Each Personality Performs Its Own Function: Boldness and Exploration Lead to Differences in the Territoriality of Swimming Crabs (Portunus trituberculatus)

 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection and Maintenance
2.2. Behavior Acquisition and Quantification
2.2.1. Measurement and Classification of Boldness and Exploration
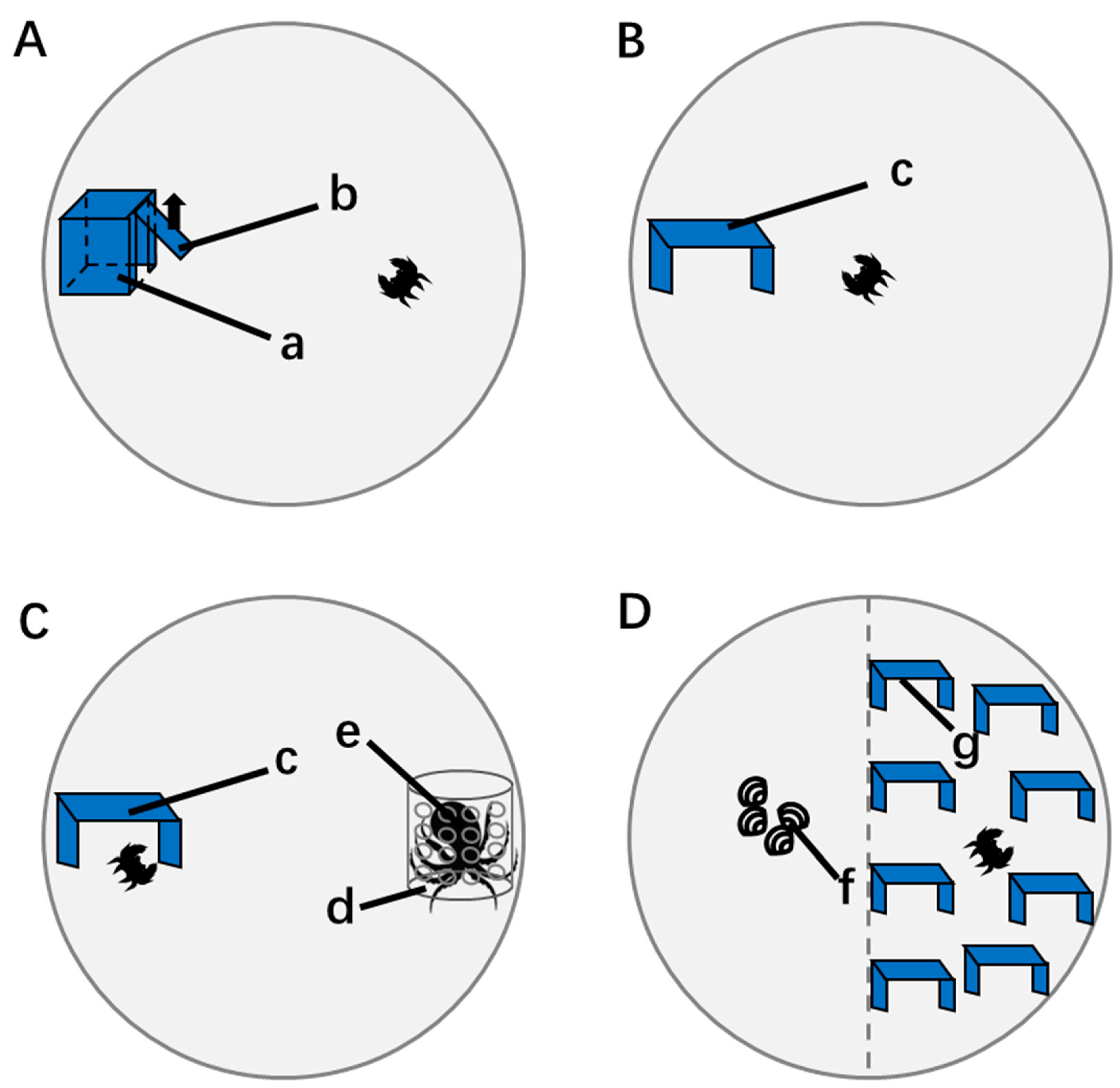
2.2.2. Behavior Measurement in a Safe Environment
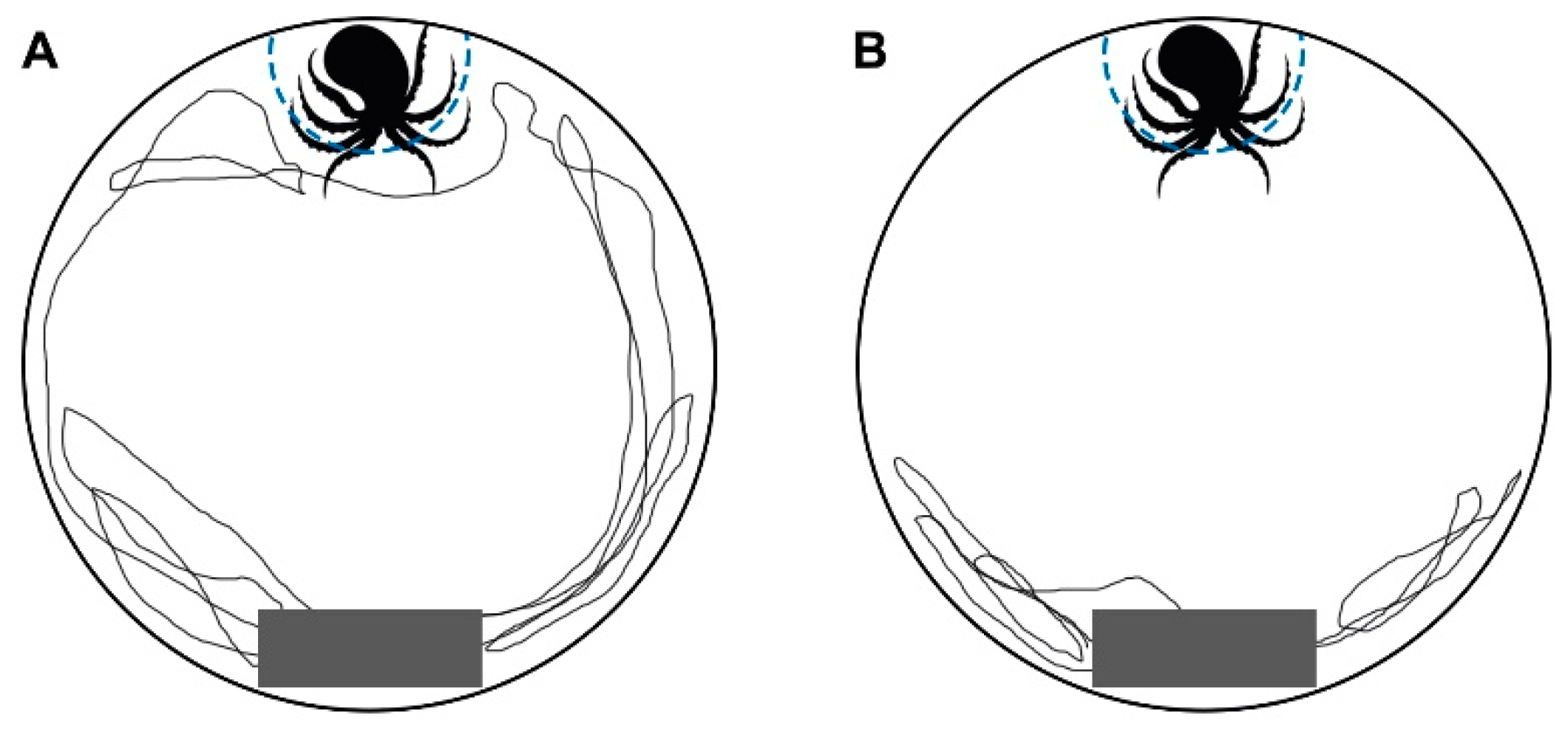
2.2.3. Predator Response Test
2.2.4. Habitat Selection Test
2.2.5. Territorial Behavior Scores Calculation
2.3. Data Analysis
3. Results
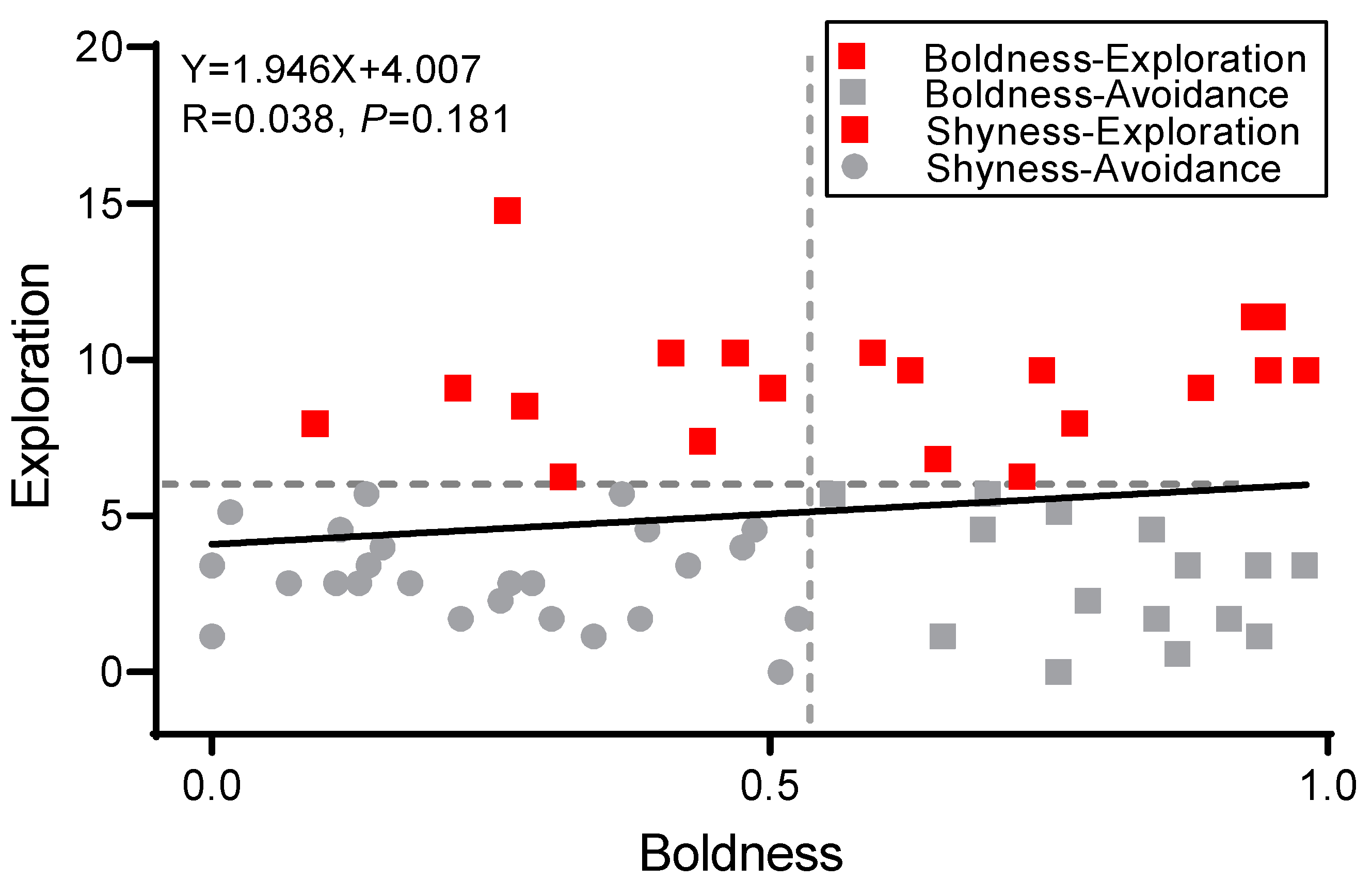
3.1. Clustering Analysis of Boldness and Exploration
3.2. Behavior Measurement in a Safe Environment
3.3. Predator Response Test
3.4. Habitat Selection Test
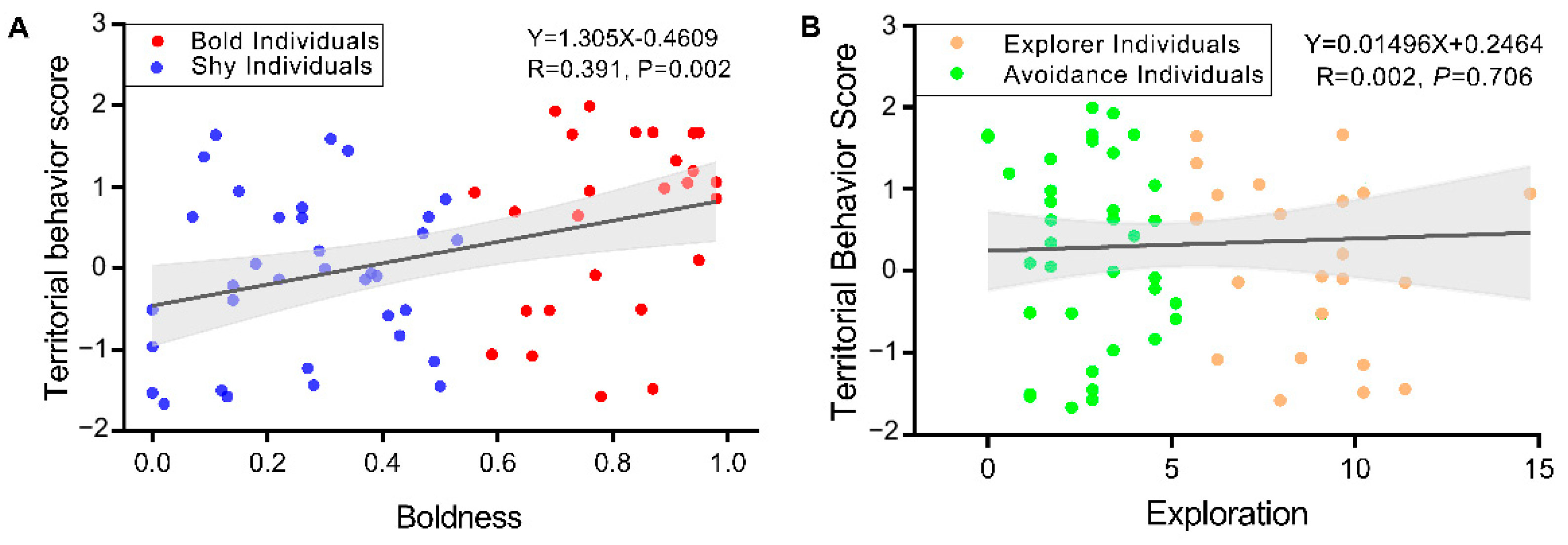
3.5. Correlation Analysis of Boldness, Exploration, and Territorial Behavior Scores
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabrera, D.; Nilsson, J.R.; Griffen, B.D. The development of animal personality across ontogeny: A cross-species review. Anim. Behav. 2021, 173, 137–144. [Google Scholar] [CrossRef]
- Reale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. Camb. Philos. Soc. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.R.; Goodman, A.C.; Santo, J.B.; Wong, R.Y. Repeatability and reliability of exploratory behavior in proactive and reactive zebrafish, Danio rerio. Sci. Rep. 2018, 8, 12114. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.F.; Gilliam, J.F.; Daley, M.J.; Le, A.N.; Skalski, G.T. Explaining Leptokurtic Movement Distributions: Intrapopulation Variation in Boldness and Exploration. Am. Nat. 2001, 158, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Mazue, G.P.F.; Dechaume-Moncharmont, F.X.; Godin, J.G.J. Boldness-exploration