
Soil Fungal Diversity and Ecology Assessed Using DNA Metabarcoding along a Deglaciated Chronosequence at Clearwater Mesa, James Ross Island, Antarctic Peninsula
 , ,
, ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Sites and Sediment Sampling
2.2. Soil Physical and Chemical Analyses
2.3. DNA Extraction, Illumina Library Construction and Sequencing
2.4. Data Analysis and Fungal Identification
2.5. Fungal Diversity and Ecology
3. Results
3.1. Fungal Taxonomy
3.2. Geological and Physicochemical Characteristics of Soil Sampling Sites in Relation to Fungal Assemblages Detected
3.3. Fungal Assemblage Distribution along the Soil Chronosequence
4. Discussion
4.1. Fungal Taxonomy
4.2. Fungal Distribution along the Chronosequence
4.3. Ecology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruisi, S.; Barreca, D.; Selbmann, L.; Zucconi, L.; Onofri, S. Fungi in Antarctica. Rev. Environ. Sci. Biotechnol. 2007, 6, 127–141. [Google Scholar] [CrossRef]
- Rosa, L.H.; Zani, C.L.; Cantrell, C.L.; Duke, S.O.; Dijck, P.V.; Desideri, A.; Rosa, C.A. Fungi in Antarctica: Diversity, ecology, effects of climate change, and bioprospection for bioactive compounds. In Fungi of Antarctica: Diversity, Ecology and Biotechnological Applications; Rosa, L.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–18. [Google Scholar]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.S.; Pupo, D.; Alves, T.M.A.; Junior, P.A.S.; Romanha, A.J.; Zani, C.L.; et al. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 1434–1451. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.N.; de Souza, L.M.D.; Lirio, J.M.; Coria, S.H.; Lopes, F.A.C.; Convey, P.; Carvalho-Silva, M.; de Oliveira, F.S.; Câmara, P.E.A.S.; Rosa, L.H. Diversity and ecology of fungal assemblages present in lake sediments at Clearwater Mesa, James Ross Island, Antarctica, assessed using metabarcoding of environmental DNA. Fungal Biol. 2022, 126, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Smellie, J.L.; Johnson, J.S.; Nelson, A.E. Geological Map of James Ross Island. I. James Ross Island Volcanic Group. (1:125,000 scale). BAS GEOMAP 2013, 2, 5. [Google Scholar]
- Tscherko, D.; Hammesfahr, U.; Zeltner, G.; Kandeler, E.; Böcker, R. Plant succession and rhizosphere microbial communities in a recently deglaciated alpine terrain. Basic Appl. Ecol. 2005, 6, 367–383. [Google Scholar] [CrossRef]
- Fell, J.W.; Scorzetti, G.; Connell, L.; Craig, S. Biodiversity of microeukaryotes in Antarctic Dry Valley soils with >5% soil moisture. Soil Biol. Biochem. 2006, 38, 3107–3119. [Google Scholar] [CrossRef]
- Siegert, M.; Atkinson, A.; Banwell, A.; Brandon, M.; Convey, P.; Davies, B.; Downie, R.; Edwards, T.; Hubbard, B.; Marshall, G.; et al. The Antarctic Peninsula under a 1.5 °C global warming scenario. Front. Environ. Sci. 2019, 7, 102. [Google Scholar] [CrossRef]
