
Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12




Abstract
:Simple Summary
Abstract
1. Introduction
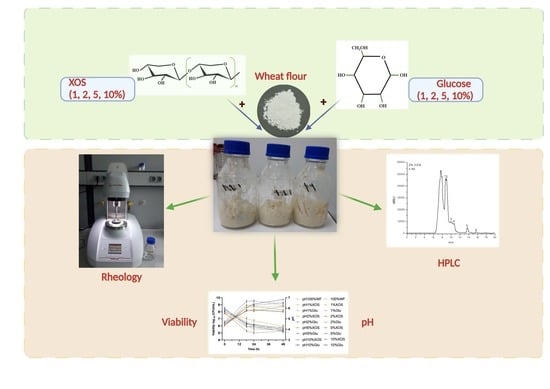
2. Materials and Methods
2.1. Materials and Formulations
2.2. Microorganisms and Culture Environment
2.3. Sourdough Formulation and Fermentation
2.4. pH and Cell Viability
2.5. Secondary Metabolite Analysis by HPLC-RID
2.6. Rheological Measurements
2.7. Statistical Analysis
3. Results and Discussions
3.1. pH and Cell Viability
3.2. Organic Acids and Secondary Metabolites Production
3.3. Rheological Analysis


4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Snigdha, S.; Ha, K.; Tsai, P.; Dinan, T.G.; Bartos, J.D.; Shahid, M. Probiotics: Potential novel therapeutics for microbiota-gut-brain axis dysfunction across gender and lifespan. Pharmacol. Ther. 2022, 231, 107978. [Google Scholar] [CrossRef] [PubMed]
- Plamada, D.; Vodnar, D.C. Polyphenols—Gut Microbiota Interrelationship: A Transition to a New Generation of Prebiotics. Nutrients 2022, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Mitrea, L.; Nemes, S.-A.; Szabo, K.; Teleky, B.-E.; Vodnar, D.-C. Guts Imbalance Imbalances the Brain: A Review of Gut Microbiota Association With Neurological and Psychiatric Disorders. Front. Med. 2022, 9, 1–21. [Google Scholar] [CrossRef]
- Precup, G.; Pocol, C.B.; Teleky, B.-E.; Vodnar, D.C. Awareness, Knowledge, and Interest about Prebiotics—A Study among Romanian Consumers. Int. J. Environ. Res. Public Health 2022, 19, 1208. [Google Scholar] [CrossRef]
- Simon, E.; Călinoiu, L.F.; Mitrea, L.; Vodnar, D.C. Probiotics, prebiotics, and synbiotics: Implications and beneficial effects against irritable bowel syndrome. Nutrients 2021, 13, 2112. [Google Scholar] [CrossRef]
- Shimizu, K.; Ojima, M.; Ogura, H. Gut microbiota and probiotics/synbiotics for modulation of immunity in critically ill patients. Nutrients 2021, 13, 2439. [Google Scholar] [CrossRef]
- Duque, A.L.R.F.; Demarqui, F.M.; Santoni, M.M.; Zanelli, C.F.; Adorno, M.A.T.; Milenkovic, D.; Mesa, V.; Sivieri, K. Effect of probiotic, prebiotic, and synbiotic on the gut microbiota of autistic children using an in vitro gut microbiome model. Food Res. Int. 2021, 149, 110657. [Google Scholar] [CrossRef]
- Vodnar, D.C.; Mitrea, L.; Teleky, B.E.; Szabo, K.; Călinoiu, L.F.; Nemeş, S.A.; Martău, G.A. Coronavirus Disease (COVID-19) Caused by (SARS-CoV-2) Infections: A Real Challenge for Human Gut Microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 575559. [Google Scholar] [CrossRef]
- An, P.; Ji, M.; Ren, H.; Su, J.; Ding, N.S.; Kang, J.; Yin, A.; Zhou, Q.; Shen, L.; Zhao, L.; et al. Prevention of COVID-19 in patients with inflammatory bowel disease in Wuhan, China. Lancet Gastroenterol. Hepatol. 2020, 5, 525–527. [Google Scholar] [CrossRef]
- Pascuta, M.S.; Vodnar, D.C. Nanocarriers for sustainable active packaging: An overview during and post COVID-19. Coatings 2022, 12, 102. [Google Scholar] [CrossRef]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Farias, D.d.P.; de Araújo, F.F.; Neri-Numa, I.A.; Pastore, G.M. Prebiotics: Trends in food, health and technological applications. Trends Food Sci. Technol. 2019, 93, 23–35. [Google Scholar] [CrossRef]
- Precup, G.; Venus, J.; Heiermann, M.; Schneider, R.; Pop, I.D.; Vodnar, D.C. Chemical and Enzymatic Synthesis of Biobased Xylo-Oligosaccharides and Fermentable Sugars from Wheat Straw for Food Applications. Polymers 2022, 14, 1336. [Google Scholar] [CrossRef]
