
Morphological and Tissue Characterization with 3D Reconstruction of a 350-Year-Old Austrian Ardea purpurea Glacier Mummy
 , , , , , and
, , , , , and 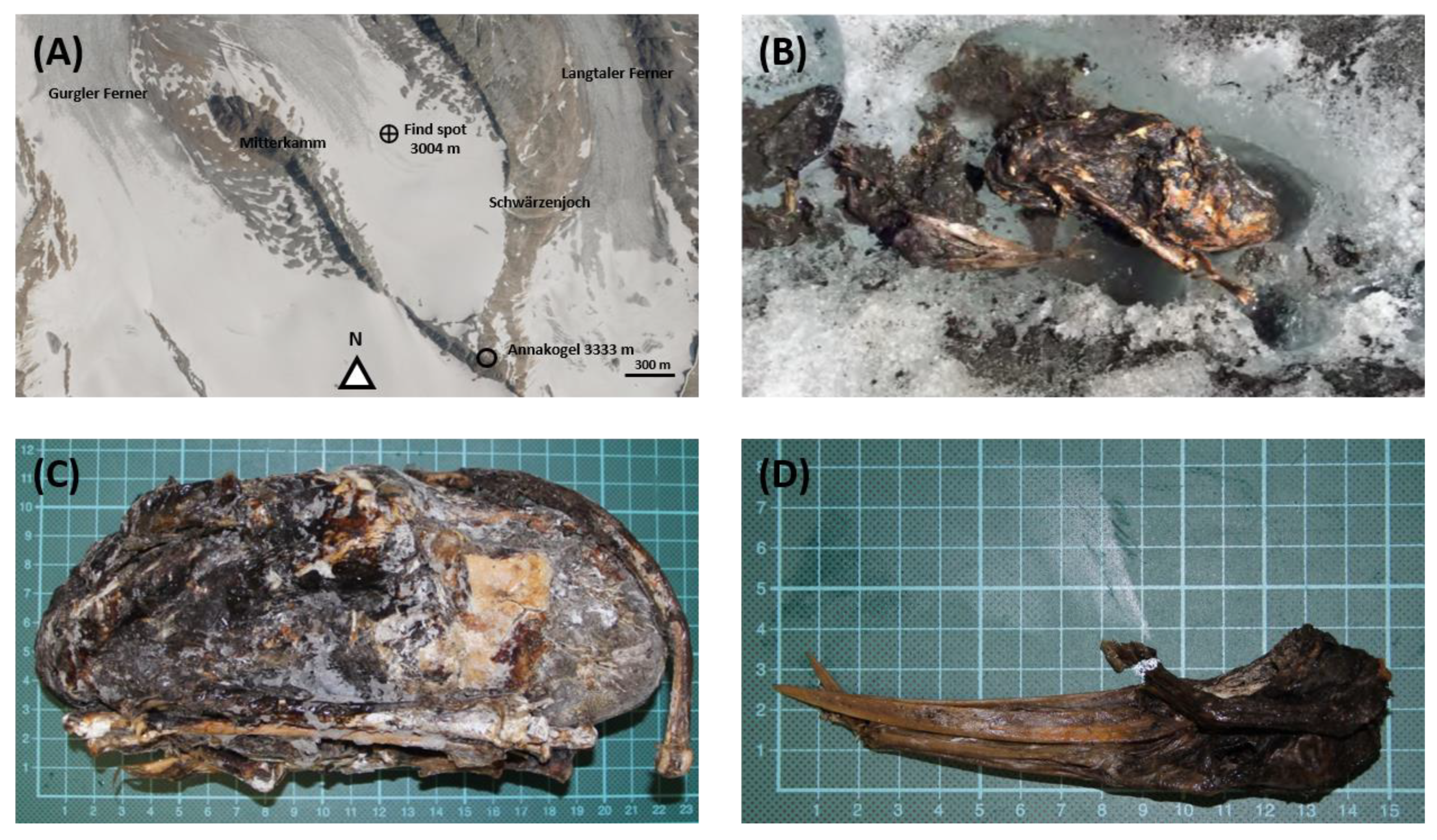{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Find Spot
2.2. Micro-Computed Tomography (Micro-CT)
- Planar CT slice images: reconstructed CT slice images along axial, sagittal, or coronal planes.