- Newsham, K.K.; Davey, M.L.; Hopkins, D.W.; Dennis, P.G. Regional diversity of maritime Antarctic soil fungi and predicted responses of guilds and growth forms to climate change. Front. Microbiol. 2021, 11, 615659. [Google Scholar] [CrossRef]
- Gomes, E.C.Q.; Godinho, V.M.; Silva, D.A.S.; de Paula, M.T.R.; Vitoreli, G.A.; Zani, C.L.; Alves, T.M.A.; Junior, P.A.S.; Murta, S.M.F.; Barbosa, E.C.; et al. Cultivable fungi present in Antarctic soils: Taxonomy, phylogeny, diversity, and bioprospecting of antiparasitic and herbicidal metabolites. Extremophiles 2018, 22, 381–393. [Google Scholar] [CrossRef]
- Rosa, L.H.; da Silva, T.H.; Ogaki, M.B.; Pinto, O.H.B.; Stech, M.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S. DNA metabarcoding uncovers fungal diversity in soils of protected and non-protected areas on Deception Island, Antarctica. Sci. Rep. 2020, 10, 21986. [Google Scholar] [CrossRef]
- Baeza, M.; Alcaíno, J.; Cifuntes, V. Amplicon-metagenomic analysis of fungi from Antarctic terrestrial habitats. Front. Microbiol. 2017, 8, 2235. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo, 3rd ed.; Centro Nacional de Pesquisa de Solos: Rio de Janeiro, Brazil, 2017. [Google Scholar]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Leon, C. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef]
- Richardson, R.T.; Lin, C.H.; Sponsler, D.B.; Quijia, J.O.; Goodell, K.; Johnson, R.M. Application of ITS2 metabarcoding to determine the provenance of pollen collected by honey bees in an agroecosystem. Appl. Plant Sci. 2015, 3, 1400066. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner (No. LBNL-7065E); Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Abarenkov, K.; Allan, Z.; Timo, P.; Raivo, P.; Filipp, I.; Nilsson, H.R.; Urmas, K. UNITE QIIME Release for Eukaryotes. Version 04.02.2020; UNITE Community: Salt Lake City, UT, USA, 2020. [Google Scholar]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.J.; Tappu, R. MEGAN community edition-interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-Enabled Heat Mapping for All. Nucleic Acids Res. 2016, 44, 147–153. [Google Scholar] [CrossRef]
- Medinger, R.; Nolte, V.; Pandey, R.V.; Jost, S.; Ottenwälder, B.; Schlötterer, C.; Boenigk, J. Diversity in a hidden world: Potential and limitation of next-generation sequencing for surveys of molecular diversity of eukaryotic microorganisms. Mol. Ecol. 2010, 19, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.A.; Pawlowski, J. Can abundance of protists be inferred from sequence data: A case study of Foraminifera. PLoS ONE 2013, 8, e56739. [Google Scholar] [CrossRef]
