
SPIM-Flow: An Integrated Light Sheet and Microfluidics Platform for Hydrodynamic Studies of Hydra

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
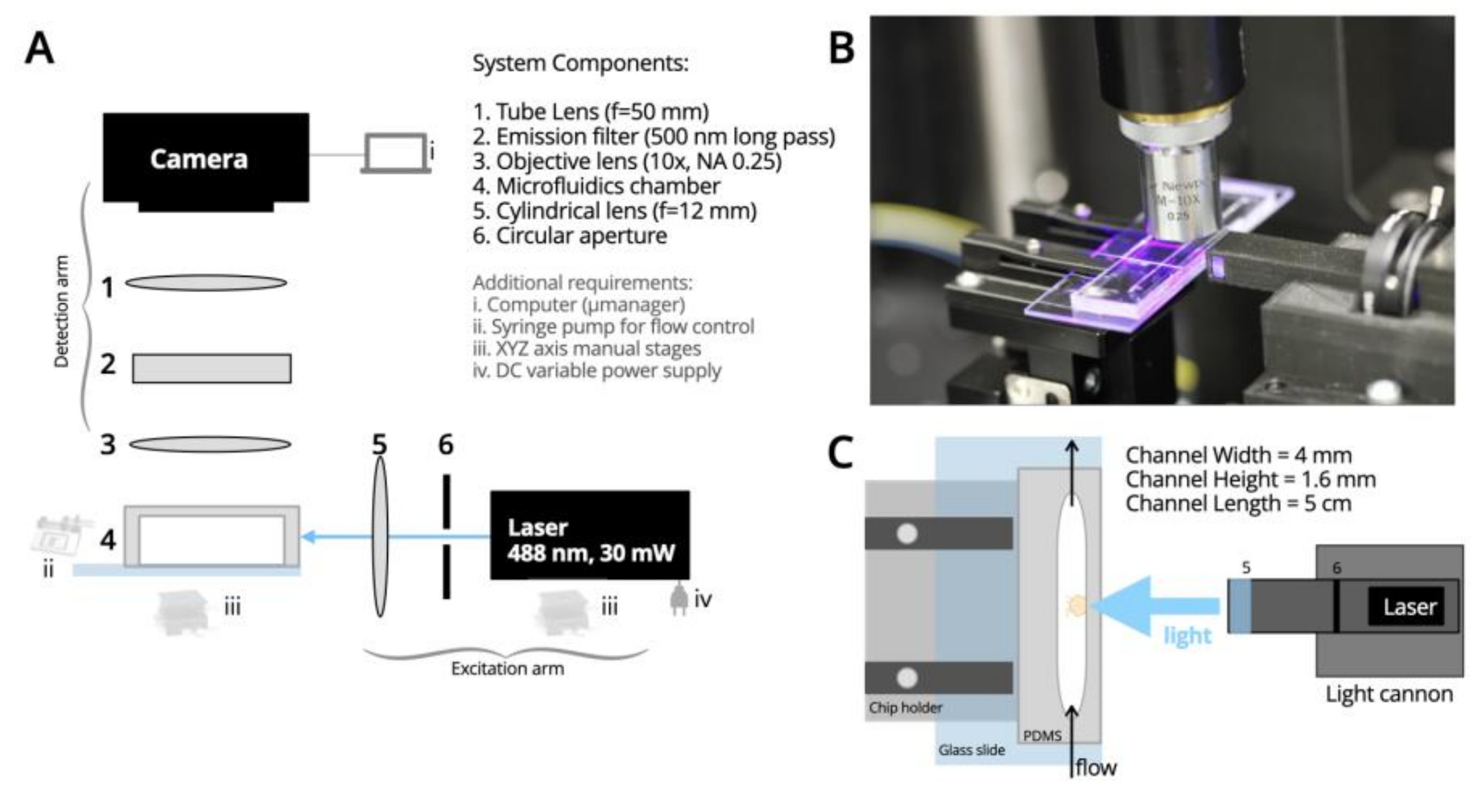
2. Materials and Methods
3. Results
3.1. Health and Feeding
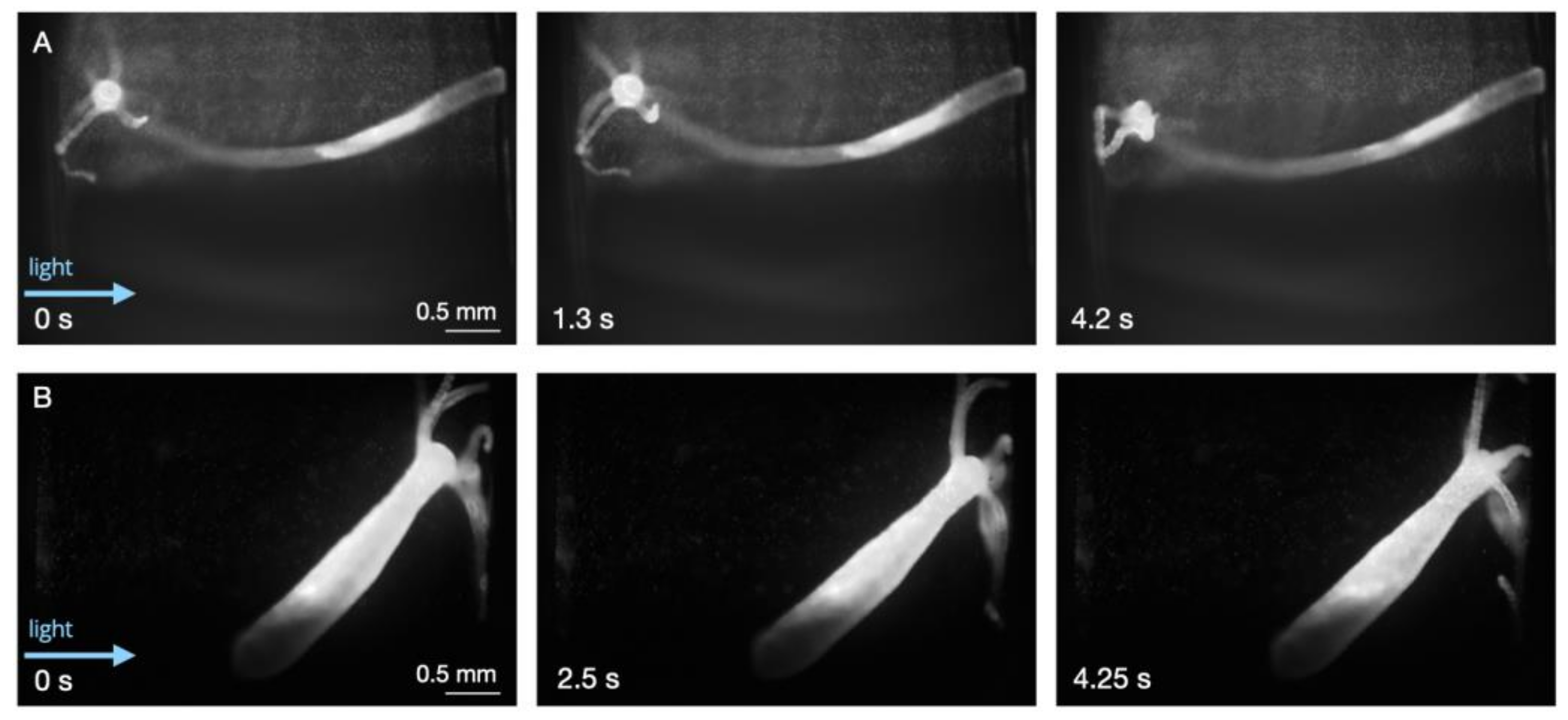
3.2. Static Conditions
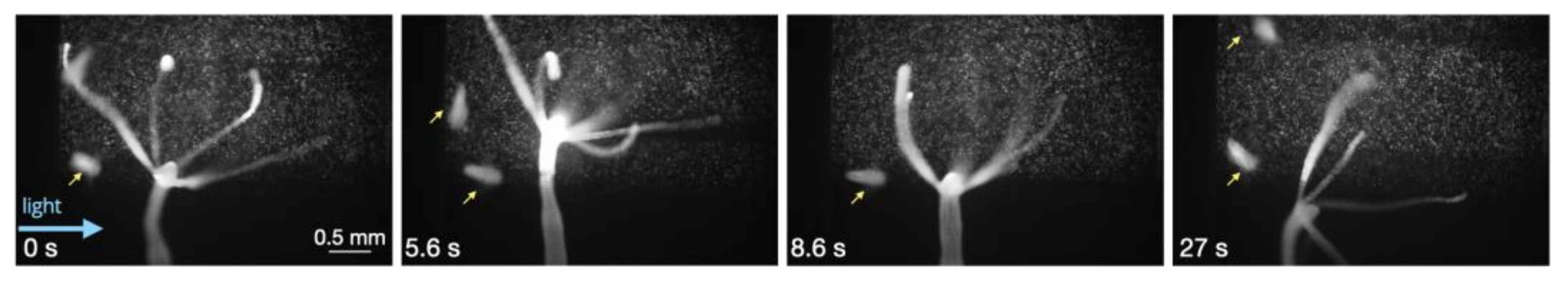
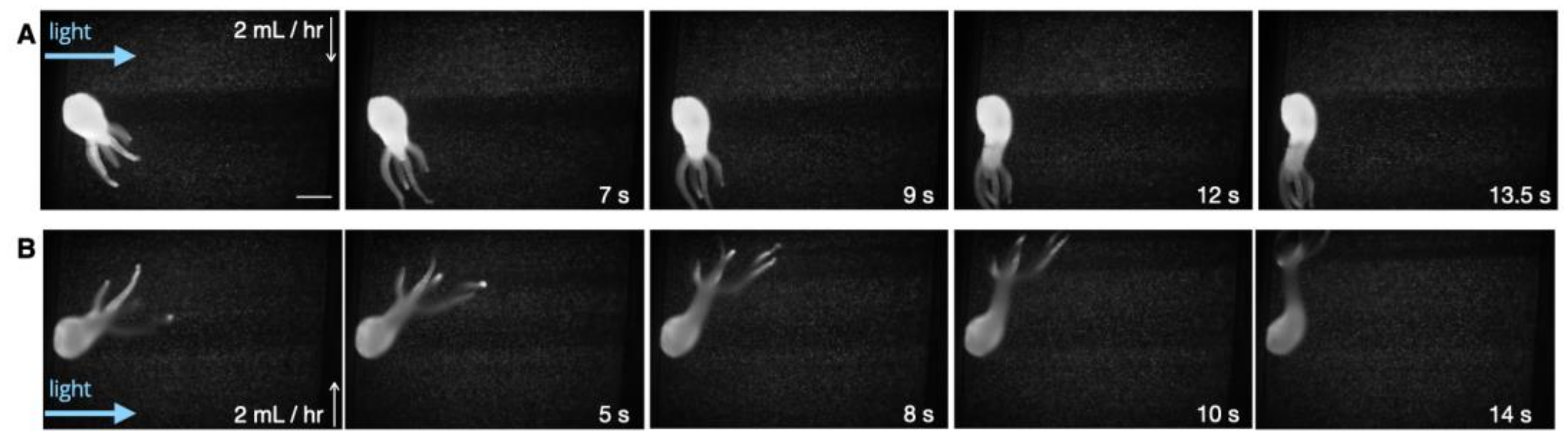
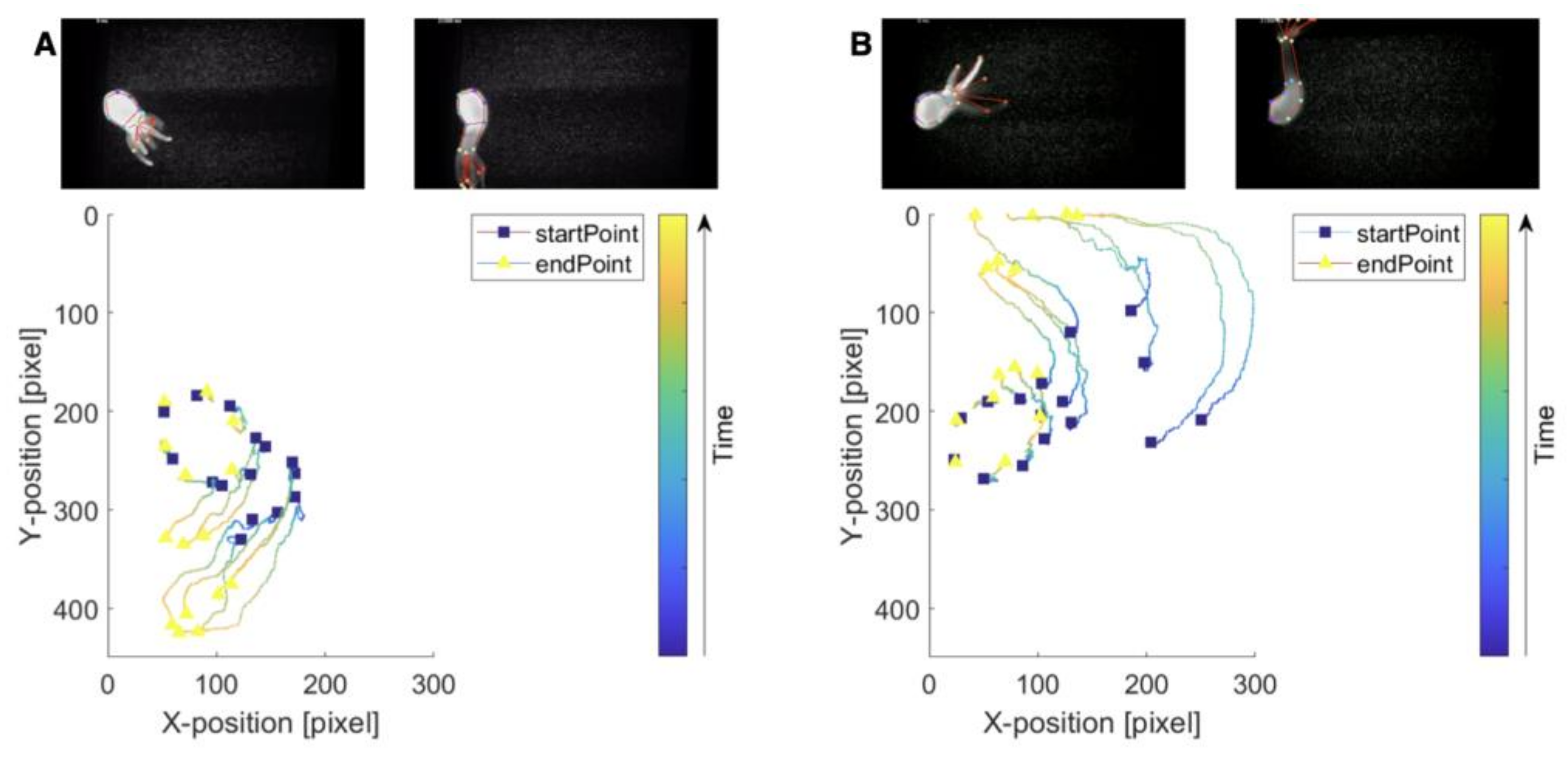
3.3. Response to Flow
3.4. Adhesion and Detachment from Surfaces
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stelzer, E.H.K.; Strobl, F.; Chang, B.-J.; Preusser, F.; Preibisch, S.; McDole, K.; Fiol, R. Light sheet fluorescence microscopy. Nat. Rev. Methods Prim. 2021, 1, 73. [Google Scholar] [CrossRef]
- Huisken, J.; Swoger, J.; Del Bene, F.; Wittbrodt, J.; Stelzer, E.H.K. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science 2004, 305, 1007–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olarte, O.E.; Andilla, J.; Gualda, E.J.; Loza-Alvarez, P. Light-sheet microscopy: A tutorial. Adv. Opt. Photonics 2018, 10, 111–179. [Google Scholar] [CrossRef]
- Voigt, F.F.; Kirschenbaum, D.; Platonova, E.; Campbell, R.A.A.; Kastli, R.; Schaettin, M.; Egolf, L.; Van der Bourg, A.; Bethge, P.; Haenraets, K.; et al. The mesospim initiative: Open-source light-sheet microscopes for imaging cleared tissue. Nat. Methods 2019, 16, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Gualda, E.J.; Pereira, H.; Vale, T.; Estrada, M.F.; Brito, C.; Moreno, N. Spim-fluid: Open source light-sheet based platform for high-throughput imaging. Biomed. Opt. Express 2015, 6, 4447–4456. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Lange, M.; Millett-Sikking, A.; Zhao, X.; Bragantini, J.; VijayKumar, S.; Kamb, M.; Gómez-Sjöberg, R.; Solak, A.C.; Wang, W.; et al. Daxi—High-resolution, large imaging volume and multi-view single-objective light-sheet microscopy. Nat. Methods 2022, 19, 461–469. [Google Scholar] [CrossRef]