- Multi-planar reconstruction: reconstructed CT slice images along planes of freely selectable position and angle.
- Curved planar reconstruction: reconstructed CT slice images along planes of arbitrary orientation.
- Maximum intensity projection: the voxel with the highest intensity is displayed along a specific projection through the volume dataset.
- Surface Rendering: according to a defined mean value, the surface is rendered along a certain projection of the volume data set.
- Volume Rendering: assignment of a color value to a voxel according to its X-ray density. It is then possible to make certain regions transparent.
- Segmentation: semi-automatic segmentation tools such as Threshold Volume, Region Grow, and Object Extractor were used to segment the calcifications and internal organs. The procedure consists of selecting a pixel within an area called a seed. Neighboring pixels of similar density are automatically added or connected. A density threshold is chosen to capture the total volume in the pmCT layer. This process is repeated for each layer, and the total volume is automatically added.
2.3. Magnetic Resonance Imaging (MRI) Data Acquisition and Processing
2.4. Sample Collection and Tissue Specimens
2.5. Radiocarbon Dating
2.6. DNA Analysis
3. Results
3.1. Radiocarbon Dating
3.2. Morphological Analysis via Micro-CT and MRI
- BB; broad basal = 20.67 mm.
- BF; broad of the facies articularis basalis = 14.23 mm.
- GL; greatest diagonal length (acrocoracoid–apex lateralis) = 54.10 mm.
- LM; length medial (acrocoracoid–apex medialis) = 51.90 mm.
3.3. DNA Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Hafner, A. Archaeological discoveries on Schnidejoch and at other ice sites in the European Alps. Arctic 2012, 65, 189–202. [Google Scholar] [CrossRef]
- Aufderheide, A.C. The Scientific Study of Mummies; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Lynnerup, N. Methods in mummy research. Anthropol. Anz. 2009, 67, 357–384. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, K.C. Advances in paleopathology in context: A focus on soft tissue paleopathology. Int. J. Paleopathol. 2020, 29, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Licata, M.; Tosi, A.; Larentis, O.; Rossetti, C.; Lorio, S.; Pinto, A. Radiology of Mummies. Semin. Ultrasound CT MR 2019, 40, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Ruhli, F.J.; Chhem, R.K.; Boni, T. Diagnostic paleoradiology of mummified tissue: Interpretation and pitfalls. Can. Assoc. Radiol. J. 2004, 55, 218–227. [Google Scholar]
- Ruhli, F.J. Short Review: Magnetic Resonance Imaging of Ancient Mummies. Anat. Rec. 2015, 298, 1111–1115. [Google Scholar] [CrossRef]
- Baldock, C.; Hughes, S.W.; Whittaker, D.K.; Taylor, J.; Davis, R.; Spencer, A.J.; Tonge, K.; Sofat, A. 3-D reconstruction of an ancient Egyptian mummy using X-ray computer tomography. J. R. Soc. Med. 1994, 87, 806–808. [Google Scholar]
- Sebes, J.I.; Langston, J.W.; Gavant, M.L.; Rothschild, B. MR imaging of growth recovery lines in fossil vertebrae. AJR Am. J. Roentgenol. 1991, 157, 415–416. [Google Scholar] [CrossRef]
- Piepenbrink, H.; Frahm, J.; Haase, A.; Matthaei, D. Nuclear magnetic resonance imaging of mummified corpses. Am. J. Phys. Anthropol. 1986, 70, 27–28. [Google Scholar] [CrossRef]