- Giner, C.R.; Forn, I.; Romac, S.; Logares, R.; de Vargas, C.; Massana, R. Environmental sequencing provides reasonable estimates of the relative abundance of specific picoeukaryotes. Appl. Environ. Microbiol. 2016, 82, 4757–4766. [Google Scholar] [CrossRef] [PubMed]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Hering, D.; Borja, A.; Jones, J.I.; Pont, D.; Boets, P.; Bouchez, A.; Bruce, K.; Drakare, S.; Hänfling, B.; Kahlert, M.; et al. Implementation options for DNA-based identification into ecological status assessment under the European Water Framework Directive. Water Res. 2018, 138, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2011. [Google Scholar]
- Tedersoo, L.; Sánchez-Ramírez, S.; Koljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Rosa, L.H.; Pinto, O.H.B.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S. DNA metabarcoding to assess the diversity of airborne fungi present over Keller Peninsula, King George Island, Antarctica. Microb. Ecol. 2021, 82, 165–172. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Rojas-Jimenez, K.; Wurzbacher, C.; Bourne, E.C.; Chiuchiolo, A.; Priscu, J.C.; Grossart, H.P. Early diverging lineages within Cryptomycota and Chytridiomycota dominate the fungal communities in ice-covered lakes of the McMurdo Dry Valleys, Antarctica. Sci. Rep. 2017, 7, 15348. [Google Scholar] [CrossRef]
- Kagami, M.; Miki, T.; Takimoto, G. Mycoloop: Chytrids in aquatic food webs. Front. Microbiol. 2014, 5, 166. [Google Scholar] [CrossRef]
- Letcher, P.M.; Powell, M.J. A taxonomic summary and revision of Rozella (Cryptomycota). IMA Fungus 2018, 9, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Hajek, A.E.; Longcore, J.E.; Simmons, D.R.; Peters, K.; Humber, R.A. Chytrid mycoparasitism of entomophthoralean azygospores. J. Invertebr. Pathol. 2013, 114, 333–336. [Google Scholar] [CrossRef]
- Letcher, P.M.; Velez, C.G.; Schultz, S.; Powell, M.J. New taxa are delineated in Alphamycetaceae (Rhizophydiales, Chytridiomycota). Nova Hedwig. 2012, 94, 9–29. [Google Scholar] [CrossRef]
- Rosa, L.H.; Coelho, L.C.; Pinto, O.H.B.; Carvalho-Silva, M.; Convey, P.; Rosa, C.A.; Câmara, P.E.A.S. Ecological succession of fungal and bacterial communities in Antarctic mosses affected by a fairy ring disease. Extremophiles 2021, 25, 471–481. [Google Scholar] [CrossRef]
- McRae, C.F.; Hocking, A.D.; Seppelt, R.D. Penicillium species from terrestrial habitats in the Windmill Islands, east Antarctica, including a new species, Penicillium antarcticum. Polar Biol. 1999, 21, 97–111. [Google Scholar] [CrossRef]
- Godinho, V.M.; Gonçalves, V.N.; Santiago, I.F.; Figueredo, H.M.; Vitoreli, G.A.; Schaefer, C.E.G.R.; Barbosa, E.C.; Oliveira, J.G.; Alves, T.M.A.; Zani, C.L.; et al. Diversity and bioprospection of fungal community present in oligotrophic soil of continental Antarctica. Extremophiles 2015, 19, 585–596. [Google Scholar] [CrossRef]
- Zucconi, L.; Selbmann, L.; Buzzini, P.; Turchetti, B.; Guglielmin, M.; Frisvad, J.C.; Onofri, S. Searching for eukaryotic life preserved in Antarctic permafrost. Polar Biol. 2012, 35, 749–757. [Google Scholar] [CrossRef]
