Effects of the Density of Invasive Lantana camara Plants on the Biodiversity of Large and Small Mammals in the Groenkloof Nature Reserve (GNR) in South Africa

Abstract
:Simple Summary
Abstract
1. Introduction
Problem
2. Materials and Methods
2.1. Study Area
2.2. Site Selection
2.3. Treatment Design
2.4. Mammal Sampling
2.4.1. Large Mammals
2.4.2. Small Mammals
2.4.3. Mammal Identifications
2.5. Statistical Analysis
2.5.1. Large Mammals
2.5.2. Small Mammals
3. Results
3.1. Large Mammals Results
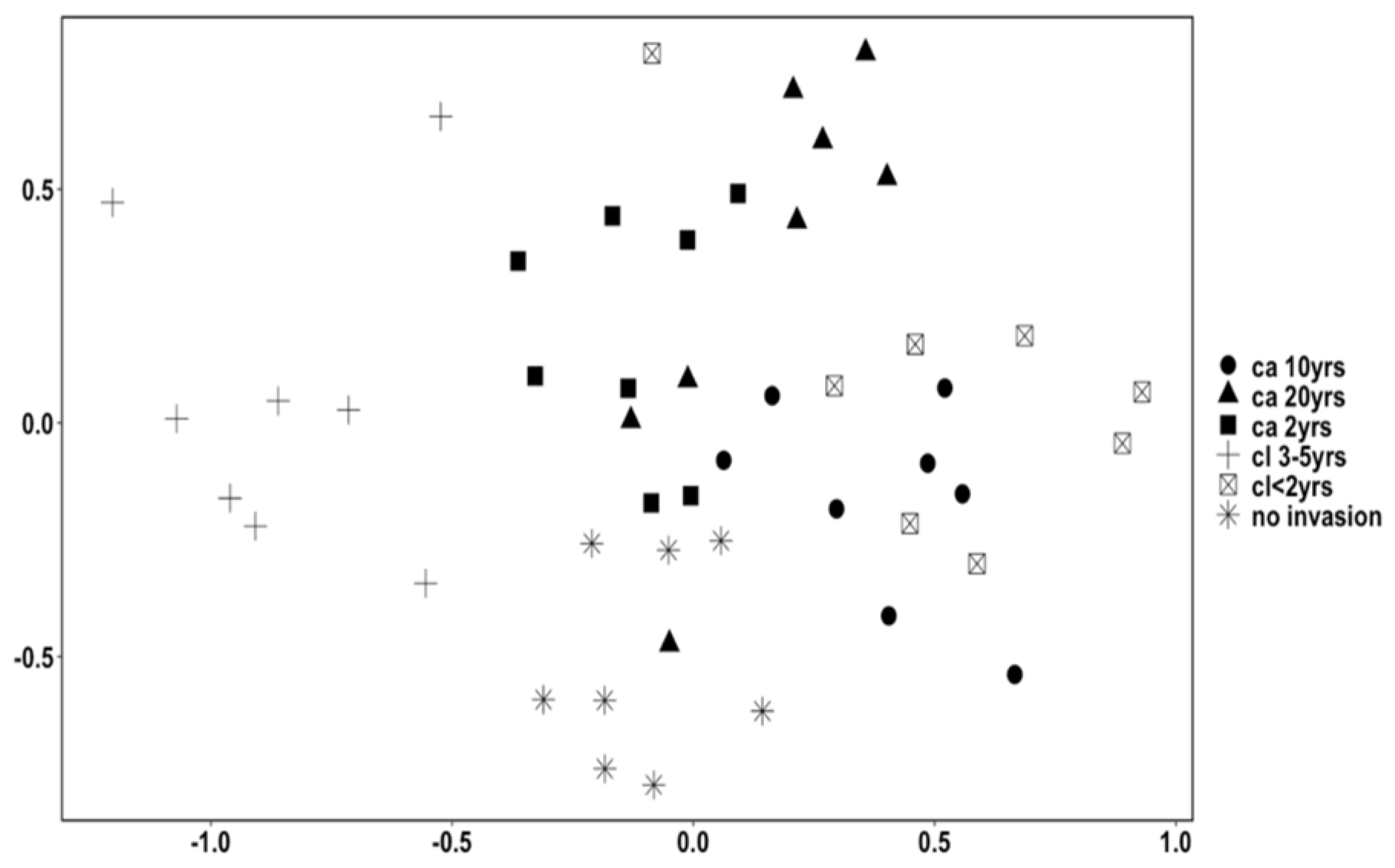
3.1.1. Differences between Treatments
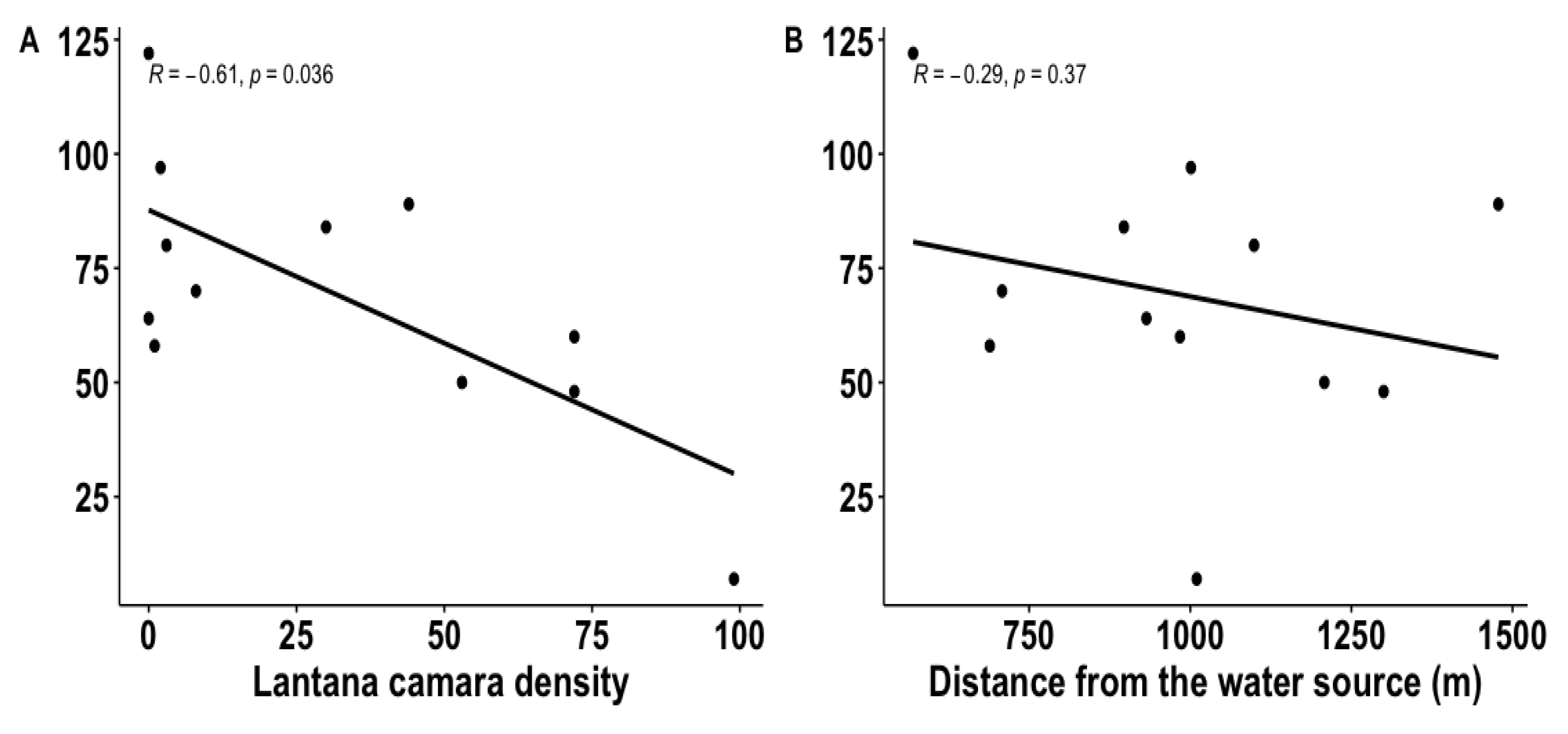
3.1.2. Treatments’ Proximity to the Water Source and L. Camara Density
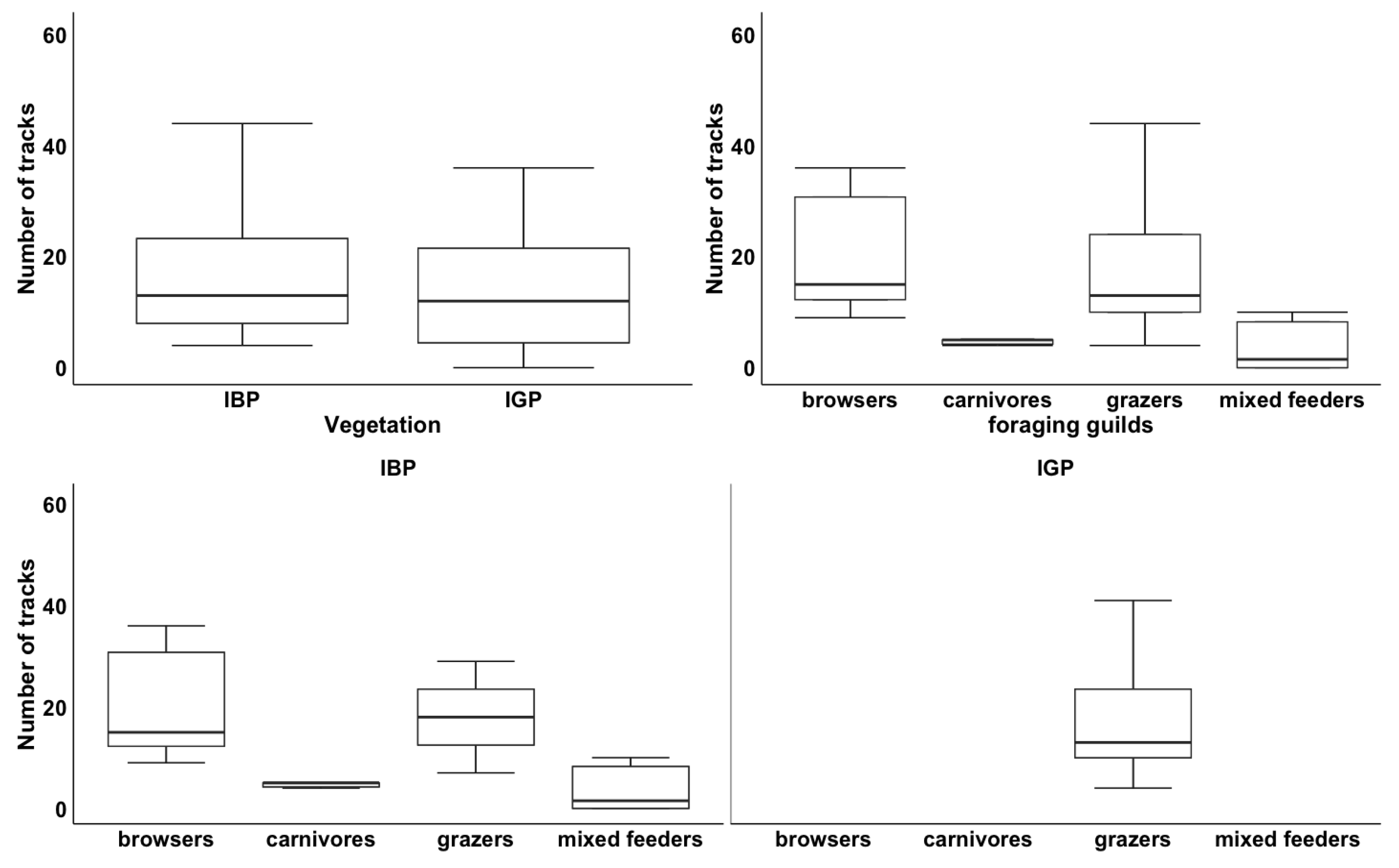
3.1.3. Variation in Species Richness by Vegetation Type and Foraging Guilds
3.2. Small Mammals Results
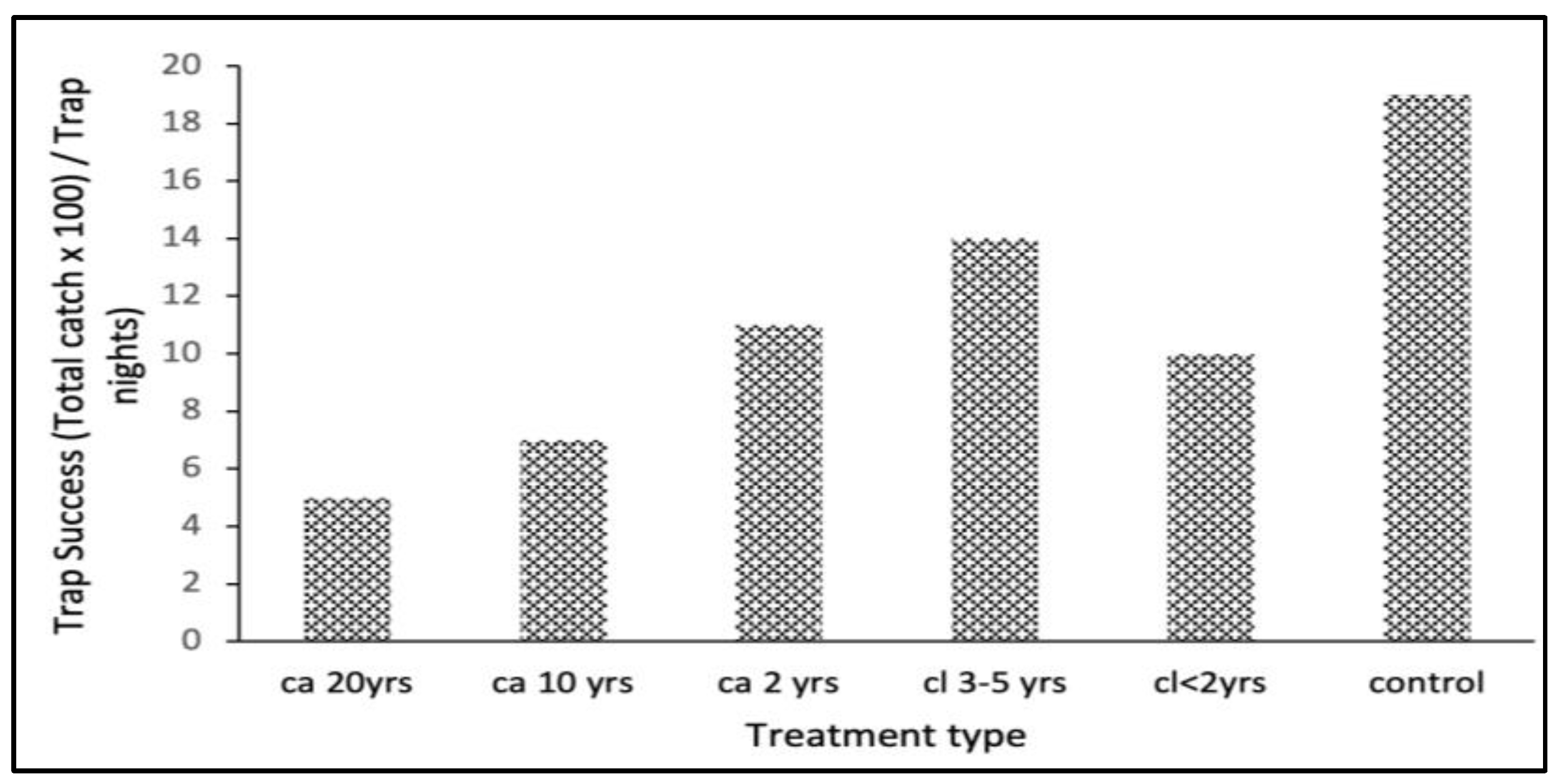
3.2.1. Rodents Trapping
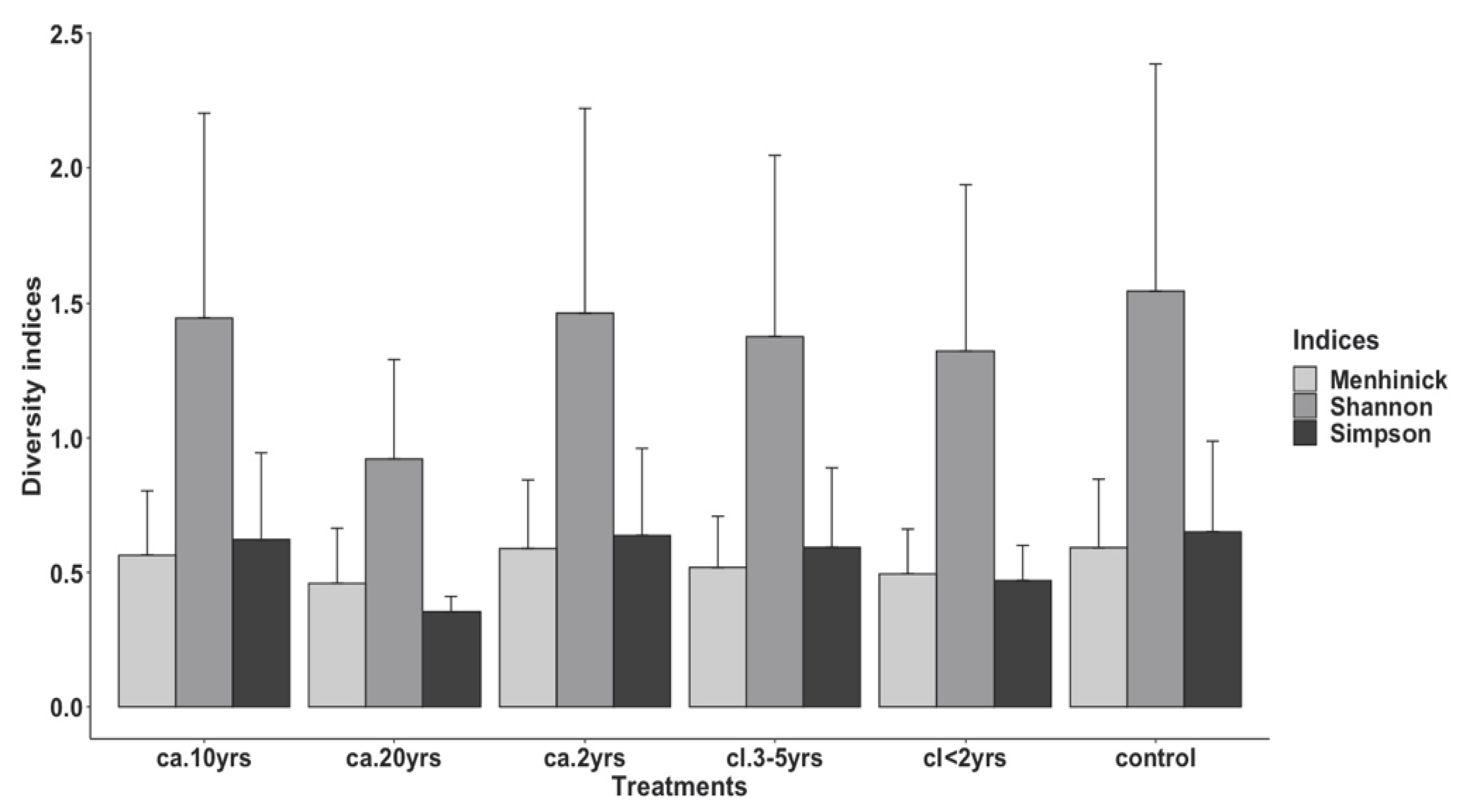
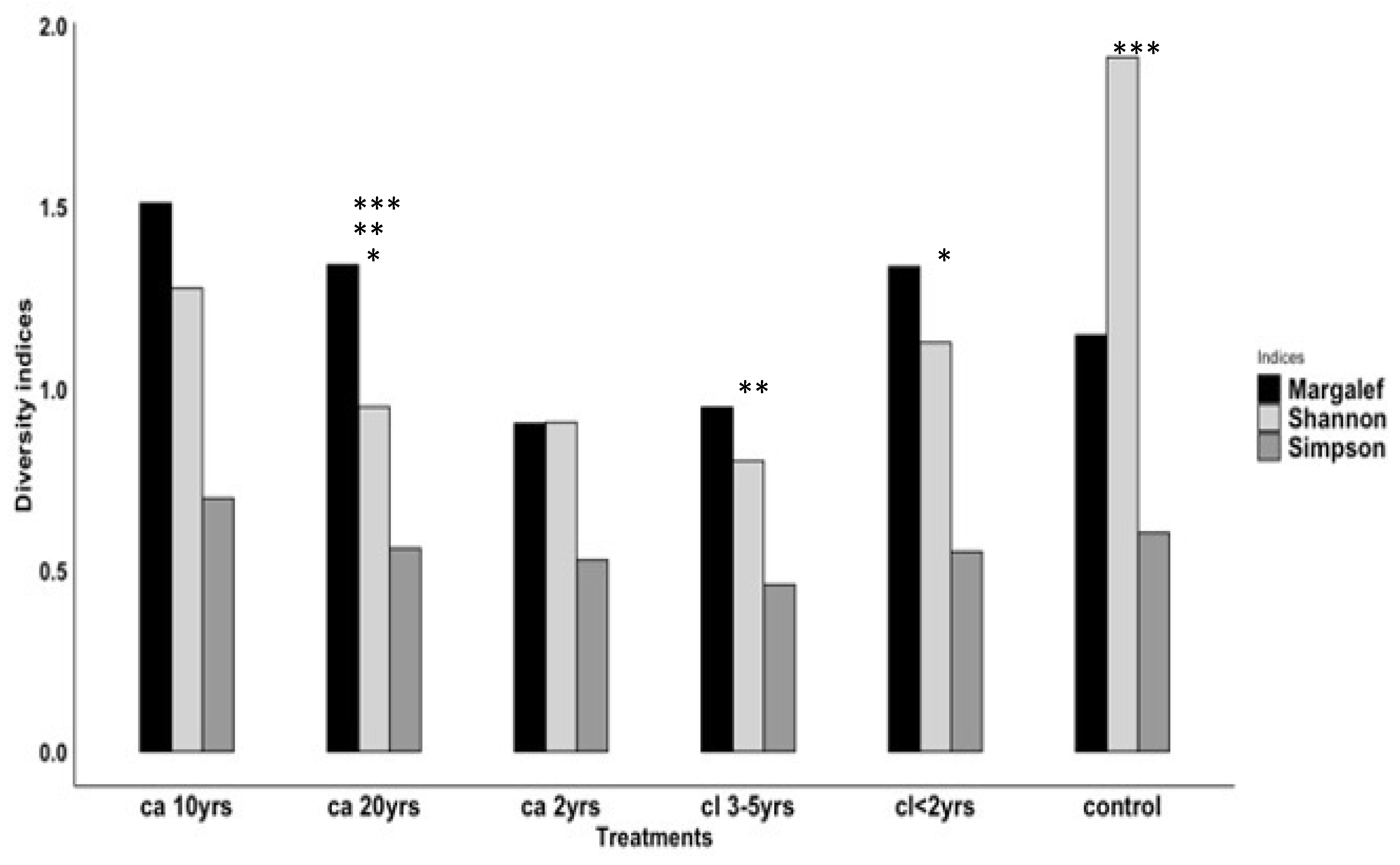
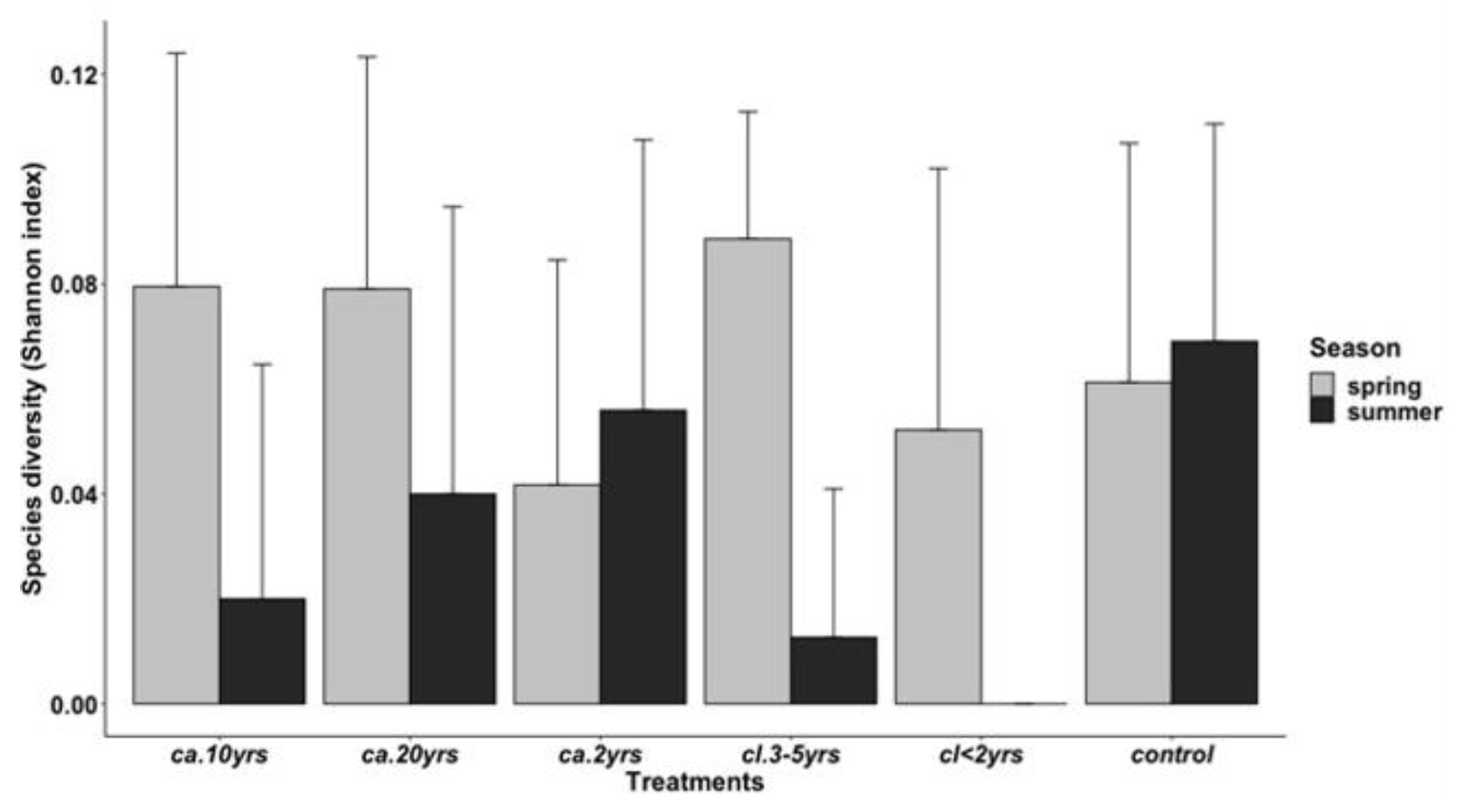
3.2.2. Species Diversity by Treatments and Season
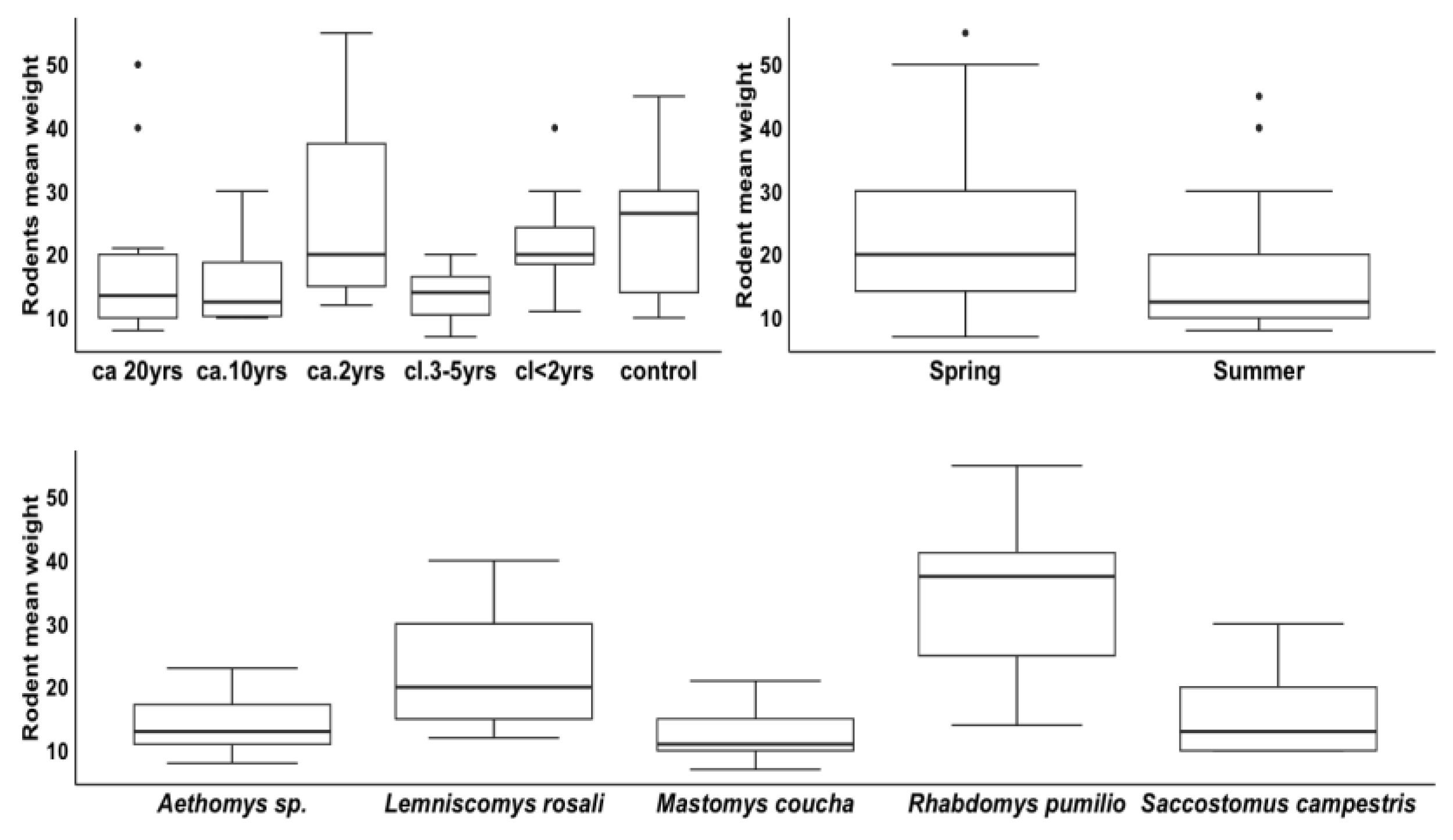