- Ohrstrom, L.M.; von Waldburg, H.; Speier, P.; Bock, M.; Suri, R.E.; Ruhli, F.J. Scenes from the past: MR imaging versus CT of ancient Peruvian and Egyptian mummified tissues. Radiographics 2013, 33, 291–296. [Google Scholar] [CrossRef]
- William, A.; Murphy, J.; Nedden, D.z.; Gostner, P.; Knapp, R.; Recheis, W.; Seidler, H. The Iceman: Discovery and Imaging. Radiology 2003, 226, 614–629. [Google Scholar] [CrossRef]
- Keller, A.; Graefen, A.; Ball, M.; Matzas, M.; Boisguerin, V.; Maixner, F.; Leidinger, P.; Backes, C.; Khairat, R.; Forster, M.; et al. New insights into the Tyrolean Iceman’s origin and phenotype as inferred by whole-genome sequencing. Nat. Commun. 2012, 3, 698. [Google Scholar] [CrossRef]
- Callaway, E. Famous ancient iceman had familiar stomach infection. Nature 2016. [Google Scholar] [CrossRef]
- Callaway, E. Iceman’s DNA reveals health risks and relations. Nature 2012. [Google Scholar] [CrossRef]
- Coia, V.; Cipollini, G.; Anagnostou, P.; Maixner, F.; Battaggia, C.; Brisighelli, F.; Gómez-Carballa, A.; Destro Bisol, G.; Salas, A.; Zink, A. Whole mitochondrial DNA sequencing in Alpine populations and the genetic history of the Neolithic Tyrolean Iceman. Sci. Rep. 2016, 6, 18932. [Google Scholar] [CrossRef] [Green Version]
- Engelke, K.; Karolczak, M.; Lutz, A.; Seibert, U.; Schaller, S.; Kalender, W. Micro-CT. Technology and application for assessing bone structure. Radiologe 1999, 39, 203–212. [Google Scholar] [CrossRef]
- Cavanaugh, D.; Johnson, E.; Price, R.E.; Kurie, J.; Travis, E.L.; Cody, D.D. In vivo respiratory-gated micro-CT imaging in small-animal oncology models. Mol. Imaging 2004, 3, 55–62. [Google Scholar] [CrossRef]
- Kalender, W.A.; Deak, P.; Kellermeier, M.; van Straten, M.; Vollmar, S.V. Application- and patient size-dependent optimization of x-ray spectra for CT. Med. Phys. 2009, 36, 993–1007. [Google Scholar] [CrossRef]
- Brouwers, J.E.; van Rietbergen, B.; Huiskes, R. No effects of in vivo micro-CT radiation on structural parameters and bone marrow cells in proximal tibia of wistar rats detected after eight weekly scans. J. Orthop. Res. 2007, 25, 1325–1332. [Google Scholar] [CrossRef]
- Brouwers, J.E.; Lambers, F.M.; Gasser, J.A.; van Rietbergen, B.; Huiskes, R. Bone degeneration and recovery after early and late bisphosphonate treatment of ovariectomized wistar rats assessed by in vivo micro-computed tomography. Calcif. Tissue Int. 2008, 82, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Stock, S.R.; Nagaraja, S.; Barss, J.; Dahl, T.; Veis, A. X-ray microCT study of pyramids of the sea urchin Lytechinus variegatus. J. Struct. Biol. 2003, 141, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Bogart, S.J.; Spiers, G.; Cholewa, E. X-ray microCT imaging technique reveals corm microstructures of an arctic-boreal cotton-sedge, Eriophorum vaginatum. J. Struct. Biol. 2010, 171, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Mirbach, T.; Hess, M.; Metscher, B.D.; Ladich, F. A unique swim bladder-inner ear connection in a teleost fish revealed by a combined high-resolution microtomographic and three-dimensional histological study. BMC Biol. 2013, 11, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulwetter, S.; Vasileiadou, A.; Kouratoras, M.; Thanos, D.; Arvanitidis, C. Micro-computed tomography: Introducing new dimensions to taxonomy. ZooKeys 2013, 263, 1–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinley, J.; Hawkins, L.; Paterson, G.; Ball, A.D.; Sinclair, I.; Sinnett-Jones, P.; Lanham, S. Micro-computed X-ray tomography: A new non-destructive method of assessing sectional, fly-through and 3D imaging of a soft-bodied marine worm. J. Microsc. 2010, 238, 123–133. [Google Scholar] [CrossRef]