behavioral syndrome: Interfamily variability and repeatability of personality traits in the young of the convict cichlid (Amatitlania siquia). Behav. Ecol. 2015, 26, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Kudo, H.; Nishizawa, H.; Uchida, K.; Sato, K. Boldness–exploration behavioral syndrome in wild sub-adult green sea turtles caught at Oita, Japan. Appl. Anim. Behav. Sci. 2021, 236, 105216. [Google Scholar] [CrossRef]
- Cote, J.; Clobert, J.; Brodin, T.; Fogarty, S.; Sih, A. Personality-dependent dispersal: Characterization, ontogeny and consequences for spatially structured populations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 4065–4076. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.D.M.; Whattam, E.M.; Bennett, R.; Visanuvimol, L.; Lauzon, C.; Bertram, S.M. Behavioral correlations across activity, mating, exploration, aggression, and antipredator contexts in the European house cricket, Acheta domesticus. Behav. Ecol. Sociobiol. 2009, 64, 703–715. [Google Scholar] [CrossRef] [Green Version]
- Gasparini, C.; Speechley, E.M.; Polverino, G. The bold and the sperm: Positive association between boldness and sperm number in the guppy. R. Soc. Open. Sci. 2019, 6, 190474. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.A.C.; Moura, R.R.; Ogawa, C.Y.; Zanette, L.R.S.; Silva, J.R.F.; Rezende, C.F. Never forget where you came from: Microhabitat of origin influences boldness and exploration in the hermit crab Clibanarius symmetricus (Diogenidae). J. Exp. Mar. Biol. Ecol. 2020, 527, 151365. [Google Scholar] [CrossRef]
- Aliperti, J.R.; Davis, B.E.; Fangue, N.A.; Todgham, A.E.; Van Vuren, D.H. Bridging animal personality with space use and resource use in a free-ranging population of an asocial ground squirrel. Anim. Behav. 2021, 180, 291–306. [Google Scholar] [CrossRef]