- Lelia, P.O.; Suharoschi, R. Emerging Food Processing Technologies: Probiotics and Prebiotics. In Nutraceutical and Functional Food Components, 2nd ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 509–536. [Google Scholar] [CrossRef]
- Juhász, R.; Penksza, P.; Sipos, L. Effect of xylo-oligosaccharides (XOS) addition on technological and sensory attributes of cookies. Food Sci. Nutr. 2020, 8, 5452–5460. [Google Scholar] [CrossRef]
- Amorim, C.; Cardoso, B.B.; Silvério, S.C.; Silva, J.C.; Alves, J.I.; Pereira, M.A.; Moreira, R.; Rodrigues, L.R. Designing a functional rice muffin formulated with prebiotic oligosaccharides and sugar reduction. Food Biosci. 2021, 40, 100858. [Google Scholar] [CrossRef]
- Palaniappan, A.; Antony, U.; Emmambux, M.N. Current status of xylo-oligosaccharides: Production, characterization, health benefits and food application. Trends Food Sci. Technol. 2021, 111, 506–519. [Google Scholar] [CrossRef]
- Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of xylo-oligosaccharides (XOS) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2018, 16, e05361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, C.A.; Kopit, L.; Doyle, C.; Yu, A.O.; Hugenholtz, J.; Marco, M.L. Polyol production during heterofermentative growth of the plant isolate Lactobacillus florum 2F. J. Appl. Microbiol. 2016, 120, 1336–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christophersen, C.T.; Petersen, A.; Licht, T.R.; Conlon, M.A. Xylo-oligosaccharides and inulin affect genotoxicity and bacterial populations differently in a human colonic simulator challenged with soy protein. Nutrients 2013, 5, 3740–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, V.T.; Calatayud, M.; Rotsaert, C.; Seifert, N.; Richard, N.; Abbeele, P.V.D.; Marzorati, M.; Steinert, R.E. Differentially Shift Microbiota Composition and Function and Improve Intestinal Epithelial Barrier In Vitro. Nutrients 2021, 13, 1125. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.H.; Peck, B.; Terry, D. The influence of probiotics on gastrointestinal tract infections among children attending childcare: A systematic review and meta-analysis. J. Appl. Microbiol. 2021, 132, 1636–1651. [Google Scholar] [CrossRef] [PubMed]
- Lensu, S.; Pariyani, R.; Mäkinen, E.; Yang, B.; Saleem, W.; Munukka, E.; Lehti, M.; Driuchina, A.; Lindén, J.; Tiirola, M.; et al. Prebiotic xylo-oligosaccharides ameliorate high-fat-diet-induced hepatic steatosis in rats. Nutrients 2020, 12, 3225. [Google Scholar] [CrossRef]
- Vodnar, D.C.; Calinoiu, L.-F.; Mitrea, L.; Precup, G.; Bindea, M.; Pacurar, A.M.; Szabo, K.; Stefanescu, B.E. A new generation of probiotic functional beverages using bioactive compounds from agro-industrial waste. In Functional and Medicinal Beverages; Academic Press: Cambridge, MA, USA, 2019; Volume 11, pp. 483–528. ISBN 9780128163979. [Google Scholar]
- Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.E.; Stuer-Lauridsen, B.; Eskesen, D. The science behind the probiotic strain bifidobacterium animalis subsp. Lactis bb-12®. Microorganisms 2014, 2, 92–110. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://isappscience.org/for-scientists/resources/probiotics/ (accessed on 31 March 2022).
