
The Preyssler-Type Polyoxotungstate Exhibits Anti-Quorum Sensing, Antibiofilm, and Antiviral Activities

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
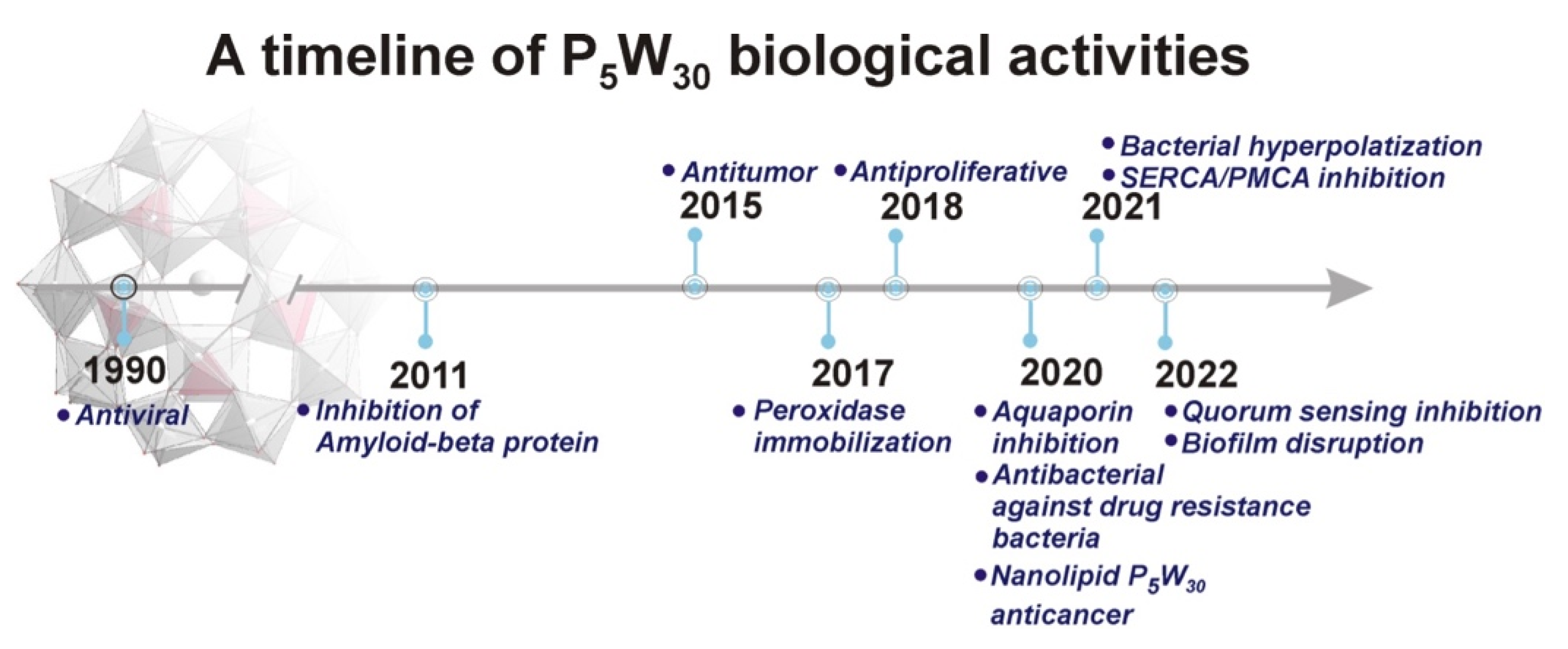
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Polyoxometalates
NMR Spectroscopy
2.3. Screening of Antibacterial Activity by Agar Diffusion
Determination of the Minimum Inhibitory Concentration
- Equation (1):
- Equation (2):The ƩFIC was calculated according to the equation ƩFIC = FIC(P5W30) + FIC(antibiotic)
2.4. Inhibition of Quorum Sensing
2.5. Inhibition of Biofilm Formation
2.6. Antiviral Activity
2.7. Transmission Electron Microscopy
2.8. Statistical Analysis
3. Results and Discussion
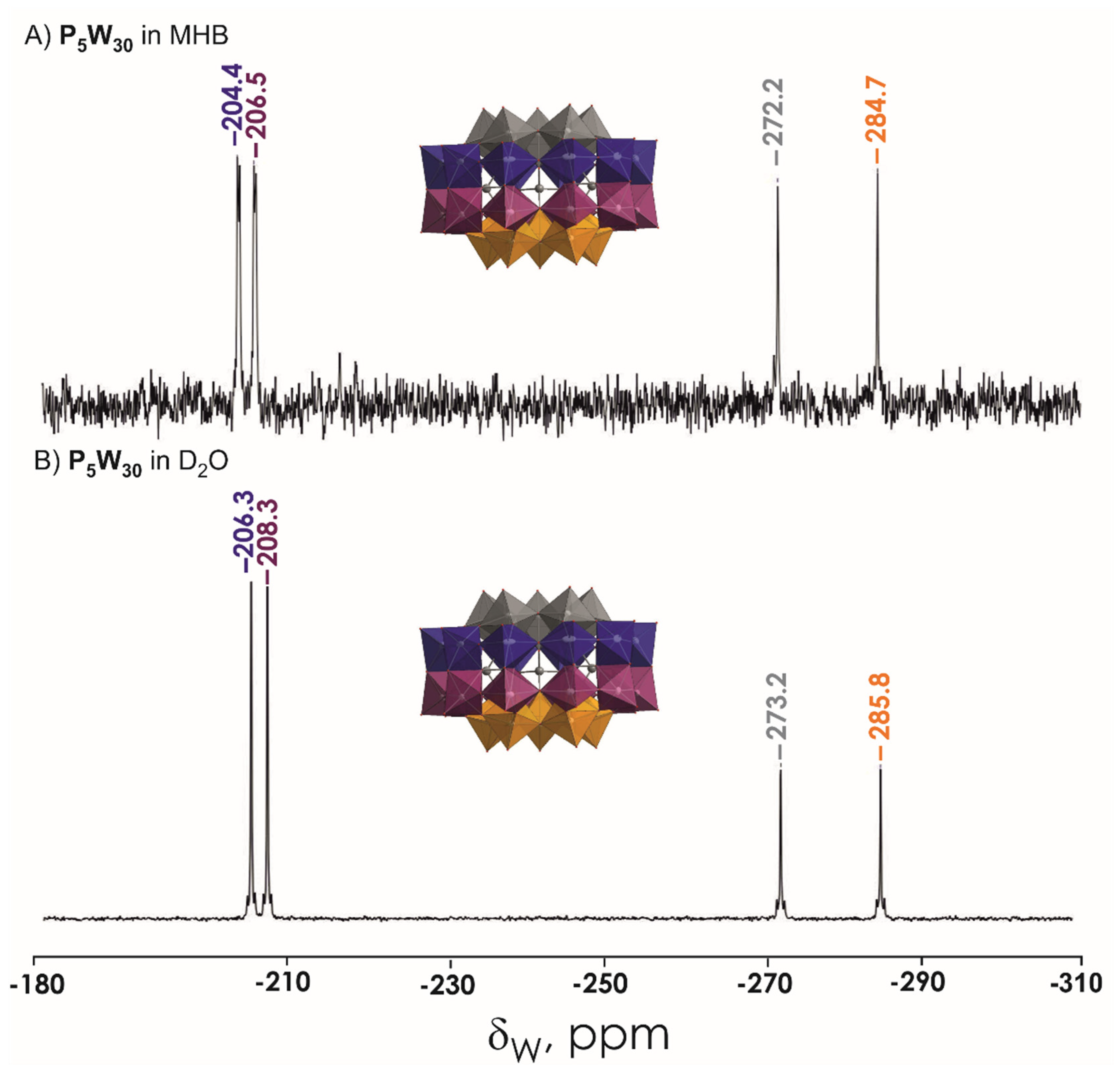
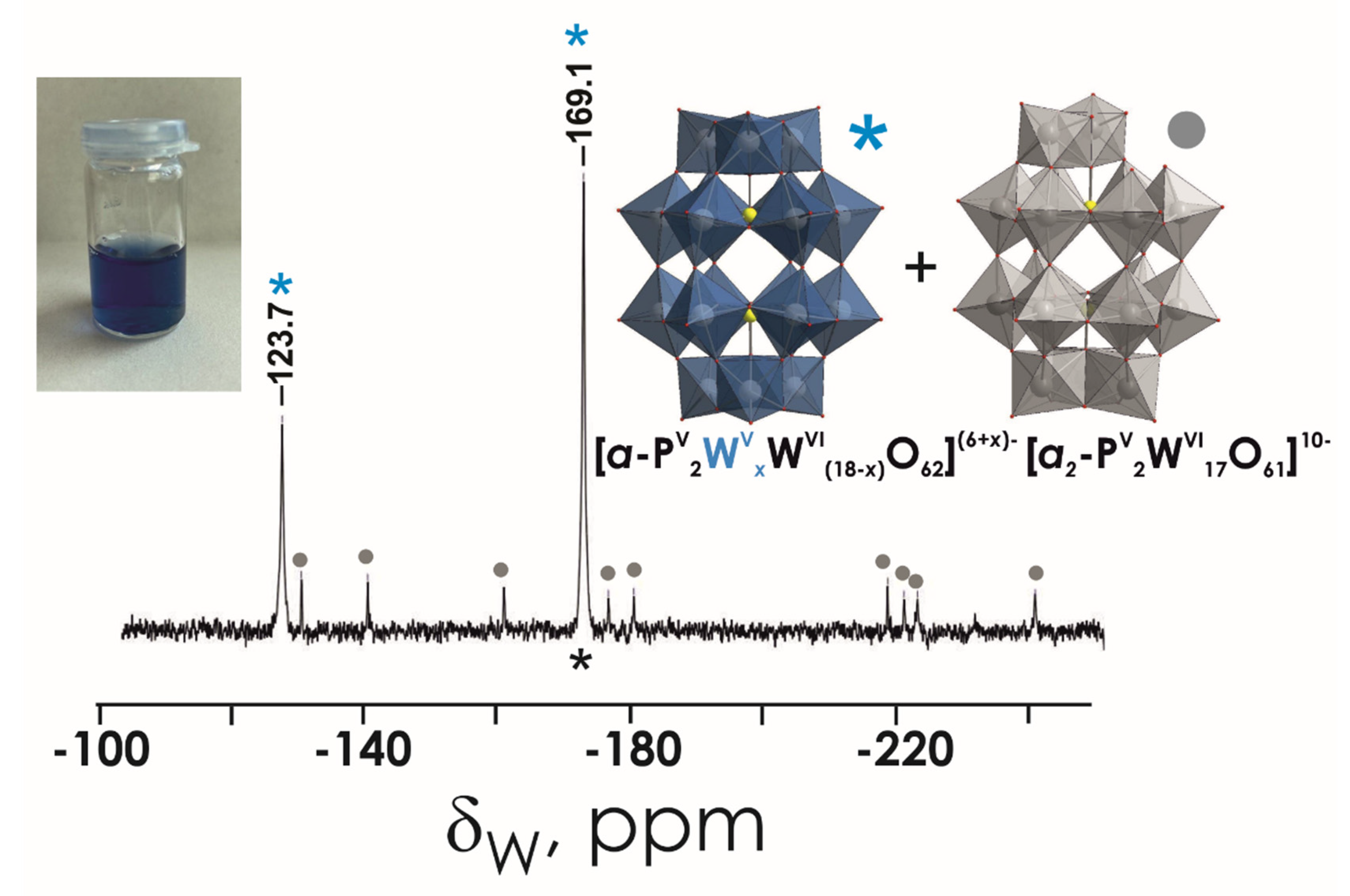
3.1. Stability Studies
3.2. Antibacterial Studies
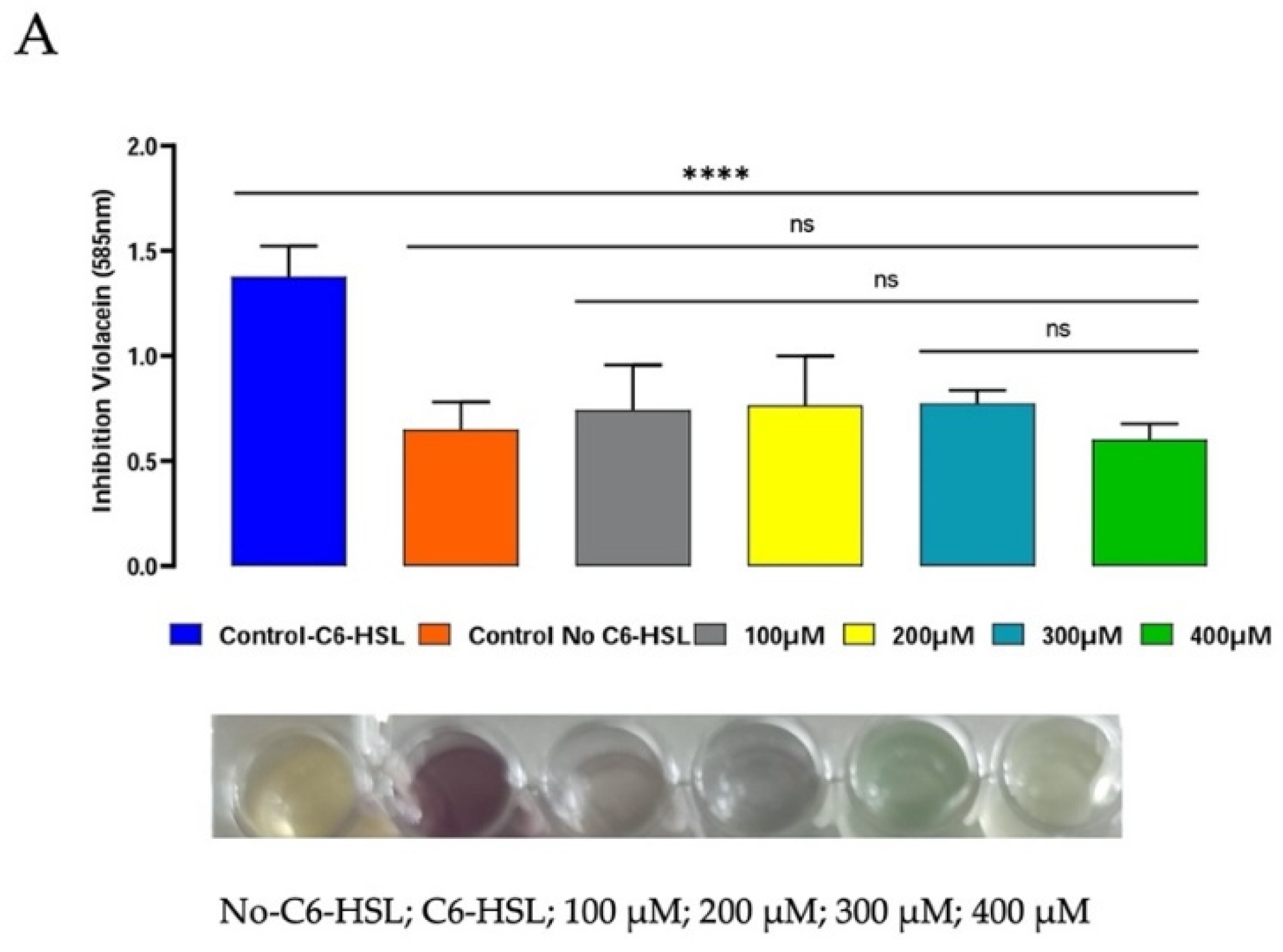
3.3. Anti-Quorum-Sensing Activity
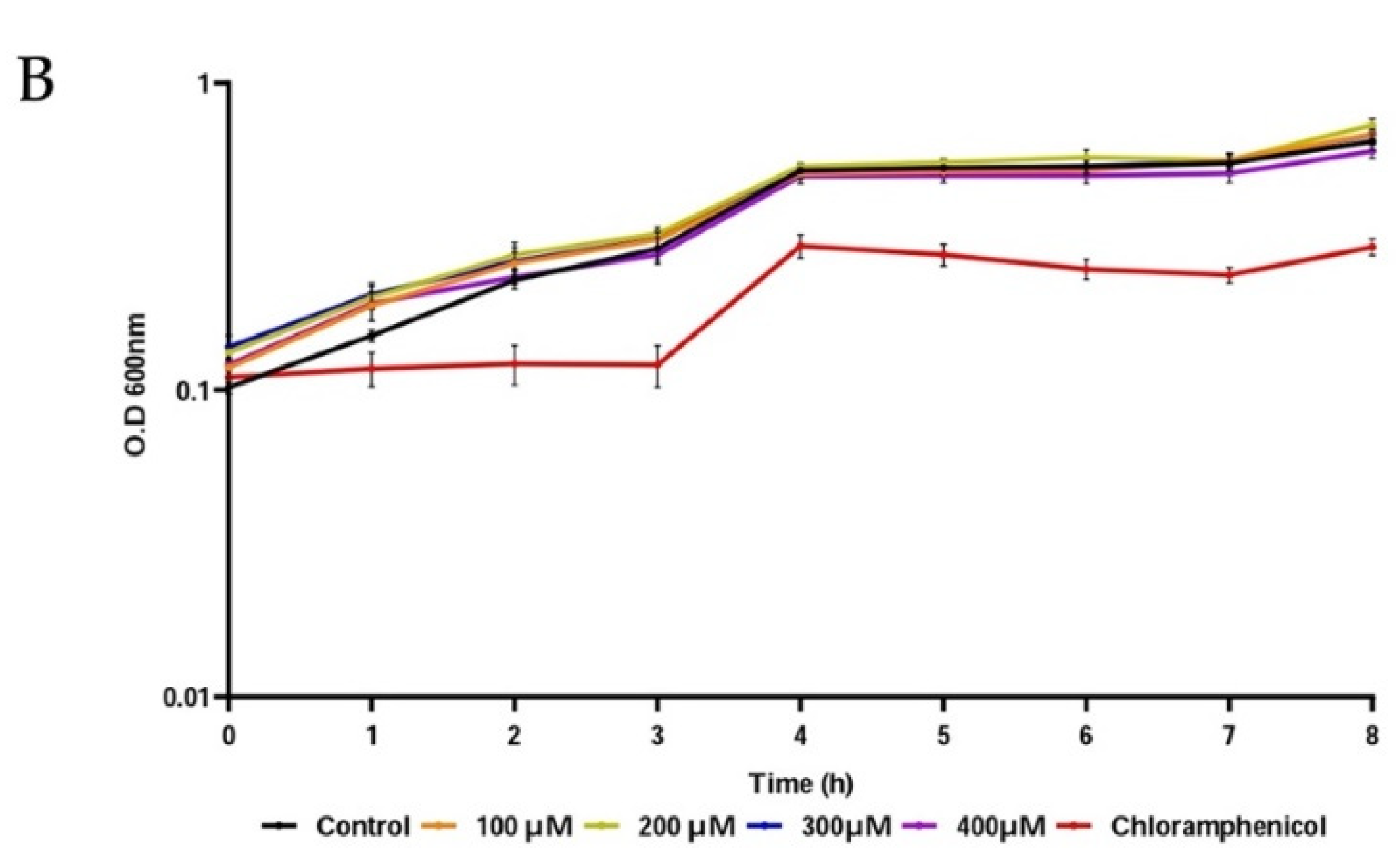
3.4. Antibiofilm Activity
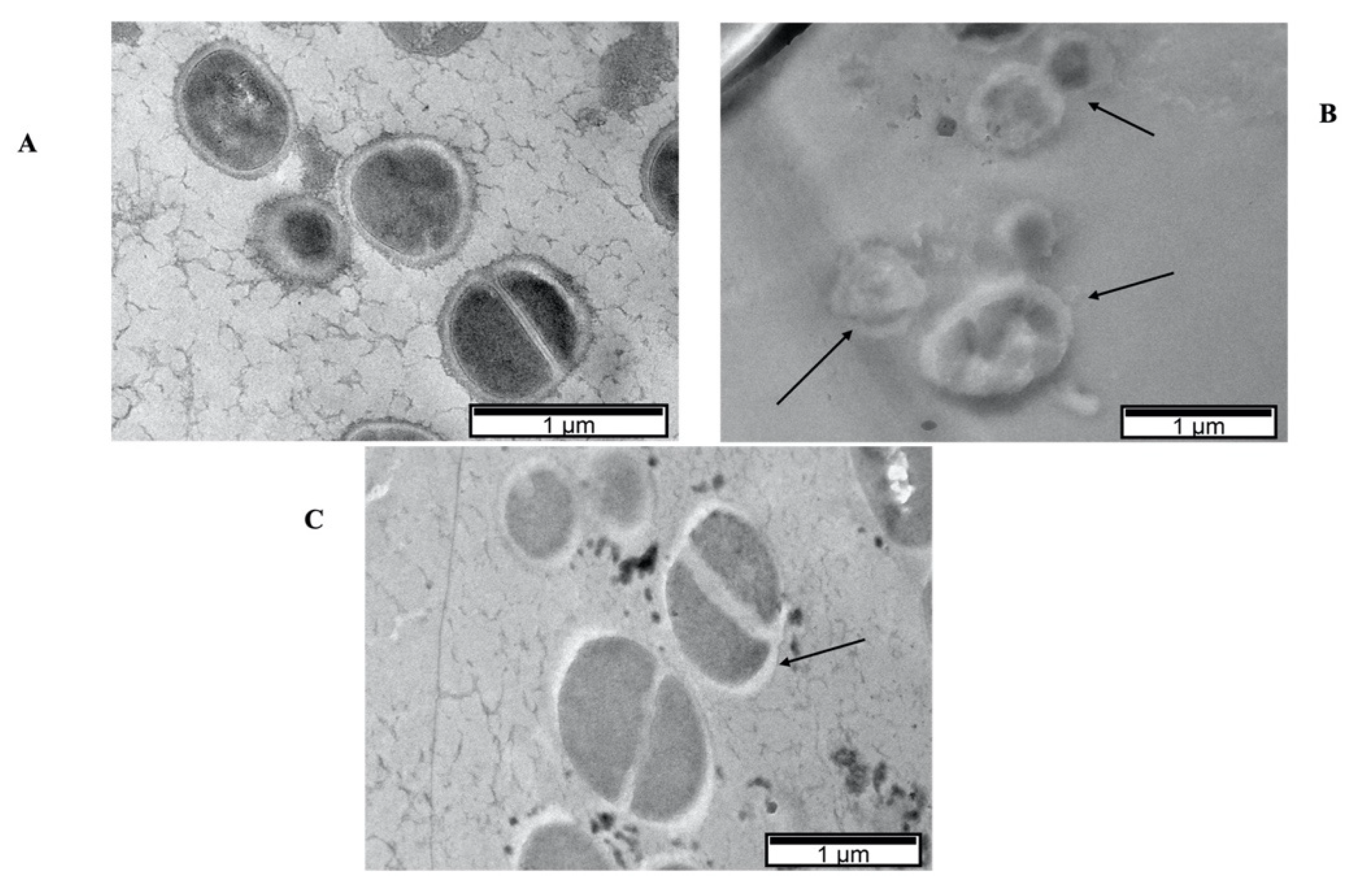
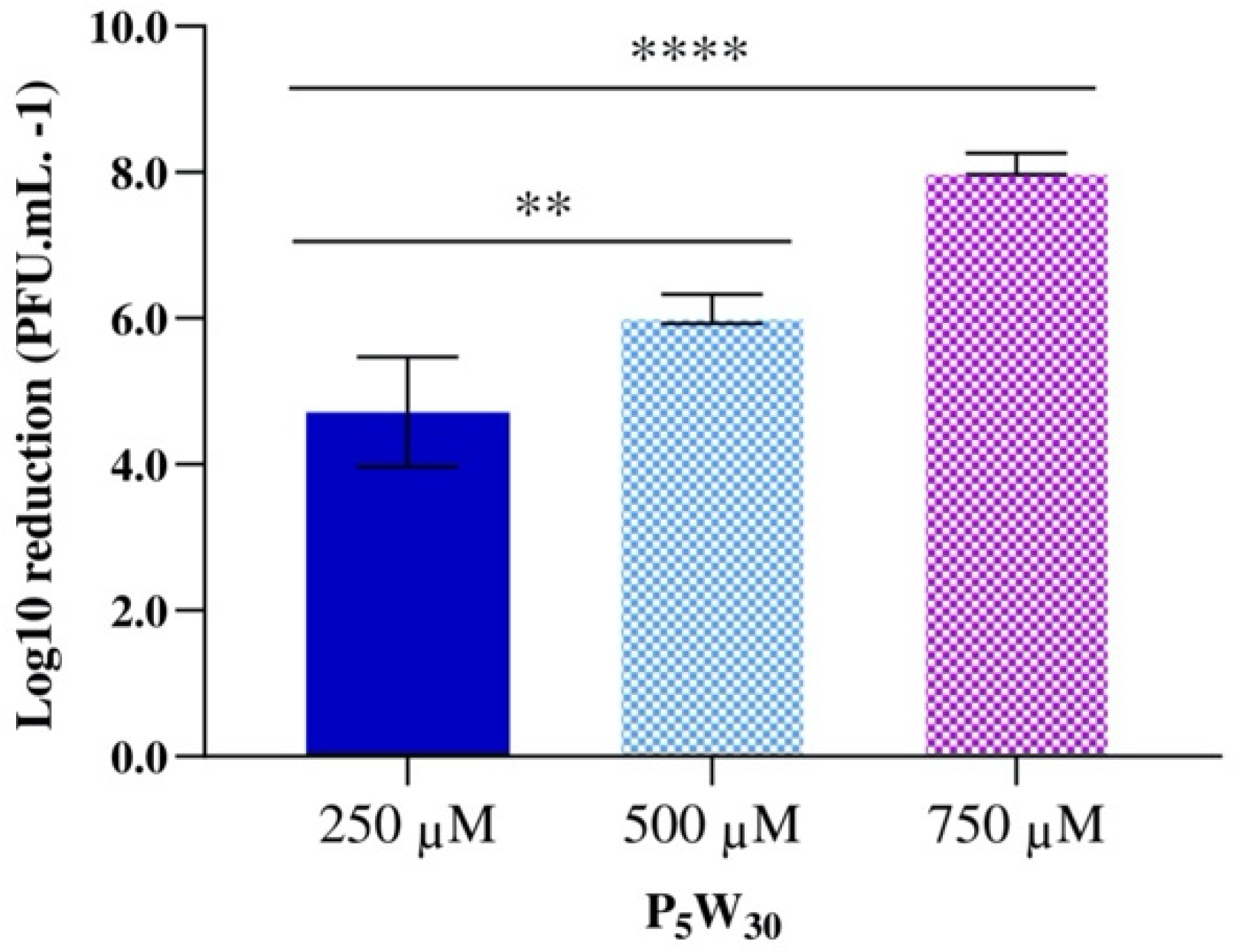
3.5. Antiviral Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, J. Origins and evolution of antibiotic resistance. Microbiologia 1996, 12, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Gudda, F.O.; Waigi, M.G.; Odinga, E.S.; Yang, B.; Carter, L.; Gao, Y. Antibiotic-contaminated wastewater irrigated vegetables pose resistance selection risks to the gut microbiome. Environ. Pollut. 2020, 264, 114752. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Hernandez, A.L.; DePas, W.H.; Park, J.H.; Teschler, J.K.; Hartmann, R.; Jeckel, H.; Drescher, K.; Beyhane, S.; Newman, D.K.; Yildiz, F.H. Upregulation of virulence genes promotes Vibrio cholerae biofilm hyperinfectivity. Proc. Natl. Acad. Sci. USA 2020, 117, 11010–11017. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Miquel, S.; Lagrafeuille, R.; Souweine, B.; Forestier, C. Anti-biofilm activity as a health issue. Front. Microbiol. 2016, 7, 592. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; Angle, A.; Aldrich, A.; Williams, S.H.; Engebretsen, I.L.; Bayles, K.W.; Horswill, A.R.; Kielian, T. Staphylococcus aureus Biofilms Prevent Macrophage Phagocytosis and Attenuate Inflammation In Vivo. J. Immunol. 2011, 186, 6585–6596. [Google Scholar] [CrossRef] [Green Version]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-Associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Aframian, N.; Eldar, A. A Bacterial Tower of Babel: Quorum-Sensing Signaling Diversity and Its Evolution. Annu. Rev. Microbiol. 2020, 74, 587–606. [Google Scholar] [CrossRef] [PubMed]