- Saher, N.U.; Qureshi, N.A.; Siddiqui, A.S. Influence of sediment characteristics on density and distribution of Ocypodoid crab burrows (superfamily: Ocypodoidea) along the coastal areas of Pakistan. Acta Ecol. Sin. 2018, 38, 234–241. [Google Scholar] [CrossRef]
- Fuxjager, M.J.; Montgomery, J.L.; Becker, E.A.; Marler, C.A. Deciding to win: Interactive effects of residency, resources and ‘boldness’ on contest outcome in white-footed mice. Anim. Behav. 2010, 80, 921–927. [Google Scholar] [CrossRef]
- Rudin, F.S.; Briffa, M. Is boldness a resource-holding potential trait? Fighting prowess and changes in startle response in the sea anemone, Actinia equina. Proc. Biol. Sci. 2012, 279, 1904–1910. [Google Scholar] [CrossRef] [Green Version]
- Mills, L. Evidence for Individual Variation and the Role of Personality in the Assessment Strategies of Teleogryllus commodus Contests. Ph.D. Thesis, The University of Waikato, Hamilton, New Zealand, 2022. [Google Scholar]
- Naguib, M.; Titulaer, M.; Waas, J.R.; van Oers, K.; Sprau, P.; Snijders, L. Prior territorial responses and home range size predict territory defense in radio-tagged great tits. Behav. Ecol. Sociobiol. 2022, 76, 35. [Google Scholar] [CrossRef]
- Sommer-Trembo, C.; Petry, A.C.; Gomes Silva, G.; Vurusic, S.M.; Gismann, J.; Baier, J.; Krause, S.; Iorio, J.A.C.; Riesch, R.; Plath, M. Predation risk and abiotic habitat parameters affect personality traits in extremophile populations of a neotropical fish (Poecilia vivipara). Ecol. Evol. 2017, 7, 6570–6581. [Google Scholar] [CrossRef] [Green Version]
- Yang, H. Construction of marine ranching in China: Reviews and prospects. J. Fish. China 2016, 40, 1133–1140. [Google Scholar] [CrossRef]