3.2.3. Rodent Weight by Seasons across Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cilliers, C.J.; Neser, S. Biological control of Lantana camara (Verbenaceae) in South Africa. Agric. Ecosyst. Environ. 1991, 37, 57–75. [Google Scholar] [CrossRef]
- De Lacy, P.; Shackleton, C.M. Woody plant species richness, composition, and structure in urban sacred sites, Grahamstown, South Africa. Urban Ecosyst. 2017, 20, 1169–1179. [Google Scholar] [CrossRef]
- Waits, L.P.; Paetkau, D. Non-invasive genetic sampling tools for wildlife biologists: A review of applications and recommendations for accurate data collection. J. Wildl. Manag. 2005, 69, 1419–1433. [Google Scholar] [CrossRef]
- Van Wilgen, B.W.; Reyers, B.; Le Maitre, D.C.; Richardson, D.M.; Schonegevel, L. A biome-scale assessment of the impact of invasive alien plants on ecosystem services in South Africa. J. Environ. Manag. 2016, 89, 336–349. [Google Scholar] [CrossRef]
- Howison, R.A.; Olff, H.; van de Koppel, J.; Smit, C. Biotically driven vegetation mosaics in grazing ecosystems: The battle between bioturbation and biocompaction. Ecol. Monogr. 2017, 87, 363–378. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Skinner, J.D.; Chimimba, C.T. The Mammals of the Southern African Sub-Region, 3rd ed.; Skinner, J.D., Chimimba, C.T., Eds.; Cambridge University Press: Pretoria, South Africa, 2005. [Google Scholar]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Sisk, T.D.; Launer, A.E.; Switky, K.R.; Ehrlich, P.R. Identifying extinction threats: Global analyses of the distribution of biodiversity and the expansion of the human enterprise. In Ecosystem Management; Springer: New York, NY, USA, 1994; pp. 53–68. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Amin, R.; Baral, H.S.; Lamichhane, B.R.; Poudyal, L.P.; Lee, S.; Jnawali, S.R.; Acharya, K.P.; Upadhyaya, G.P.; Pandey, M.B.; Shrestha, R.; et al. The status of Nepal’ s mammals. J. Threat. Taxa 2018, 10, 11361–11378. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Rylands, A.B.; Konstant, W.R.; Flick, P.; Pilgrim, J.; Oldfield, S.; Magin, G.; et al. Habitat loss and extinction in the hotspots of biodiversity. Conserv. Biol. 2002, 16, 909–923. [Google Scholar] [CrossRef] [Green Version]