- Ampomah-Wireko, M.; Luo, C.; Cao, Y.; Wang, H.; Nininahazwe, L.; Wu, C. Chemical probe of AHL modulators on quorum sensing in Gram-Negative Bacteria and as antiproliferative agents: A review. Eur. J. Med. Chem. 2021, 226, 113864. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Rewak-Soroczyńska, J.; Jędrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of biofilm formation by quorum quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Ding, Y.; Cao, J.; Sun, Y.; Wang, F.; Ju, S.; Yu, J. Non-antibiotic methods against Pseudomonas aeruginosa include QS inhibitors: A narrative review. Ann. Palliat. Med. 2021, 10, 6926–6935. [Google Scholar] [CrossRef]
- Chadha, J.; Harjai, K.; Chhibber, S. Repurposing phytochemicals as anti-virulent agents to attenuate quorum sensing-regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Microb. Biotechnol. 2021, 15, 1695–1718. [Google Scholar] [CrossRef] [PubMed]
- Anju, V.T.; Busi, S.; Ranganathan, S.; Ampasala, D.R.; Kumar, S.; Suchiang, K.; Kumavath, R.; Dyavaiah, M. Sesamin and sesamolin rescues Caenorhabditis elegans from Pseudomonas aeruginosa infection through the attenuation of quorum sensing regulated virulence factors. Microb. Pathog. 2021, 155, 104912. [Google Scholar] [CrossRef]
- Horváth, M.; Kovács, T.; Koderivalappil, S.; Ábrahám, H.; Rákhely, G.; Schneider, G. Identification of a newly isolated lytic bacteriophage against K24 capsular type, carbapenem resistant Klebsiella pneumoniae isolates. Sci. Rep. 2020, 10, 5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, J.; Melo, L.D.R.; Poeta, P.; Igrejas, G.; Ferraz, M.P.; Azeredo, J.; Monteiro, F.J. Lytic bacteriophages against multidrug-resistant Staphylococcus aureus, Enterococcus faecalis and Escherichia coli isolates from orthopaedic implant-associated infections. Int. J. Antimicrob. Agents 2019, 54, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Garza-Cervantes, J.A.; Mendiola-Garza, G.; de Melo, E.M.; Dugmore, T.I.J.; Matharu, A.S.; Morones-Ramirez, J.R. Antimicrobial activity of a silver-microfibrillated cellulose biocomposite against susceptible and resistant bacteria. Sci. Rep. 2020, 10, 7281. [Google Scholar] [CrossRef]
- García-Lara, B.; Saucedo-Mora, M.A.; Roldán-Sánchez, J.A.; Pérez-Eretza, B.; Ramasamy, M.; Lee, J.; Coria-Jimenez, R.; Tapia, M.; Varela-Guerrero, V.; García-Contreras, R. Inhibition of quorum-sensing-dependent virulence factors and biofilm formation of clinical and environmental Pseudomonas aeruginosa strains by ZnO nanoparticles. Lett. Appl. Microbiol. 2015, 61, 299–305. [Google Scholar] [CrossRef]
