
Endoglin and Activin Receptor-like Kinase 1 (Alk1) Modify Adrenomedullin Expression in an Organ-Specific Manner in Mice

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Collection
2.2. Chemiluminescent Enzyme Immunoassay (EIA)
2.3. RNA Extraction, Reverse Transcription, and Real Time Polymerase Chain Reaction
2.4. Immunohistochemistry
2.5. Quantification of Immunohistochemical Signals
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
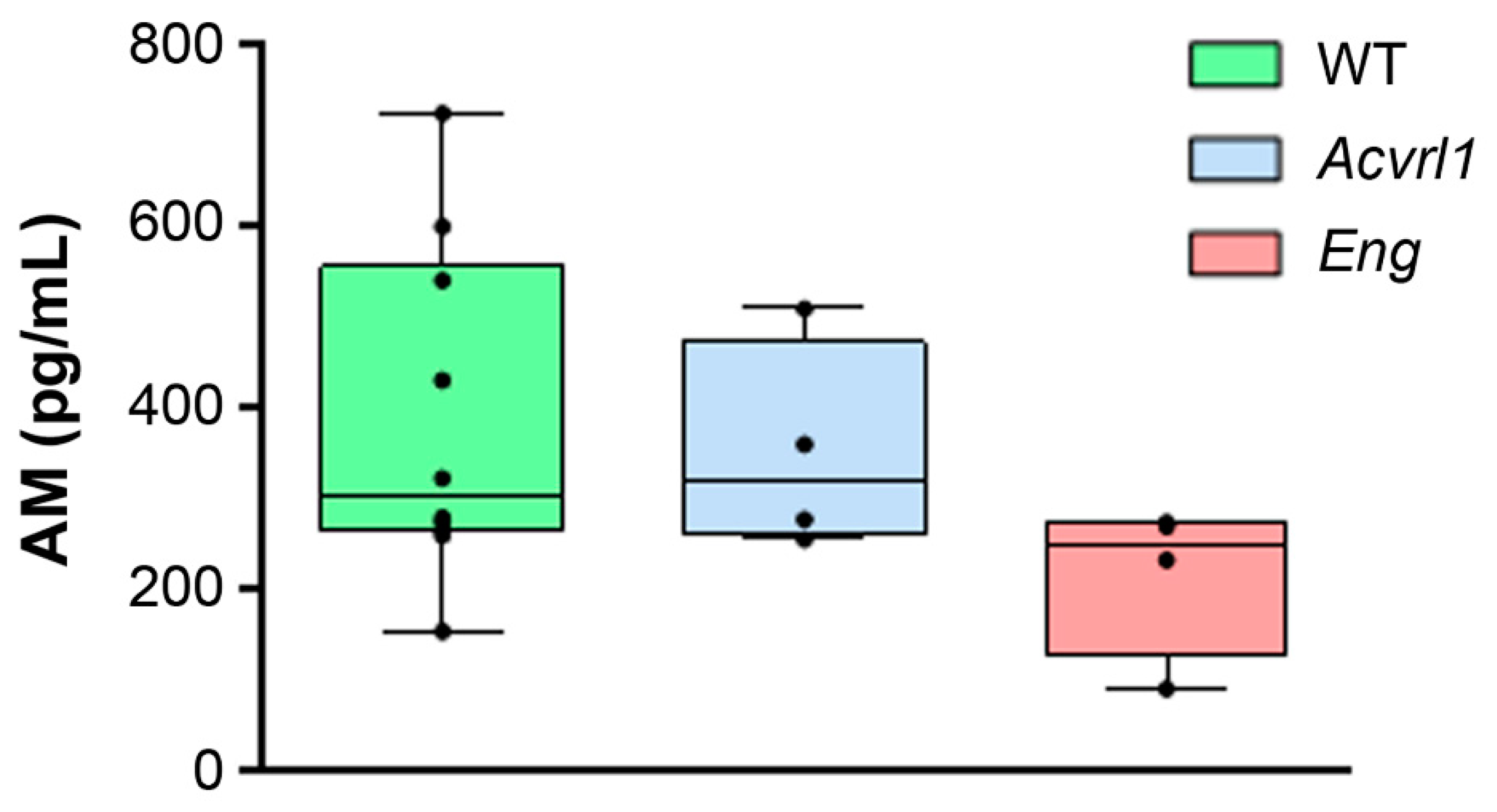
3.1. AM Blood Levels
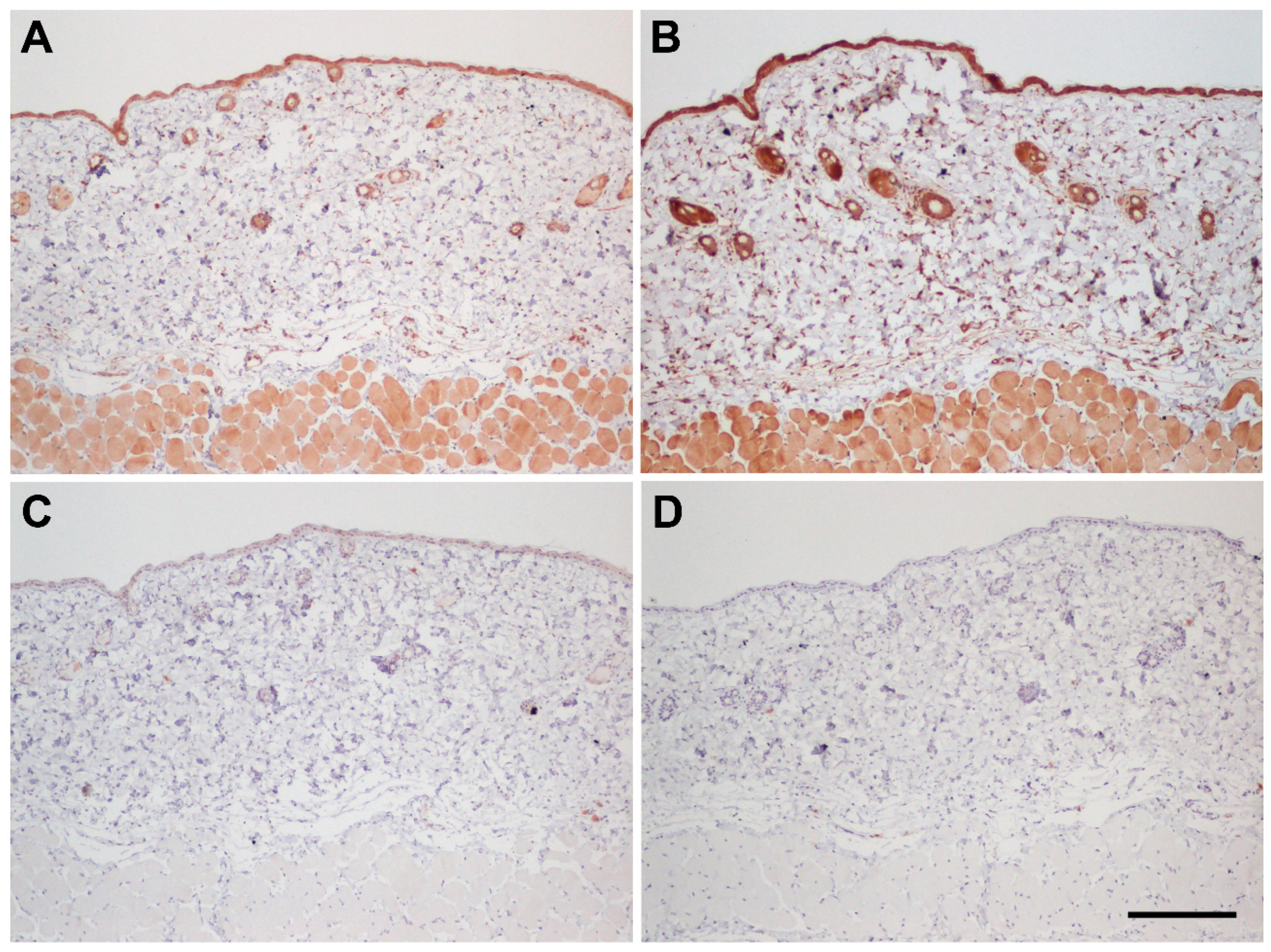
3.2. Immunohistochemical Controls
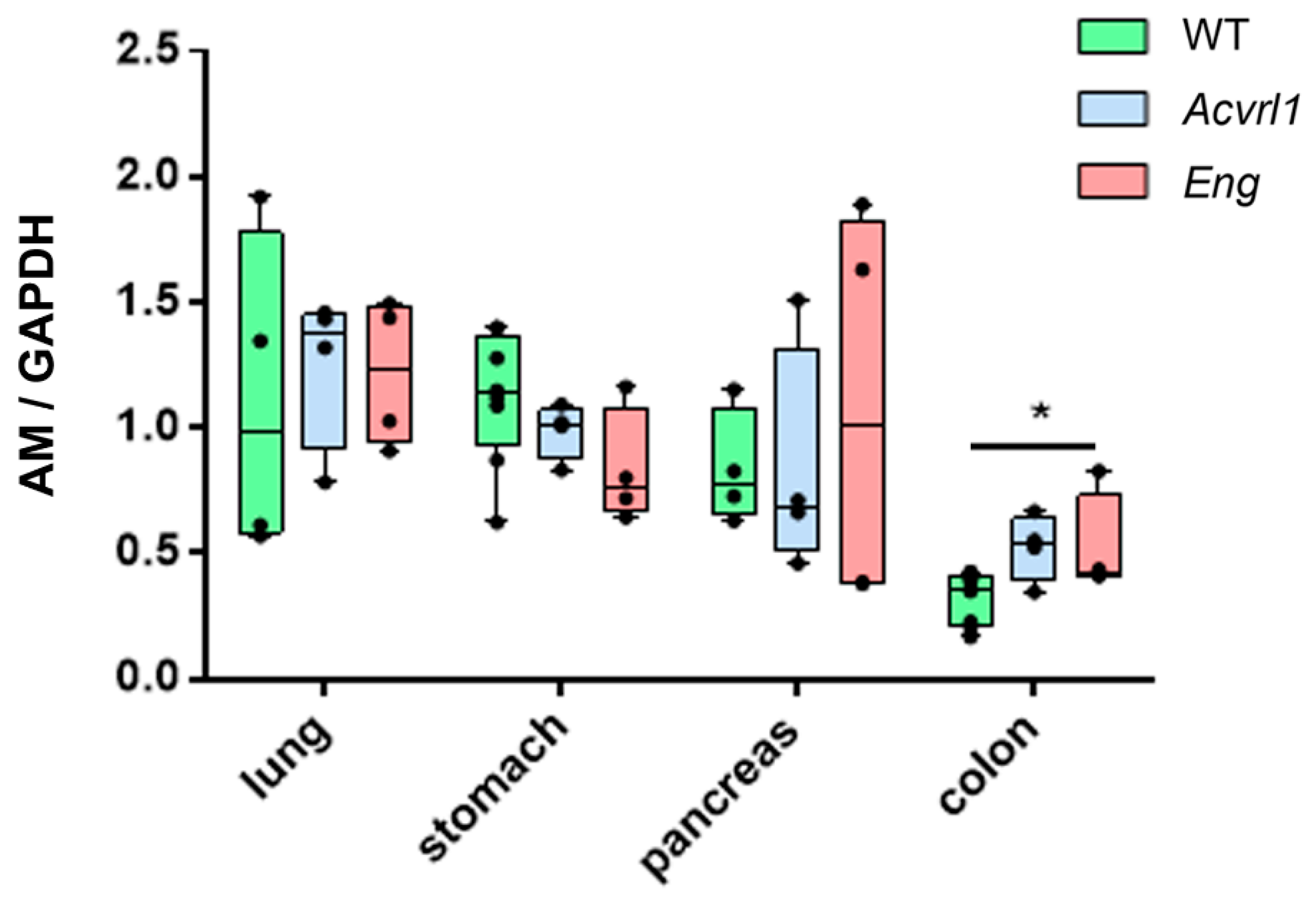
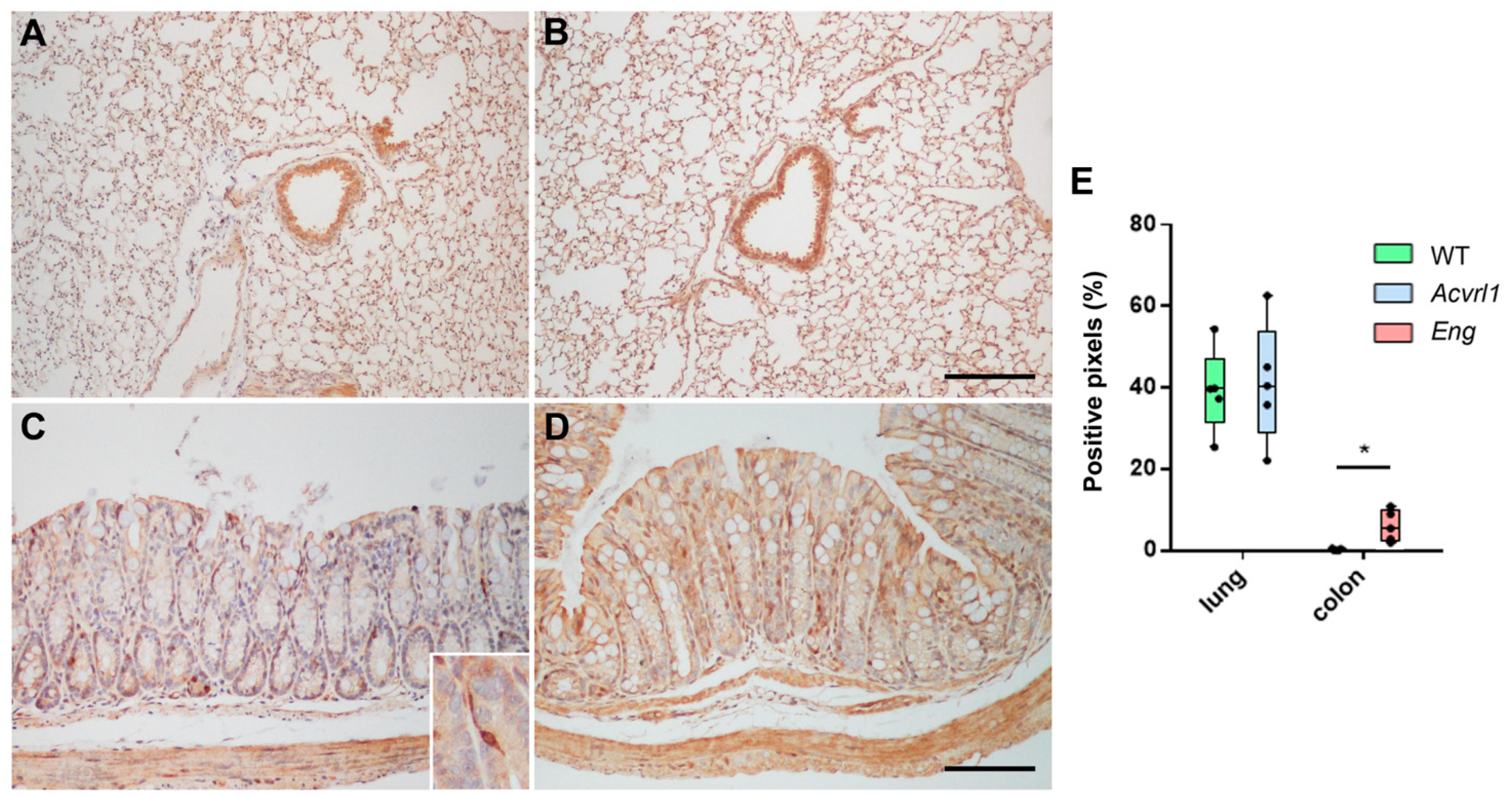
3.3. Expression of AM in Organs of Endodermic Origin
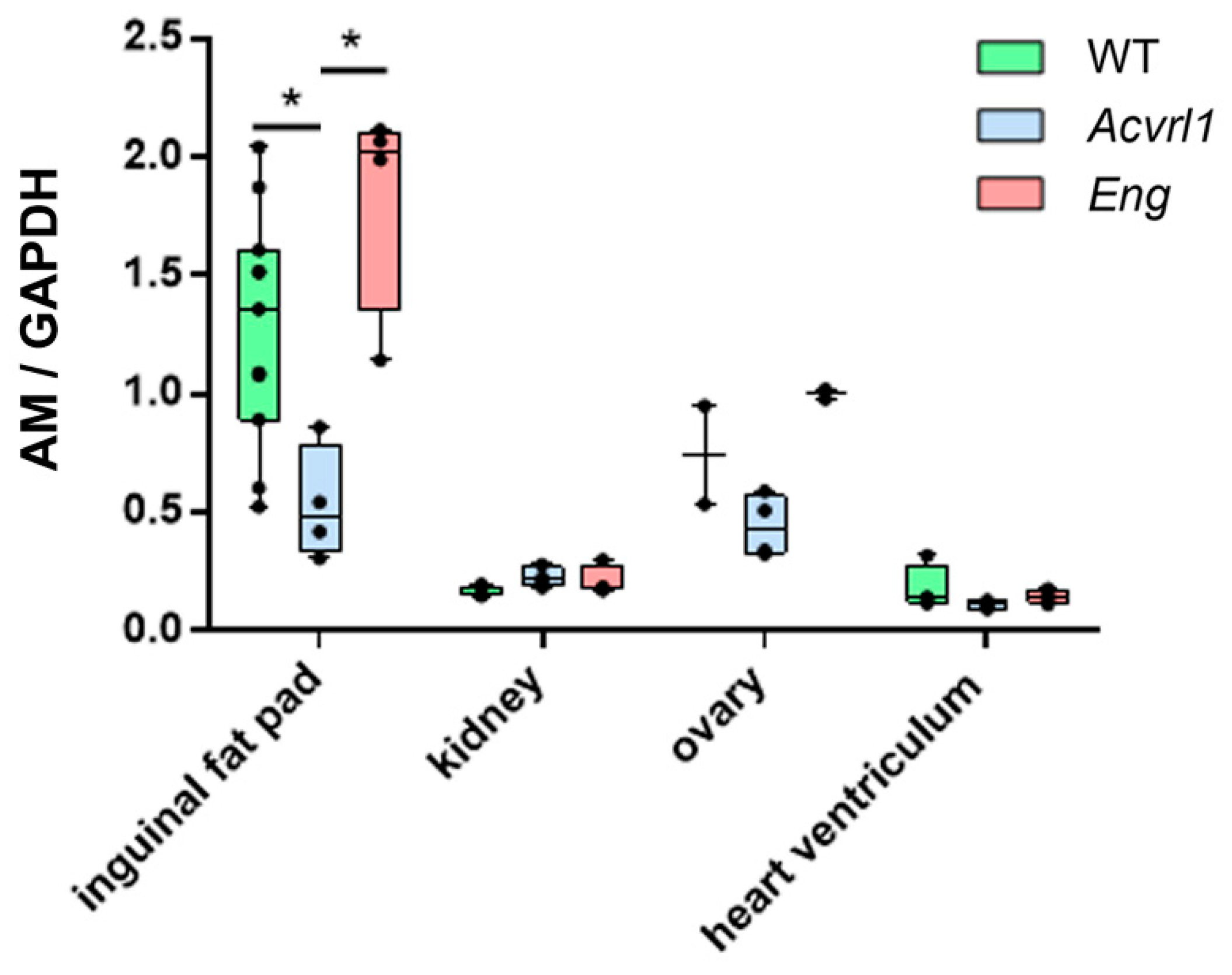
3.4. Expression of AM in Organs of Mesodermic Origin
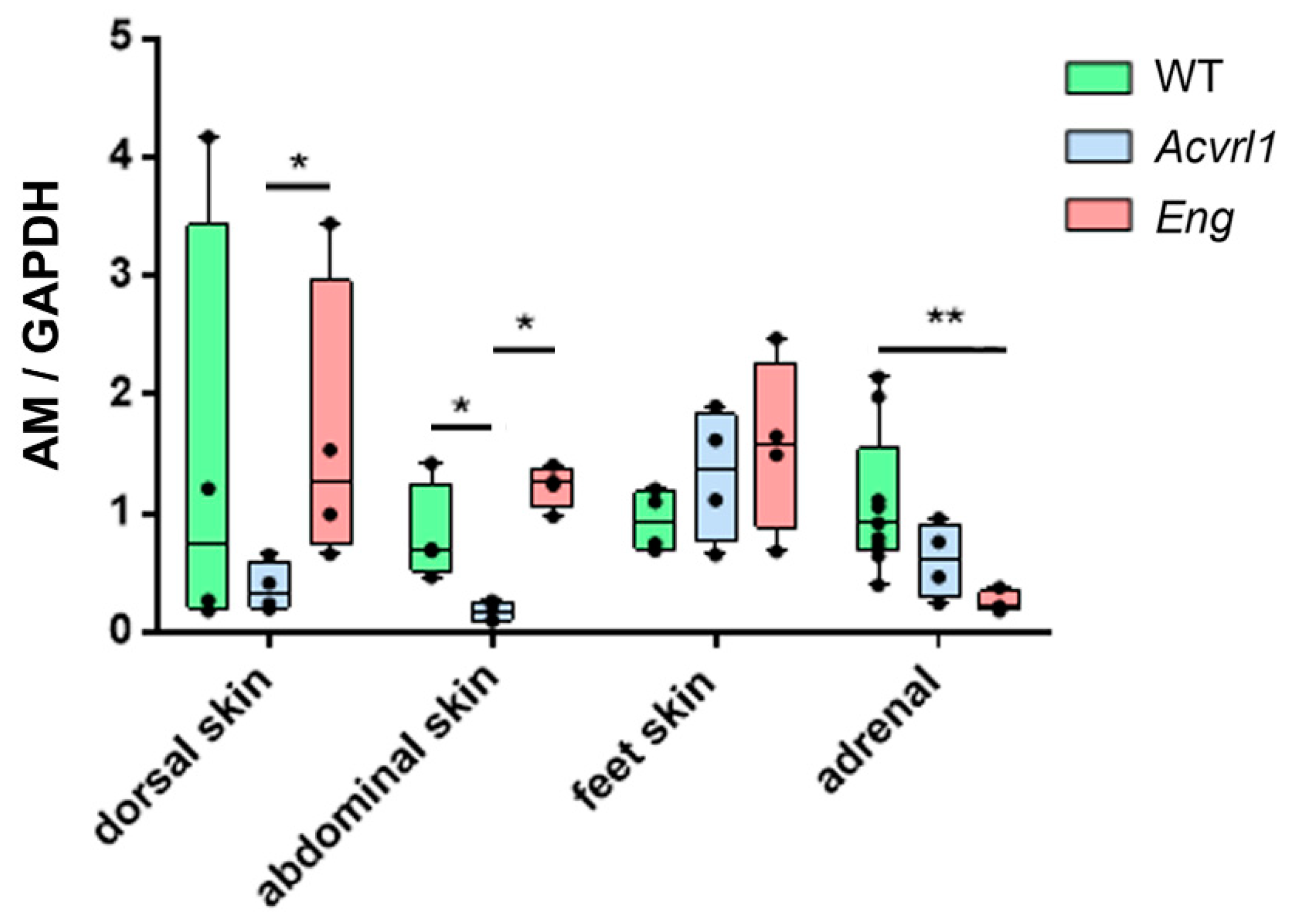
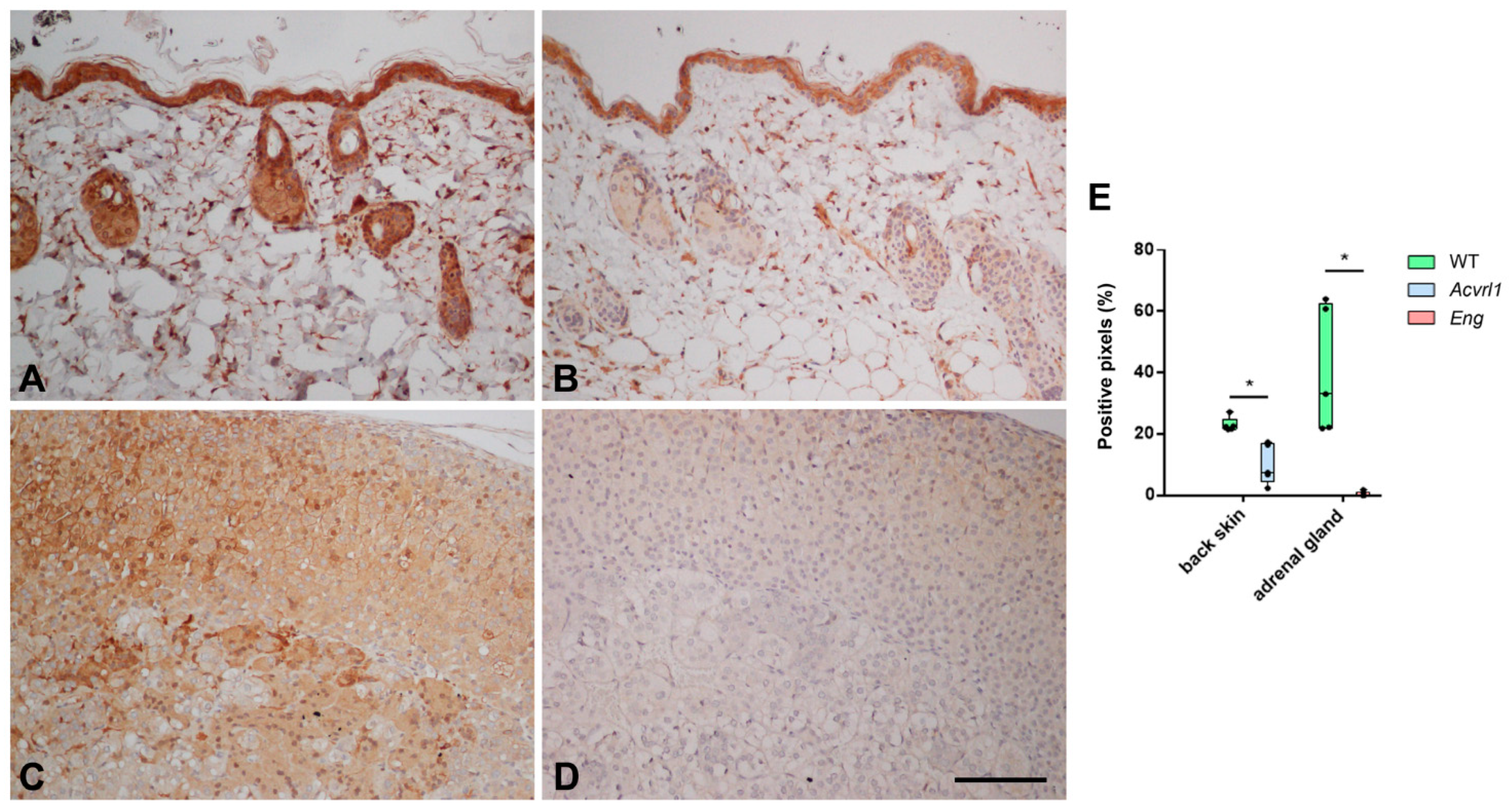
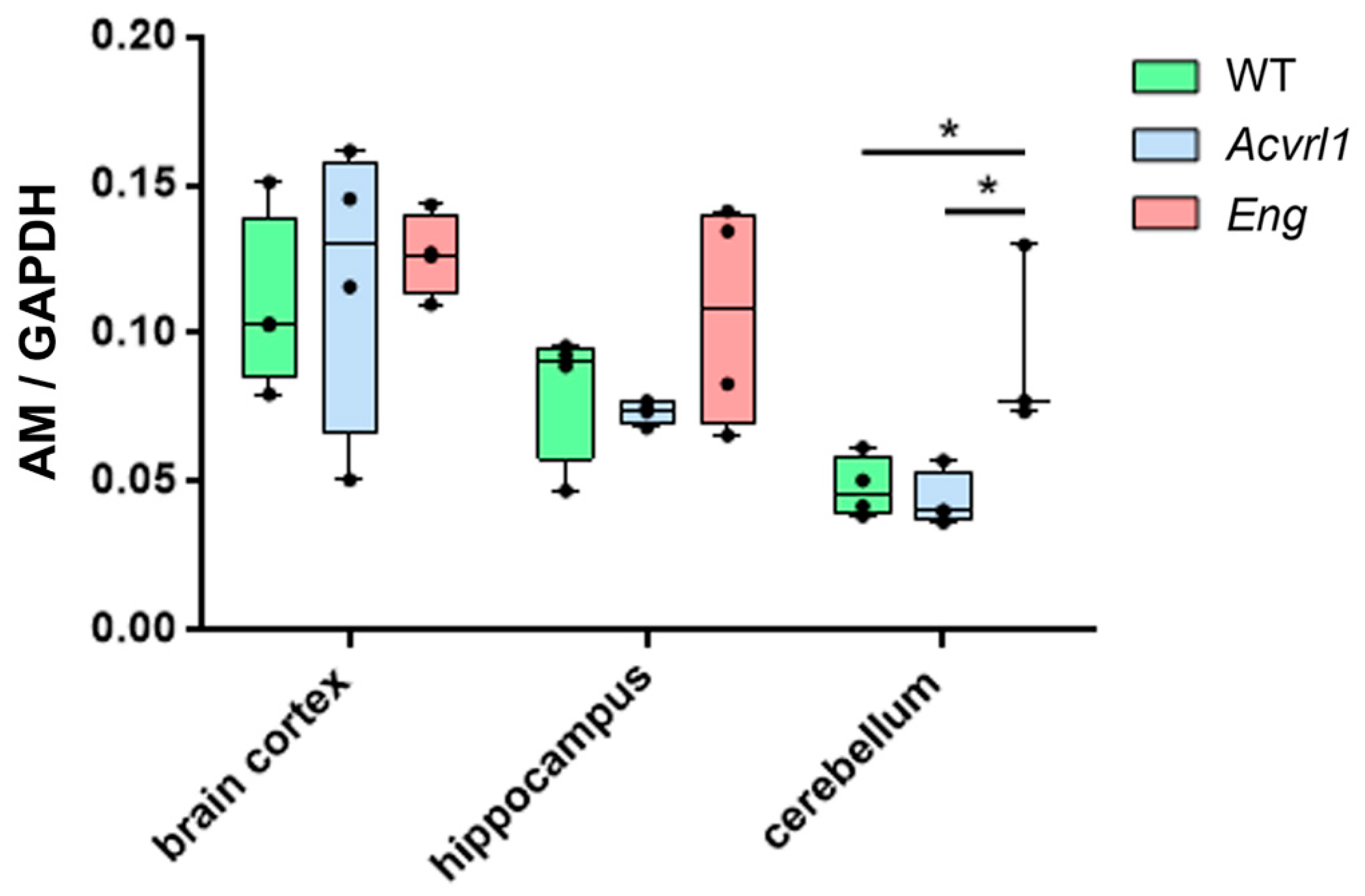
3.5. Expression of AM in Organs of Ectodermic Origin
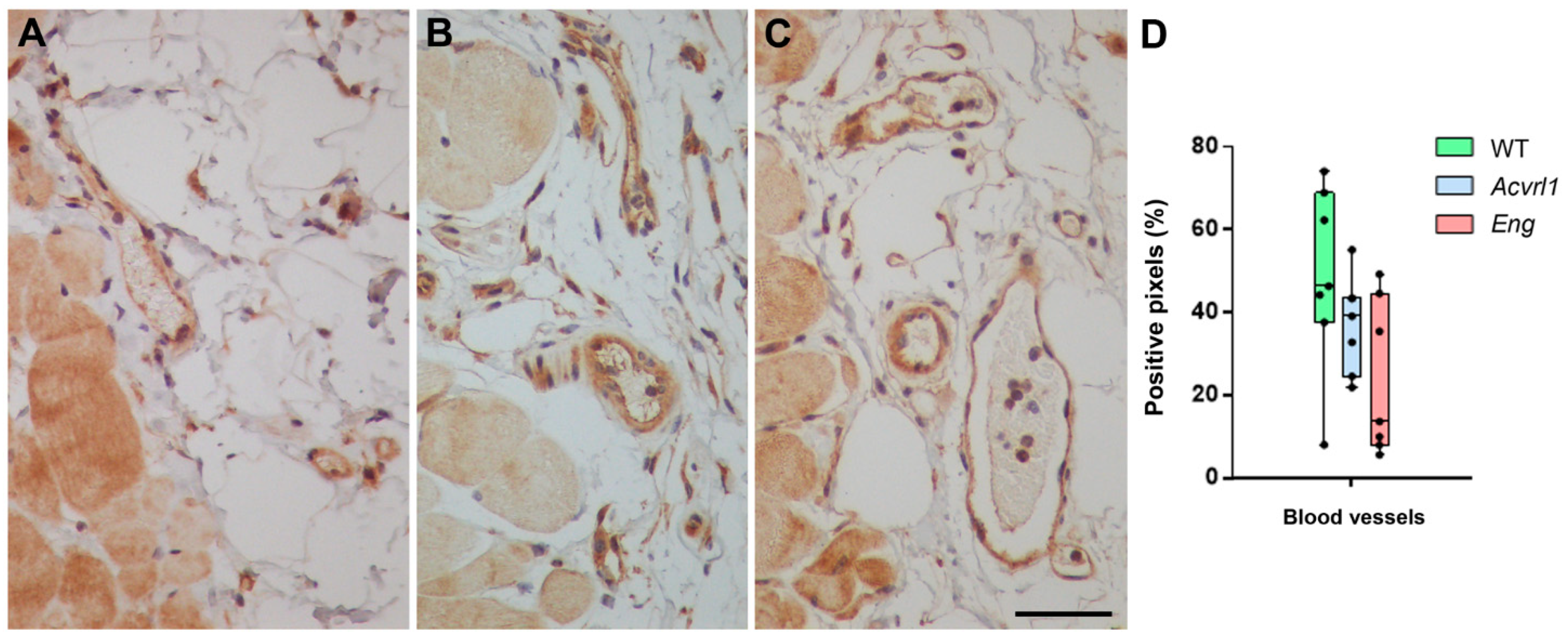
3.6. Expression of AM in Blood Vessels
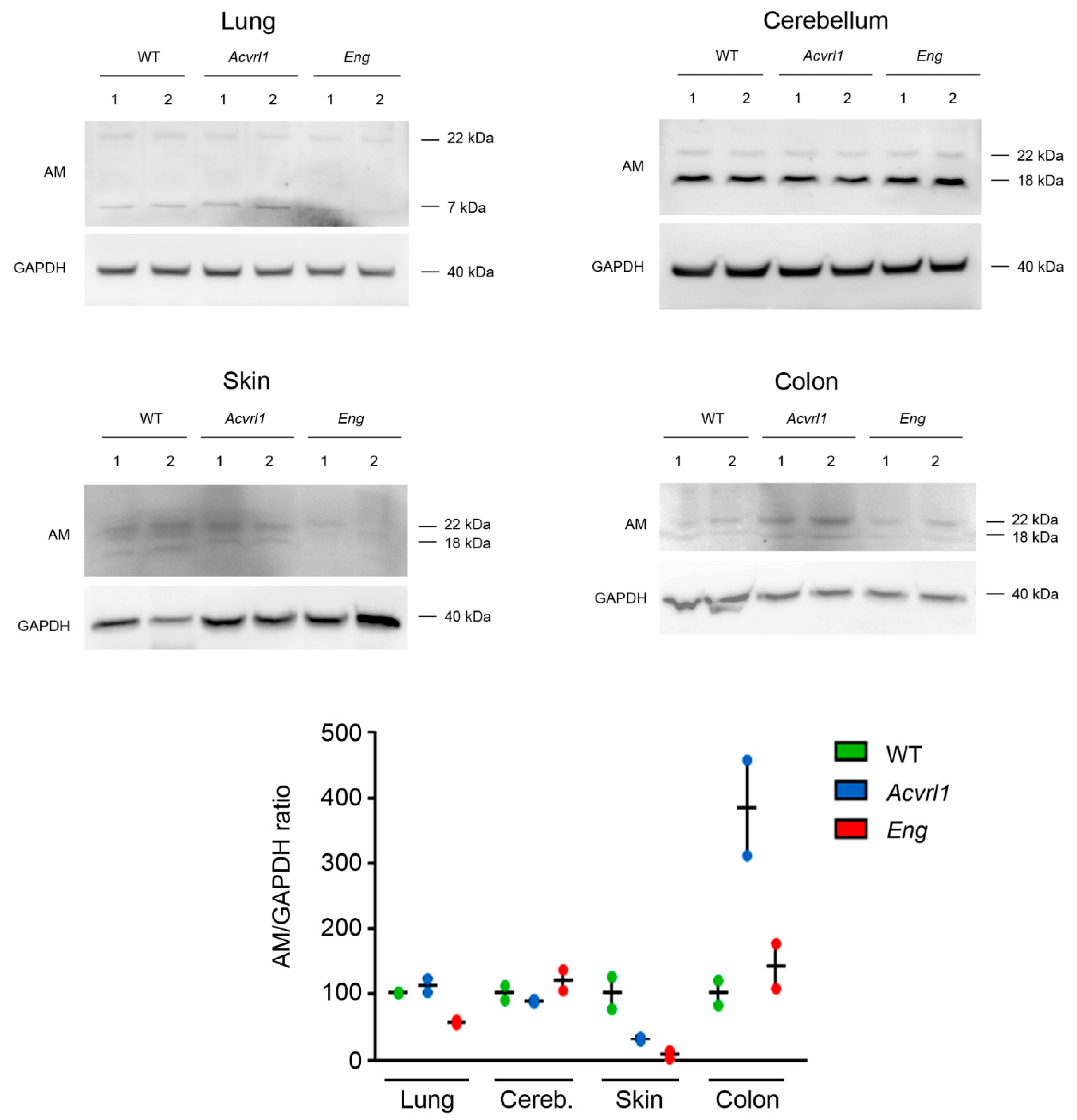
3.7. Western Blots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Orphanet. The Portal for Rare Diseases and Orphan Drugs. Available online: https://www.orpha.net/consor/cgi-bin/index.php (accessed on 22 December 2021).
- Faughnan, M.E.; Mager, J.J.; Hetts, S.W.; Palda, V.A.; Lang-Robertson, K.; Buscarini, E.; Deslandres, E.; Kasthuri, R.S.; Lausman, A.; Poetker, D.; et al. Second International Guidelines for the Diagnosis and Management of Hereditary Hemorrhagic Telangiectasia. Ann. Intern. Med. 2020, 173, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martínez, R.; Iriarte, A.; Mora-Luján, J.M.; Patier, J.L.; López-Wolf, D.; Ojeda, A.; Torralba, M.A.; Juyol, M.C.; Gil, R.; Añón, S.; et al. Current HHT genetic overview in Spain and its phenotypic correlation: Data from RiHHTa registry. Orphanet J. Rare Dis. 2020, 15, 138. [Google Scholar] [CrossRef] [PubMed]