- Ewel, J.J.; O’Dowd, D.J.; Bergelson, J.; Daehler, C.C.; D’Antonio, C.M.; Gómez, L.D.; Gordon, D.R.; Hobbs, R.J.; Holt, A.; Hopper, K.R.; et al. Deliberate introductions of species: Research needs: Benefits can be reaped, but risks are high. BioScience 1999, 49, 619–630. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Invasion of deciduous forest preserves in the New York metropolitan region by Japanese barberry (Berberis thunbergii DC.). J. Torrey Bot. Soc. 1997, 124, 210–215. [Google Scholar] [CrossRef]
- Dumalisile, L.; Somers, M.J. The effects of an invasive alien plant (Chromolaena odorata) on large African mammals. Nat. Conserv. Res. 2017, 2, 102–108. [Google Scholar] [CrossRef]
- Ramaswami, G.; Sukumar, R. Invasive plants in the tropics and the Case of Lantana camara. In Tropical Conservation: Perspectives on Local and Global Priorities; Aguirre, A.A., Sukumar, R., Eds.; Oxford University Press: New York, NY, USA, 2016; p. 154. [Google Scholar]
- Maluleke, X.G. Modeling and Explaining the Distribution of Lantana camara in South African Savanna Ecosystems. Ph.D. Thesis, University of KwaZulu-Natal, Durban, South Africa, 2019. [Google Scholar]
- Lasmar, C.J.; Rosa, C.; Queiroz, A.; Nunes, C.A.; Imata, M.M.; Alves, G.P.; Nascimento, G.B.; Ázara, L.N.; Vieira, L.; Louzada, J.; et al. Temperature and productivity distinctly affect the species richness of ectothermic and endothermic multitrophic guilds along a tropical elevational gradient. Oecologia 2021, 197, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Paredes, O.S.L.; Norris, D.; Oliveira, T.G.D.; Michalski, F. Water availability not fruitfall modulates the dry season distribution of frugivorous terrestrial vertebrates in a lowland Amazon forest. PLoS ONE 2017, 12, e0174049. [Google Scholar] [CrossRef]
- Weidlich, E.W.; Flórido, F.G.; Sorrini, T.B.; Brancalion, P.H. Controlling invasive plant species in ecological restoration: A global review. J. Appl. Ecol. 2020, 57, 1806–1817. [Google Scholar] [CrossRef]
- Mills, J.N.; Childs, J.E.; Ksiazek, T.G.; Peters, C.J.; Velleca, W.M. Methods for Trapping and Sampling Small Mammals for Virologic Testing; U.S. Department of Health & Human Services: Atlanta, GA, USA, 1995.