- Schulz-Mirbach, T.; Metscher, B.; Ladich, F. Relationship between swim bladder morphology and hearing abilities—A case study on Asian and African cichlids. PLoS ONE 2012, 7, e42292. [Google Scholar] [CrossRef]
- Collareta, A.; Landini, W.; Lambert, O.; Post, K.; Tinelli, C.; Di Celma, C.; Panetta, D.; Tripodi, M.; Salvadori, P.A.; Caramella, D.; et al. Piscivory in a Miocene Cetotheriidae of Peru: First record of fossilized stomach content for an extinct baleen-bearing whale. Die Nat. 2015, 102, 70. [Google Scholar] [CrossRef]
- Ketcham, R.A.; Carlson, W.D. Acquisition, optimization and interpretation of X-ray computed tomographic imagery: Applications to the geosciences. Comput. Geosci. 2001, 27, 381–400. [Google Scholar] [CrossRef]
- Evans, N.J.; McInnes, B.I.; Squelch, A.P.; Austin, P.J.; McDonald, B.J.; Wu, Q. Application of X-ray micro-computed tomography in (U–Th)/He thermochronology. Chem. Geol. 2008, 257, 101–113. [Google Scholar] [CrossRef]
- Wildenschild, D.; Vaz, C.; Rivers, M.; Rikard, D.; Christensen, B. Using X-ray computed tomography in hydrology: Systems, resolutions, and limitations. J. Hydrol. 2002, 267, 285–297. [Google Scholar] [CrossRef]
- Elliot, T.R.; Heck, R.J. A comparison of 2D vs. 3D thresholding of X-ray CT imagery. Can. J. Soil Sci. 2007, 87, 405–412. [Google Scholar] [CrossRef]
- Elliot, T.R.; Heck, R.J. A comparison of optical and X-ray CT technique for void analysis in soil thin section. Geoderma 2007, 141, 60–70. [Google Scholar] [CrossRef]
- Jassogne, L.; McNeill, A.; Chittleborough, D. 3D-visualization and analysis of macro-and meso-porosity of the upper horizons of a sodic, texture-contrast soil. Eur. J. Soil Sci. 2007, 58, 589–598. [Google Scholar] [CrossRef]
- Torrance, J.; Elliot, T.; Martin, R.; Heck, R. X-ray computed tomography of frozen soil. Cold Reg. Sci. Technol. 2008, 53, 75–82. [Google Scholar] [CrossRef]
- Wang, Y. Morphological Characterization of Wood Plastic Composite (WPC) with Advanced Imaging Tools: Developing Methodologies for Reliable Phase and Internal Damage Characterization. Masters Thesis, Oregon State University, Corvallis, OR, USA, 2007. [Google Scholar]
- Tondi, G.; Blacher, S.; Léonard, A.; Pizzi, A.; Fierro, V.; Leban, J.; Celzard, A. X-ray microtomography studies of tannin-derived organic and carbon foams. Microsc. Microanal. 2009, 15, 384. [Google Scholar] [CrossRef] [Green Version]
- Cnudde, V.; Masschaele, B.; De Cock, H.E.; Olstad, K.; Vlaminck, L.; Vlassenbroeck, J.; Dierick, M.; Witte, Y.D.; Van Hoorebeke, L.; Jacobs, P. Virtual histology by means of high-resolution X-ray CT. J. Microsc. 2008, 232, 476–485. [Google Scholar] [CrossRef]