- Liu, H.; Lv, H.; Cui, F.; Xie, W.; Li, B.; Wang, L.; Liu, Q.; Chen, L.; Xing, K.; Wang, X.; et al. Parental Contribution and Genetic Diversity between Broodstock and Offsprings in Swimming Crab (Portunus trituberculatus) Releasing into Natural Waters. Fish. Sci. 2016, 35, 613–619. [Google Scholar]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China. China Fisheries Yearbook; Chinese Agriculture Express: Beijing, China, 2021. [Google Scholar]
- Ord, T.J. Costs of territoriality: A review of hypotheses, meta-analysis, and field study. Oecologia 2021, 197, 615–631. [Google Scholar] [CrossRef]
- Su, X.; Sun, Y.; Liu, D.; Wang, F.; Liu, J.; Zhu, B. Agonistic behaviour and energy metabolism of bold and shy swimming crabs Portunus trituberculatus. J. Exp. Biol. 2019, 222, jeb188706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Zhang, H.; Lu, Y.; Wang, F.; Liu, D. The Effect of Intruder Density on Territoriality and Dominance in Male Swimming Crab (Portunus trituberculatus). Animals 2022, 12, 314. [Google Scholar] [CrossRef] [PubMed]
- Mazzamuto, M.V.; Cremonesi, G.; Santicchia, F.; Preatoni, D.; Martinoli, A.; Wauters, L.A. Rodents in the arena: A critical evaluation of methods measuring personality traits. Ethol. Ecol. Evol. 2018, 31, 38–58. [Google Scholar] [CrossRef]
- Su, X.; Zhu, B.; Ren, Z.; Wang, F. Differences in Agonistic Behavior and Energy Metabolism between Male and Female Swimming Crab Portunus trituberculatus Based on the Analysis of Boldness. Animals 2022, 12, 2363. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Su, X.; Liu, D.; Guo, Z.; Wang, F.; Lu, Y. A New Method of Aquatic Animal Personality Analysis Based on Machine Learning (PAML): Taking Swimming Crab Portunus trituberculatus as an Example. Front. Mar. Sci. 2020, 7, 32. [Google Scholar] [CrossRef]