- da Silva, T.H.; Silva, D.A.S.; de Oliveira, F.S.; Schaefer, C.E.G.R.; Rosa, C.A.; Rosa, L.H. Diversity, distribution, and ecology of viable fungi in permafrost and active layer of Maritime Antarctica. Extremophiles 2020, 24, 565–576. [Google Scholar] [CrossRef]
- de Menezes, G.C.A.; Câmara, P.E.A.S.; Pinto, O.H.B.; Carvalho-Silva, M.; Oliveira, F.S.; Schaefer, C.D.R.; Convey, P.; Rosa, C.A.; Rosa, L.H. (Fungal diversity present on rocks from a polar desert in continental Antarctica assessed using DNA metabarcoding. Extremophiles 2021, 25, 193–202. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Pinto, O.H.B.; Vieira, R.; Neto, A.A.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Fungi present in Antarctic deep-sea sediments assessed using DNA metabarcoding. Microb. Ecol. 2021, 82, 157–164. [Google Scholar] [CrossRef]
- Rosa, L.H.; de Menezes, G.C.A.; Pinto, O.H.B.; Convey, P.; Carvalho-Silva, M.; Simões, J.C.; Rosa, C.A.; Câmara, P.E.A.S. Fungal diversity in seasonal snow of Martel Inlet, King George Island, South Shetland Islands, assessed using DNA metabarcoding. Polar Biol. 2022, 45, 627–636. [Google Scholar] [CrossRef]
- de Souza, L.M.D.; Lirio, J.M.; Coria, S.H.; Lopes, F.A.C.; Convey, P.; Carvalho-Silva, M.; Oliveira, F.S.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Diversity, distribution and ecology of fungal communities present in Antarctic lake sediments uncovered by DNA metabarcoding. Sci. Rep. 2022, 12, 8407. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.H.; Ogaki, M.B.; Lirio, J.M.; Vieira, R.; Coria, S.H.; Pinto, O.H.B.; Carvalho-Silva, M.; Convey, P.; Rosa, C.A.; Câmara, P.E.A.S. Fungal diversity in a sediment core from climate change impacted Boeckella Lake, Hope Bay, north-eastern Antarctic Peninsula assessed using metabarcoding. Extremophiles 2022, 26, 16. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [PubMed]
- Golzar, H.; Thomas, G.; Jayasena, K.W. Neoascochyta species cause leaf scorch on wheat in Australia. Australas. Plant Dis. Notes 2019, 914, 1. [Google Scholar] [CrossRef]
- Gonçalves, V.N.; Vaz, A.B.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of fungal communities in lakes of Antarctica. FEMS Microbiol. Ecol. 2012, 82, 459–471. [Google Scholar] [CrossRef]
- Rosa, L.H.; Pinto, O.H.B.; Šantl-Temkiv, T.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S. DNA metabarcoding of fungal diversity in air and snow of Livingston Island, South Shetland Islands, Antarctica. Sci. Rep. 2020, 10, 21793. [Google Scholar] [CrossRef]
- de Souza, L.D.M.; Ogaki, M.B.; Câmara, P.E.A.S.; Pinto, O.H.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Rosa, L.H. Assessment of fungal diversity present in lakes of Maritime Antarctica using DNA metabarcoding: A temporal microcosm experiment. Extremophiles 2021, 25, 77–84. [Google Scholar] [CrossRef]
