
Two New Species and Two New Records of the Lichen-Forming Fungal Genus Peltula (Ascomycota: Peltulaceae) from China


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Sequence Alignment
2.4. Congruence among Loci
2.5. Phylogeny of the Genus Peltula
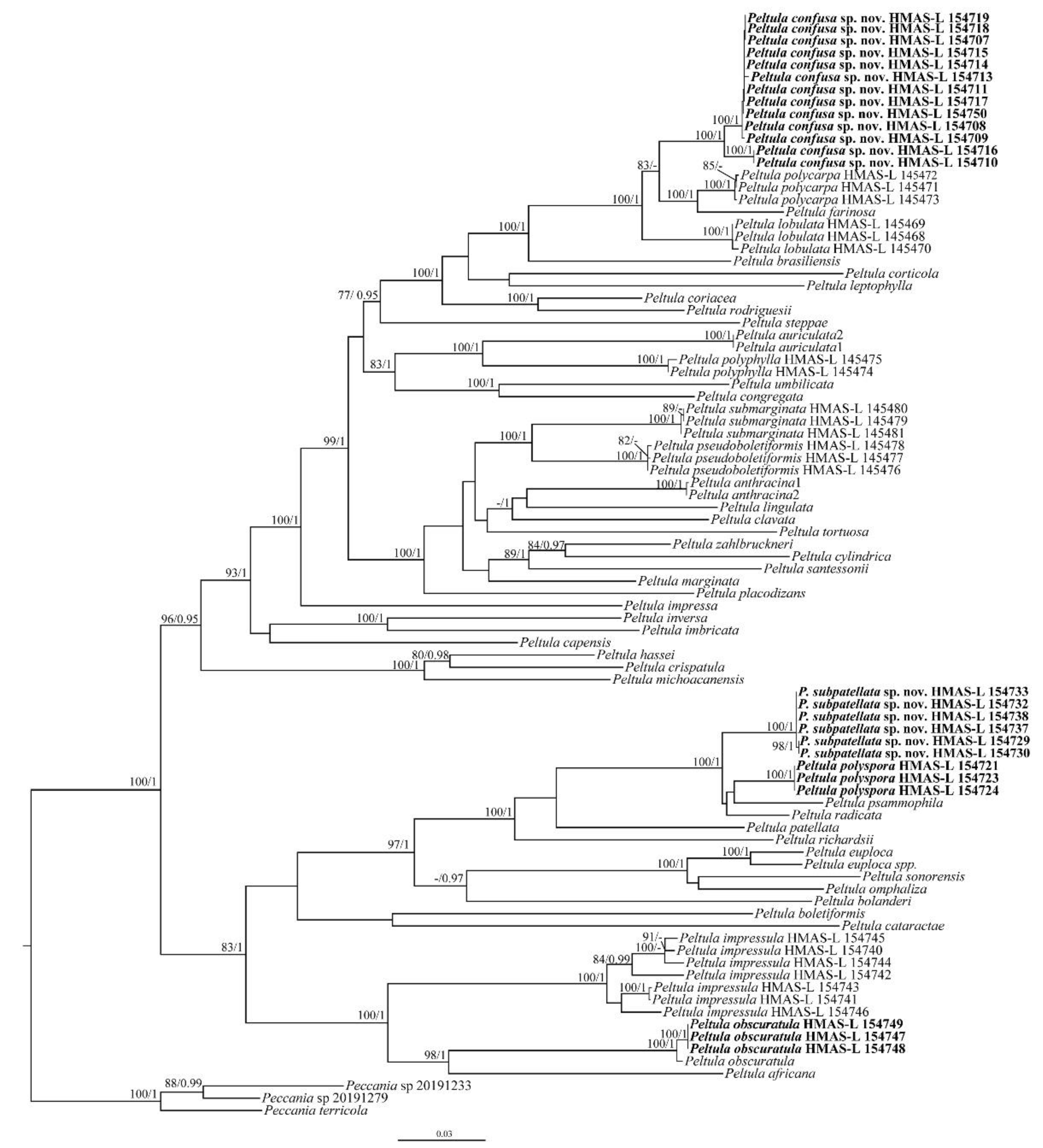
3. Results
3.1. Phylogenetic Analysis
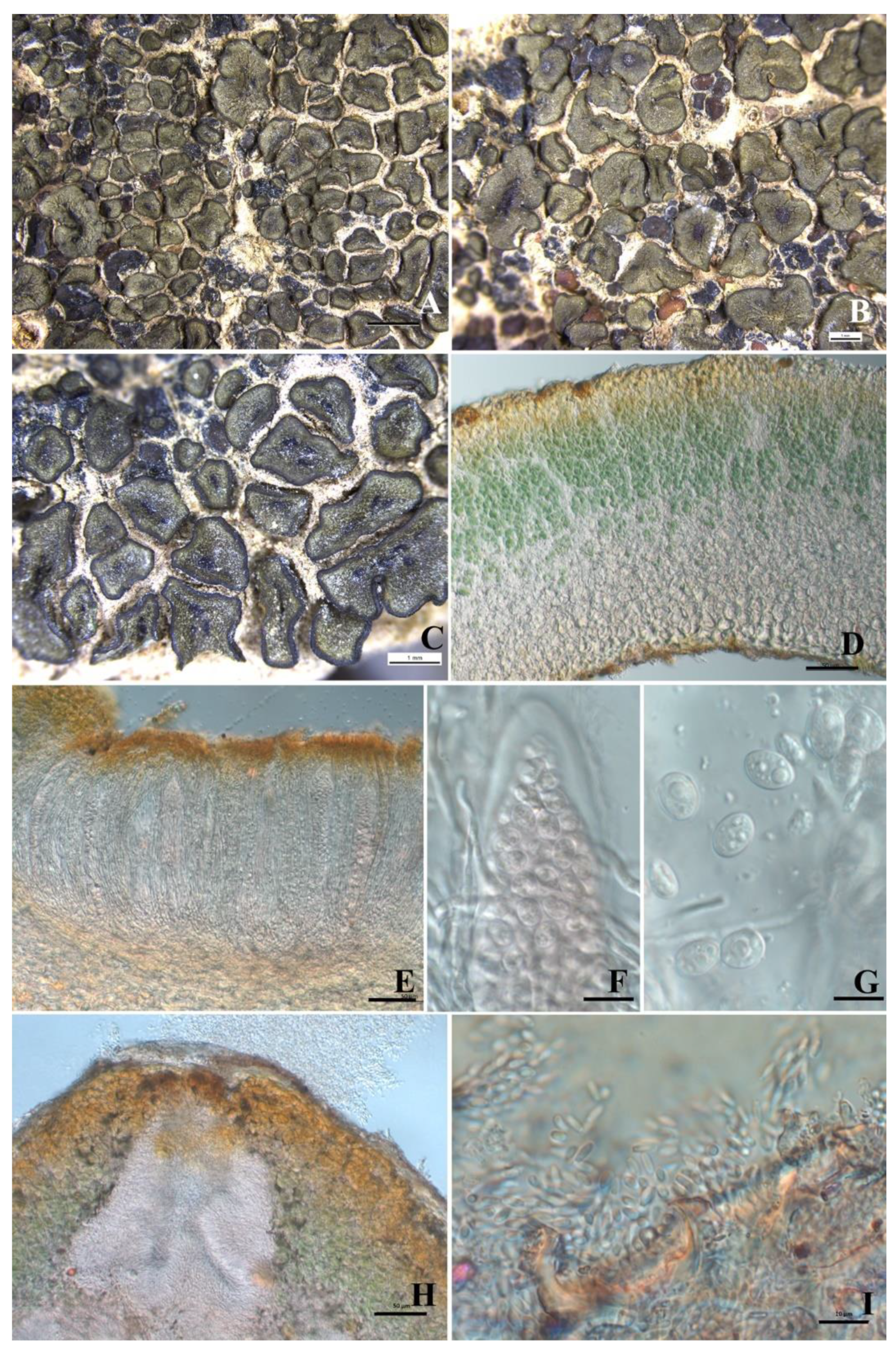
3.2. Taxonomy
3.3. New Records for China
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gyelnik, V. De familia Heppiacearum 1, 2. Repert. Specierum Nov. Regni Veg. 1935, 38, 153–157, 307–313. [Google Scholar]
- Büdel, B. Taxonomy and biology of the lichen genus Peltula Nyl. Bibl. Lichenol. 1987, 25, 209–217. [Google Scholar]
- Büdel, B. The lichen genus Neoheppia. Mycotaxon 1995, 54, 137–145. [Google Scholar]
- Kalb, K. New or otherwise interesting lichens. I. Bibl. Lichenol. 2001, 78, 141–167. [Google Scholar]
- Kauff, F.; Bachran, A.; Schultz, M.; Hofstetter, V.; Lutzoni, F.; Büdel, B. Molecular data favours a monogeneric Peltulaceae (Lichinomycetes). Lichenologist 2018, 50, 313–327. [Google Scholar] [CrossRef]
- Magnusson, A.H. Lichens from Central Asia I; Publ. 13; The Sino-Swedish Expedition: Stockholm, Sweden, 1940; pp. 42–45. [Google Scholar]
- Wu, J.N.; Kang, R.C.; Abbas, A. The lichen genera Cyphelium and Pseudevernia and lichen species Peltula tortuosa first recorded in China from Hanas of Xinjiang. Arid. Zone Res. 1997, 14, 13–15. [Google Scholar]
- Sahedat, G.; Xu, J.; Memet, B.; Nurtai, L.; Abbas, A. New record species of lichen Genus Peltula Nyl. From Inner Mongolia, China. J. Inn. Mogolia Univ. (Nat. Sci. Ed.) 2016, 47, 647–651. [Google Scholar]
- Zahlbruckner, A. Catalogus lichenum universalis. Leipzig 1930, 8, 295. [Google Scholar]
- Aptroot, A.; Seaward, M.D. Annotated checklist of Hongkong Lichens. Trop. Bryol. 1999, 17, 57–101. [Google Scholar]
- Aptroot, A.; Sipman, H.J.M. New Hong Kong lichens, ascomycets and lichenicolous fungi. J. Hattori Bot. Lab. 2001, 91, 317–343. [Google Scholar]
- Aptroot, A.; Sparrius, L.B. New microlichens from Taiwan. Fungal Divers. 2003, 14, 1–50. [Google Scholar]
- Wu, J.L. Lichen Iconography of China; China Prospect Publishing house: Beijing, China, 1987; pp. 60–61. [Google Scholar]
