
Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
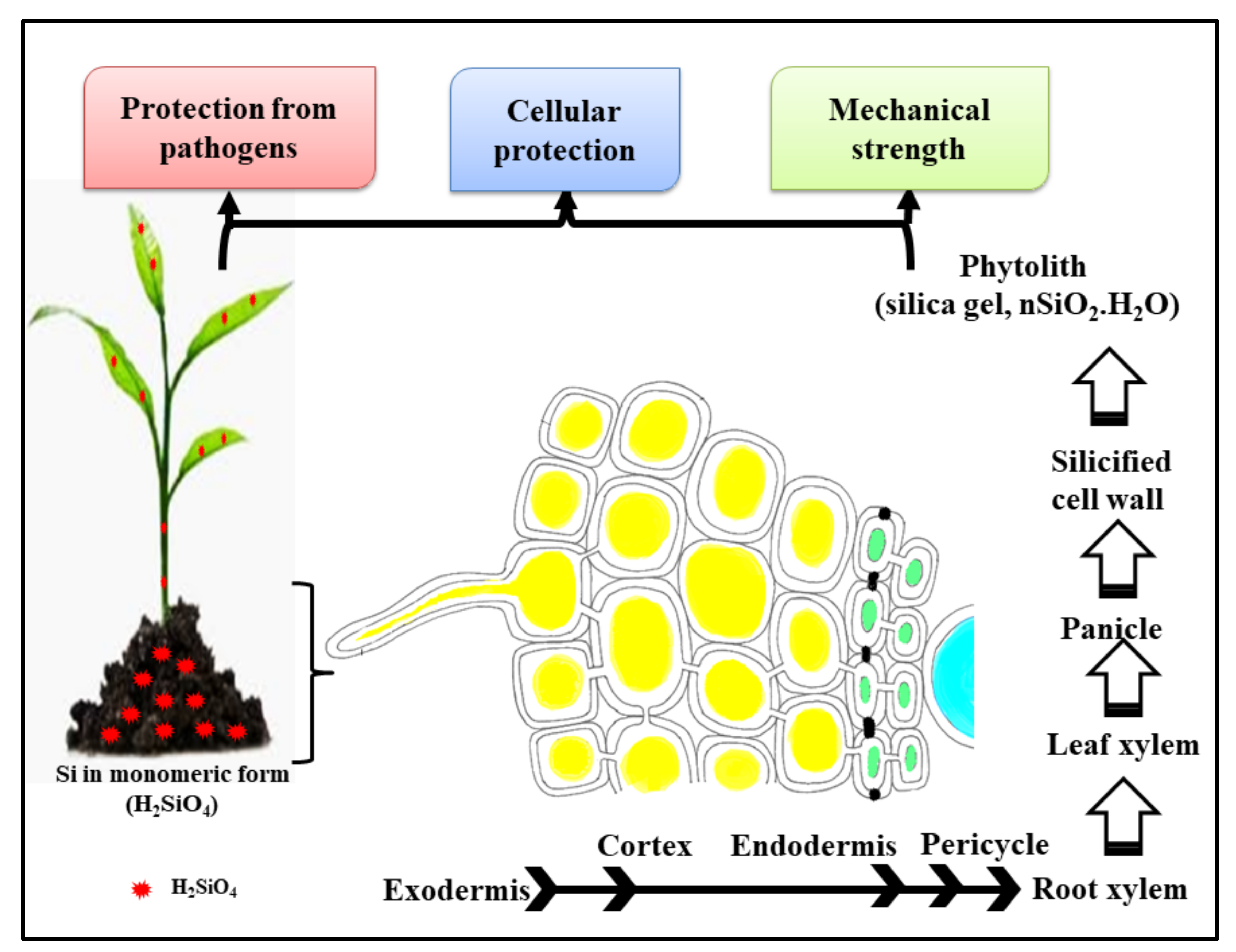
2. Source of Si and Si-NPs and Their Uptake
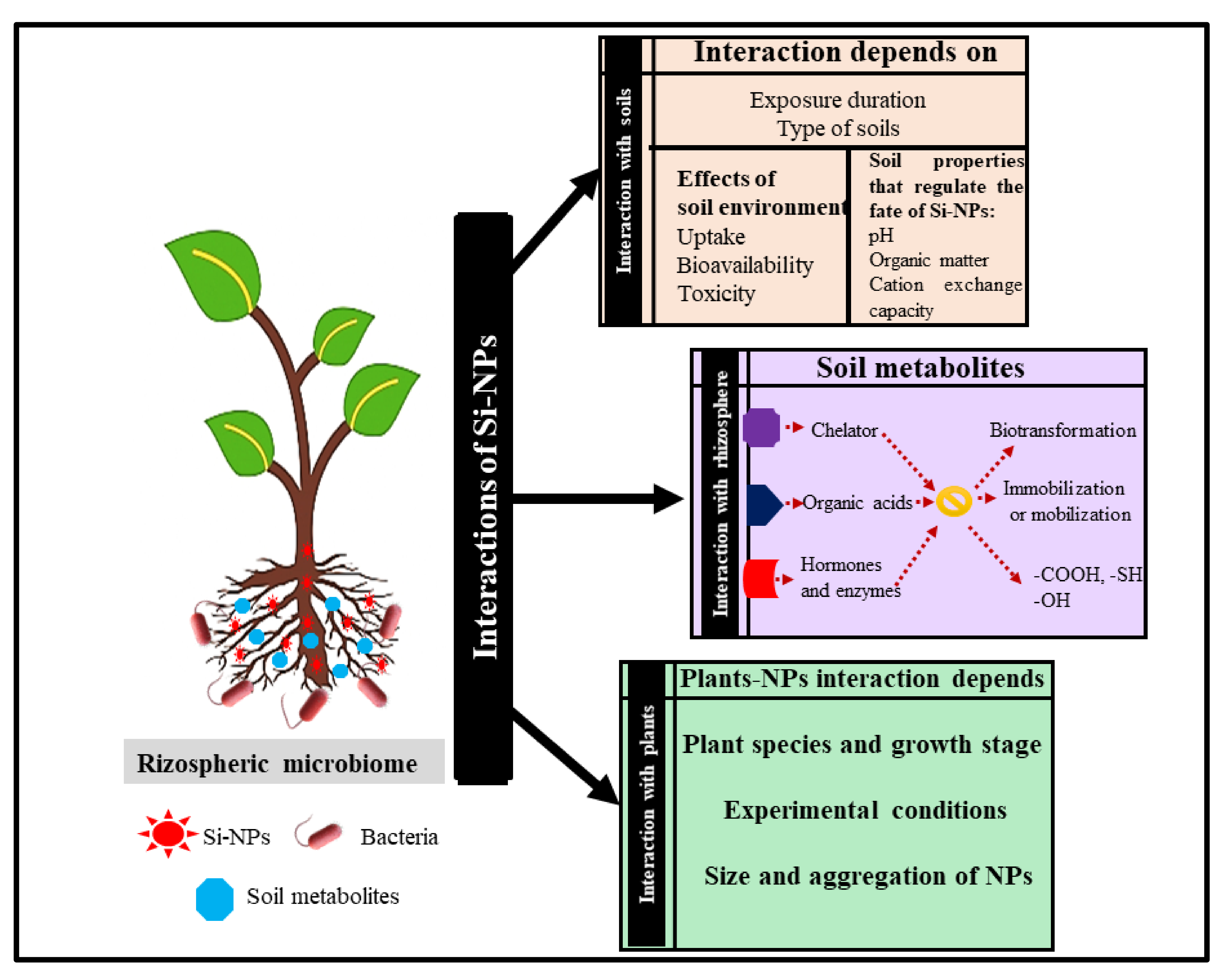
3. Impacts of Si-NPs on Soil Properties
4. Effects of Si-NPs on the Rhizospheric Microbiome
5. Effects of Si-NPs on Plant Growth and Development
6. Role of Si and Si-NPs in Abiotic Stress Tolerance in Plants
7. Role of Si and Si-NPs in Plant Biotic Stress Management
8. Molecular Mechanism of Si and Si-NP Uptake and Their Applications in Agriculture
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, J.F.; Yamaji, N. Silicon Uptake and Accumulation in Higher Plants. Trends Plant. Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, H.; Ahmed, K.B.M.; Sami, F.; Hayat, S. Silicon nanoparticles and plants: Current knowledge and future perspectives. In Sustainable Agriculture Reviews 41: Nanotechnology for Plant Growth and Development; Hayat, S., Pichtel, J., Faizan, M., Fariduddin, Q., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 129–142. [Google Scholar]
- Verma, K.K.; Song, X.-P.; Li, D.-M.; Singh, M.; Rajput, V.D.; Malviya, M.K.; Minkina, T.; Singh, R.K.; Singh, P.; Li, Y.-R. Interactive Role of Silicon and Plant–Rhizobacteria Mitigating Abiotic Stresses: A New Approach for Sustainable Agriculture and Climate Change. Plants 2020, 9, 1055. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.K.; Song, X.-P.; Verma, C.L.; Chen, Z.-L.; Rajput, V.D.; Wu, K.-C.; Liao, F.; Chen, G.-L.; Li, Y.-R. Functional Relationship Between Photosynthetic Leaf Gas Exchange in Response to Silicon Application and Water Stress Mitigation in Sugarcane. Biol. Res. 2021, 54, 15. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wong, J.W.; Wei, L. Silicon-mediated Enhancement of Cadmium Tolerance in Maize (Zea mays L.) Grown in Cadmium Contaminated Soil. Chemosphere 2005, 58, 475–483. [Google Scholar] [CrossRef]
- Liu, P.; Yin, L.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S.; Tanaka, K. Enhanced Root Hydraulic Conductance by Aquaporin Regulation Accounts for Silicon Alleviated Salt-Induced Osmotic Stress in Sorghum Bicolor L. Environ. Exp. Bot. 2015, 111, 42–51. [Google Scholar] [CrossRef]
- Bao-Shan, L.; Shao-Qi, D.; Chun-Hui, L.; Li-Jun, F.; Shu-Chun, Q.; Min, Y. Effect of TMS (nanostructured silicon dioxide) on growth of Changbai larch seedlings. J. For. Res. 2004, 15, 138–140. [Google Scholar] [CrossRef]
- Yuvakkumar, R.; Elango, V.; Rajendran, V.; Kannan, N.S.; Prabu, P. Influence of Nanosilica Powder on the Growth of Maize Crop (Zea Mays L.). Int. J. Green Nanotechnol. 2011, 3, 180–190. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Nano-silicon alters antioxidant activities of soybean seedlings under salt toxicity. Protoplasma 2018, 255, 953–962. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Foliar Application of Silica Nanoparticles on the Phytochemical Responses of Maize (Zea mays L.) and Its Toxicological Behavior. Synth. React. Inorg. Met. Org. Nano Met. Chem. 2014, 44, 1128–1131. [Google Scholar] [CrossRef]
- Mukarram, M.; Khan, M.M.A.; Corpas, F.J. Silicon nanoparticles elicit an increase in lemongrass (Cymbopogon flexuosus (Steud.) Wats) agronomic parameters with a higher essential oil yield. J. Hazard. Mater. 2021, 412, 125254. [Google Scholar] [CrossRef]
- Asgari, F.; Majd, A.; Jonoubi, P.; Najafi, F. Effects of silicon nanoparticles on molecular, chemical, structural and ultrastructural characteristics of oat (Avena sativa L.). Plant Physiol. Biochem. 2018, 127, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Suciaty, T.; Purnomo, D.; Sakya, A.T.; Supriyadi. The effect of nano-silica fertilizer concentration and rice hull ash doses on soybean (Glycine max (L.) Merrill) growth and yield. IOP Conf. Ser. Earth Environ. Sci. 2018, 129, 012009. [Google Scholar] [CrossRef] [Green Version]
- Janmohammadi, M.; Sabaghnia, N. Effect of Pre-Sowing Seed Treatments with Silicon Nanoparticles on Germinability of Sunflower (Helianthus Annuus). Botanica 2015, 21, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Azimi, R.; Borzelabad, M.J.; Feizi, H.; Azimi, A. Interaction of SiO Nanoparticles with Seed Prechilling on Germination and Early Seedling Growth of Tall Wheatgrass (Agropyron Elongatum L.). Pol. J. Chem. Technol. 2014, 16, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Lin, B.; Guo, D.-J.; Singh, M.; Rajput, V.D.; Singh, R.K.; Singh, P.; Sharma, A.; Malviya, M.K.; et al. Silicon Induced Drought Tolerance in Crop Plants: Physiological Adaptation Strategies. Silicon 2021. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Mushinskiy, A.A.; Aminova, E.V.; Korotkova, A.M. Evaluation of tolerance of tubers Solanum tuberosum to silica nanoparticles. Environ. Sci. Pollut. Res. Int. 2018, 25, 34559–34569. [Google Scholar] [CrossRef] [PubMed]