- Martau, G.A.; Teleky, B.-E.; Ranga, F.; Pop, I.D.; Vodnar, D.C. Apple Pomace as a Sustainable Substrate in Sourdough Fermentation. Front. Microbiol. 2021, 12, 742020. [Google Scholar] [CrossRef] [PubMed]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa sourdough fermented with lactobacillus plantarum ATCC 8014 designed for gluten-free muffins—A powerful tool to enhance bioactive compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Ashwini, A.; Ramya, H.N.; Ramkumar, C.; Reddy, K.R.; Kulkarni, R.V.; Abinaya, V.; Naveen, S.; Raghu, A.V. Reactive mechanism and the applications of bioactive prebiotics for human health: Review. J. Microbiol. Methods 2019, 159, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Duboux, S.; Van Wijchen, M.; Kleerebezem, M. The Possible Link Between Manufacturing and Probiotic Efficacy; a Molecular Point of View on Bifidobacterium. Front. Microbiol. 2021, 12, 812536. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.P.d.S.; Perego, P.; de Oliveira, M.N.; Converti, A. Growth, organic acids profile and sugar metabolism of Bifidobacterium lactis in co-culture with Streptococcus thermophilus: The inulin effect. Food Res. Int. 2012, 48, 21–27. [Google Scholar] [CrossRef]
- Mäkeläinen, H.; Forssten, S.; Saarinen, M.; Stowell, J.; Rautonen, N.; Ouwehand, A.C. Xylo-oligosaccharides enhance the growth of bifidobacteria and Bifidobacterium lactis in a simulated colon model. Benef. Microbes 2010, 1, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Hesam, F.; Tarzi, B.G.; Honarvar, M.; Jahadi, M. Valorization of sugarcane bagasse to high value-added xylo-oligosaccharides and evaluation of their prebiotic function in a synbiotic pomegranate juice. Biomass Convers. Biorefinery 2020. [Google Scholar] [CrossRef]
- Cho, S.S. The Generally Recognized as Safe [GRAS] Determination of Bifidobacterium Animalis subsp. lactis AD011 (B. lactis AD011) as a Food Ingredient; BIFIDO CO., LTD.: Hongcheon, Korea, 2020. [Google Scholar]
- Cronin, M.; Ventura, M.; Fitzgerald, G.F.; Van Sinderen, D. Progress in genomics, metabolism and biotechnology of bifidobacteria. Int. J. Food Microbiol. 2011, 149, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Penksza, P.; Juhász, R.; Szabó-Nótin, B.; Sipos, L. Xylo-oligosaccharides as texture modifier compounds in aqueous media and in combination with food thickeners. Food Sci. Nutr. 2020, 8, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Huang, J.S.; Xie, Q.T.; Zou, Y.M.; Wang, H.L.; Wu, X.Y.; Xu, X.M. Effect of multiple freezing-thawing cycles on structural and functional properties of starch granules isolated from soft and hard wheat. Food Chem. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Wang, P.; Wu, F.; Jin, Z.; Xu, X. Particle size distribution of wheat starch granules in relation to baking properties of frozen dough. Carbohydr. Polym. 2016, 137, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Park, E.Y.; Jang, S.B.; Lim, S.T. Effect of fructo-oligosaccharide and isomalto-oligosaccharide addition on baking quality of frozen dough. Food Chem. 2016, 213, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Tu, J.; Zheng, M.; Deng, K.; Miao, S.; Zeng, S.; Zheng, B.; Lu, X. Effects of oligosaccharides on particle structure, pasting and thermal properties of wheat starch granules under different freezing temperatures. Food Chem. 2020, 315, 126209. [Google Scholar] [CrossRef] [PubMed]