- Alsina-Sanchís, E.; García-Ibáñez, Y.; Figueiredo, A.; Riera-Domingo, C.; Figueras, A.; Matias-Guiu, X.; Casanovas, O.; Botella, L.M.; Pujana, M.A.; Riera-Mestre, A.; et al. ALK1 Loss Results in Vascular Hyperplasia in Mice and Humans Through PI3K Activation. Arter. Thromb. Vasc. Biol. 2018, 38, 1216–1229. [Google Scholar] [CrossRef] [Green Version]
- Tillet, E.; Bailly, S. Emerging roles of BMP9 and BMP10 in hereditary hemorrhagic telangiectasia. Front. Genet. 2015, 5, 456. [Google Scholar] [CrossRef] [Green Version]
- Bourdeau, A.; Dumont, D.J.; Letarte, M. A murine model of hereditary hemorrhagic telangiectasia. J. Clin. Investig. 1999, 104, 1343–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, T.; Yun, J.; Oh, S.P. Arterial Endothelium-Specific Activin Receptor-Like Kinase 1 Expression Suggests Its Role in Arterialization and Vascular Remodeling. Circ. Res. 2003, 93, 682–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerkic, M.; Rivas-Elena, J.V.; Santibanez, J.F.; Prieto, M.; Rodríguez-Barbero, A.; Perez-Barriocanal, F.; Pericacho, M.; Arévalo, M.; Vary, C.P.; Letarte, M.; et al. Endoglin Regulates Cyclooxygenase-2 Expression and Activity. Circ. Res. 2006, 99, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerkic, M.; Kabir, M.G.; Davies, A.; Yu, L.X.; McIntyre, B.A.; Husain, N.W.; Enomoto, M.; Sotov, V.; Husain, M.; Henkelman, M.; et al. Pulmonary hypertension in adult Alk1 heterozygous mice due to oxidative stress. Cardiovasc. Res. 2011, 92, 375–384. [Google Scholar] [CrossRef]