- Hedde, P.N. minispim—A miniaturized light-sheet microscope. ACS Sens. 2021, 6, 2654–2663. [Google Scholar] [CrossRef]
- Hedde, P.N.; Stakic, M.; Gratton, E. Rapid measurement of molecular transport and interaction inside living cells using single plane illumination. Sci. Rep. 2014, 4, 7048. [Google Scholar] [CrossRef] [Green Version]
- Sapoznik, E.; Chang, B.-J.; Huh, J.; Ju, R.J.; Azarova, E.V.; Pohlkamp, T.; Welf, E.S.; Broadbent, D.; Carisey, A.F.; Stehbens, S.J.; et al. A versatile oblique plane microscope for large-scale and high-resolution imaging of subcellular dynamics. Elife 2020, 9, e57681. [Google Scholar] [CrossRef]
- Lemon, W.C.; Pulver, S.R.; Höckendorf, B.; McDole, K.; Branson, K.; Freeman, J.; Keller, P.J. Whole-central nervous system functional imaging in larval drosophila. Nat. Commun. 2015, 6, 7924. [Google Scholar] [CrossRef]
- Silvestri, L.; Bria, A.; Costantini, I.; Sacconi, L.; Peng, H.; Iannello, G.; Pavone, F.S. Micron-scale resolution optical tomography of entire mouse brains with confocal light sheet microscopy. J. Vis. Exp. 2013, 80, e50696. [Google Scholar] [CrossRef] [Green Version]
- Von Wangenheim, D.; Hauschild, R.; Friml, J. Light sheet fluorescence microscopy of plant roots growing on the surface of a gel. J. Vis. Exp. 2017, 119, e55044. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.R.; Dodt, H.-U.; Osten, P.; Economo, M.; Chandrashekar, J.; Keller, P. Whole-brain profiling of cells and circuits in mammals by tissue clearing and light-sheet microscopy. Neuron 2020, 106, 369–387. [Google Scholar] [CrossRef]
- Zhou, P.; He, H.; Ma, H.; Wang, S.; Hu, S. A review of optical imaging technologies for microfluidics. Micromachines 2022, 13, 274. [Google Scholar] [CrossRef]