- Chiș, M.S.; Păucean, A.; Stan, L.; Mureșan, V.; Vlaic, R.A.; Man, S.; Biriș-Dorhoi, E.S.; Muste, S. Lactobacillus plantarum ATCC 8014 in quinoa sourdough adaptability and antioxidant potential. Rom. Biotechnol. Lett. 2018, 23, 13581–13591. [Google Scholar]
- Teleky, B.-E.; Martău, G.-A.; Vodnar, D.-C. Physicochemical Effects of Lactobacillus plantarum and Lactobacillus casei Cocultures on Soy–Wheat Flour Dough Fermentation. Foods 2020, 9, 1894. [Google Scholar] [CrossRef] [PubMed]
- Le Bleis, F.; Chaunier, L.; Chiron, H.; Della Valle, G.; Saulnier, L. Rheological properties of wheat flour dough and French bread enriched with wheat bran. J. Cereal Sci. 2015, 65, 167–174. [Google Scholar] [CrossRef]
- Simpson, P.J.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Intrinsic tolerance of Bifidobacterium species to heat and oxygen and survival following spray drying and storage. J. Appl. Microbiol. 2005, 99, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Śliżewska, K.; Chlebicz-Wójcik, A. Growth kinetics of probiotic lactobacillus strains in the alternative, cost-efficient semi-solid fermentation medium. Biology 2020, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Teleky, B.E.; Martău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C.; Gheorghe, A.; Chet, F. Exploitation of lactic acid bacteria and Baker’s yeast as single or multiple starter cultures of wheat flour dough enriched with soy flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef]
- Mitrea, L.; Trif, M.; Vodnar, D.-C.; Laura, M.; Monica, T.; Dan-Cristian, V. The effect of crude glycerol impurities on 1,3-propanediol biosynthesis by Klebsiella pneumoniae DSMZ 2026. Renew. Energy 2020, 153, 1418–1427. [Google Scholar] [CrossRef]
- Mitrea, L.; Teleky, B.-E.; Leopold, L.-F.; Nemes, S.-A.; Plamada, D.; Dulf, F.V.; Pop, I.-D.; Vodnar, D.C. The physicochemical properties of five vegetable oils exposed at high temperature for a short-time-interval. J. Food Compos. Anal. 2022, 106, 104305. [Google Scholar] [CrossRef]
- Stefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crisan, G. The Chemical and Biological Profiles of Leaves from Commercial Blueberry Varieties. Plants 2020, 9, 1193. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Teleky, B.-E.; Ranga, F.; Simon, E.; Pop, O.L.; Babalau-Fuss, V.; Kapsalis, N.; Vodnar, D.C. Bioaccessibility of microencapsulated carotenoids, recovered from tomato processing industrial by-products, using in vitro digestion model. LWT—Food Sci. Technol. 2021, 152, 112285. [Google Scholar] [CrossRef]