- Yang, Q.X.; Cheng, X.M.; Zhang, T.T.; Liu, X.Z.; Wei, X.L. Five New Species of the Lichen-Forming Fungal Genus Peltula from China. J. Fungi 2022, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual A6; Gelvin, S.B., Schilperoort, R.A., Eds.; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 1–10. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, A Guide to Methods and Application; Academic Press: San Diego, CA, USA, 1990; Volume 1, pp. 315–322. [Google Scholar]
- Thompson, J.; Higgins, D.; Gibson, T. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Mason-Gamer, R.J.; Kellogg, E.A. Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Graminaeae). Syst. Biol. 1996, 45, 524–545. [Google Scholar] [CrossRef]
- Prieto, M.; Martínez, I.; Aragón, G.; Gueidan, C.; Lutzoni, F. Molecular phylogeny of Heteroplacidium, Placidium, and related catapyrenioid genera (Verrucariaceae, lichen-forming Ascomycota). Am. J. Bot. 2012, 99, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Schultz, M.; Van den Boom, P.P.G. Notes on cyanobacterial lichens (mostly Lichinales, Ascomycota) of the Canary Islands. Nova Hedwig. 2007, 84, 113–133. [Google Scholar] [CrossRef]
- Büdel, B.; Nash, T.H., III. A new species of Peltula from the Sonoran Desert. Lichenologist 1993, 25, 279–284. [Google Scholar] [CrossRef]
- Magnusson, A.H. Lichens from Central Asia II; Publ. 22; The Sino-Swedish Expedition: Stockholm, Sweden, 1944; pp. 1–71. [Google Scholar]
- Marton, K.; Galun, M. The cyanophilous lichen population of the Arava Valley and the Judean Desert (Israel). Isr. J. Bot. 1981, 30, 125–155. [Google Scholar]
- Swinscow, T.D.V.; Krog, H. The lichen genera Heppia and Peltula in East Africa. Nor. J. Bot. 1979, 26, 213–224. [Google Scholar]
- Filson, R.B. The lichen genera Heppia and Peltula in Australia. Muelleria 1988, 6, 495–517. [Google Scholar] [CrossRef]
- Upreti, D.K.; Büdel, B. The lichen genera Heppia and Peltula in India. J. Hattori Bot. Lab. 1990, 68, 279–284. [Google Scholar]
- Wetmore, C.M. The lichen family Heppiaceae in North America. Ann. Mo. Bot. Gard. 1970, 57, 158–209. [Google Scholar] [CrossRef]
- Egea, J.M. Los géneros Heppia y Peltula (Líquenes) en Europa Occidental y Norte de Africa. Bibl. Lichenol. 1989, 31, 1–122. [Google Scholar]