- Sahebi, M.; Hanafi, M.M.; Siti Nor Akmar, A.; Rafii, M.Y.; Azizi, P.; Tengoua, F.F.; Nurul Mayzaitul Azwa, J.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. Biomed. Res. Int. 2015, 2015, 396010. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Hashmi, A.; Khan, M.R.; Parveen, A. Management of bacteria Pectobacterium carotovorum, Xanthomonas campestris pv. carotae, and fungi Rhizoctonia solani, Fusarium solani and Alternaria dauci with silicon dioxide nanoparticles on carrot. Int. J. Veg. Sci. 2020, 26, 547–557. [Google Scholar] [CrossRef]
- Gattullo, C.E.; Allegretta, I.; Medici, L.; Fijan, R.; Pii, Y.; Cesco, S.; Mimmo, T.; Terzano, R. Silicon dynamics in the rhizosphere: Connections with iron mobilization. J. Plant Nutr. Soil Sci. 2016, 179, 409–417. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.Y.; Su, Y.H.; McGrath, S.P.; Zhao, F.J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Song, Y.; Long, J.; Wang, R.; Baerson, S.R.; Pan, Z.; Zhu-Salzman, K.; Xie, J.; Cai, K.; Luo, S.; et al. Priming of jasmonate-mediated antiherbivore defense responses in rice by silicon. Proc. Natl. Acad. Sci. USA 2013, 110, E3631–E3639. [Google Scholar] [CrossRef] [Green Version]
- Carré-Missio, V.; Rodrigues, F.A.; Schurt, D.A.; Resende, R.S.; Souza, N.F.A.; Rezende, D.C.; Moreira, W.R.; Zambolim, L. Effect of foliar-applied potassium silicate on coffee leaf infection by Hemileia vastatrix. Ann. Appl. Biol. 2014, 164, 396–403. [Google Scholar] [CrossRef]
- Karunakaran, G.; Suriyaprabha, R.; Manivasakan, P.; Yuvakkumar, R.; Rajendran, V.; Prabu, P.; Kannan, N. Effect of nanosilica and silicon sources on plant growth promoting rhizobacteria, soil nutrients and maize seed germination. IET Nanobiotechnol. 2013, 7, 70–77. [Google Scholar] [CrossRef]
- Khan, M.R.; Siddiqui, Z.A. Use of silicon dioxide nanoparticles for the management of Meloidogyne incognita, Pectobacterium betavasculorum and Rhizoctonia solani disease complex of beetroot (Beta vulgaris L.). Sci. Hortic. 2020, 265, 109211. [Google Scholar] [CrossRef]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of Silicon on Plant–Pathogen Interactions. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Ghareeb, H.; Bozsó, Z.; Ott, P.G.; Repenning, C.; Stahl, F.; Wydra, K. Transcriptome of silicon-induced resistance against Ralstonia solanacearum in the silicon non-accumulator tomato implicates priming effect. Physiol. Mol. Plant Pathol. 2011, 75, 83–89. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, M.; Wang, L.; Lin, W.; Fan, X.; Cai, K. Proteomic characterization of silicon-mediated resistance against Ralstonia solanacearum in tomato. Plant Soil 2015, 387, 425–440. [Google Scholar] [CrossRef]
- Brunings, A.M.; Datnoff, L.E.; Ma, J.F.; Mitani, N.; Nagamura, Y.; Rathinasabapathi, B.; Kirst, M. Differential gene expression of rice in response to silicon and rice blast fungus Magnaporthe oryzae. Ann. Appl. Biol. 2009, 155, 161–170. [Google Scholar] [CrossRef]
- Chain, F.; Côté-Beaulieu, C.; Belzile, F.; Menzies, J.G.; Bélanger, R.R. A Comprehensive Transcriptomic Analysis of the Effect of Silicon on Wheat Plants Under Control and Pathogen Stress Conditions. Mol. Plant Microbe Interact. 2009, 22, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayfeh, M.H.; Mitas, L. Chapter one—Silicon nanoparticles: New Photonic and electronic material at the transition between solid and molecule. In Nanosilicon; Kumar, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 1–78. [Google Scholar]
- Kaur, H.; Greger, M. A Review on Si Uptake and Transport System. Plants 2019, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- El-Ramady, H.; Alshaal, T.; Elhawat, N.; El-Nahrawy, E.; Omara, A.E.-D.; El-Nahrawy, S.; Elsakhawy, T.; Ghazi, A.; Abdalla, N.; Fári, M. Biological aspects of selenium and silicon nanoparticles in the terrestrial environments. In Phytoremediation: Management of Environmental Contaminants; Ansari, A.A., Gill, S.S., Gill, R., Lanza, G., Newman, L., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 6, pp. 235–264. [Google Scholar]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Ma, J.; Miyake, Y.; Takahashi, E. Chapter 2 Silicon as a beneficial element for crop plants. Stud. Plant Sci. 2001, 8, 17–39. [Google Scholar]
- Jadhao, K.R.; Bansal, A.; Rout, G.R. Silicon amendment induces synergistic plant defense mechanism against pink stem borer (Sesamia inferens Walker.) in finger millet (Eleusine coracana Gaertn.). Sci. Rep. 2020, 10, 4229. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic Difference in Silicon Uptake and Expression of Silicon Transporter Genes in Rice. Plant. Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Yamaji, N. Functions and transport of silicon in plants. Cell. Mol. Life Sci. CMLS 2008, 65, 3049–3057. [Google Scholar] [CrossRef]