- Zhu, B.; Su, X.; Yu, W.; Wang, F. What Forms, Maintains, and Changes the Boldness of Swimming Crabs (Portunus trituberculatus)? Animals 2022, 12, 1618. [Google Scholar] [CrossRef]
- Church, K.D.W.; Grant, J.W.A.; Wright, J. Effects of habitat complexity, dominance and personality on habitat selection: Ideal despotic cichlids. Ethology 2019, 125, 832–845. [Google Scholar] [CrossRef]
- Landys, M.M.; Goymann, W.; Schwabl, I.; Trapschuh, M.; Slagsvold, T. Impact of season and social challenge on testosterone and corticosterone levels in a year-round territorial bird. Horm. Behav. 2010, 58, 317–325. [Google Scholar] [CrossRef]
- Zhu, B.; Lu, Y.; Wang, F.; Liu, D.; Su, X.; Shang, Y. Will swimming crab change their territoriality when prey density increases? J. Exp. Mar. Biol. Ecol. 2021, 539, 151546. [Google Scholar] [CrossRef]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef]
- Reaney, L.T.; Backwell, P.R.Y. Risk-taking behavior predicts aggression and mating success in a fiddler crab. Behav. Ecol. 2007, 18, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Courtene-Jones, W.; Briffa, M. Boldness and asymmetric contests: Role- and outcome-dependent effects of fighting in hermit crabs. Behav. Ecol. 2014, 25, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Suzuki, N.; Koga, T. Burrow defense behaviors in a sand-bubbler crab, Scopimera globosa, in relation to body size and prior residence. J. Ethol. 2001, 19, 93–96. [Google Scholar] [CrossRef]
- Daly, B.J.; Eckert, G.L.; Long, W.C.; Pernet, F. Moulding the ideal crab: Implications of phenotypic plasticity for crustacean stock enhancement. ICES J. Mar. Sci. 2021, 78, 421–434. [Google Scholar] [CrossRef]