- da Silva, T.H.; Câmara, P.E.A.S.; Pinto, O.H.B.; Carvalho-Silva, M.; Oliveira, F.S.; Convey, P.; Rosa, C.A.; Rosa, L.H. Diversity of fungi present in permafrost in the South Shetland Islands, maritime Antarctic. Microb. Ecol. 2022, 83, 58–67. [Google Scholar] [CrossRef]
- Schütte, U.M.; Henning, J.A.; Ye, Y.; Bowling, A.; Ford, J.; Genet, H.; Waldrop, M.P.; Turetsky, M.R.; White, J.R.; Bever, J.D. Effect of permafrost thaw on plant and soil fungal community in a boreal forest: Does fungal community change mediate plant productivity response? J. Ecol. 2019, 107, 1737–1752. [Google Scholar] [CrossRef]
- Al-Tohamy, R.; Kenawy, E.R.; Sun, J.; Ali, S.S. Performance of a newly isolated salt-tolerant yeast strain Sterigmatomyces halophilus SSA-1575 for azo dye decolorization and detoxification. Front. Microbiol. 2020, 11, 1163. [Google Scholar] [CrossRef]
- Fleet, G.; Prakitchaiwattana, C.; Beh, A.; Heard, G. The yeast ecology of wine grapes. In Biodiversity and Biotechnology of Wine Yeasts; Ciani, M., Ed.; Research Signpost: Kerala, India, 2022; pp. 1–17. [Google Scholar]
- Karpov, S.A.F.L.; Alacid, E. Dinomyces arenysensis gen. et sp. nov. (Rhizophydiales, Dinomycetaceae fam. nov.), a chytrid infecting marine dinoflagellates. Protist 2014, 165, 230–244. [Google Scholar]
- Li, A.H.; Yan, F.X.; Groenewald, M. Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: Proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud. Mycol. 2020, 96, 17–140. [Google Scholar] [CrossRef]
- Ming, C.; Huang, J.; Wang, Y. Revision of the medically relevant species of the yeast genus Diutina. Med. Mycol. 2019, 57, 226–233. [Google Scholar] [CrossRef]
- Pykälä, J.; Kantelinen, A.; Myllys, L. Taxonomy of Verrucaria species characterized by large spores, perithecia leaving pits in the rock and a pale thin thallus in Finland. MycoKeys 2020, 72, 43. [Google Scholar] [CrossRef]
- Sampaio, J.P. Leucosporidium Fell, Statzell, IL Hunter & Phaff (1969). In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1485–1494. [Google Scholar]
- Saubin, M.; Devillers, H.; Proust, L. Investigation of genetic relationships between Hanseniaspora species found in grape musts revealed interspecific hybrids with dynamic genome structures. Front. Microbiol. 2020, 10, 2960. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D. Endophytic fungi VI. Ciliophora quercus sp. nov. from China. Nova Hedwigia 2007, 85, 403–406. [Google Scholar] [CrossRef]



{kind=link}
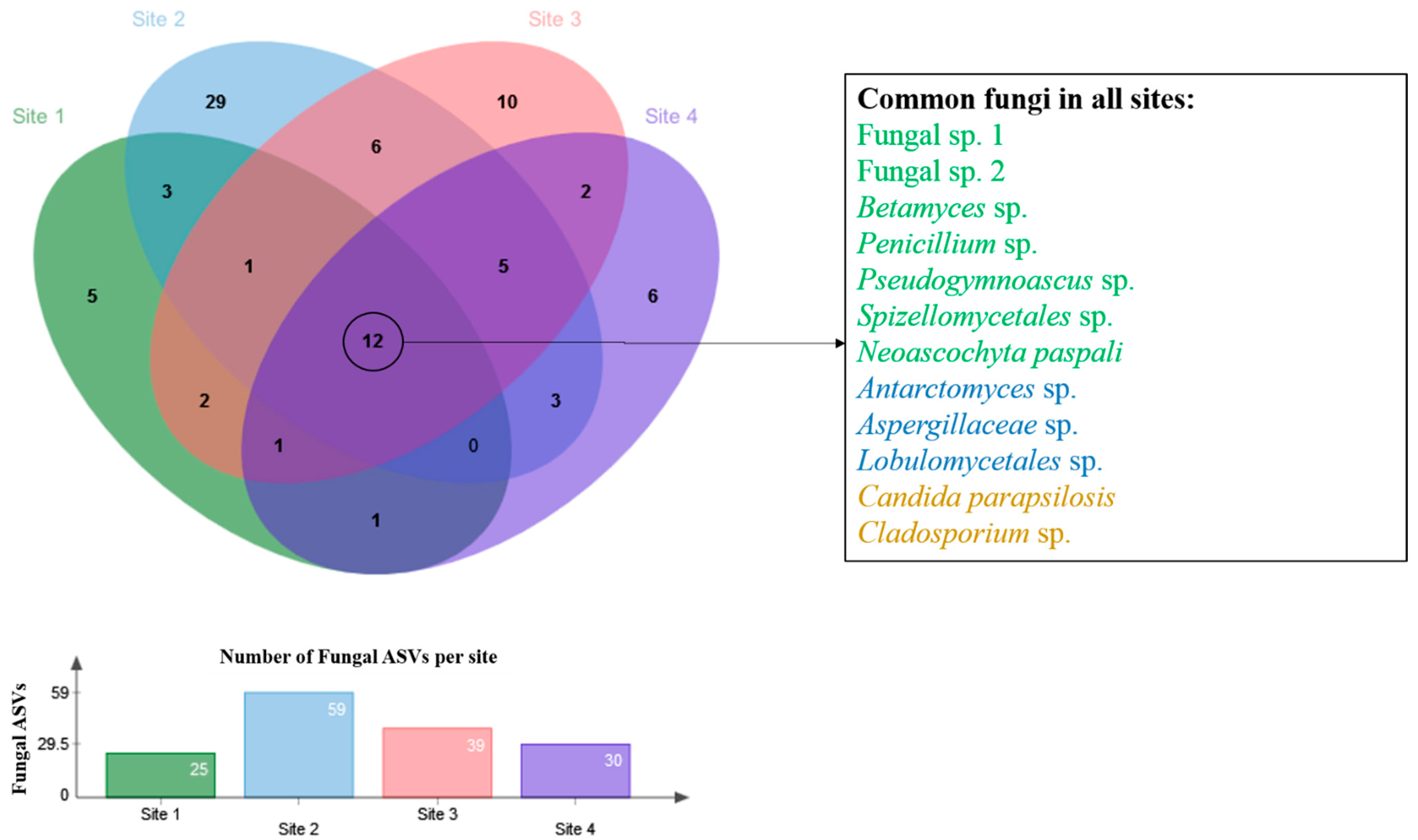{kind=link}
| Clearwater Mesa Soils | ||||
|---|---|---|---|---|
| Parameters | Site 1 | Site 2 | Site 3 | Site 4 |
| Geology | ||||
| Location | 64°01′16.5″ S; 57°43′49.6″ W | 64°01′53.5″ S; 57°40′47.0″ W | 64°02′03.6″ S; 57°40′05.9″ W | 64°02′09.5″ S; 57°39′22.1″ W |