- Yamaji, N.; Sakurai, G.; Mitani-Ueno, N.; Ma, J.F. Orchestration of three transporters and distinct vascular structures in node for intervascular transfer of silicon in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11401–11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of maize silicon influx transporters. Plant. Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Foy, C.D. Soil chemical factors limiting plant root growth. In Limitations to Plant Root Growth; Hatfield, J.L., Stewart, B.A., Eds.; Springer: New York, NY, USA, 1992; pp. 97–149. [Google Scholar]
- Snyder, G.H.; Jones, D.B.; Gascho, G.J. Silicon Fertilization of Rice on Everglades Histosols. Soil Sci. Soc. Am. J. 1986, 50, 1259–1263. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Deren, C.W.; Snyder, G.H. Silicon fertilization for disease management of rice in Florida. Crop. Prot. 1997, 16, 525–531. [Google Scholar] [CrossRef]
- Sauer, D.; Burghardt, W. The occurrence and distribution of various forms of silica and zeolites in soils developed from wastes of iron production. CATENA 2006, 65, 247–257. [Google Scholar] [CrossRef]
- Farooq, M.A.; Dietz, K.-J. Silicon as Versatile Player in Plant and Human Biology: Overlooked and Poorly Understood. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Savvas, D.; Ntatsi, G. Biostimulant activity of silicon in horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Zargar, S.M.; Mahajan, R.; Bhat, J.A.; Nazir, M.; Deshmukh, R. Role of silicon in plant stress tolerance: Opportunities to achieve a sustainable cropping system. 3 Biotech 2019, 9, 73. [Google Scholar] [CrossRef]
- Buzea, C.; Pacheco, I. Nanomaterials and their classification. In EMR/ESR/EPR Spectroscopy for Characterization of Nanomaterials; Shukla, A.K., Ed.; Springer India: New Delhi, India, 2017; pp. 3–45. [Google Scholar]
- Jilling, A.; Keiluweit, M.; Contosta, A.R.; Frey, S.; Schimel, J.; Schnecker, J.; Smith, R.G.; Tiemann, L.; Grandy, A.S. Minerals in the rhizosphere: Overlooked mediators of soil nitrogen availability to plants and microbes. Biogeochemistry 2018, 139, 103–122. [Google Scholar] [CrossRef] [Green Version]
- Richard Drees, L.; Wilding, L.P.; Smeck, N.E.; Senkayi, A.L. Silica in soils: Quartz and disordered silica polymorphs. In Minerals in Soil Environments; Soil Science Society of America, Inc.: Madison, WI, USA, 1989; pp. 913–974. [Google Scholar]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Fraysse, F.; Pokrovsky, O.S.; Schott, J.; Meunier, J.-D. Surface properties, solubility and dissolution kinetics of bamboo phytoliths. Geochim. Cosmochim. Acta 2006, 70, 1939–1951. [Google Scholar] [CrossRef]
- Tubaña, B.S.; Heckman, J.R. Silicon in soils and plants. In Silicon and Plant Diseases; Rodrigues, F.A., Datnoff, L.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 7–51. [Google Scholar]
- Afshari, M.; Pazoki, A.; Sadeghipour, O. Foliar-applied Silicon and its Nanoparticles Stimulate Physio-chemical Changes to Improve Growth, Yield and Active Constituents of Coriander (Coriandrum Sativum L.) Essential Oil Under Different Irrigation Regimes. Silicon 2021. [Google Scholar] [CrossRef]
- Ahmadian, K.; Jalilian, J.; Pirzad, A. Nano-fertilizers improved drought tolerance in wheat under deficit irrigation. Agric. Water Manag. 2021, 244, 106544. [Google Scholar] [CrossRef]
- Banerjee, A.; Singh, A.; Sudarshan, M.; Roychoudhury, A. Silicon nanoparticle-pulsing mitigates fluoride stress in rice by fine-tuning the ionomic and metabolomic balance and refining agronomic traits. Chemosphere 2021, 262, 127826. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, R.V.; Saharan, V.; Kumari, S.; Chandra Choudhary, R.; Pal, A.; Sharma, S.S.; Rakshit, S.; Raliya, R.; Biswas, P. Chitosan-silicon nanofertilizer to enhance plant growth and yield in maize (Zea mays L.). Plant Physiol. Biochem. 2021, 159, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.; Wang, L.; Feng, Q.; Niu, L.; Zhao, Z.; Wang, P.; Song, C.; Li, X.; Zhang, Z. Thiol-functionalized nano-silica for in-situ remediation of Pb, Cd, Cu contaminated soils and improving soil environment. Environ. Pollut. Barking Essex 1987 2021, 280, 116879. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Desoky, E.-S.M.; Saad, A.M.; Eid, R.S.M.; Selem, E.; Elrys, A.S. Biological silicon nanoparticles improve Phaseolus vulgaris L. yield and minimize its contaminant contents on a heavy metals-contaminated saline soil. J. Environ. Sci. 2021, 106, 1–14. [Google Scholar] [CrossRef]
- Farajzadeh Memari-Tabrizi, E.; Yousefpour-Dokhanieh, A.; Babashpour-Asl, M. Foliar-applied silicon nanoparticles mitigate cadmium stress through physio-chemical changes to improve growth, antioxidant capacity, and essential oil profile of summer savory (Satureja hortensis L.). Plant Physiol. Biochem. 2021, 165, 71–79. [Google Scholar] [CrossRef]