- Jones, K.A.; Godin, J.G. Are fast explorers slow reactors? Linking personality type and anti-predator behaviour. Proc. Biol. Sci. 2010, 277, 625–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, X.; Wang, W.; Xu, R.; Li, C.; Zhang, F. Effects of Personality Traits on the Food-Scratching Behaviour and Food Intake of Japanese Quail (Coturnix japonica). Animals 2021, 11, 3423. [Google Scholar] [CrossRef]
- Bamber, J.A.; Shuttleworth, C.M.; Hayward, M.W. Do Differing Levels of Boldness Influence the Success of Translocation? A Pilot Study on Red Squirrels (Sciurus vulgaris). Animals 2020, 10, 1748. [Google Scholar] [CrossRef]
- Oldham, R.C.; Pintor, L.M.; Gray, S.M. Behavioral differences within and among populations of an African cichlid found in divergent and extreme environments. Curr. Zool. 2019, 65, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Church, K.D.W.; Matte, J.-M.; Grant, J.W.A.; Pinter-Wollman, N. Territoriality modifies the effects of habitat complexity on animal behavior: A meta-analysis. Behav. Ecol. 2022, 33, 455–466. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Component | Boldness | F | p | df1, df2 | AIC | Intercept | Residual | |
|---|---|---|---|---|---|---|---|---|
| Boldness Individuals | Shyness Individuals | |||||||
| Territory size (m2) | 0.46 ± 0.14 | 0.46 ± 0.13 | 0.010 | 0.921 | 1, 59 | 6.187 | 0.467 | 0.014 |
| Occupying time (s) | 1939 ± 646 | 1929 ± 468 | 0.001 | 0.971 | 1, 59 | 404.792 | 1230.055 | 2231.553 |
| Defense time (s) | 2158 ± 176 | 1564 ± 838 | 7.735 | 0.008 | 1, 59 | 314.512 | 1364.813 | 872.252 |
| Behavioral Component | Exploration | F | p | df1, df2 | AIC | Intercept | Residual | |
| Explorer Individuals | Avoidance Individuals | |||||||
| Territory size (m2) | 0.53 ± 0.04 | 0.43 ± 0.13 | 3.134 | 0.082 | 1, 59 | 9.335 | 0.433 | 0.013 |
| Occupying time (s) | 2064 ± 222 | 1869 ± 154 | 0.508 | 0.478 | 1, 59 | 404.182 | 1170.358 | 2212.5 |