- Rao, H.; Choo, S.; Mahalingam, S.R.R.; Adisuri, D.S.; Madhavan, P.; Akim, A.M.; Chong, P.P. Approaches for mitigating microbial biofilm-related drug resistance: A focus on micro- and nanotechnologies. Molecules 2021, 26, 1870. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Suzuki, T.; Fujita, Y.; Oda, M.; Matsumoto, N.; Yamase, T. Enhancement of antibacterial activity of β-lactam antibiotics by [P2W18O62]6−, [SiMo12O40]4−, and [PTi2W10O40]7− against methicillin-resistant and vancomycin-resistant Staphylococcus aureus. J. Inorg. Biochem. 2006, 100, 1225–1233. [Google Scholar] [CrossRef]
- Yu, X.; Chen, C.; Peng, J.; Shi, Z.; Shen, Y.; Mei, J.; Ren, Z. Antibacterial-active multilayer films composed of polyoxometalate and Methyl Violet: Fabrication, characterization and properties. Thin Solid Films 2014, 571, 69–74. [Google Scholar] [CrossRef]
- Gumerova, N.; Al-Sayed, E.; Krivosudský, L.; Čipčić-Paljetak, H.; Verbanac, D.; Rompel, A. Antibacterial Activity of Polyoxometalates Against Moraxella catarrhalis. Front. Chem. 2018, 6, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Farzana, R.; Iqra, P.; Hunaiza, T. Antioxidant and antimicrobial effects of polyoxometalates. Microbiol. Curr. Res. 2018, 2, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Leite, A.; Bessa, L.J.; Silva, A.M.G.; Gameiro, P.; de Castro, B.; Rangel, M. Antibacterial activity of naphthyl derived bis-(3-hydroxy-4-pyridinonate) copper(II) complexes against multidrug-resistant bacteria. J. Inorg. Biochem. 2019, 197, 110704. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, G.; Saoncella, O.; Kaner, P.; Altinkaya, S.A.; Figoli, A.; Bonchio, M.; Carraro, M. Chitosan-polyoxometalate nanocomposites: Synthesis, characterization and application as antimicrobial agents. J. Clust. Sci. 2014, 25, 839–854. [Google Scholar] [CrossRef] [Green Version]
- Daima, H.K.; Selvakannan, P.R.; Kandjani, A.E.; Shukla, R.; Bhargava, S.K.; Bansal, V. Synergistic influence of polyoxometalate surface corona towards enhancing the antibacterial performance of tyrosine-capped Ag nanoparticles. Nanoscale 2014, 6, 758–765. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Yu, Q.; Liu, Y.; Yin, P. Bacterial hyperpolarization modulated by polyoxometalates for solutions of antibiotic resistance. J. Inorg. Biochem. 2021, 220, 111463. [Google Scholar] [CrossRef]
- Pope, M. Heteropoly and Isopoly Oxometalates, Inorganic; Springer: Berlin/Heidelberg, Germany, 1983; ISBN 978-3-662-12006-4. [Google Scholar]