- Valizadeh-rad, K.; Motesharezadeh, B.; Alikhani, H.A.; Jalali, M. Direct and Residual Effects of Water Deficit Stress, Different Sources of Silicon and Plant-Growth Promoting Bacteria on Silicon Fractions in the Soil. Silicon 2021. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, J.; Dou, F.; Sun, W.; Ma, X. Simultaneous mitigation of arsenic and cadmium accumulation in rice (Oryza sativa L.) seedlings by silicon oxide nanoparticles under different water management schemes. Paddy Water Environ. 2021. [Google Scholar] [CrossRef]
- Asadpour, S.; Madani, H.; Mohammadi, G.N.; Heravan, I.M.; Abad, H.H.S. Improving Maize Yield with Advancing Planting Time and Nano-Silicon Foliar Spray Alone or Combined with Zinc. Silicon 2020. [Google Scholar] [CrossRef]
- Cao, P.; Qiu, K.; Zou, X.; Lian, M.; Liu, P.; Niu, L.; Yu, L.; Li, X.; Zhang, Z. Mercapto propyltrimethoxysilane- and ferrous sulfate-modified nano-silica for immobilization of lead and cadmium as well as arsenic in heavy metal-contaminated soil. Environ. Pollut. Barking Essex 1987 2020, 266, 115152. [Google Scholar] [CrossRef]
- Esmaili, S.; Tavallali, V.; Amiri, B. Nano-Silicon Complexes Enhance Growth, Yield, Water Relations and Mineral Composition in Tanacetum parthenium under Water Deficit Stress. Silicon 2020. [Google Scholar] [CrossRef]
- Namjoyan, S. Nano-silicon protects sugar beet plants against water deficit stress by improving the antioxidant systems and compatible solutes. Acta Physiol. Plant 2020, 42, 157. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Zhan, W.; Zheng, K.; Lian, M.; Zhang, C.; Ruan, X.; Li, T. Long-term stabilization of Cd in agricultural soil using mercapto-functionalized nano-silica (MPTS/nano-silica): A three-year field study. Ecotoxicol. Environ. Saf. 2020, 197, 110600. [Google Scholar] [CrossRef]
- Zuo, R.; Liu, H.; Xi, Y.; Gu, Y.; Ren, D.; Yuan, X.; Huang, Y. Nano-SiO2 combined with a surfactant enhanced phenanthrene phytoremediation by Erigeron annuus (L.) Pers. Environ. Sci. Pollut. Res. Int. 2020, 27, 20538–20544. [Google Scholar] [CrossRef]
- Oliveira, E.M.D.; Garcia-Rojas, E.E.; Valadão, I.C.R.P.; Araújo, A.D.S.F.; Castro, J.A.D. Effects of the silica nanoparticles (NPSiO2) on the stabilization and transport of hazardous nanoparticle suspensions into landfill soil columns. REM Int. Eng. J. 2017, 70, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Santos, L.F.; Olivares, F.L. Plant microbiome structure and benefits for sustainable agriculture. Curr. Plant Biol. 2021, 26, 100198. [Google Scholar] [CrossRef]
- Brolsma, K.M.; Vonk, J.A.; Mommer, L.; Van Ruijven, J.; Hoffland, E.; De Goede, R.G.M. Microbial catabolic diversity in and beyond the rhizosphere of plant species and plant genotypes. Pedobiologia 2017, 61, 43–49. [Google Scholar] [CrossRef]
- McGee, C.F.; Storey, S.; Clipson, N.; Doyle, E. Soil microbial community responses to contamination with silver, aluminium oxide and silicon dioxide nanoparticles. Ecotoxicology 2017, 26, 449–458. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Singh, M.; Joshi, D.; Singh, J.; Suyal, D.C.; Kumar, A.; Rajput, V.D.; et al. Beneficial microbiomes for bioremediation of diverse contaminated environments for environmental sustainability: Present status and future challenges. Environ. Sci. Pollut. Res. 2021, 28, 24917–24939. [Google Scholar] [CrossRef] [PubMed]
- Barea, J.M.; Pozo, M.J.; Azcón, R.; Azcón-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef] [Green Version]
- Theng, B.K.G.; Yuan, G. Nanoparticles in the Soil Environment. Elements 2008, 4, 395–399. [Google Scholar] [CrossRef]
- Tian, L.; Shen, J.; Sun, G.; Wang, B.; Ji, R.; Zhao, L. Foliar Application of SiO2 Nanoparticles Alters Soil Metabolite Profiles and Microbial Community Composition in the Pakchoi (Brassica chinensis L.) Rhizosphere Grown in Contaminated Mine Soil. Environ. Sci. Technol. 2020, 54, 13137–13146. [Google Scholar] [CrossRef]
- Liu, X.; Li, Q.; Li, Y.; Guan, G.; Chen, S. Paenibacillus strains with nitrogen fixation and multiple beneficial properties for promoting plant growth. PeerJ 2019, 7, e7445. [Google Scholar] [CrossRef] [Green Version]
- Withers, E.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Use of untargeted metabolomics for assessing soil quality and microbial function. Soil Biol. Biochem. 2020, 143, 107758. [Google Scholar] [CrossRef]
- Kumar, N.; Shah, V.; Walker, V.K. Influence of a nanoparticle mixture on an arctic soil community. Environ. Toxicol. Chem. 2012, 31, 131–135. [Google Scholar] [CrossRef]