| Defense time (s) | 1801 ± 169 | 1692 ± 118 | 0.277 | 0.601 | 1, 59 | 379.662 | 993.05 | 1446.512 |
| Behavioral Component | Boldness | F | p | df1, df2 | AIC | Intercept | Residual | |
|---|---|---|---|---|---|---|---|---|
| Boldness Individuals | Shyness Individuals | |||||||
| Territory size (m2) | 0.55 ± 0.02 | 0.43 ± 0.12 | 8.625 | 0.035 | 1, 59 | −45.523 | 0.439 | 0.007 |
| Occupying time (s) | 1672 ± 639 | 2362 ± 313 | 2.427 | 0.012 | 1, 59 | 400.621 | 1263.820 | 2079.338 |
| Defense time (s) | 955 ± 121 | 781 ± 105 | 1.170 | 0.284 | 1, 59 | 348.599 | 689.619 | 861.242 |
| Times attacking the predator | 5.87 ± 1.01 | 2.25 ± 1.42 | 2.251 | 0.016 | 1, 10 | 62.84 | 2.25 | 2.721 |
| Behavioral Component | Exploration | F | p | df1, df2 | AIC | Intercept | Residual | |
| Explorer Individuals | Avoidance Individuals | |||||||
| Territory size (m2) | 0.51 ± 0.03 | 0.47 ± 0.02 | 0.873 | 0.354 | 1, 59 | −38.443 | 0.473 | 0.008 |
| Occupying time (h) | 2199 ± 220 | 2196 ± 153 | 0.001 | 0.992 | 1, 59 | 402.895 | 1196.341 | 2164.897 |
| Defense time (h) | 913 ± 139 | 826 ± 98 | 0.259 | 0.613 | 1, 59 | 349.395 | 627.550 | 874.381 |
| Times attacking the predator | 5.25 ± 1.50 | 3.37 ± 1.06 | 1.034 | 0.033 | 1, 10 | 63.886 | 3.375 | 3.021 |
| Behavioral Component | Boldness | F | p | df1, df2 | AIC | Intercept | Residual | |
|---|---|---|---|---|---|---|---|---|
| Boldness Individuals | Shyness Individuals | |||||||
| Time occupying complex habitat (s) | 2108.70 ± 210.05 | 2181.00 ± 146.70 | 0.005 | 0.945 | 1, 59 | 988.317 | 2181 | 294.146 |
| Feeding time (s) | 301.73 ± 32.29 | 265.28 ± 27.83 | 0.731 | 0.396 | 1, 59 | 782.959 | 265.286 | 9040.114 |
| Behavioral component | Exploration | F | p | df1, df2 | AIC | Intercept | Residual | |
| Explorer Individuals | Avoidance Individuals | |||||||
