
Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guájares (Granada, Spain) Wildfire

 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
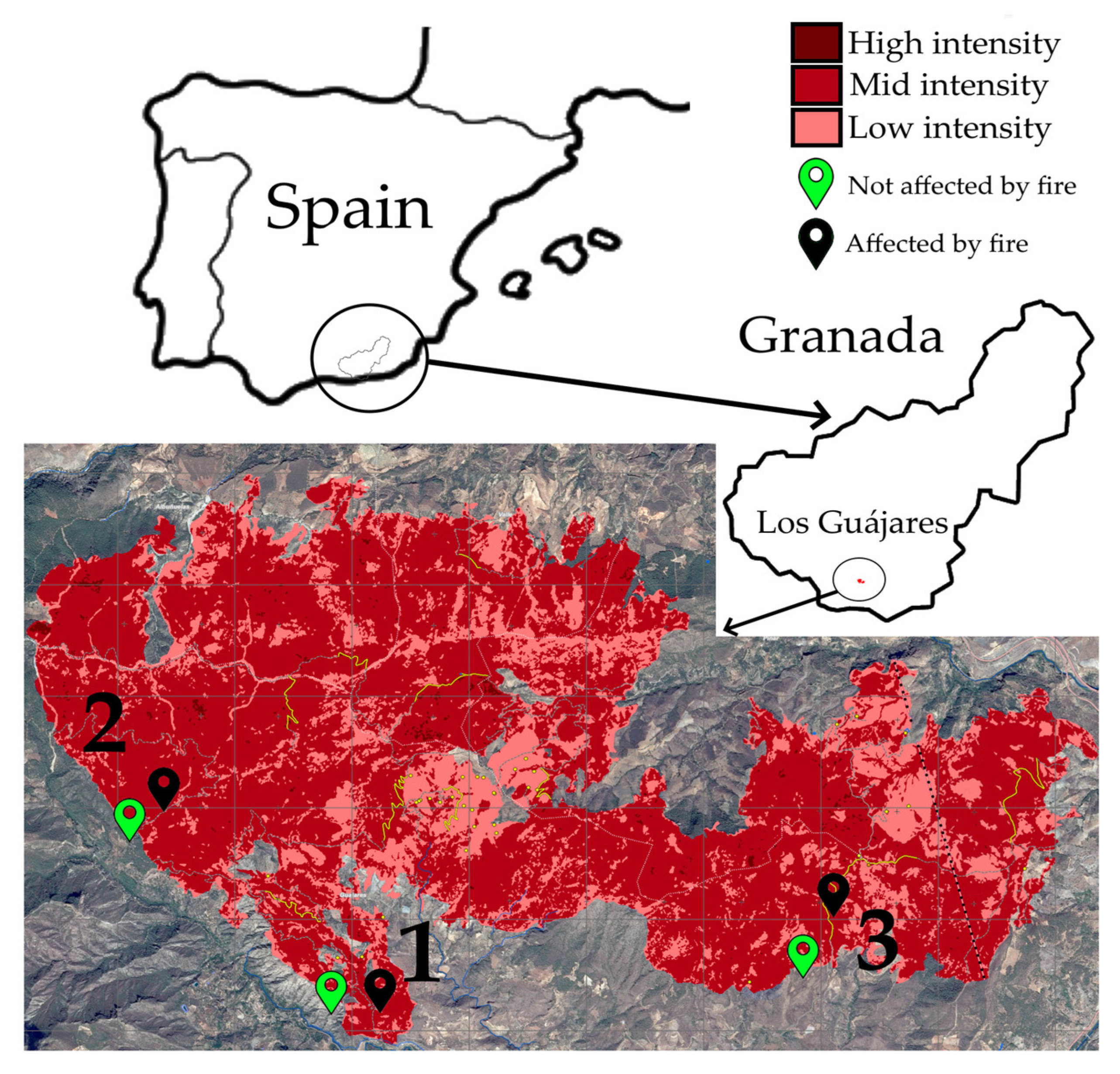
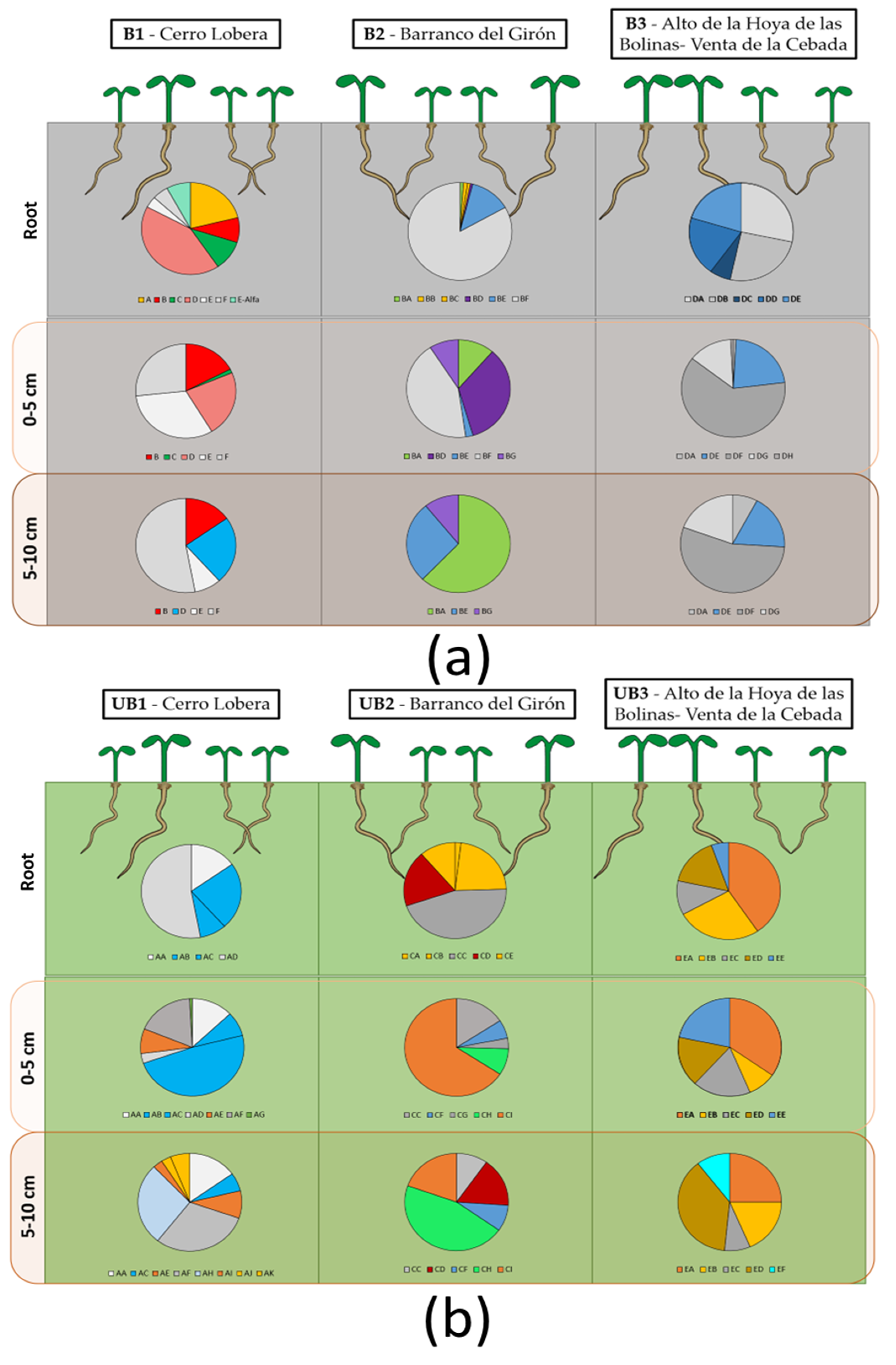
2.1. Sampling
2.2. Soil Characterization
2.3. Isolation, Identification and Analysis of the Culturable Populations in Ashes, Soil and Roots