- Khan, M.; Khan, A.U.; Hasan, M.A.; Yadav, K.K.; Pinto, M.M.C.; Malik, N.; Yadav, V.K.; Khan, A.H.; Islam, S.; Sharma, G.K. Agro-Nanotechnology as an Emerging Field: A Novel Sustainable Approach for Improving Plant Growth by Reducing Biotic Stress. Appl. Sci. 2021, 11. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Chourasiya, V.; Nehra, A.; Shukla, P.; Singh, K.; Singh, P. Impact of Mesoporous Nano-Silica (SiO2) on Seed Germination and Seedling Growth of Wheat, Pea and Mustard Seed. J. Nanosci. Nanotechnol. 2021. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Zeng, Y.; Guo, D.-J.; Singh, M.; Rajput, V.D.; Malviya, M.K.; Wei, K.-J.; Sharma, A.; Li, D.-P.; et al. Foliar application of silicon boosts growth, photosynthetic leaf gas exchange, antioxidative response and resistance to limited water irrigation in sugarcane (Saccharum officinarum L.). Plant Physiol. Biochem. 2021, 166, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Kumari, A.; Harish; Singh, V.K.; Verma, K.K.; Mandzhieva, S.; Sushkova, S.; Srivastava, S.; Keswani, C. Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles. Plants 2021, 10. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Sunoj, V.S.J.; Wen, Y.; Zhu, J.J.; Muralidharan, G.; Cao, K.F. Foliar application of nanoparticles mitigates the chilling effect on photosynthesis and photoprotection in sugarcane. Plant Physiol. Biochem. 2020, 149, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, N.; Liang, X.; Bai, X.; Zheng, L.; Zhao, J.; Li, Y.-F.; Zhang, Z.; Gao, Y. Silica nanoparticles alleviate mercury toxicity via immobilization and inactivation of Hg(ii) in soybean (Glycine max). Environ. Sci. Nano 2020, 7, 1807–1817. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Mohammadi, H.; Kariman, K. Nanosilicon-based recovery of barley (Hordeum vulgare) plants subjected to drought stress. Environ. Sci. Nano 2020, 7, 443–461. [Google Scholar] [CrossRef]
- Cui, J.; Liu, T.; Li, F.; Yi, J.; Liu, C.; Yu, H. Silica nanoparticles alleviate cadmium toxicity in rice cells: Mechanisms and size effects. Environ. Pollut. Barking Essex 1987 2017, 228, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Nazaralian, S.; Majd, A.; Irian, S.; Najafi, F.; Ghahremaninejad, F.; Landberg, T.; Greger, M. Comparison of silicon nanoparticles and silicate treatments in fenugreek. Plant Physiol. Biochem. 2017, 115, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef]
- Sun, D.; Hussain, H.I.; Yi, Z.; Rookes, J.E.; Kong, L.; Cahill, D.M. Mesoporous silica nanoparticles enhance seedling growth and photosynthesis in wheat and lupin. Chemosphere 2016, 152, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci. Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-Ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon dioxide nanoparticles ameliorate the phytotoxic hazards of aluminum in maize grown on acidic soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.H.; Qiu, H.; Tian, T.; Zhan, S.S.; Deng, T.H.; Chaney, R.L.; Wang, S.Z.; Tang, Y.T.; Morel, J.L.; Qiu, R.L. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil. Chemosphere 2011, 83, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, Y.; Jin, Q.; Li, F. Silica nanoparticles inhibit arsenic uptake into rice suspension cells via improving pectin synthesis and the mechanical force of the cell wall. Environ. Sci. Nano 2020, 7, 162–171. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Khlestkina, E.K. Anthocyanins Participate in the Protection of Wheat Seedlings against Cadmium Stress. Cereal Res. Commun. 2018, 46, 242–252. [Google Scholar] [CrossRef]
- Hayat, S.; Hasan, S.A.; Hayat, Q.; Ahmad, A. Brassinosteroids protect Lycopersicon esculentum from cadmium toxicity applied as shotgun approach. Protoplasma 2010, 239, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Hussain, B.; Lin, Q.; Hamid, Y.; Sanaullah, M.; Di, L.; Hashmi, M.; Khan, M.B.; He, Z.; Yang, X. Foliage application of selenium and silicon nanoparticles alleviates Cd and Pb toxicity in rice (Oryza sativa L.). Sci. Total Environ. 2020, 712, 136497. [Google Scholar] [CrossRef] [PubMed]
- Merwad, A.-R.M.A.; Desoky, E.-S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Rai, P.; Guerriero, G.; Sharma, S.; Corpas, F.J.; Singh, V.P. Silicon induces adventitious root formation in rice under arsenate stress with involvement of nitric oxide and indole-3-acetic acid. J. Exp. Bot. 2021, 72, 4457–4471. [Google Scholar] [CrossRef] [PubMed]
- Burachevskaya, M.; Minkina, T.; Fedorenko, A.; Fedorenko, G.; Chernikova, N.; Rajput, V.D.; Mandzhieva, S.; Bauer, T. Accumulation, translocation, and toxicity of arsenic in barley grown in contaminated soil. Plant Soil 2021. [Google Scholar] [CrossRef]