- Gumerova, N.I.; Rompel, A. Polyoxometalates in solution: Speciation under spotlight. Chem. Soc. Rev. 2020, 49, 7568–7601. [Google Scholar] [CrossRef]
- Stuckart, M.; Monakhov, K.Y. Polyoxometalates as components of supramolecular assemblies. Chem. Sci. 2019, 10, 4364–4376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rompuy, L.S.; Parac-Vogt, T.N. Interactions between polyoxometalates and biological systems: From drug design to artificial enzymes. Curr. Opin. Biotechnol. 2019, 58, 92–99. [Google Scholar] [CrossRef]
- Aureliano, M.; Gumerova, N.I.; Sciortino, G.; Garribba, E.; Rompel, A.; Crans, D.C. Polyoxovanadates with emerging biomedical activities. Coord. Chem. Rev. 2021, 447, 214143. [Google Scholar] [CrossRef]
- Bijelic, A.; Aureliano, M.; Rompel, A. Polyoxometalates as potential next-generation metallodrugs in the combat against cancer. Angew. Chem. Int. Ed. 2019, 58, 2980–2999. [Google Scholar] [CrossRef] [Green Version]
- Bijelic, A.; Aureliano, M.; Rompel, A. The antibacterial activity of polyoxometalates: Structures, antibiotic effects and future perspectives. Chem. Commun. 2018, 54, 1153–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasenknopf, B. Polyoxometalates: Introduction to a class of inorganic compounds and their biomedical applications. Front. Biosci. 2005, 10, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Lara, E.; Treviño, S.; Sánchez-Gaytán, B.L.; Sánchez-Mora, E.; Castro, M.E.; Meléndez-Bustamante, F.J.; Méndez-Rojas, M.A.; González-Vergara, E. Decavanadate salts of cytosine and metformin: A combined experimental-theoretical study of potential metallodrugs against diabetes and cancer. Front. Chem. 2018, 6, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aureliano, M.; Gumerova, N.I.; Sciortino, G.; Garribba, E.; McLauchlan, C.C.; Rompel, A.; Crans, D.C. Polyoxidovanadates’ interactions with proteins: An overview. Coord. Chem. Rev. 2022, 454, 214344. [Google Scholar] [CrossRef]
- Haider, A.; Zarschler, K.; Joshi, S.A.; Smith, R.M.; Lin, Z.; Mougharbel, A.S.; Herzog, U.; Müller, C.E.; Stephan, H.; Kortz, U. Preyssler-Pope-Jeannin polyanions [NaP5W30O110]14– and [AgP5W30O110]14–: Microwave-assisted synthesis, structure, and biological activity. Z. Anorg. Allg. Chem. 2018, 644, 752–758. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, K.; Li, M.; Hu, C.; Yin, P. Sustained release of Ag+ confined inside polyoxometalates for long-lasting bacterial resistance. Chem. Commun. 2020, 56, 5287–5290. [Google Scholar] [CrossRef]