| Altitude (meters above sea level) | 247 | 182 | 186 | 201 |
| Distance to coastline (m) | 650 | 3350 | 3990 | 4720 |
| Sampling site characteristics | Located in patterned ground over basaltic lava flow | Located in patterned ground over basaltic hyaloclastite breccias | Located in elongated patterned ground, very rich in clay sediment, formed over basaltic hyaloclastite breccias | Located in basal till deposit composed mainly by basaltic angular clast, with incipient formation of patterned ground, over basaltic hyaloclastite breccias |
| Sediment physical parameters | ||||
| Clay (%) | 25 | 21 | 14 | 24 |
| Silt | 26 | 31 | 37 | 20 |
| Coarse sand | 25 | 27 | 21 | 28 |
| Fine sand | 24 | 20 | 28 | 22 |
| Textural class | Clay-sand-loam | Loam | Loam | Loam |
| Sediment chemical parameters | ||||
| pH in H2O | 8.8 | 8.3 | 8.9 | 7.6 |
| Exchangeable P—mg dm3 | 103.2 | 13.6 | 6.7 | 10.7 |
| Sum of exchangeable bases Ca+K+Mg (SB)—cmolc dm3 | 8.81 | 4.54 | 6.54 | 8.75 |
| Percentage of base saturation (PBS)-% | 100.0 | 100.0 | 100.0 | 89.8 |
| H+Al—potential acidity—cmolc dm3 | 0.00 | 0.00 | 0.00 | 0.99 |
| Cation exchange capacity at pH 7 (CEC)—cmolc dm3 | 8.81 | 4.54 | 6.54 | 9.74 |
| Total organic carbon (TOC)—dag kg−1 | 0.7 | 0.7 | 0.93 | 0.54 |
| Micronutrient Fe—mg dm3-1 | 214.2 | 14.3 | 41.0 | 14.8 |
| Micronutrient Mn—mg dm3-1 | 72.1 | 27.4 | 58.7 | 24.1 |
| Fungal diversity indices | ||||
| Number of DNA reads | 15,636 | 56,473 | 38,575 | 17,090 |
| Number of taxa assigned | 25 | 59 | 39 | 31 |
| Fisher’s-α (diversity) | 10.7 | 60.43 | 23.51 | 15.39 |
| Margalef (richness) | 5.21 | 12.6 | 8.25 | 6.51 |
| Simpson’s (dominance) | 0.81 | 0.83 | 0.84 | 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonçalves, V.N.; Lirio, J.M.; Coria, S.H.; Lopes, F.A.C.; Convey, P.; de Oliveira, F.S.; Carvalho-Silva, M.; Câmara, P.E.A.S.; Rosa, L.H. Soil Fungal Diversity and Ecology Assessed Using DNA Metabarcoding along a Deglaciated Chronosequence at Clearwater Mesa, James Ross Island, Antarctic Peninsula. Biology 2023, 12, 275. https://doi.org/10.3390/biology12020275
Gonçalves VN, Lirio JM, Coria SH, Lopes FAC, Convey P, de Oliveira FS, Carvalho-Silva M, Câmara PEAS, Rosa LH. Soil Fungal Diversity and Ecology Assessed Using DNA Metabarcoding along a Deglaciated Chronosequence at Clearwater Mesa, James Ross Island, Antarctic Peninsula. Biology. 2023; 12(2):275. https://doi.org/10.3390/biology12020275
Chicago/Turabian StyleGonçalves, Vivian N., Juan M. Lirio, Silvia H. Coria, Fabyano A. C. Lopes, Peter Convey, Fábio S. de Oliveira, Micheline Carvalho-Silva, Paulo E. A. S. Câmara, and Luiz H. Rosa. 2023. "Soil Fungal Diversity and Ecology Assessed Using DNA Metabarcoding along a Deglaciated Chronosequence at Clearwater Mesa, James Ross Island, Antarctic Peninsula" Biology 12, no. 2: 275. https://doi.org/10.3390/biology12020275