- Kuiper, T.R.; Parker, D.M. Grass height is the determinant of sheep grazing effects on small mammals in a savanna ecosystem. Rangel. J. 2013, 35, 403–408. [Google Scholar] [CrossRef]
- Osborn, T.C.; Butrulle, D.V.; Sharpe, A.G.; Pickering, K.J.; Parkin, I.A.; Parker, J.S.; Lydiate, D.J. Detection and effects of a homeologous reciprocal transposition in Brassica napus. Genetics 2003, 165, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Ceradini, J.P.; Chalfoun, A.D. Species’ traits help predict small mammal responses to habitat homogenization by an invasive grass. Ecol. Appl. 2017, 27, 1451–1465. [Google Scholar] [CrossRef]
- Haack, N.; Grimm-Seyfarth, A.; Schlegel, M.; Wirth, C.; Bernhard, D.; Brunk, I.; Henle, K. Patterns of richness across forest beetle communities—A methodological comparison of observed and estimated species numbers. Ecol. Evol. 2021, 11, 626–635. [Google Scholar] [CrossRef]
- McHugh, D.; Goldingay, R.L.; Parkyn, J.; Goodwin, A.; Letnic, M. Short-term response of threatened small macropods and their predators to prescribed burns in subtropical Australia. Ecol. Manag. Restor. 2020, 21, 97–107. [Google Scholar] [CrossRef]
- Apolloni, N.; Grüebler, M.U.; Arlettaz, R.; Gottschalk, T.K.; Naef-Daenzer, B. Habitat selection and range use of little owls in relation to habitat patterns at three spatial scales. Anim. Conserv. 2018, 21, 65–75. [Google Scholar] [CrossRef]
- Austad, S.N.; Kristan, D.M. Are mice calorically restricted in nature? Aging Cell 2003, 2, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Malo, A.F.; Godsall, B.; Prebble, C.; Grange, Z.; McCandless, S.; Taylor, A.; Coulson, T. Positive effects of an invasive shrub on aggregation and abundance of a native small rodent. Behav. Ecol. 2013, 24, 759–767. [Google Scholar] [CrossRef]
- Van Wyk, J. Groenkloof Nature Reserve The oldest in Africa? Afr. Wildl. Eppindust- 2004, 58, 10–14. [Google Scholar]
- Tauoatsoala, P.; Coetzee, W.J.L.; Hermann, U.P. Perceptions of visitors on sustainable tourism practices in an urban nature reserve: The case of Groenkloof Nature Reserve, South Africa. Afr. J. Phys. Act. Health Sci. 2017, 23, 608–620. [Google Scholar]
- Eckhardt, H.C.; Van Rooyen, N.; Bredenkamp, G.J. Plant communities and species richness of the Agrostis lachnantha–Eragrostis plana wetlands of northern KwaZulu-Natal. S. Afr. J. Bot. 1996, 62, 306–315. [Google Scholar] [CrossRef]
- Grobler, C.H. The Vegetation Ecology of Urban Open Spaces in Gauteng. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2006. [Google Scholar]
- Kamffer, D. ECOCHECK. 2006. Available online: https://sahris.sahra.org.za/sites/default/files/additionaldocs/Appendix%20D-%20Biodiversity%20EIA%20Report%28EIA%29.pdf (accessed on 10 February 2023).
- Rovero, F.; Martin, E.; Rosa, M.; Ahumada, J.A.; Spitale, D. Estimating species richness and modeling habitat preferences of tropical forest mammals from camera trap data. PLoS ONE 2014, 9, e103300. [Google Scholar] [CrossRef]
- Espartosa, K.D.; Pinotti, B.T.; Pardini, R. Performance of camera trapping and track counts for surveying large mammals in rainforest remnants. Biodivers. Conserv. 2011, 20, 2815–2829. [Google Scholar] [CrossRef]
- Schwarz, C.J.; Seber, G.A. Estimating animal abundance: Review III. Stat. Sci. 1999, 14, 427–456. [Google Scholar] [CrossRef]
- Suryawanshi, K.R.; Bhatnagar, Y.V.; Mishra, C. Standardizing the double-observer survey method for estimating mountain ungulate prey of the endangered snow leopard. Oecologia 2012, 169, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C. Field Guide to Tracks and Signs of Southern, Central, and East African Wildlife; Penguin Random House South Africa: Cape Town, South Africa, 2013. [Google Scholar]
- Conway, T.M.; Bourne, K.S. A comparison of neighborhood characteristics related to canopy cover, stem density, and species richness in an urban forest. Landsc. Urban Plan. 2013, 113, 10–18. [Google Scholar] [CrossRef]
- Aurélio, M.; Faleiro, F.; Lopes, V.M.; Pires, V.; Lopes, A.R.; Pimentel, M.S.; Repolho, T.; Baptista, M.; Narciso, L.; Rosa, R. Physiological and behavioral responses of temperate seahorses (Hippocampus guttulatus) to environmental warming. Mar. Biol. 2013, 160, 2663–2670. [Google Scholar] [CrossRef]
- Brehm, A.M.; Tironi, S.; Mortelliti, A. Effects of trap confinement on personality measurements in two terrestrial rodents. PLoS ONE 2020, 15, e0221136. [Google Scholar]
- Dumalisile, L. Effects of Chromolaena odorata on Mammalian Biodiversity in Hluhluwe-iMfolozi Park, South Africa. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2009. [Google Scholar]
- Jenkins, K.J.; Manly, B.F.J. A double-observer method for reducing bias in fecal pellet surveys of forest ungulates. J. Appl. Ecol. 2008, 45, 1339–1348. [Google Scholar] [CrossRef]
- Twigg, G.I. Marking mammals by tissue removal. In Animal Marking; Palgrave: London, UK, 1978; pp. 109–118. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Harper and Row: New York, NY, USA, 1989. [Google Scholar]
- Magnussen, S.; Nord-Larsen, T. A jack-knife estimator of variance for a random tessellated stratified sampling design. For. Sci. 2019, 65, 543–547. [Google Scholar]