- Liu, C.; Wei, L.; Zhang, S.; Xu, X.; Li, F. Effects of nanoscale silica sol foliar application on arsenic uptake, distribution and oxidative damage defense in rice (Oryza sativa L.) under arsenic stress. RSC Adv. 2014, 4, 57227–57234. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Daoliang, Y. Chromium Bioaccumulation and Its Impacts on Plants: An Overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Prabu, P.; Rajendran, V.; Kannan, N. Growth and physiological responses of maize (Zea mays L.) to porous silica nanoparticles in soil. J. Nanopart. Res. 2012, 14, 1294. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Tian, D.-D.; Singh, M.; Verma, C.L.; Rajput, V.D.; Singh, R.K.; Sharma, A.; Singh, P.; Malviya, M.K.; et al. Investigation of Defensive Role of Silicon during Drought Stress Induced by Irrigation Capacity in Sugarcane: Physiological and Biochemical Characteristics. ACS Omega 2021. [Google Scholar] [CrossRef]
- Singhal, R.K.; Jatav, H.S.; Aftab, T.; Pandey, S.; Mishra, U.N.; Chauhan, J.; Chand, S.; Indu; Saha, D.; Dadarwal, B.K.; et al. Roles of Nitric Oxide in Conferring Multiple Abiotic Stress Tolerance in Plants and Crosstalk with Other Plant Growth Regulators. J. Plant. Growth Regul. 2021. [Google Scholar] [CrossRef]
- Matysik, J.; Alia, N.; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Iyer, S.; Caplan, A. Products of Proline Catabolism Can Induce Osmotically Regulated Genes in Rice1. Plant Physiol. 1998, 116, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Almutairi, Z.M. Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (’Solanum lycopersicum’ L.) seedlings under salt stress. South. Cross J. 2016, 9, 106–114. [Google Scholar]
- Wang, Y.; Stass, A.; Horst, W.J. Apoplastic binding of aluminum is involved in silicon-induced amelioration of aluminum toxicity in maize. Plant Physiol. 2004, 136, 3762–3770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, R.; Fretté, X.; Jensen, B.; Shetty, N.P.; Jensen, J.D.; Jørgensen, H.J.L.; Newman, M.-A.; Christensen, L.P. Silicon-induced changes in antifungal phenolic acids, flavonoids, and key phenylpropanoid pathway genes during the interaction between miniature roses and the biotrophic pathogen Podosphaera pannosa. Plant Physiol. 2011, 157, 2194–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mushtaq, A.; Jamil, N.; Rizwan, S.; Mandokhel, F.; Riaz, M.; Hornyak, G.L.; Najam Malghani, M.; Naeem Shahwani, M. Engineered Silica Nanoparticles and silica nanoparticles containing Controlled Release Fertilizer for drought and saline areas. IOP Conf. Ser. Mater. Sci. Eng. 2018, 414, 012029. [Google Scholar] [CrossRef]
- Faizan, M.; Rajput, V.D.; Al-Khuraif, A.A.; Arshad, M.; Minkina, T.; Sushkova, S.; Yu, F. Effect of Foliar Fertigation of Chitosan Nanoparticles on Cadmium Accumulation and Toxicity in Solanum lycopersicum. Biology 2021, 10, 666. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, D. Silicon-induced cell wall fortification of rice leaves: A possible cellular mechanism of enhanced host resistance to blast. Phytopathology 2002, 92, 1095–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takó, M.; Kerekes, E.B.; Zambrano, C.; Kotogán, A.; Papp, T.; Krisch, J.; Vágvölgyi, C. Plant Phenolics and Phenolic-Enriched Extracts as Antimicrobial Agents against Food-Contaminating Microorganisms. Antioxidants 2020, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G.; Hausman, J.-F.; Legay, S. Silicon and the Plant Extracellular Matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, F.A.; Benhamou, N.; Datnoff, L.E.; Jones, J.B.; Bélanger, R.R. Ultrastructural and cytochemical aspects of silicon-mediated rice blast resistance. Phytopathology 2003, 93, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebold, K.W.; Datnoff, L.E.; Correa-Victoria, F.J.; Kucharek, T.A.; Snyder, G.H. Effects of Silicon and Fungicides on the Control of Leaf and Neck Blast in Upland Rice. Plant Dis. 2004, 88, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Hayasaka, T.; Fujii, H.; Ishiguro, K. The Role of Silicon in Preventing Appressorial Penetration by the Rice Blast Fungus. Phytopathology 2008, 98, 1038–1044. [Google Scholar] [CrossRef] [Green Version]
- Menzies, J.G.; Ehret, D.L.; Glass, A.D.M.; Samuels, A.L. The influence of silicon on cytological interactions between Sphaerotheca fuliginea and Cucumis sativus. Physiol. Mol. Plant Pathol. 1991, 39, 403–414. [Google Scholar] [CrossRef]
- Ponmurugan, P.; Manjukarunambika, K.; Elango, V.; Gnanamangai, B.M. Antifungal activity of biosynthesised copper nanoparticles evaluated against red root-rot disease in tea plants. J. Exp. Nanosci. 2016, 11, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Elmer, W.; Shen, Y.; Zuverza-Mena, N.; Ma, C.; Botella, P.; White, J.C.; Haynes, C.L. Silica Nanoparticle Dissolution Rate Controls the Suppression of Fusarium Wilt of Watermelon (Citrullus lanatus). Environ. Sci. Technol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Siddiqui, Z.A. Effect of silicon dioxide nanoparticles and Rhizobium leguminosarum alone and in combination on the growth and bacterial blight disease complex of pea caused by Meloidogyne incognita and Pseudomonas syringae pv. pisi. Arch. Phytopathol. Plant Prot. 2020, 1–17. [Google Scholar] [CrossRef]