- Guo, L.; Fang, F.; Zhang, Y.; Xu, D.; Jin, Z.; Xu, X. Glutathione affects rheology and water distribution of wheat dough by changing gluten conformation and protein depolymerisation. Int. J. Food Sci. Technol. 2020, 56, 3157–3165. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Gholamhosseinpour, A.; Mousavi Khaneghah, A. Fermentation of acorn dough by lactobacilli strains: Phytic acid degradation and antioxidant activity. LWT—Food Sci. Technol. 2019, 100, 144–149. [Google Scholar] [CrossRef]
- Finegold, S.M.; Li, Z.; Summanen, P.H.; Downes, J.; Thames, G.; Corbett, K.; Dowd, S.; Krak, M.; Heber, D. Xylooligosaccharide increases bifidobacteria but not lactobacilli in human gut microbiota. Food Funct. 2014, 5, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Fujikawa, S.; Matsumoto, N. Effect of Xylooligosaccharide on the Growth of Bifidobacteria. Bifidobact. Microflora 1990, 9, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Van Der Meulen, R.; Adriany, T.; Verbrugghe, K.; De Vuyst, L. Kinetic analysis of bifidobacterial metabolism reveals a minor role for succinic acid in the regeneration of NAD+ through its growth-associated production. Appl. Environ. Microbiol. 2006, 72, 5204–5210. [Google Scholar] [CrossRef] [Green Version]
- Paucean, A.; Vodnar, D.C.; Socaci, S.A.; Socaciu, C. Carbohydrate metabolic conversions to lactic acid and volatile derivatives, as influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 efficiency during in vitro and sourdough fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Gioulatos, S.; Tsakalidou, E.; Kalantzopoulos, G. Interactions between Saccharomyces cerevisiae and lactic acid bacteria in sourdough. Process Biochem. 2006, 41, 2429–2433. [Google Scholar] [CrossRef]
- Dysvik, A.; La Rosa, S.L.; Buffetto, F.; Liland, K.H.; Myhrer, K.S.; Rukke, E.O.; Wicklund, T.; Westereng, B. Secondary Lactic Acid Bacteria Fermentation with Wood-Derived Xylo-oligosaccharides as a Tool to Expedite Sour Beer Production. J. Agric. Food Chem. 2020, 68, 301–314. [Google Scholar] [CrossRef]
- Kim, M.J.; Yoo, S.H.; Jung, S.; Park, M.K.; Hong, J.H. Relative sweetness, sweetness quality, and temporal profile of xylo-oligosaccharides and luo han guo (Siraitia grosvenorii) extract. Food Sci. Biotechnol. 2015, 24, 965–973. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Zhang, Y.; Yang, W.; Ma, G.; Ma, N.; Hu, Q.; Pei, F. A novel lactic acid bacterium for improving the quality and shelf life of whole wheat bread. Food Control 2020, 109, 106914. [Google Scholar] [CrossRef]
- Aachary, A.A.; Prapulla, S.G. Xylo-oligosaccharides (XOS) as an Emerging Prebiotic: Microbial Synthesis, Utilization, Structural Characterization, Bioactive Properties, and Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Albano, K.M.; Franco, C.M.L.; Telis, V.R.N. Rheological behavior of Peruvian carrot starch gels as affected by temperature and concentration. Food Hydrocoll. 2014, 40, 30–43. [Google Scholar] [CrossRef]