- Symonds, M.R.; Moussalli, A. A brief guide to model selection, multimodel inference, and model averaging in behavioral ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- De Winton, W.E. XXXV.—On a collection of rodents from Angola. J. Nat. Hist. 1897, 20, 320–324. [Google Scholar] [CrossRef]
- Avery, D.M.; Avery, G. Micromammals in the Northern Cape Province of South Africa, past and present. Afr. Nat. Hist. 2011, 7, 9–39. [Google Scholar]
- Dickman, C.R. Habitat fragmentation and vertebrate species richness in an urban environment. J. Appl. Ecol. 1987, 24, 337–351. [Google Scholar] [CrossRef]
- Lamprey, H.F. Ecological separation of the large mammal species in the Tarangire game reserve, Tanganyika. Afr. J. Ecol. 1963, 1, 63–92. [Google Scholar] [CrossRef]
- O’Brien, C.S.; Waddell, R.B.; Rosenstock, S.S.; Rabe, M.J. Wildlife use of water catchments in southwestern Arizona. Wildl. Soc. Bull. 2006, 34, 582–591. [Google Scholar] [CrossRef]
- Kochenderfer, J.D.; Kochenderfer, J.N.; Miller, G.W. Controlling beech root and stump sprouts using the cut-stump treatment. North. J. Appl. For. 2006, 23, 155–165. [Google Scholar] [CrossRef]
- Schirmel, J.; Bundschuh, M.; Entling, M.H.; Kowarik, I.; Buchholz, S. Impacts of invasive plants on resident animals across ecosystems, taxa, and feeding types: A global assessment. Glob. Change Biol. 2016, 22, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Change Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Dutra, H.P.; Barnett, K.; Reinhardt, J.R.; Marquis, R.J.; Orrock, J.L. Invasive plant species alter consumer behavior by providing refuge from predation. Oecologia 2011, 166, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Mattos, K.J.; Orrock, J.L. Behavioral consequences of plant invasion: An invasive plant alters rodent antipredator behavior. Behav. Ecol. 2010, 21, 556–561. [Google Scholar] [CrossRef]
- Sampson, C.; Leimgruber, P.; Tonkyn, D.; Pastorini, J.; Janaka, H.K.; Sotherden, E.; Fernando, P. Effects of illegal grazing and invasive Lantana camara on Asian elephant habitat use. Biol. Conserv. 2018, 220, 50–59. [Google Scholar] [CrossRef]
- Te Beest, M.; Esler, K.J.; Richardson, D.M. Linking functional traits to impacts of invasive plant species: A case study. Plant Ecol. 2015, 216, 293–305. [Google Scholar] [CrossRef]
- Shannon, G.; Larson, C.L.; Reed, S.E.; Crooks, K.R.; Angeloni, L.M. Ecological consequences of ecotourism for wildlife populations and communities. In Ecotourism’s Promise and Peril; Springer: Cham, Switzerland, 2017; pp. 29–46. [Google Scholar]
- Do Linh San, E.; Malongwe, N.B.; Fike, B.; Somers, M.J.; Walters, M. A diverse autumn diet without dominant prey for opportunistic black-backed jackals Canis mesomelas. Wildl. Biol. Pract. 2009, 5, 96–103. [Google Scholar] [CrossRef]
- Plumptre, A.J.; Kirunda, B.; Mugabe, H.; Stabach, J.; Driciru, M.; Picton-Phillipps, G.; Ayebare, S.; Nangendo, G.; Laporte, N. The Impact of Fire and Large Mammals on the Ecology of Queen Elizabeth National Park; Wildlife Conservation Society and Woods Hole Research Centre: New York, NY, USA, 2010. [Google Scholar]
- Walton, C. Lantana camara (shrub); Department of Natural Resources, Queensland, Australia and IUCN/SSC, Invasive Species Specialist Group (ISSG). Available online: https://vital.seals.ac.za/vital/access/services/Download/vital:42893/SOURCE1 (accessed on 10 February 2023).
- Julius, R.S.; Schwan, E.V.; Chimimba, C.T. Helminth composition and prevalence of indigenous and invasive synanthropic murid rodents in urban areas of Gauteng Province, South Africa. J. Helminthol. 2018, 92, 445–454. [Google Scholar] [CrossRef]
- Taylor, P.J.; Munyai, A.; Gaigher, I.; Baxter, R. Afromontane small mammals do not follow the hump-shaped rule: Altitudinal variation in the Soutpansberg Mountains, South Africa. J. Trop. Ecol. 2015, 31, 37–48. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, J.; Chen, M.; Huang, X.; Xie, X.; Li, W.; Cao, Q.; Kan, H.; Xu, Y.; Ying, Z. Exposure to concentrated ambient PM 2.5 alters the composition of gut microbiota in a murine model. Part. Fiber Toxicol. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Hagenah, N. Among Rodents and Rhinos: Interplay between Small Mammals and Large Herbivores in a South African Savanna. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2006. [Google Scholar]
- Shackleton, R.T.; Witt, A.B.; Piroris, F.M.; van Wilgen, B.W. Distribution and socio-ecological impacts of the invasive alien cactus Opuntia stricta in eastern Africa. Biol. Invasions 2017, 19, 2427–2441. [Google Scholar] [CrossRef]
- MacDonald, I.A.W. Alien trees, shrubs and creepers invading indigenous vegetation in the Hluhluwe-Umfolozi Game Reserve Complex in Natal. Bothalia 1983, 14, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.F.; Frischknecht, N.C. Small mammals increase on recently cleared and seeded juniper rangeland. J. Range Manag. 1973, 26, 101–103. [Google Scholar] [CrossRef]
- Isabirye-Basuta, G.; Kasenene, J.M. Small rodent populations in selectively felled and mature tracts of Kibale Forest, Uganda. Biotropica 1987, 19, 260–266. [Google Scholar] [CrossRef]
- Negi, G.C.; Sharma, S.; Vishvakarma, S.C.; Samant, S.S.; Maikhuri, R.K.; Prasad, R.C.; Palni, L.M. Ecology and use of Lantana camara in India. Bot. Rev. 2019, 85, 109–130. [Google Scholar] [CrossRef]