- Cui, J.; Liang, Y.; Yang, D.; Liu, Y. Facile fabrication of rice husk based silicon dioxide nanospheres loaded with silver nanoparticles as a rice antibacterial agent. Sci. Rep. 2016, 6, 21423. [Google Scholar] [CrossRef] [Green Version]
- Ramouthar, P.V.; Caldwell, P.M.; McFarlane, S.A. Effect of silicon on the severity of brown rust of sugarcane in South Africa. Eur. J. Plant Pathol. 2016, 145, 53–60. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Kavitha, K.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Application of silica nanoparticles in maize to enhance fungal resistance. IET Nanobiotechnol. 2014, 8, 133–137. [Google Scholar] [CrossRef]
- Resende, R.S.; Rodrigues, F.A.; Gomes, R.J.; Nascimento, K.J.T. Microscopic and biochemical aspects of sorghum resistance to anthracnose mediated by silicon. Ann. Appl. Biol. 2013, 163, 114–123. [Google Scholar] [CrossRef]
- Li, Y.C.; Bi, Y.; Ge, Y.H.; Sun, X.J.; Wang, Y. Antifungal activity of sodium silicate on Fusarium sulphureum and its effect on dry rot of potato tubers. J. Food Sci. 2009, 74, M213–M218. [Google Scholar] [CrossRef] [PubMed]
- Deepak, S.; Manjunath, G.; Manjula, S.; Niranjan-Raj, S.; Geetha, N.P.; Shetty, H.S. Involvement of silicon in pearl millet resistance to downy mildew disease and its interplay with cell wall proline/hydroxyproline-rich glycoproteins. Australas. Plant Pathol. 2008, 37, 498–504. [Google Scholar] [CrossRef]
- Mandlik, R.; Thakral, V.; Raturi, G.; Shinde, S.; Nikolić, M.; Tripathi, D.K.; Sonah, H.; Deshmukh, R. Significance of silicon uptake, transport, and deposition in plants. J. Exp. Bot. 2020, 71, 6703–6718. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Bokor, B.; Bokorová, S.; Ondoš, S.; Švubová, R.; Lukačová, Z.; Hýblová, M.; Szemes, T.; Lux, A. Ionome and expression level of Si transporter genes (Lsi1, Lsi2, and Lsi6) affected by Zn and Si interaction in maize. Environ. Sci. Pollut. Res. Int. 2015, 22, 6800–6811. [Google Scholar] [CrossRef] [PubMed]
- Parisi, C.; Vigani, M.; Rodríguez-Cerezo, E. Agricultural Nanotechnologies: What are the current possibilities? Nano Today 2015, 10, 124–127. [Google Scholar] [CrossRef]
- Li, Z.Z.; Chen, J.F.; Liu, F.; Liu, A.Q.; Wang, Q.; Sun, H.Y.; Wen, L.X. Study of UV-shielding properties of novel porous hollow silica nanoparticle carriers for avermectin. Pest. Manag. Sci. 2007, 63, 241–246. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Details of Si-NPs | Soil Properties | Main Effect | Reference | ||||
|---|---|---|---|---|---|---|---|
| Nano-Si Dose and Type of Preparation | Size (nm) | Texture | pH | SOM (g kg−1) | EC (dS m−1) | ||
| Chemical nano-SiO2 (1.5 mM mg Si L−1) | 20–35 | Clay loam | 7.84 | 10.3 | 1.03 | Improves growth and oil yield of coriander under drought stress | [62] |
| Chemical nano-SiO2 fertilizer (2%) | 80–90 | Silty clay | 7.99 | 9.2 | 1.1 | Records the best yield of wheat under water deficit | [63] |
| Chemical nano-SiO2 (60 mg L−1) | 20 | Conditioned soil (data not available) | Mitigates stress in rice by enhancing antioxidant system | [64] | |||
| Chemical chitosan-Si-nano-fertilizer (from 0.01 to 0.16%, w/v) | 100 | Clay | 8.20 | 13.78 | 0.56 | Enhances maize growth and yield by inducing antioxidant defense system | [65] |
| Chemical thiol functionalized nano-SiO2 (4%) | 20 | Silt loam | 7.93 | 13.78 | NA | Remediates polluted soil from heavy metals and improves growth of lettuce | [66] |
| Biological Si-NPs (2.5 and 5.0 mmol L−1) | 38.78 | Loamy | 7.30 | 6.88 | 7.81 | Promotes common bean under saline and polluted soil with Pb, Ni and Cd | [67] |
| Chemical SiO2-NPs (0.75, 1.5 and 2.25 mM) | 10–20 | Sandy loam | 7.10 | NA | 1.20 | Mitigates Cd stress by improving antioxidants and growth of summer savory | [68] |
| Chemical Si-NPs (100–200 mg kg−1) | 8.3 | Clay loam | 6.60 | 7.0 | 0.70 | The 200 mg kg−1 nano-Si + PGPB recorded highest level of Si-soluble and exchangeable fractions under water deficit stress | [69] |
| Chemical SiO2-NPs (150–2000 mg kg−1) | 10 | Commercial soil | 7.35 | NA | NA | Treatment of 500 mg kg−1 lowered the content of As and Cd to 70 and 50% under the water regimes in rice shoots | [70] |
| Chemical SiO2-NPs (at 2 mM) | 30 | Clay loam | 7.40 | 2.80 | 1.7 | Improves maize yield under applied nano-Si in combined with Zn nutrient | [71] |
| Surface-modified nano-silica (3.0%) | 18.0 | NA | 7.61 | 16.9 | NA | Immobilizes bioavailable As, Pb, Cd, by 85, 97.1, 80.1%, res. in polluted soil | [72] |
| Chemical nano-Si complex with glycine, glutamine, histidine | 10–40 | Silty loam | 7.02 | NA | 0.62 | Nano-Si enhances growth of feverfew under drought stress at foliar 1.5 or 3 mM | [73] |