- Albert-Smet, I.; Marcos-Vidal, A.; Vaquero, J.J.; Desco, M.; Muñoz-Barrutia, A.; Ripoll, J. Applications of light-sheet microscopy in microdevices. Front. Neuroanat. 2019, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Paiè, P.; Bragheri, F.; Bassi, A.; Osellame, R. Selective plane illumination microscopy on a chip. Lab A Chip 2016, 16, 1556–1560. [Google Scholar] [CrossRef] [Green Version]
- Regmi, R.; Mohan, K.; Mondal, P.P. Light sheet based imaging flow cytometry on a microfluidic platform. Microsc. Res. Tech. 2013, 76, 1101–1107. [Google Scholar] [CrossRef]
- Lin, M.; Liu, Q.; Liu, C.; Qiao, X.; Shao, C.; Su, X. Label-free light-sheet microfluidic cytometry for the automatic identification of senescent cells. Biomed. Opt. Express 2018, 9, 1692–1703. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.-J.; Hsieh, H.-Y.; Tsai, S.-F.; Wu, C.-H.; Lee, C.-M.; Liu, Y.-T.; Lu, C.-H.; Chang, S.-W.; Chen, B.-C. Microfluidic channel integrated with a lattice lightsheet microscopic system for continuous cell imaging. Lab A Chip 2021, 21, 344–354. [Google Scholar] [CrossRef]
- Sala, F.; Castriotta, M.; Paiè, P.; Farina, A.; D’Annunzio, S.; Zippo, A.; Osellame, R.; Bragheri, F.; Bassi, A. High-throughput 3d imaging of single cells with light-sheet fluorescence microscopy on chip. Biomed. Opt. Express. 2020, 11, 4397–4407. [Google Scholar] [CrossRef]
- Jiang, H.; Zhu, T.; Zhang, H.; Nie, J.; Guan, Z.; Ho, C.-M.; Liu, S.; Fei, P. Droplet-based light-sheet fluorescence microscopy for high-throughput sample preparation, 3-d imaging and quantitative analysis on a chip. Lab A Chip 2017, 17, 2193–2197. [Google Scholar] [CrossRef] [PubMed]