- Iuga, M.; Boestean, O.; Ghendov-Mosanu, A.; Mironeasa, S. Impact of Dairy Ingredients on Wheat Flour Dough Rheology and Bread Properties. Foods 2020, 9, 828. [Google Scholar] [CrossRef] [PubMed]
- Nammakuna, N.; Barringer, S.A.; Ratanatriwong, P. The effects of protein isolates and hydrocolloids complexes on dough rheology, physicochemical properties and qualities of gluten-free crackers. Food Sci. Nutr. 2016, 4, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Ding, B.; Ding, W.; Xiao, S.; Song, J. Effect of hydrocolloids on physical, thermal and microstructure properties of par-baked baguette during frozen storage. Int. J. Biol. Macromol. 2020, 163, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Tong, J.; Ding, M.; Wang, K.; Wang, L.; Cheng, D.; Li, H.; Liu, A.; Liu, J.; Zhao, Z.; et al. Physicochemical properties of starch in relation to rheological properties of wheat dough (Triticum aestivum L.). Food Chem. 2019, 297, 125000. [Google Scholar] [CrossRef]
- Wang, P.; Jin, Z.; Xu, X. Physicochemical alterations of wheat gluten proteins upon dough formation and frozen—A review from gluten, glutenin and gliadin perspectives. Trends Food Sci. Technol. 2015, 46, 189–198. [Google Scholar] [CrossRef]
- Ferrero, C. Hydrocolloids in wheat breadmaking: A concise review. Food Hydrocoll. 2017, 68, 15–22. [Google Scholar] [CrossRef]
- Nuzzo, D.; Picone, P.; Sanchez, J.L.; Borras-linares, I.; Guiducci, A.; Muscolino, E.; Giacomazza, D.; Sanfilippo, T.; Guggino, R.; Bulone, D.; et al. Recovery from Food Waste—Biscuit Doughs Enriched with Pomegranate Peel Powder as a Model of Fortified Aliment. Biology 2022, 11, 416. [Google Scholar] [CrossRef]
- Mellado-Mojica, E.; Seeram, N.P.; López, M.G. Comparative analysis of maple syrups and natural sweeteners: Carbohydrates composition and classification (differentiation) by HPAEC-PAD and FTIR spectroscopy-chemometrics. J. Food Compos. Anal. 2016, 52, 1–8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | 100% WF | 1% Glu | 2% Glu | 5% Glu | 10% Glu | 1% XOS | 2% XOS | 5% XOS | 10% XOS | |
|---|---|---|---|---|---|---|---|---|---|---|
| Glucose | 0 | 2.04 ± 0.14 c | 2.93 ± 0.14 c | 7.25 ± 0.22 bc | 11.38 ± 0.16 b | 29.16 ± 0.32 a | 1.24 ± 0.08 cd | 0.31 ± 0.04 d | 0.32 ± 0.06 d | 0.15 ± 0.03 d |
| 18 | 0.59 ± 0.06 d | 2.61 ± 0.13 c | 4.05 ± 0.19 b | 11.26 ± 0.25 b | 42.36 ± 0.29 a | 0.33 ± 0.05 d | 0.21 ± 0.02 d | 0.19 ± 0.02 d | 0.06 ± 0.01 d | |
| 24 | 0.20 ± 0.07 d | 1.66 ± 0.09 c | 4.03 ± 0.16 b,c | 12.03 ± 0.24 b | 41.63 ± 0.36 a | 0.21 ± 0.03 d | 0.21 ± 0.02 d | 0.16 ± 0.02 d | 0.09 ± 0.01 d | |
| 48 | 0.63 ± 0.09 e | 1.84 ± 0.09 d | 4.36 ± 0.14 c | 9.59 ± 0.14 b | 33.20 ± 0.35 a | 0.17 ± 0.02 e | 0.17 ± 0.01 e | N.D. | N.D. | |
| Maltose | 0 | 0.41 ± 0.04 d | 5.69 ± 0.15 a,b | 3.59 ± 0.21 b | 2.14 ± 0.16 b,c | 1.32 ± 0.10 c | 5.13 ± 0.12 a,b | 2.18 ± 0.09 b,c | 1.21 ± 0.10 b | 6.59 ± 0.24 a |