- Iriarte, A.; Ochoa-Callejero, L.; García-Sanmartín, J.; Cerdà, P.; Garrido, P.; Narro-Íñiguez, J.; Mora-Luján, J.; Jucglà, A.; Sánchez-Corral, M.; Cruellas, F.; et al. Adrenomedullin as a potential biomarker involved in patients with hereditary hemorrhagic telangiectasia. Eur. J. Intern. Med. 2021, 88, 89–95. [Google Scholar] [CrossRef]
- Kitamura, K.; Kangawa, K.; Kawamoto, M.; Ichiki, Y.; Nakamura, S.; Matsuo, H.; Eto, T. Adrenomedullin: A Novel Hypotensive Peptide Isolated from Human Pheochromocytoma. Biochem. Biophys. Res. Commun. 1993, 192, 553–560. [Google Scholar] [CrossRef]
- Ichiki, Y.; Kitamura, K.; Kangawa, K.; Kawamoto, M.; Matsuo, H.; Eto, T. Distribution and characterization of immunoreactive adrenomedullin in human tissue and plasma. FEBS Lett. 1994, 338, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Ochoa-Callejero, L.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Iinuma, N.; Arai, T.; Yoshizawa, T.; Iesato, Y.; Lei, Y.; et al. Vascular Endothelial Adrenomedullin-RAMP2 System Is Essential for Vascular Integrity and Organ Homeostasis. Circulation 2013, 127, 842–853. [Google Scholar] [CrossRef]
- Ochoa-Callejero, L.; Pozo-Rodrigálvarez, A.; Martínez-Murillo, R.; Martínez, A. Lack of adrenomedullin in mouse endothelial cells results in defective angiogenesis, enhanced vascular permeability, less metastasis, and more brain damage. Sci. Rep. 2016, 6, 33495. [Google Scholar] [CrossRef]
- Geven, C.; Bergmann, A.; Kox, M.; Pickkers, P. Vascular Effects of Adrenomedullin and the Anti-Adrenomedullin Antibody Adrecizumab in Sepsis. Shock 2018, 50, 132–140. [Google Scholar] [CrossRef]
- López, J.; Martínez, A. Cell and molecular biology of the multifunctional peptide, adrenomedullin. Int. Rev. Cytol. 2002, 221, 1–92. [Google Scholar] [CrossRef]