- Fu, L.; Gao, H.; Yan, M.; Li, S.; Li, X.; Dai, Z.; Liu, S. Polyoxometalate-based organic-inorganic hybrids as antitumor drugs. Small 2015, 11, 2938–2945. [Google Scholar] [CrossRef] [PubMed]
- Pimpão, C.; da Silva, I.V.; Mósca, A.F.; Pinho, J.O.; Gaspar, M.M.; Gumerova, N.I.; Rompel, A.; Aureliano, M.; Soveral, G. The aquaporin-3-inhibiting potential of polyoxotungstates. Int. J. Mol. Sci. 2020, 21, 2467. [Google Scholar] [CrossRef] [Green Version]
- Razavi, S.F.; Bamoharram, F.F.; Hashemi, T.; Shahrokhabadi, K.; Davoodnia, A. Nanolipid-loaded Preyssler polyoxometalate: Synthesis, characterization and invitro inhibitory effects on HepG2 tumor cells. Toxicol. Vitr. 2020, 68, 104917. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.L.; Schinazi, R.F.; Weeks, M.S. Anti-HIV-1 activity, toxicity, and stability studies of representative structural families of polyoxometalates. J. Med. Chem. 1990, 33, 2767–2772. [Google Scholar] [CrossRef] [PubMed]
- Aureliano, M.; Fraqueza, G.; Gumerova, N.I.; Rompel, A.; Cordoba-Granados, J.J.; Berrocal, M.; Mata, A.M. Comparison of SERCA and PMCA Inhibition Potential of Polyoxotungstates; Antunes, C., Costa, A., Palma, P., Galveias, A., Gastalho, C., Marques, M., Cancela, L., Aureliano, M., Eds.; Universidade de Évora: Évora, Portugal, 2021; ISBN 978-972-778-215-4. [Google Scholar]
- Shigeta, S.; Mori, S.; Yamase, T.; Yamamoto, N.; Yamamoto, N. Anti-RNA virus activity of polyoxometalates. Biomed. Pharmacother. 2006, 60, 211–219. [Google Scholar] [CrossRef]
- Müller, A.; Fedin, V.P.; Kuhlmann, C.; Bögge, H.; Hauptfleisch, B.; Fenske, H.-D.; Baum, G. ‘Adding’ stable functional complementary, nucleophilic and electrophilic clusters: A synthetic route to [{(SiW11O39)Mo3S4(H2O)3(μ-OH)}2]10− and [{(P2W17O61)Mo3S4(H2O)3(μ-OH)}2]14− as examples. Chem. Commun. 1999, 1189–1190. [Google Scholar] [CrossRef]
- Du, J.; Cao, M.D.; Feng, S.L.; Su, F.; Sang, X.J.; Zhang, L.C.; You, W.S.; Yang, M.; Zhu, Z.M. Two New Preyssler-Type Polyoxometalate-Based Coordination Polymers and Their Application in Horseradish Peroxidase Immobilization. Chem. A Eur. J. 2017, 23, 14614–14622. [Google Scholar] [CrossRef]
- Contant, R.; Klemperer, W.; Yaghi, O. Potassium octadecatungstodiphosphates (V) and related lacunary compounds. In Inorganic Syntheses; Ginsberg, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; pp. 104–111. [Google Scholar]
- Jeannin, Y.; Martin-Frere, J.; Choi, D.; Pope, M. The sodium pentaphosphato (V)-triacontatungstate anion isolated as the ammonium salt. In Inorganic Syntheses; Ginsberg, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; pp. 115–118. [Google Scholar]