- Akhter, M.H.; Mathur, M.; Bhide, N.K. Skin and liver toxicity in experimental Lantana camara poisoning in albino rats. Ind. J. Physiol. Pharmacol. 1990, 34, 13–16. [Google Scholar]
- Quirici, V.; Castro, R.A.; Ortiz-Tolhuysen, L.; Chesh, A.S.; Burger, J.R.; Miranda, E.; Cortés, A.; Hayes, L.D.; Ebensperger, L.A. Seasonal variation in the range areas of the diurnal rodent Octodon degus. J. Mammal. 2010, 91, 458–466. [Google Scholar] [CrossRef]
- Pour, B.M.; Latha, L.Y.; Sasidharan, S. Cytotoxicity and oral acute toxicity studies of Lantana camara leaf extract. Molecules 2011, 16, 3663–3674. [Google Scholar] [CrossRef] [PubMed]
- Schradin, C.; Pillay, N. Female striped mice (Rhabdomys pumilio) change their home ranges in response to seasonal variation in food availability. Behav. Ecol. 2006, 17, 452–458. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | Values and SI Units |
|---|---|
| Mean annual temperature | 18.7 °C–24.9 °C |
| Average Maximum Temperature recorded | 24.5 °C–36 °C |
| Mean Annual precipitation | 670 mm |
| Altitude | 1082 m–1899 m above sea level |
| Simper Pairwise Tests between Treatments | Test Statistics (R) | Significant level% |
|---|---|---|
| control—ca. 2 yrs | 0.941 | 0.1 * |
| control—ca. 10 yrs | 0.993 | 0.1 * |
| control—ca. 20 yrs | 0.873 | 0.1 * |
| control—cl. < 2 yrs | 1.000 | 0.1 * |
| control—cl. < 3–5 yrs | 0.806 | 0.1 * |
| ca. 2 yrs–ca. 10 yrs | 0.895 | 0.1 * |
| ca. 2 yrs–ca. 20 yrs | 0.883 | 0.1 * |
| ca. 2 yrs–cl. < 2 yrs | 0.998 | 0.1 * |
| ca. 2 yrs–cl. < 3–5 yrs | 0.88 | 0.1 * |
| ca. 10 yrs–ca. 20 yrs | 0.115 | 3.2 * |
| ca. 10 yrs–cl. < 2 yrs | 0.996 | 0.1 * |
| ca. 10 yrs–cl. < 3–5 yrs | 0.671 | 0.1 * |
| ca. 20 yrs–cl. < 2 yrs | 0.849 | 0.1 * |
| ca. 20 yrs–cl. 3–5 yrs | 0.531 | 0.1 * |
| cl. < 2 yrs–cl. 3–5 yrs | 0.621 | 0.1 * |
| Control | ca. 2 yrs | ca. 10 yrs | ca. 20 yrs | cl. < 2 yrs | |
|---|---|---|---|---|---|
| ca. 2 yrs | 0.211 | ||||
| ca. 10 yrs | 0.235 | 0.874 | |||
| ca. 20 yrs | 0.065 | 0.393 | 0.186 | ||
| cl. < 2 yrs | 0.019 * | 0.034 * | 0.051 | 0.187 | |
| cl. 3–5 yrs | 0.014 * | 0.019 * | 0.027 * | 0.056 | 0.335 |
| Variables | Estimate | Std Error | Z Value | p Value |
|---|---|---|---|---|
| Treatments—ca. 20 yrs | 0.040 | 0.057 | 0.709 | p = 0.478 |
| Treatments—ca. 2 yrs | 2.967 | 0.355 | 8.341 | p < 0.001 * |
| Treatments—cl. 3–5 yrs | 5.760 | 0.278 | 7.090 | p < 0.001 * |
| Treatments—cl. < 2 yrs | 7.078 | 0.954 | 74.163 | p < 0.001 * |
| Treatments—control | 6.552 | 0.139 | 46.848 | p < 0.001 * |
| Vegetation–IGP | 0.208 | 0.997 | 2.134 | p = 0.032 * |
| Parameters | Estimates | Std Error | Z Value | p Value |
|---|---|---|---|---|
| treatments—ca. 20 yrs | 1.099 | 5.774 × 10−1 | 1.903 | p = 0.061 |
| treatments—ca. 2 yrs | −2.340 × 101 | 6.965 × 104 | 0.000 | p = 0.040 * |
| treatments—cl. 3–5 yrs | 2.877 × 10−1 | 7.638 × 10−1 | 0.377 | p < 0.001 * |
| treatments—cl. < 2 yrs | −4.055 × 10−1 | 9.129 × 10 −1 | −0.444 | p = 0.001 * |
| treatments—control | −1.099 | 1.155 | −0.951 | p = 0.023 * |
| Variables | Estimate | Std Error | Z Value | p Value |
|---|---|---|---|---|
| Treatments: ca. 10 yrs | −0.211 | 0.113 | −1.855 | p = 0.063 |
| Treatments: ca. 2 yrs | 0.356 | 0.112 | 3.171 | p = 0.001 * |
| Treatments: cl. 3–5 yrs | −0.275 | 0.135 | −2.031 | p = 0.042 * |
| Treatments: cl. < 2 yrs | 0.188 | 0.117 | 1.600 | p = 0.109 |
| Treatments: control | 0.309 | 0.118 | 2.619 | p = 0.008 * |
| Season: summer | −0.356 | 0.161 | −2.205 | p = 0.027 * |
| Species: Lemniscomys rosalia | 0.390 | 0.135 | 2.873 | p = 0.004 * |
| Species: Mastomys coucha | −0.142 | 0.150 | −0.947 | p = 0.343 * |
| Species: Rhabdomys pumilio | 0.845 | 0.127 | 6.617 | p < 0.001 |
| Species: Saccostomus campestris | 0.042 | 0.146 | 0.293 | p = 0.769 * |
| Season: Summer—Species: Lemniscomys rosali | 0.172 | 0.201 | 0.855 | p = 0.392 |
| Season: Summer—Species: Mastomys coucha | 0.026 | 0.236 | 0.114 | p = 0.909 |
| Season: Summer—Species: Rhabdomys pumilio | 0.159 | 0.188 | 0.843 | p = 0.398 |
| Season: Summer—Species: Saccostomus campestris | 0.098 | 0.218 | 0.451 | p = 0.652 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raphela, T.D.; Duffy, K.J. Effects of the Density of Invasive Lantana camara Plants on the Biodiversity of Large and Small Mammals in the Groenkloof Nature Reserve (GNR) in South Africa. Biology 2023, 12, 296. https://doi.org/10.3390/biology12020296
Raphela TD, Duffy KJ. Effects of the Density of Invasive Lantana camara Plants on the Biodiversity of Large and Small Mammals in the Groenkloof Nature Reserve (GNR) in South Africa. Biology. 2023; 12(2):296. https://doi.org/10.3390/biology12020296
Chicago/Turabian StyleRaphela, Tlou D., and Kevin J. Duffy. 2023. "Effects of the Density of Invasive Lantana camara Plants on the Biodiversity of Large and Small Mammals in the Groenkloof Nature Reserve (GNR) in South Africa" Biology 12, no. 2: 296. https://doi.org/10.3390/biology12020296
APA StyleRaphela, T. D., & Duffy, K. J. (2023). Effects of the Density of Invasive Lantana camara Plants on the Biodiversity of Large and Small Mammals in the Groenkloof Nature Reserve (GNR) in South Africa. Biology, 12(2), 296. https://doi.org/10.3390/biology12020296