- Julián-Villaverde, F.; Ochoa-Callejero, L.; Siles, E.; Martínez-Lara, E.; Martínez, A. Adrenomedullin Is a Diagnostic and Prognostic Biomarker for Acute Intracerebral Hemorrhage. Curr. Issues Mol. Biol. 2021, 43, 324–334. [Google Scholar] [CrossRef]
- Oh, S.P.; Seki, T.; Goss, K.A.; Imamura, T.; Yi, Y.; Donahoe, P.K.; Li, L.; Miyazono, K.; Dijke, P.T.; Kim, S.; et al. Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Jerkic, M.; Rivas-Elena, J.V.; Prieto, M.; Carrón, R.; Sanz-Rodríguez, F.; Pérez-Barriocanal, F.; Rodríguez-Barbero, A.; Bernabeu, C.; Lopez-Novoa, J.M. Endoglin regulates nitric oxide-dependent vasodilatation. FASEB J. 2004, 18, 609–611. [Google Scholar] [CrossRef]
- Martínez, A.; Elsasser, T.; Bhathena, S.; Pio, R.; Buchanan, T.; Macri, C.; Cuttitta, F. Is adrenomedullin a causal agent in some cases of type 2 diabetes? Peptides 1999, 20, 1471–1478. [Google Scholar] [CrossRef]
- Ochoa-Callejero, L.; García-Sanmartín, J.; Martínez-Herrero, S.; Rubio-Mediavilla, S.; Narro-Íñiguez, J.; Martínez, A. Small molecules related to adrenomedullin reduce tumor burden in a mouse model of colitis-associated colon cancer. Sci. Rep. 2017, 7, 17488. [Google Scholar] [CrossRef] [Green Version]
- García-Sanmartín, J.; Rubio-Mediavilla, S.; Gallego, J.J.S.; Martínez, A. Glu-tubulin is a marker for Schwann cells and can distinguish between schwannomas and neurofibromas. Histochem. Cell Biol. 2016, 146, 467–477. [Google Scholar] [CrossRef]
- Martínez, A.; Pío, R.; López, J.; Cuttitta, F. Expression of the adrenomedullin binding protein, complement factor H, in the pancreas and its physiological impact on insulin secretion. J. Endocrinol. 2001, 170, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Crowe, A.R.; Yue, W. Semi-quantitative Determination of Protein Expression Using Immunohistochemistry Staining and Analysis: An Integrated Protocol. Bio. Protocol. 2019, 9, e3465. [Google Scholar] [CrossRef]