- Tuckerman, E. Genera Lichenum: An Arrangement of the North American Lichens; Edwin Nelson: Amherst, MA, USA, 1872; pp. 114–115. [Google Scholar]
- Büdel, B. Zur Biologie und Systematik der Flechtengattung Heppia und Peltula im sudlichen Afrika. Bibl. Lichenol. 1987, 23, 1–105. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Numbers | |||||
|---|---|---|---|---|---|
| Species | Voucher Information | nrSSU | nrLSU | ITS | RPB2 |
| Peltula africana | South Africa, 1990, B.B. 14304b | MF766261 | MF766384 | MF766343 | MF804877 |
| P. anthracina1 | Brazil, 2010, CGMS 385 | — | — | MW267988 | — |
| P. anthracina2 | Brazil, 2010, CGMS 384 | — | — | MW267989 | — |
| P. auriculata1 | Venezuela, 1992, B.B. 24901 | — | DQ832330 | DQ832329 | — |
| P. auriculata2 | Venezuela, 1992, B.B. 24902 | MF766262 | MF766385 | MF766344 | MF804860 |
| P. bolanderi | Mexico, 1993, B.B. 20196e | MF766263 | MF766386 | MF766345 | MF804891 |
| P. boletiformis | South Africa, 2003, B.B. 14911a-1 | MF766264 | MF766387 | MF766346 | MF804892 |
| P. brasiliensis | South Africa, 1983, B.B. 14083a | MF766298 | MF766421 | MF766380 | MF804874 |
| P. capensis | South Africa, 1994, B.B. 14382b 2 | MF766265 | MF766388 | MF766347 | MF804893 |
| P. cataractae | Dem. Rp. Congo, 1947, B.B. 1329 | MF766299 | MF766422 | MF766381 | MF804875 |
| P. clavata | Australia, 1987, DUKE 164 (18047a) | MF766266 | MF766389 | MF766348 | MF804861 |
| P. confusa | 20191485 (HMAS-L 154714) | — | OP429688 | OP429718 | — |
| P. confusa | 20191619 (HMAS-L 154710) | OP429768 | OP429710 | OP429719 | — |
| P. confusa | 20191620 (HMAS-L 154707) | OP429760 | OP429685 | OP429720 | OP348234 |
| P. confusa | 20191621 (HMAS-L 154711) | OP429761 | OP429689 | OP429721 | OP348228 |
| P. confusa | 20191622 (HMAS-L 154708) | OP429762 | OP429686 | OP429722 | OP348233 |
| P. confusa | 20191623 (HMAS-L 154709) | OP429763 | OP429690 | OP429723 | OP348229 |
| P. confusa | 20191646-2 (HMAS-L 154715) | — | OP429691 | OP429724 | — |
| P. confusa | 20191648 (HMAS-L 154713) | — | OP429692 | OP429726 | OP348231 |
| P. confusa | 20191928 (HMAS-L 154716) | OP429765 | OP429693 | OP429727 | OP348232 |
| P. confusa | 20191944 (HMAS-L 154717) | OP429764 | OP429687 | OP429725 | OP348230 |
| P. confusa | 20192000 (HMAS-L 154718) | — | OP429694 | OP429728 | OP348226 |
| P. confusa | 20192011 (HMAS-L 154719) | — | OP429695 | OP429729 | OP348227 |