- Behrouzi, M.; Rahimpouresfahani, F.; Youssef, K.; Rezai, P.; Tabatabaei, N. Optofluidic device for light-sheet fluorescence microscopy of c. elegans with a conventional wide-field microscope. In Proceedings of the Imaging, Manipulation, and Analysis of Biomolecules, Cells, and Tissues XX, San Francisco, CA, USA, 3 March 2022; Volume 11964, p. 1196402. [Google Scholar]
- Memeo, R.; Paiè, P.; Sala, F.; Castriotta, M.; Guercio, C.; Vaccari, T.; Osellame, R.; Bassi, A.; Bragheri, F. Automatic imaging of drosophila embryos with light sheet fluorescence microscopy on chip. J. Biophotonics 2021, 14, e202000396. [Google Scholar] [CrossRef]
- Vanwalleghem, G.; Schuster, K.; Taylor, M.A.; Favre-Bulle, I.A.; Scott, E.K. Brain-wide mapping of water flow perception in zebrafish. J. Neurosci. 2020, 40, 4130–4144. [Google Scholar] [CrossRef] [PubMed]
- Hedde, P.N.; Malacrida, L.; Ahrar, S.; Siryaporn, A.; Gratton, E. sidespim–selective plane illumination based on a conventional inverted microscope. Biomed. Opt. Express 2017, 8, 3918–3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogg, M.C.; Galliot, B.; Tsiairis, C.D. Model systems for regeneration: Hydra. Dev. 2019, 146, dev177212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kier, W.M. The diversity of hydrostatic skeletons. J. Exp. Biol. 2012, 215, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Taralova, E.; Dupre, C.; Yuste, R. Comprehensive machine learning analysis of Hydra behavior reveals a stable basal behavioral repertoire. Elife 2018, 7, e32605. [Google Scholar] [CrossRef]
- Carter, J.A.; Hyland, C.; Steele, R.E.; Collins, E.-M.S. Dynamics of mouth opening in Hydra. Biophys. J. 2016, 110, 1191–1201. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.D. Structure of the mouth of Hydra spp. a breach in the epithelium that disappears when it closes. Cell Tissue Res. 1987, 249, 189–197. [Google Scholar] [CrossRef]