- Schnabl, J.; Glueckert, R.; Feuchtner, G.; Recheis, W.; Potrusil, T.; Kuhn, V.; Wolf-Magele, A.; Riechelmann, H.; Sprinzl, G.M. Sheep as a large animal model for middle and inner ear implantable hearing devices: A feasibility study in cadavers. Otol. Neurotol. 2012, 33, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Maret, D.; Peters, O.A.; Dedouit, F.; Telmon, N.; Sixou, M. Cone-Beam Computed Tomography: A useful tool for dental age estimation? Med. Hypotheses 2011, 76, 700–702. [Google Scholar] [CrossRef]
- Jones, A.C.; Milthorpe, B.; Averdunk, H.; Limaye, A.; Senden, T.J.; Sakellariou, A.; Sheppard, A.P.; Sok, R.M.; Knackstedt, M.A.; Brandwood, A.; et al. Analysis of 3D bone ingrowth into polymer scaffolds via micro-computed tomography imaging. Biomaterials 2004, 25, 4947–4954. [Google Scholar] [CrossRef]
- Hanson, N.A.; Bagi, C.M. Alternative approach to assessment of bone quality using micro-computed tomography. Bone 2004, 35, 326–333. [Google Scholar] [CrossRef]
- Schmidt, V.-M.; Zelger, P.; Woess, C.; Pallua, A.K.; Arora, R.; Degenhart, G.; Brunner, A.; Zelger, B.; Schirmer, M.; Rabl, W.; et al. Application of Micro-Computed Tomography for the Estimation of the Post-Mortem Interval of Human Skeletal Remains. Biology 2022, 11, 1105. [Google Scholar] [CrossRef]
- Sliva, D. Cellular and physiological effects of Ganoderma lucidum (Reishi). Mini Rev. Med. Chem. 2004, 4, 873–879. [Google Scholar] [CrossRef]
- Deng, J.Y.; Chen, S.J.; Jow, G.M.; Hsueh, C.W.; Jeng, C.J. Dehydroeburicoic acid induces calcium- and calpain-dependent necrosis in human U87MG glioblastomas. Chem. Res. Toxicol. 2009, 22, 1817–1826. [Google Scholar] [CrossRef] [PubMed]
- Steppe, K.; Cnudde, V.; Girard, C.; Lemeur, R.; Cnudde, J.P.; Jacobs, P. Use of X-ray computed microtomography for non-invasive determination of wood anatomical characteristics. J. Struct. Biol. 2004, 148, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Dutilleul, P.; Lontoc-Roy, M.; Prasher, S.O. Branching out with a CT scanner. Trends Plant Sci. 2005, 10, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, A. Kapitel 5| Das Potential historischer Karten zur Rekonstruktion des Gletscherrückgangs im Gurgler Tal. Alp. Forsch. Obergurgl Band 2013, 3, 95. [Google Scholar]
- Meixner, W.; Siegl, G. Kapitel 1| Historisches zum Thema Gletscher, Gletschervorfeld und Obergurgl. Alp. Forsch. Obergurgl Band 2010, 1. [Google Scholar]
- Kellner, M. Vergleichend Morphologische Untersuchungen an Einzelknochen des Postkranialen Skelettes in Europa Vorkommender Ardeidae; Universitat Tierarztliche Fakultat: Munich, Germany, 1986. [Google Scholar]
- Grodzki, D.M.; Jakob, P.M.; Heismann, B. Ultrashort echo time imaging using pointwise encoding time reduction with radial acquisition (PETRA). Magn. Reson. Med. 2012, 67, 510–518. [Google Scholar] [CrossRef]
- Dagher, G.; Becker, K.-F.; Bonin, S.; Foy, C.; Gelmini, S.; Kubista, M.; Kungl, P.; Oelmueller, U.; Parkes, H.; Pinzani, P.; et al. Pre-analytical processes in medical diagnostics: New regulatory requirements and standards. New Biotechnol. 2019, 52, 121–125. [Google Scholar] [CrossRef]
- Stumptner, C.; Sargsyan, K.; Kungl, P.; Zatloukal, K. Crucial role of high quality biosamples in biomarker development. In Handbook of Biomarkers and Precision Medicine; Chapman and Hall/CRC: London, UK, 2019; pp. 128–134. [Google Scholar]
- Hajdas, I. Radiocarbon dating and its applications in Quaternary studies. E&G Quat. Sci. J. 2008, 57, 2–24. [Google Scholar] [CrossRef]