| P. confusa | 20192022 (HMAS-L 154750) | — | OP429696 | OP429730 | — |
| P. congregata | South Africa, 2003, B.B. 14909b-1 | MF766267 | MF766390 | MF766349 | MF804896 |
| P. coriacea | South Africa, 2003, B.B. 14500a-1 | MF766268 | MF766391 | MF766350 | MF804878 |
| P. corticola | Yemen, 2002, M.S. 14201 | MF766296 | MF766419 | MF766378 | MF804873 |
| P. crispatula | Morocco, 1987, B.B. 21001a | MF766269 | MF766392 | MF766351 | MF804862 |
| P. cylindrica | South Africa, 2003, B.B. 14920a-1 | MF766270 | MF766393 | MF766352 | — |
| P. euploca | Mexico, 1993, B.B. 20162a | MF766271 | MF766394 | MF766353 | MF804879 |
| P. euploca ssp. sorediosa | South Africa, 2003, B.B. 14921c-1 | MF766272 | MF766395 | MF766354 | MF804880 |
| P. farinosa | Mexico, 1993, B.B. 20119a | MF766273 | MF766396 | MF766355 | MF804898 |
| P. hassei | South Africa, 1994, B.B. 14354a | MF766283 | MF766406 | MF766365 | MF804895 |
| P. imbricata | Australia, 1987, B.B. 18060a | MF766274 | MF766397 | MF766356 | MF804899 |
| P. impressa | Mexico, 1993, B.B. 20140f | MF766275 | MF766398 | MF766357 | MF804863 |
| P. impressula | China, 2019, HMAS-L 154740 | OP429747 | OP429698 | OP429737 | — |
| P. impressula | China, 2019, HMAS-L 154741 | OP429746 | OP429697 | OP429738 | — |
| P. impressula | China, 2019, HMAS-L 154742 | — | OP429699 | OP429743 | — |
| P. impressula | China, 2017, HMAS-L 154743 | OP429748 | OP429700 | OP429739 | — |
| P. impressula | China, 2017, HMAS-L 154744 | OP429749 | OP429701 | OP429740 | — |
| P. impressula | China, 2017, HMAS-L 154745 | OP429750 | OP429702 | OP429741 | — |
| P. impressula | China, 2017, HMAS-L 154746 | OP429751 | OP429703 | OP429742 | — |
| P. inversa | Namibia, 2001, Pretoria 15058 | MF766276 | MF766399 | MF766358 | MF804881 |
| P. leptophylla | Mexico, 1993, B.B. 20128a | MF766277 | MF766400 | MF766359 | MF804864 |
| P. lingulata | South Africa, 1994, B.B. 14452a | MF766278 | MF766401 | MF766360 | MF804882 |
| P. lobulata | China, 2019, HMAS-L 145468 | — | MT499313 | MT499291 | — |
| P. lobulata | China, 2019, HMAS-L 145469 | — | MT499314 | MT499292 | — |
| P. lobulata | China, 2019, HMAS-L 145470 | — | MT499315 | MT499293 | — |
| P. marginata | South Africa, 2003, B.B. 14920d-1 | MF766279 | MF766402 | MF766361 | MF804883 |