| Chemical Si-NPs (at 1 and 2 mM) | 20 | Loamy | 8.08 | 8.0 | 1.11 | Improves antioxidants to protect sugar beet plants from water deficiency stress | [74] |
| Chemical mercapto-functionalized nano-silica (0.2 to 0.1%) | 20–30 | NA | 8.12 | 19.6 | NA | Increases wheat grain yield by 33.5% and soil dehydrogenase by 43.4% under Cd stress | [75] |
| Chemical nano-SiO2 (500 mg kg−1) | NA | Sandy loam | 7.67 | 2.54 | NA | Enhances remediation by Erigeron annuus L. grown in polluted soil by PAH (150 mg kg−1) | [76] |
| Structure of Si-NPs | Crop | Concentration | Adaptive Mechanism | Reference |
|---|---|---|---|---|
| SiO2 (chemical) | Saccharum officinarum L. | 300 ppm | Improves leaf photosynthetic responses, chlorophyll fluorescence yield, photosynthetic pigments and photosynthetic apparatus (PS II) during chilling stress | [94] |
| SiO2 (chemical) | Glycine max L. | 100–2000 ppm | Si-NPs increase plant performance and reduce the uptake of Hg in epidermis and pericycle of roots and stems. They enhance photosynthetic content and antioxidant enzyme activities in soybean during exposure to mercury (Hg) | [95] |
| SiO2 (chemical) | Hordeum vulgare L. | 125–250 ppm | Improves plant development, green pigments, photosynthetic activities, plant osmolyte and metabolite profiles, cellular damage and membrane stability indicators, and antioxidant enzymes are all affected. | [96] |
| Si-NPs (chemical) | Oryza sativa L. | 1 mM | Enhances gene expression and transportation of cadmium to vacuoles. | [97] |
| SiO2 (commercial) | Trigonella foenumL. | 0–2.5 mM | Increases nanoparticle translocation, accumulation, Si uptake, cell wall lignification and the formation of stress-related enzymes during metal toxicity (cadmium) | [98] |
| Si-NPs (chemical) | Triticum aestivumL. | 10 µM | Mitigates negative effects of UV radiation on plants | [99] |
| Mesoporous Si-NPs (chemical) | Triticum spp. L., Lupinus polyphyllusL. | 200–2000 ppm | Nanoparticles upregulate leaf gas exchange responses and growth development performance of plants | [100] |
| SiO2 (commercial) | Pisum sativum L. | 10 µM | Protects plant seedlings and increases enzymatic activities | [101] |
| Nanoparticle Type | Pathogen | Concentration | Effect | Reference |
|---|---|---|---|---|
| Si-NPs | Fusarium oxysporum f. sp. niveum | 100 mg L−1 | Enhances biomass and fruit yield in comparison to untreated plants | [134] |
| SiO2-NPs | Meloidogyne incognita, Pectobacterium betavasculorum and Rhizoctonia solani | 100, 200 mg L−1 | Si-NPs were most effective against test pathogens | [30] |
| SiO2-NPs | Xanthomonas campestris pv. carotae, Pectobacterium carotovorum and fungi Rhizoctonia solani, Fusarium solani and Alternaria dauci | 100 mg L−1 | Inhibits the growth of all tested pathogens | [23] |
| SiO2-Ag composites | Xanthomonas oryzae >pv. oryzae | 50, 100 and 200 μg mL−1 | Displays antibacterial activity against the tested pathogen | [136] |
| Si | Puccinia melanocephala | 400, 1200 mg L−1 | Reduces disease in sugarcane and induces resistance | [137] |
| Si-NPs | Fusarium oxysporum and Aspergillus niger | 5, 10, 15 kg ha−1 | Reduces the growth of pathogens | [138] |
| Si | Hemileia vastatrix | 0.24 and 0.30 mg kg−1 | Inhibits infection of fungus Hemileia vastatrix and urediniospore germination | [28] |
| Si | Colletotrichum sublineolum | 2 mmol L−1 | Reduces growth by around 20%, acervuli found smaller in size | [139] |
| Si | Podosphaera pannosa | 1 mg mL−1 | Reduces disease severity by 46% and induces phenolic acid formation | [123] |
| Si | Fusarium sulphureum | 100 and 200 mM | Decreases pathogen growth and reduces disease | [140] |
| SiO2 | Sclerosporagraminicola | 5, 10, 15 mM | Inhibits the growth of the fungal pathogen | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K.K.; Singh, A.; et al. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology 2021, 10, 791. https://doi.org/10.3390/biology10080791
Rajput VD, Minkina T, Feizi M, Kumari A, Khan M, Mandzhieva S, Sushkova S, El-Ramady H, Verma KK, Singh A, et al. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology. 2021; 10(8):791. https://doi.org/10.3390/biology10080791
Chicago/Turabian StyleRajput, Vishnu D., Tatiana Minkina, Morteza Feizi, Arpna Kumari, Masudulla Khan, Saglara Mandzhieva, Svetlana Sushkova, Hassan El-Ramady, Krishan K. Verma, Abhishek Singh, and et al. 2021. "Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth" Biology 10, no. 8: 791. https://doi.org/10.3390/biology10080791
APA StyleRajput, V. D., Minkina, T., Feizi, M., Kumari, A., Khan, M., Mandzhieva, S., Sushkova, S., El-Ramady, H., Verma, K. K., Singh, A., Hullebusch, E. D. v., Singh, R. K., Jatav, H. S., & Choudhary, R. (2021). Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology, 10(8), 791. https://doi.org/10.3390/biology10080791