- Dawson, B. The structure of the 9(18)-heteropoly anion in potassium 9(18)-tungstophosphate, K6(P2W18O62)·14H2O. Acta Crystallogr. 1953, 6, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Finke, R.G.; Lyon, D.K.; Nomiya, K.; Weakley, T.J.R. Structure of nonasodium—triniobatopentadecawolframatodiphosphate-acetonitrile-water (1/2/23), Na9[P2W15Nb3O62]·2CH3CN·23H2O. Acta Cryst 1990, C46, 1592–1596. [Google Scholar] [CrossRef]
- Alizadeh, M.H.; Harmalker, S.P.; Jeannin, Y.; Martin-Frer, J.; Pope, M.T. A Heteropolyanion with fivefold molecular symmetry that contains a nonlabile encapsulated sodium ion. The structure and chemistry of [NaP5W30O110]14−. J. Am. Chem. Soc. 1985, 107, 2662–2669. [Google Scholar] [CrossRef]
- EUCAST, European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2019. Available online: http://www.eucast.org (accessed on 2 December 2019).
- EUCAST, European Committee on Antimicrobial Susceptibility Testing. Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, D.; Grossmann, G.; Séquin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.N.; Horswill, A.R. A coverslip-based technique for evaluating Staphylococcus aureus biofilm formation on human plasma. Front. Cell. Infect. Microbiol. 2012, 2, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. J. Hyg. 1938, 38, 732–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, M.R. Simple colorimetric microplate test of phage lysis in Salmonella enterica. J. Microbiol. Methods 2007, 69, 394–398. [Google Scholar] [CrossRef]
- Miguel, M.G.; Faleiro, L.; Antunes, M.D.; Aazza, S.; Duarte, J.; Silvério, A.R. Antimicrobial, antiviral and antioxidant activities of “água-mel” from Portugal. Food Chem. Toxicol. 2013, 56, 136–144. [Google Scholar] [CrossRef]
- El-Guendouz, S.; Lyoussi, B.; Lourenço, J.P.; Rosa da Costa, A.M.; Miguel, M.G.; Barrocas Dias, C.; Manhita, A.; Jordao, L.; Nogueira, I.; Faleiro, M.L. Magnetite nanoparticles functionalized with propolis against methicillin resistant strains of Staphylococcus aureus. J. Taiwan Inst. Chem. Eng. 2019, 102, 25–33. [Google Scholar] [CrossRef]
- Lampl, R.; Breibeck, J.; Gumerova, N.I.; Galanski, M.S.; Rompel, A. Wells–Dawson phosphotungstates as mushroom tyrosinase inhibitors: A speciation study. Sci. Rep. 2021, 11, 19354. [Google Scholar] [CrossRef]
- Miklasińska-Majdanik, M. Mechanisms of resistance to macrolide antibiotics among Staphylococcus aureus. Antibiotics 2021, 10, 1406. [Google Scholar] [CrossRef]