| P. michoacanensis | Mexico, 1993, B.B. 20140l | MF766280 | MF766403 | MF766362 | MF804900 |
| P. obscuratula | 20191465 (HMAS-L 154747) | OP429752 | OP429708 | OP429735 | — |
| P. obscuratula | 20191491 (HMAS-L 154748) | — | OP429707 | OP429734 | — |
| P. obscuratula | 20191493 (HMAS-L 154749) | OP429753 | OP429709 | OP429736 | — |
| P. obscuratula | Morocco, 1987, B.B. ex Murcia | MF766284 | MF766407 | MF766366 | MF804867 |
| P. omphaliza | Mexico, 1993, B.B. 20148b | MF766285 | MF766408 | MF766367 | MF804884 |
| P. patellata | Mexico, 2003, M.S. 16254b | MF766286 | MF766409 | MF766368 | MF804868 |
| P. placodizans | Mexico, 1993, B.B. 20112a | MF766287 | MF766410 | MF766369 | MF804885 |
| P. polycarpa | China, 2019, HMAS-L 145471 | MT499282 | MT499319 | MT499300 | OP348224 |
| P. polycarpa | China, 2019, HMAS-L 145472 | MT499286 | MT499320 | MT499301 | — |
| P. polycarpa | China, 2019, HMAS-L 145473 | MT499287 | MT499321 | MT499302 | OP348225 |
| P. polyphylla | China, 2019, HMAS-L 145475 | — | MT499326 | MT499303 | — |
| P. polyphylla | China, 2019, HMAS-L 145474 | — | MT499325 | MT499304 | OP348221 |
| P. polyspora | 20190012 (HMAS-L 154721) | OP429758 | OP429706 | OP429732 | — |
| P. polyspora | 20190196 (HMAS-L 154723) | OP429757 | OP429704 | OP429733 | — |
| P. polyspora | 20191252 (HMAS-L 154724) | OP429759 | OP429705 | OP429731 | — |
| P. psammophila | Canary Islands, 1985, BM 761074 | MF766288 | MF766411 | MF766370 | MF804869 |
| P. pseudoboletiformis | China, 2019, HMAS-L 145476 | MT499288 | MT499322 | MT499297 | OP348222 |
| P. pseudoboletiformis | China, 2019, HMAS-L 145478 | MT499289 | MT499323 | MT499298 | — |
| P. pseudoboletiformis | China, 2019, HMAS-L 145477 | MT499290 | MT499324 | MT499299 | — |
| P. radicata | Yemen, 2002, M.S. 14241a | MF766289 | MF766412 | MF766371 | MF804870 |
| P. richardsii | Mexico, 1993, B.B. 20194a | MF766290 | MF766413 | MF766372 | MF804871 |
| P. rodriguesii | Namibia, 1990, B.B. 15901 | MF766291 | MF766414 | MF766373 | MF804872 |
| P. santessonii | South Africa, 2003, B.B. 14912b-1 | MF766292 | MF766415 | MF766374 | MF804886 |
| P. sonorensis | Mexico, 1993, B.B. 20196d | MF766293 | MF766416 | MF766375 | MF804887 |
| P. steppae | Venezuela, 1989, K.K. 23948 | MF766297 | MF766420 | MF766379 | MF804890 |