- Iachetta, R.; Ambrosone, A.; Klimovich, A.; Wittlieb, J.; Onorato, G.; Candeo, A.; D’andrea, C.; Intartaglia, D.; Scotti, N.; Blasio, M.; et al. Real time dynamics of β-catenin expression during Hydra development, regeneration and wnt signalling activation. Int. J. Dev. Biol. 2018, 62, 311–318. [Google Scholar] [CrossRef]
- Maroudas-Sacks, Y.; Garion, L.; Shani-Zerbib, L.; Livshits, A.; Braun, E.; Keren, K. Topological defects in the nematic order of actin fibres as organization centers of Hydra morphogenesis. Nat. Phys. 2021, 17, 251–259. [Google Scholar] [CrossRef]
- Badhiwala, K.N.; Gonzales, D.L.; Vercosa, D.G.; Avants, B.W.; Robinson, J.T. Microfluidics for electrophysiology, imaging, and behavioral analysis of Hydra. Lab A Chip 2018, 18, 2523–2539. [Google Scholar] [CrossRef]
- Badhiwala, K.N.; Primack, A.S.; Juliano, C.E.; Robinson, J.T. Multiple neuronal networks coordinate Hydra mechanosensory behavior. Elife 2021, 10, e64108. [Google Scholar] [CrossRef]
- Edelstein, A.D.; Tsuchida, M.A.; Amodaj, N.; Pinkard, H.; Vale, R.D.; Stuurman, N. Advanced methods of microscope control using µmanager software. J. Biol. Methods 2014, 1, e10. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Lee, J.; Jiang, H.; Dong, S.; Jen, N.; Hsiai, T.; Ho, C.-M.; Fei, P. Compact plane illumination plugin device to enable light sheet fluorescence imaging of multi-cellular organisms on an inverted wide-field microscope. Biomed. Opt. Express 2016, 7, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Steele, R.E.; Updegrove, M.D.; Kirolos, S.A.; Mowery, L.; Martinez, D.E.; Bryant, P.J. Reproductive bet-hedging and existence in vernal pools as components of Hydra life history. Biol. Bull. 2019, 237, 111–118. [Google Scholar] [CrossRef]
- Trembley, A. Mémoires Pour Servir à L’histoire D’un Genre de Ploypes D’eau Douce, à bras en Forme de Cornes; Durand: Lower Beechmont, Australia, 1744; Volume 1. [Google Scholar]