- Hajdas, I.; Bonani, G.; Furrer, H.; Mäder, A.; Schoch, W. Radiocarbon chronology of the mammoth site at Niederweningen, Switzerland: Results from dating bones, teeth, wood, and peat. Quat. Int. 2007, 164–165, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Ramsey, C.B.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, C.B.; Lee, S. Recent and Planned Developments of the Program OxCal. Radiocarbon 2013, 55, 720–730. [Google Scholar] [CrossRef]
- Stuiver, M.; Polach, H.A. Discussion Reporting of 14C Data. Radiocarbon 1977, 19, 355–363. [Google Scholar] [CrossRef]
- Tully, G.; Bar, W.; Brinkmann, B.; Carracedo, A.; Gill, P.; Morling, N.; Parson, W.; Schneider, P. Considerations by the European DNA profiling (EDNAP) group on the working practices, nomenclature and interpretation of mitochondrial DNA profiles. Forensic Sci. Int. 2001, 124, 83–91. [Google Scholar] [CrossRef]
- Gilbert, M.T.; Bandelt, H.J.; Hofreiter, M.; Barnes, I. Assessing ancient DNA studies. Trends Ecol. Evol. 2005, 20, 541–544. [Google Scholar] [CrossRef]
- Xavier, C.; Eduardoff, M.; Bertoglio, B.; Amory, C.; Berger, C.; Casas-Vargas, A.; Pallua, J.; Parson, W. Evaluation of DNA Extraction Methods Developed for Forensic and Ancient DNA Applications Using Bone Samples of Different Age. Genes 2021, 12, 146. [Google Scholar] [CrossRef]
- Parson, W.; Pegoraro, K.; Niederstätter, H.; Föger, M.; Steinlechner, M. Species identification by means of the cytochrome b gene. Int. J. Leg. Med. 2000, 114, 23–28. [Google Scholar] [CrossRef]
- University of Hawaii. The Simple Fool’s Guide to PCR; University of Hawaii: Honolulu, HI, USA, 2002. [Google Scholar]
- Cox, S.L. A Critical Look at Mummy CT Scanning. Anat. Rec. 2015, 298, 1099–1110. [Google Scholar] [CrossRef]
- Baadsvik, E.L.; Weiger, M.; Froidevaux, R.; Rösler, M.B.; Brunner, D.O.; Öhrström, L.; Rühli, F.J.; Eppenberger, P.; Pruessmann, K.P. High-resolution MRI of mummified tissues using advanced short-T2 methodology and hardware. Magn. Reson. Med. 2021, 85, 1481–1492. [Google Scholar] [CrossRef]
- Lynnerup, N. Mummies. Am. J. Phys. Anthropol. 2007, 134, 162–190. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unterberger, S.H.; Berger, C.; Schirmer, M.; Pallua, A.K.; Zelger, B.; Schäfer, G.; Kremser, C.; Degenhart, G.; Spiegl, H.; Erler, S.; et al. Morphological and Tissue Characterization with 3D Reconstruction of a 350-Year-Old Austrian Ardea purpurea Glacier Mummy. Biology 2023, 12, 114. https://doi.org/10.3390/biology12010114
Unterberger SH, Berger C, Schirmer M, Pallua AK, Zelger B, Schäfer G, Kremser C, Degenhart G, Spiegl H, Erler S, et al. Morphological and Tissue Characterization with 3D Reconstruction of a 350-Year-Old Austrian Ardea purpurea Glacier Mummy. Biology. 2023; 12(1):114. https://doi.org/10.3390/biology12010114
Chicago/Turabian StyleUnterberger, Seraphin H., Cordula Berger, Michael Schirmer, Anton Kasper Pallua, Bettina Zelger, Georg Schäfer, Christian Kremser, Gerald Degenhart, Harald Spiegl, Simon Erler, and et al. 2023. "Morphological and Tissue Characterization with 3D Reconstruction of a 350-Year-Old Austrian Ardea purpurea Glacier Mummy" Biology 12, no. 1: 114. https://doi.org/10.3390/biology12010114