- Serrano, J.; Uttenthal, L.; Martínez, A.; Fernández, A.P.; de Velasco, J.M.; Alonso, D.; Bentura, M.L.; Santacana, M.; Gallardo, J.R.; Martínez-Murillo, R.; et al. Distribution of adrenomedullin-like immunoreactivity in the rat central nervous system by light and electron microscopy. Brain Res. 2000, 853, 245–268. [Google Scholar] [CrossRef] [Green Version]
- Hinck, A.; Mueller, T.D.; Springer, T.A. Structural Biology and Evolution of the TGF-β Family. Cold Spring Harb. Perspect. Biol. 2016, 8, a022103. [Google Scholar] [CrossRef] [Green Version]
- Tu, L.; Castan, A.; Mallet, C.; Guyon, L.; Cumont, A.; Phan, C.; Robert, F.; Thuillet, R.; Bordenave, J.; Sekine, A.; et al. Selective BMP-9 Inhibition Partially Protects Against Experimental Pulmonary Hypertension. Circ. Res. 2019, 124, 846–855. [Google Scholar] [CrossRef]
- Bodegas, E.; Martínez, A.; Ozbun, L.L.; Garayoa, M.; Letterio, J.J.; Montuenga, L.; Jakowlew, S.B. Depressed adrenomedullin in the embryonic transforming growth factor-beta1 null mouse becomes elevated postnatally. Int. J. Dev. Biol. 2004, 48, 67–70. [Google Scholar] [CrossRef]
- Martínez-Herrero, S.; Larrayoz, I.; Narro-Íñiguez, J.; Villanueva-Millán, M.J.; Recio-Fernández, E.; Pérez-Matute, P.; Oteo, J.A.; Martínez, A. Lack of Adrenomedullin Results in Microbiota Changes and Aggravates Azoxymethane and Dextran Sulfate Sodium-Induced Colitis in Mice. Front. Physiol. 2016, 7, 595. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Herrero, S.; Larrayoz, I.; Narro-Íñiguez, J.; Rubio-Mediavilla, S.; Martínez, A. Lack of Adrenomedullin Aggravates Acute TNBS-Induced Colitis Symptoms in Mice, Especially in Females. Front. Physiol. 2017, 8, 1058. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Herrero, S.; Martínez, A. Adrenomedullin: Not just another gastrointestinal peptide. Biomolecules 2022, 12, 156. [Google Scholar] [CrossRef]