- Wilson, E.B. The heliotropism of Hydra. Am. Nat. 1891, 25, 413–433. [Google Scholar] [CrossRef]
- Haug, G. Die lichtreaktionen der hydren. Z. Für Vgl. Physiol. 1933, 19, 246–303. [Google Scholar] [CrossRef]
- Singer, R.H.; Rushforth, N.B.; Burnett, A.L. The photodynamic action of light on Hydra. J. Exp. Zool. 1963, 154, 169–173. [Google Scholar] [CrossRef]
- Nath, T.; Mathis, A.; Chen, A.C.; Patel, A.; Bethge, M.; Mathis, M.W. Using deeplabcut for 3d markerless pose estimation across species and behaviors. Nat. Protoc. 2019, 14, 2152–2176. [Google Scholar] [CrossRef]
- Gilpin, W.; Prakash, V.N.; Prakash, M. Flowtrace: Simple visualization of coherent structures in biological fluid flows. J. Exp. Biol. 2017, 220, 3411–3418. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, W.; Prakash, V.N.; Prakash, M. Dynamic vortex arrays created by starfish larvae. Phys. Rev. Fluids 2017, 2, 090501. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Leclère, P.; Flammang, P.; Hess, M.W.; Salvenmoser, W.; Hobmayer, B.; Ladurner, P. The cellular basis of bioadhesion of the freshwater polyp Hydra. BMC Zool. 2016, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Khetan, N.; Maheshwari, S.; Athale, C.A.A. Quantitative detachment mechanics of Hydra from substrates. Proc. Indian Natn. Sci. Acad. 2019, 85, 291–299. [Google Scholar] [CrossRef]
- Shen, Y.; Siryaporn, A.; Lecuyer, S.; Gitai, Z.; Stone, H.A. Flow directs surface-attached bacteria to twitch upstream. Biophys. J. 2012, 103, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.A.; Nekimken, A.L.; Fechner, S.; O’Brien, L.E.; Pruitt, B.L. Microfluidics for mechanobiology of model organisms. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 146, pp. 217–259. [Google Scholar]