| P. submarginata | China, 2019, HMAS-L 145480 | MT499283 | MT499316 | MT499294 | — |
| P. submarginata | China, 2019, HMAS-L 145479 | MT499284 | MT499317 | MT499296 | OP348223 |
| P. submarginata | China, 2019, HMAS-L 145481 | MT499285 | MT499318 | MT499295 | — |
| P. subpatellata | 20190050 (HMAS-L 154729) | — | OP429679 | OP429712 | — |
| P. subpatellata | 20190053 (HMAS-L 154730) | — | OP429680 | OP429713 | — |
| P. subpatellata | 20190112 (HMAS-L 154732) | OP429754 | OP429681 | OP429714 | — |
| P. subpatellata | 20190143 (HMAS-L 154733) | — | OP429682 | OP429715 | — |
| P. subpatellata | XL2017268 (HMAS-L 154738) | OP429755 | OP429683 | OP429716 | — |
| P. subpatellata | XL2017279 (HMAS-L 154737) | OP429756 | OP429684 | OP429717 | — |
| P. tortuosa | Venezuela, 1996, B.B. 24039b | MF766294 | MF766417 | MF766376 | — |
| P. umbilicata | South Africa, 2003, B.B. 14901a-1 | DQ782887 | DQ832334 | DQ832333 | DQ832335 |
| P. zahlbruckneri | Mexico, 1993, B.B. 20157a | — | MF766418 | — | MF804889 |
| Peccania terricola | China, Xinjiang University, 201899118 | — | OM523033 | OM523029 | — |
| Peccania sp | China, 2019, HMAS-L 154764 | OP429766 | OP429711 | OP429744 | — |
| Peccania sp | China, 2019, HMAS-L 154765 | OP429767 | — | OP429745 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Hollinger, J.; Leavitt, S.D.; Wei, X. Two New Species and Two New Records of the Lichen-Forming Fungal Genus Peltula (Ascomycota: Peltulaceae) from China. Biology 2022, 11, 1518. https://doi.org/10.3390/biology11101518
Yang Q, Hollinger J, Leavitt SD, Wei X. Two New Species and Two New Records of the Lichen-Forming Fungal Genus Peltula (Ascomycota: Peltulaceae) from China. Biology. 2022; 11(10):1518. https://doi.org/10.3390/biology11101518
Chicago/Turabian StyleYang, Qiuxia, Jason Hollinger, Steven D. Leavitt, and Xinli Wei. 2022. "Two New Species and Two New Records of the Lichen-Forming Fungal Genus Peltula (Ascomycota: Peltulaceae) from China" Biology 11, no. 10: 1518. https://doi.org/10.3390/biology11101518
APA StyleYang, Q., Hollinger, J., Leavitt, S. D., & Wei, X. (2022). Two New Species and Two New Records of the Lichen-Forming Fungal Genus Peltula (Ascomycota: Peltulaceae) from China. Biology, 11(10), 1518. https://doi.org/10.3390/biology11101518