- Nambu, T.; Arai, H.; Komatsu, Y.; Yasoda, A.; Moriyama, K.; Kanamoto, N.; Itoh, H.; Nakao, K. Expression of the adrenomedullin gene in adipose tissue. Regul. Pept. 2005, 132, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Van Der Walt, N.; Pennington, K.A.; Yallampalli, C. Impact of adrenomedullin blockage on lipid metabolism in female mice exposed to high-fat diet. Endocrine 2019, 65, 278–285. [Google Scholar] [CrossRef]
- Dai, H.; Wang, F.; Kang, Y.; Sun, J.; Zhou, H.; Gao, Q.; Li, Z.; Qian, P.; Zhu, G.; Zhou, Y. Adrenomedullin Attenuates Inflammation in White Adipose Tissue of Obese Rats Through Receptor-Mediated PKA Pathway. Obesity 2020, 29, 86–97. [Google Scholar] [CrossRef]
- Yang, M.; Liang, Z.; Yang, M.; Jia, Y.; Yang, G.; He, Y.; Li, X.; Gu, H.F.; Zheng, H.; Zhu, Z.; et al. Role of bone morphogenetic protein-9 in the regulation of glucose and lipid metabolism. FASEB J. 2019, 33, 10077–10088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapitány, A.; Medgyesi, B.; Jenei, A.; Somogyi, O.; Szabó, L.; Gáspár, K.; Méhes, G.; Hendrik, Z.; Dócs, K.; Szücs, P.; et al. Regional Differences in the Permeability Barrier of the Skin—Implications in Acantholytic Skin Diseases. Int. J. Mol. Sci. 2021, 22, 10428. [Google Scholar] [CrossRef]
- Martínez, A.; Elsasser, T.H.; Muro-Cacho, C.; Moody, T.W.; Miller, M.J.; Macri, C.J.; Cuttitta, F. Expression of Adrenomedullin and Its Receptor in Normal and Malignant Human Skin: A Potential Pluripotent Role in the Integument. Endocrinology 1997, 138, 5597–5604. [Google Scholar] [CrossRef] [PubMed]