- Hwang, H.; Lu, H. Microfluidic tools for developmental studies of small model organisms–nematodes, fruit flies, and zebrafish. Biotechnol. J. 2013, 8, 192–205. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, D.; Li, H.; Rey, F.B.; Cambournac, P.; Larson, A.G.; Li, E.; Prakash, M. Scale-free vertical tracking microscopy. Nat. Methods 2020, 17, 1040–1051. [Google Scholar] [CrossRef]
- Hol, F.J.H.; Lambrechts, L.; Prakash, M. Biteoscope, an open platform to study mosquito biting behavior. Elife 2020, 9, e56829. [Google Scholar] [CrossRef]
- Kumar, S.; Hol, F.J.H.; Pujhari, S.; Ellington, C.; Narayanan, H.V.; Li, H.; Rasgon, J.L.; Prakash, M. A microfluidic platform for highly parallel bite by bite profiling of mosquito-borne pathogen transmission. Nat. Commun. 2021, 12, 6018. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedde, P.N.; Le, B.T.; Gomez, E.L.; Duong, L.; Steele, R.E.; Ahrar, S. SPIM-Flow: An Integrated Light Sheet and Microfluidics Platform for Hydrodynamic Studies of Hydra. Biology 2023, 12, 116. https://doi.org/10.3390/biology12010116
Hedde PN, Le BT, Gomez EL, Duong L, Steele RE, Ahrar S. SPIM-Flow: An Integrated Light Sheet and Microfluidics Platform for Hydrodynamic Studies of Hydra. Biology. 2023; 12(1):116. https://doi.org/10.3390/biology12010116
Chicago/Turabian StyleHedde, Per Niklas, Brian T. Le, Erika L. Gomez, Leora Duong, Robert E. Steele, and Siavash Ahrar. 2023. "SPIM-Flow: An Integrated Light Sheet and Microfluidics Platform for Hydrodynamic Studies of Hydra" Biology 12, no. 1: 116. https://doi.org/10.3390/biology12010116
APA StyleHedde, P. N., Le, B. T., Gomez, E. L., Duong, L., Steele, R. E., & Ahrar, S. (2023). SPIM-Flow: An Integrated Light Sheet and Microfluidics Platform for Hydrodynamic Studies of Hydra. Biology, 12(1), 116. https://doi.org/10.3390/biology12010116