011219.2 | 74 |
| RPTPβ/ζ CA_rev | gtagtatccataagcccagtcca | |||
| RPTPβ/ζ PTP1_for | ttgacggttccttcatgttg | NM_001311064.1 | 101 | |
| RPTPβ/ζ PTP1_rev | ttgacggttccttcatgttg | |||
| Tenascin-C_for | Tnc | cagggatagactgctctgagg | NM_001369211.1 | 90 |
| Tenascin-C_rev | cattgtcccatgccagattt | |||
| Tenascin-R_for | Tnr | gatggaagccgcaaagag | NM_022312.3 | 68 |
| Tenascin-R_rev | tctgacaggccctctagtcg |
| Primary Antibody | Dilution | Reference/Source | Secondary Antibody | Dilution | Source |
|---|---|---|---|---|---|
| Aggrecan | 1:250 | Millipore | Goat anti-rabbit Cy3 | 1:250 | Jackson ImmunoResearch Labs |
| Brevican | 1:300 | [46] | Goat anti-guinea pig Cy2 | 1:250 | Jackson ImmunoResearch Labs |
| Fibronectin | 1:300 | [47] | Goat anti-rabbit Cy3 | 1:250 | Jackson ImmunoResearch Labs |
| Laminin | 1:300 | [47] | Goat anti-rabbit Cy2 | 1:250 | Jackson ImmunoResearch Labs |
| Tenascin-C (KAF14 antibody) | 1:250 | [48] | Goat anti-rabbit Cy3 | 1:250 | Jackson ImmunoResearch Labs |
| Tenascin-R (23-14 antibody) | 1:100 | [49] | Goat anti-mouse Cy2 | 1:250 | Jackson ImmunoResearch Labs |
| Phosphacan/RPTPβ/ζ (473HD antibody) | 1:200 | [50] | Goat anti-rat Cy2 | 1:250 | Jackson ImmunoResearch Labs |
| RPTPβ/ζ-isoforms (KAF13 antibody) | 1:200 | [50] | Goat anti-rabbit Cy3 | 1:250 | Jackson ImmunoResearch Labs |
| Protein | Tissue | Background Subtraction | Lower Threshold | Upper Threshold |
|---|---|---|---|---|
| Aggrecan | Retina | 50 | 7.55 | 78.88 |
| Optic nerve | 50 | 13.84 | 80.10 | |
| Brevican | Retina | 20 | 5.21 | 35.24 |
| Optic nerve | 20 | 12.18 | 72.30 | |
| Fibronectin | Retina | 50 | 23.45 | 78.31 |
| Optic nerve | 50 | 10.90 | 78.85 | |
| Laminin | Retina | 30 | 11.25 | 75.65 |
| Optic nerve | 30 | 33.98 | 79.83 | |
| Tenascin-C | Retina | 50 | 25.33 | 80.11 |
| Optic nerve | 50 | 18.50 | 73.50 | |
| Tenascin-R | Retina | 20 | 20.00 | 80.00 |
| Optic nerve | 50 | 21.22 | 79.99 | |
| RPTPβ/ζ (473HD) | Retina | 50 | 8.77 | 79.22 |
| Optic nerve | 20 | 18.80 | 75.30 | |
| RPTPβ/ζ (KAF13) | Retina | 50 | 18.01 | 77.02 |
| Optic nerve | 50 | 22.53 | 80.00 |
| Primary Antibody | Molecular Weight | Dilution | Reference/Source | Secondary Antibody | Dilution | Source |
|---|---|---|---|---|---|---|
| Aggrecan | >100 kDa, >150 kDa | 1:1000 | Millipore | Goat anti-rabbit HRP | 1:10,000 | Jackson ImmunoResearch Labs |
| Brevican | ~50 kDa, >100 kDa | 1:1000 | [46] | Goat anti-guinea pig HRP | 1:5000 | Jackson ImmunoResearch Labs |
| Fibronectin | >250 kDa | 1:10,000 | [47] | Goat anti-rabbit HRP | 1:10,000 | Jackson ImmunoResearch Labs |
| Laminin | 200 kDa, 400 kDa | 1:10,000 | [47] | Goat anti-rabbit HRP | 1:5000 | Jackson ImmunoResearch Labs |
| Tenascin-C (KAF14 antibody) | ~250 kDa, >250 kDa | 1:5000 | [48] | Goat anti-rabbit HRP | 1:10,000 | Jackson ImmunoResearch Labs |
| Tenascin-R (23–14 antibody) | 160 kDa, 180 kDa | 1:1000 | [49] | Goat anti-mouse HRP | 1:5000 | Jackson ImmunoResearch Labs |
| Phosphacan/RPTPβ/ζ (473HD antibody) | >150 kDa | 1:100 | [50] | Goat anti-rat HRP | 1:5000 | Jackson ImmunoResearch Labs |
| RPTPβ/ζ-isoforms (KAF13 antibody) | >150 kDa | 1:5000 | [50] | Goat anti-rabbit HRP | 1:5000 | Jackson ImmunoResearch Labs |
| α-Tubulin | ~50 kDa | 1:20,000 | Sigma-Aldrich | Goat anti-mouse HRP | 1:10,000 | Jackson ImmunoResearch Labs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinhard, J.; Wiemann, S.; Hildebrandt, S.; Faissner, A. Extracellular Matrix Remodeling in the Retina and Optic Nerve of a Novel Glaucoma Mouse Model. Biology 2021, 10, 169. https://doi.org/10.3390/biology10030169
Reinhard J, Wiemann S, Hildebrandt S, Faissner A. Extracellular Matrix Remodeling in the Retina and Optic Nerve of a Novel Glaucoma Mouse Model. Biology. 2021; 10(3):169. https://doi.org/10.3390/biology10030169
Chicago/Turabian StyleReinhard, Jacqueline, Susanne Wiemann, Sebastian Hildebrandt, and Andreas Faissner. 2021. "Extracellular Matrix Remodeling in the Retina and Optic Nerve of a Novel Glaucoma Mouse Model" Biology 10, no. 3: 169. https://doi.org/10.3390/biology10030169
APA StyleReinhard, J., Wiemann, S., Hildebrandt, S., & Faissner, A. (2021). Extracellular Matrix Remodeling in the Retina and Optic Nerve of a Novel Glaucoma Mouse Model. Biology, 10(3), 169. https://doi.org/10.3390/biology10030169