- Ojo, K.K.; Striplin, M.J.; Ulep, C.C.; Close, N.S.; Zittle, J.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M.C. Staphylococcus efflux msr(A) gene characterized in Streptococcus, Enterococcus, Corynebacterium, and Pseudomonas isolates. Antimicrob. Agents Chemother. 2006, 50, 1089–1091. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udo, E.E.; Boswihi, S.S.; Mathew, B.; Noronha, B.; Verghese, T. Resurgence of chloramphenicol resistance in methicillin-resistant Staphylococcus aureus due to the acquisition of a variant florfenicol exporter (Fexav)-mediated chloramphenicol resistance in Kuwait hospitals. Antibiotics 2021, 10, 1250. [Google Scholar] [CrossRef] [PubMed]
- Lamret, F.; Varin-Simon, J.; Velard, F.; Terryn, C.; Mongaret, C.; Colin, M.; Gangloff, S.C.; Reffuveille, F. Staphylococcus aureus Strain-Dependent Biofilm Formation in Bone-Like Environment. Front. Microbiol. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuhashi, M.; Tamaki, S. Enzymatic studies on the mechanism of action of cefoxitin. J. Antibiot. 1982, 2, 888–901. [Google Scholar]
- Bastin, G.; Loison, P.; Vernex-Loset, L.; Dupire, F.; Challant, J.; Majou, D.; Boudaud, N.; Krier, G.; Gantzer, C. Structural Organizations of Qβ and MS2 Phages Affect Capsid Protein Modifications by Oxidants Hypochlorous Acid and Peroxynitrite. Front. Microbiol. 2020, 11, 1157. [Google Scholar] [CrossRef] [PubMed]
- Aureliano, M. The future is bright for polyoxometalates. BioChem 2022, 2, 8–26. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Origin and Characteristics | Source |
|---|---|---|
| Acinetobacter baumannii DSM 3007 | Urine | German Collection of Microorganisms |
| Acinetobacter baumannii I73775 | Urine, multiresistant | Laboratory Microbiology, ABC-RI, UAlg 1 |
| Escherichia coli DSM 1077 | K12 galR arg nad | German Collection of Microorganisms |
| Escherichia coli DSM 5210 | Hfr 3000 U432. Host of phage Qß (DSM 5696) | German Collection of Microorganisms |
| Escherichia coli I73194 | Urine, multiresistant | Laboratory Microbiology, ABC-RI, UAlg |
| Klebsiella pneumoniae 70923/4 | Urine, multiresistant | Laboratory Microbiology, ABC-RI, UAlg |
| Pseudomonas aeruginosa C46281 | Urine, multiresistant | Laboratory Microbiology, ABC-RI, UAlg |
| Staphylococcus aureus ATCC 6538 | Wound | American Type Culture Collection |
| Staphylococcus aureus methicillin resistant 15 (MRSA15) | Clinical | Laboratory Microbiology, ABC-RI, UAlg |
| Staphylococcus aureus methicillin resistant 16 (MRSA16) | Clinical | Laboratory Microbiology, ABC-RI, UAlg |