- Chai, P.; Yu, J.; Wang, X.; Ge, S.; Jia, R. BMP9 promotes cutaneous wound healing by activating Smad1/5 signaling pathways and cytoskeleton remodeling. Clin. Transl. Med. 2021, 11, e271. [Google Scholar] [CrossRef]
- Serrano, J.; Alonso, D.; Fernández, A.; Encinas, J.M.; López, J.; Castro-Blanco, S.; Fernández-Vizarra, P.; Richart, A.; Santacana, M.; Uttenthal, L.; et al. Adrenomedullin in the central nervous system. Microsc. Res. Tech. 2002, 57, 76–90. [Google Scholar] [CrossRef]
- Fernández, A.P.; Serrano, J.; Tessarollo, L.; Cuttitta, F.; Martínez, A. Lack of adrenomedullin in the mouse brain results in behavioral changes, anxiety, and lower survival under stress conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 12581–12586. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.P.; Serrano, J.; Martínez-Murillo, R.; Martínez, A. Lack of Adrenomedullin in the Central Nervous System Results in Apparently Paradoxical Alterations on Pain Sensitivity. Endocrinology 2010, 151, 4908–4915. [Google Scholar] [CrossRef] [Green Version]
- Kapas, S.; Martínez, A.; Cuttitta, F.; Hinson, J.P. Local production and action of adrenomedullin in the rat adrenal zona glomerulosa. J. Endocrinol. 1998, 156, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iring, A.; Jin, Y.-J.; Albarrán-Juárez, J.; Siragusa, M.; Wang, S.; Dancs, P.T.; Nakayama, A.; Tonack, S.; Chen, M.; Künne, C.; et al. Shear stress–induced endothelial adrenomedullin signaling regulates vascular tone and blood pressure. J. Clin. Investig. 2019, 129, 2775–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Ponz, M.; Rodrigo-Gasqué, C.; Siles, E.; Martínez-Lara, E.; Ochoa-Callejero, L.; Martínez, A. Temporal profiles of blood pressure, circulating nitric oxide, and adrenomedullin as predictors of clinical outcome in acute ischemic stroke patients. Mol. Med. Rep. 2016, 13, 3724–3734. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rice, M.; Swist, S.; Kubin, T.; Wu, F.; Wang, S.; Kraut, S.; Weissmann, N.; Böttger, T.; Wheeler, M.; et al. BMP9 and BMP10 Act Directly on Vascular Smooth Muscle Cells for Generation and Maintenance of the Contractile State. Circulation 2021, 143, 1394–1410. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of Interest | Sense Primer | Antisense Primer | Annealing Temp. |
|---|---|---|---|
| Adm | TTGGGTTCACTCGCTTTCCT | TTAGCGCCCACTTATTCCAC | 60 °C |
| Gapdh | CATGTTCCAGTATGACTCCACTC | GGCCTCACCCCATTTGATGT | 60 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Sanmartín, J.; Narro-Íñiguez, J.; Rodríguez-Barbero, A.; Martínez, A. Endoglin and Activin Receptor-like Kinase 1 (Alk1) Modify Adrenomedullin Expression in an Organ-Specific Manner in Mice. Biology 2022, 11, 358. https://doi.org/10.3390/biology11030358
García-Sanmartín J, Narro-Íñiguez J, Rodríguez-Barbero A, Martínez A. Endoglin and Activin Receptor-like Kinase 1 (Alk1) Modify Adrenomedullin Expression in an Organ-Specific Manner in Mice. Biology. 2022; 11(3):358. https://doi.org/10.3390/biology11030358
Chicago/Turabian StyleGarcía-Sanmartín, Josune, Judit Narro-Íñiguez, Alicia Rodríguez-Barbero, and Alfredo Martínez. 2022. "Endoglin and Activin Receptor-like Kinase 1 (Alk1) Modify Adrenomedullin Expression in an Organ-Specific Manner in Mice" Biology 11, no. 3: 358. https://doi.org/10.3390/biology11030358
APA StyleGarcía-Sanmartín, J., Narro-Íñiguez, J., Rodríguez-Barbero, A., & Martínez, A. (2022). Endoglin and Activin Receptor-like Kinase 1 (Alk1) Modify Adrenomedullin Expression in an Organ-Specific Manner in Mice. Biology, 11(3), 358. https://doi.org/10.3390/biology11030358