| 18 | 10.11 ± 0.11 a | 4.41 ± 0.12 c,d | 4.58 ± 0.19 c,d | 2.64 ± 0.12 d,e | 1.86 ± 0.13 e | 7.75 ± 0.14 b | 1.93 ± 0.11 c | 1.35 ± 0.09 e | 2.23 ± 0.12 d,e | |
| 24 | 6.71 ± 0.23 a,b | 2.11 ± 0.09 c | 4.83 ± 0.14 b | 2.89 ± 0.10 c | 1.85 ± 0.09 c | 7.97 ± 0.21 a | 1.27 ± 0.08 c | 0.92 ± 0.05 c | 2.39 ± 0.10 c | |
| 48 | 10.37 ± 0.19 a | 0.45 ± 0.06 d | 6.01 ± 0.18 b | 2.41 ± 0.10 c | 1.09 ± 0.06 d | 9.72 ± 0.10 a | 0.87 ± 0.05 d | 0.67 ± 0.04 d | 0.18 ± 0.08 d | |
| Lactic acid | 0 | 0.01 ± 0.00 a | 0.06 ± 0.00 a | 0.06 ± 0.01 a | 0.09 ± 0.01 a | 0.03 ± 0.00 a | 0.02 ± 0.00 a | 0.12 ± 0.01 a | 0.01 ± 0.00 a | 0.09 ± 0.01 a |
| 18 | 0.53 ± 0.02 a,b | 0.83 ± 0.06 a | 0.43 ± 0.05 b | 0.37 ± 0.03 b | 0.51 ± 0.04 | 0.76 ± 0.04 a | 0.24 ± 0.01 b | 0.56 ± 0.04 a,b | 1.07 ± 0.09 a | |
| 24 | 0.74 ± 0.03 b | 0.93 ± 0.08 a,b | 0.52 ± 0.05 b | 0.51 ± 0.05 b | 0.66 ± 0.05 b | 1.08 ± 0.10 | 0.23 ± 0.02 c | 0.90 ± 0.07 a,b | 1.44 ± 0.11 a | |
| 48 | 1.98 ± 0.07 a | 0.95 ± 0.10 b | 1.22 ± 0.08 a,b | 0.74 ± 0.06 b,c | 1.24 ± 0.10 a,b | 1.50 ± 0.10 a | 0.30 ± 0.03 c | 1.17 ± 0.09 a,b | 1.28 ± 0.11 a,b |
| Time (h) | 100% WF | 1% XOS | 2% XOS | 5% XOS | 10% XOS | |
|---|---|---|---|---|---|---|
| Acetic acid | 0 | N.D. | N.D. | N.D. | N.D. | 0.16 ± 0.01 |
| 18 | N.D. | N.D. | 0.11 ± 0.01 b | 0.26 ± 0.02 b | 0.77 ± 0.07 a | |
| 24 | N.D. | 0.12 ± 0.01 b | 0.12 ± 0.01 b | 0.40 ± 0.03 b | 0.95 ± 0.08 a | |
| 48 | 0.19 ± 0.01 b | 0.22 ± 0.02 b | 0.16 ± 0.02 b | 0.59 ± 0.04 a,b | 0.87 ± 0.03 a |
| Time (h) | 1% XOS | 2% XOS | 5% XOS | 10% XOS | |
|---|---|---|---|---|---|
| XOS | 0 | 0.04 ± 0.00 d | 2.78 ± 0.08 c | 5.83 ± 0.09 b | 13.08 ± 0.15 a |
| 18 | N.D. | 2.92 ± 0.06 c | 5.11 ± 0.11 b,c | 17.29 ± 0.18 a | |
| 24 | N.D. | 2.44 ± 0.09 c | 4.24 ± 0.08 b | 14.43 ± 0.12 a | |
| 48 | N.D. | 2.72 ± 0.04 b | 2.67 ± 0.06 b | 5.93 ± 0.11 a | |
| Xylose | 0 | 0.75 ± 0.04 a | 0.52 ± 0.04 a | 0.40 ± 0.02 a | 0.746 ± 0.06 a |
| 18 | 0.58 ± 0.02 b | 0.28 ± 0.02 b | 0.45 ± 0.02 b | 1.207 ± 0.06 a | |
| 24 | 0.78 ± 0.04 a,b | 0.25 ± 0.02 c | 0.64 ± 0.03 b | 1.20 ± 0.02 a | |
| 48 | 1.12 ± 0.06 a,b | 0.29 ± 0.02 b | 1.03 ± 0.04 a,b | 1.62 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Precup, G.; Teleky, B.-E.; Ranga, F.; Vodnar, D.C. Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12. Biology 2022, 11, 553. https://doi.org/10.3390/biology11040553
Precup G, Teleky B-E, Ranga F, Vodnar DC. Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12. Biology. 2022; 11(4):553. https://doi.org/10.3390/biology11040553
Chicago/Turabian StylePrecup, Gabriela, Bernadette-Emőke Teleky, Floricuța Ranga, and Dan Cristian Vodnar. 2022. "Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12" Biology 11, no. 4: 553. https://doi.org/10.3390/biology11040553
APA StylePrecup, G., Teleky, B.-E., Ranga, F., & Vodnar, D. C. (2022). Assessment of Physicochemical and Rheological Properties of Xylo-Oligosaccharides and Glucose-Enriched Doughs Fermented with BB-12. Biology, 11(4), 553. https://doi.org/10.3390/biology11040553