| Streptococcus pneumoniae D39 | Clinical | UL, UK 2 |
| Chromobacterium violaceum (CV026) | (HgR, cvil:Tn5 xylE, KanR, higher spontaneous resistance StrR) | Gift of Professor Mondher El Jaziri of the University Libre of Brussels |
| Formula | Net Charge in Solid State | Charge Density (Charge/Number of Addenda Atoms Ratio) | First Structural Report in | Synthesis | Anions Present in Aqueous Solution Based on NMR Studies | Anions Present in MHB Based on NMR Studies |
|---|---|---|---|---|---|---|
| K6[α-PV2WVI18O62]·14H2O (MW 4849.6) P2W18 | −6 | 0.33 | [52] | [50] | [α-PV2WVI18O62]6− | [α-PV2WVI18O62]6− + [α2-PV2WVI17O61]10− |
| K10[α2-PV2WVI17O61]·20H2O (MW 4914.2) P2W17 | −10 | 0.56 | [47] | [50] | [α2-PV2WVI17O61]10− | [α2-PV2WVI17O61]10− |
| K12[α-PV2WVI15O56]·24H2O (MW 4617.1) P2W15 | −12 | 0.80 | [53] | [50] | [α2-PV2WVI17O61]10− | [α2-PV2WVI17O61]10− |
| (NH4)14[NaPV5WVI30O110]·31H2O (MW 8264.0) P5W30 | −14 | 0.47 | [54] | [51] | [NaPV5WVI30O110]14− | [NaPV5WVI30O110]14− |
| Microorganisms | P2W15 | P2W17 | P2W18 | P5W30 |
|---|---|---|---|---|
| S. aureus ATCC 6538 | 3.54 (800 µM) | 2.95 (600 µM) | 2.91 (600 µM) | <0.83 (<100 µM) |
| MRSA15 | 2.65 (600 µM) | 2.95 (600 µM) | 2.91 (600 µM) | 3.31 (400 µM) |
| MRSA16 | 7.01 (1600 µM) | 7.87 (1600 µM) | 7.76 (1600 µM) | 4.96 (600 µM) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faleiro, L.; Marques, A.; Martins, J.; Jordão, L.; Nogueira, I.; Gumerova, N.I.; Rompel, A.; Aureliano, M. The Preyssler-Type Polyoxotungstate Exhibits Anti-Quorum Sensing, Antibiofilm, and Antiviral Activities. Biology 2022, 11, 994. https://doi.org/10.3390/biology11070994
Faleiro L, Marques A, Martins J, Jordão L, Nogueira I, Gumerova NI, Rompel A, Aureliano M. The Preyssler-Type Polyoxotungstate Exhibits Anti-Quorum Sensing, Antibiofilm, and Antiviral Activities. Biology. 2022; 11(7):994. https://doi.org/10.3390/biology11070994
Chicago/Turabian StyleFaleiro, Leonor, Ana Marques, João Martins, Luísa Jordão, Isabel Nogueira, Nadiia I. Gumerova, Annette Rompel, and Manuel Aureliano. 2022. "The Preyssler-Type Polyoxotungstate Exhibits Anti-Quorum Sensing, Antibiofilm, and Antiviral Activities" Biology 11, no. 7: 994. https://doi.org/10.3390/biology11070994
APA StyleFaleiro, L., Marques, A., Martins, J., Jordão, L., Nogueira, I., Gumerova, N. I., Rompel, A., & Aureliano, M. (2022). The Preyssler-Type Polyoxotungstate Exhibits Anti-Quorum Sensing, Antibiofilm, and Antiviral Activities. Biology, 11(7), 994. https://doi.org/10.3390/biology11070994