| Time occupying complex habitat (s) | 1664.88 ± 183.04 | 2277.37 ± 157.76 | 0.773 | 0.018 | 1, 59 | 987.657 | 2277.371 | 290.364 |
| Feeding time (s) | 293.35 ± 36.99 | 274.70 ± 25.84 | 0.171 | 0.681 | 1, 59 | 783.41 | 274.707 | 9125.667 |
| Component Loadings | PC1 |
| Behavior measurement in a safe environment | |
| Territory size | 0.112 |
| Occupying time | −0.080 |
| Defense time | 0.008 |
| Predator response test | |
| Territory size | −0.884 |
| Occupying time | 0.967 |
| Defense time | −0.834 |
| Habitat selection test | |
| Feeding time | 0.102 |
| Time occupying complex habitat | 0.930 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, B.; Wang, X.; Ren, Z.; Zhang, H.; Liu, D.; Wang, F. Each Personality Performs Its Own Function: Boldness and Exploration Lead to Differences in the Territoriality of Swimming Crabs (Portunus trituberculatus). Biology 2023, 12, 883. https://doi.org/10.3390/biology12060883
Zhu B, Wang X, Ren Z, Zhang H, Liu D, Wang F. Each Personality Performs Its Own Function: Boldness and Exploration Lead to Differences in the Territoriality of Swimming Crabs (Portunus trituberculatus). Biology. 2023; 12(6):883. https://doi.org/10.3390/biology12060883
Chicago/Turabian StyleZhu, Boshan, Xin Wang, Ziwen Ren, Hanzun Zhang, Dapeng Liu, and Fang Wang. 2023. "Each Personality Performs Its Own Function: Boldness and Exploration Lead to Differences in the Territoriality of Swimming Crabs (Portunus trituberculatus)" Biology 12, no. 6: 883. https://doi.org/10.3390/biology12060883
APA StyleZhu, B., Wang, X., Ren, Z., Zhang, H., Liu, D., & Wang, F. (2023). Each Personality Performs Its Own Function: Boldness and Exploration Lead to Differences in the Territoriality of Swimming Crabs (Portunus trituberculatus). Biology, 12(6), 883. https://doi.org/10.3390/biology12060883