2.4. Nutrient Cycling Skills
2.4.1. Screening of Nitrogen Fixing Activity
2.4.2. Phosphate Solubilization
2.4.3. Potassium Solubilization
2.4.4. Sulfur-Oxidation
2.4.5. Siderophores Production
2.5. Biofilm Production
2.6. Plant Growth Promoting (PGP) and Stress-Tolerance Enhancing Traits
2.7. Microcosm Test
2.8. Microscopy
2.9. Statistics and Analyses
3. Results
3.1. Soil Characterization
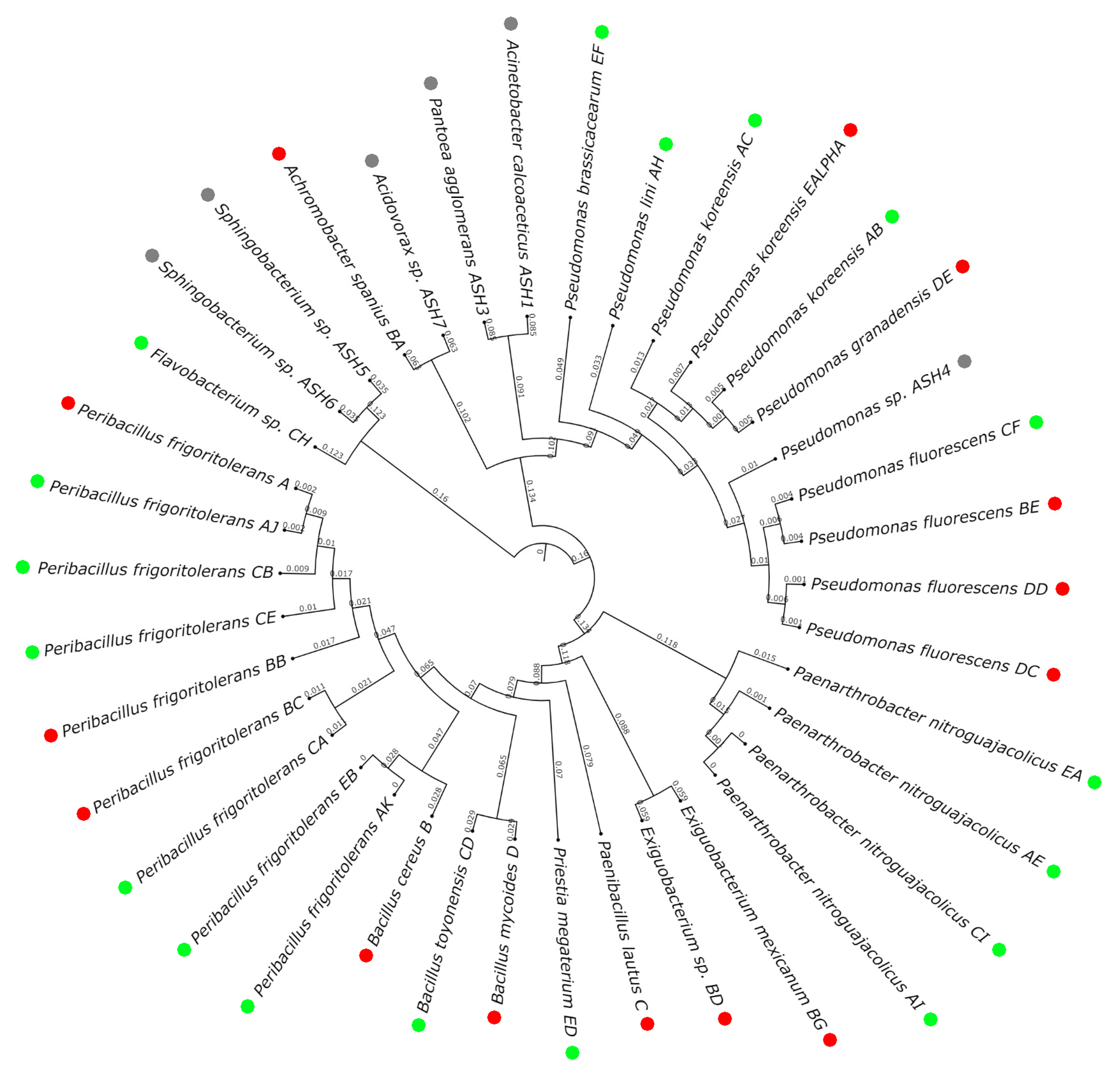
3.2. Population Analysis: Differences Caused by Wildfire Events
3.3. Screening of Nutrient-Related, Structuring and Plant Growth-Promoting Skills
3.4. Selection of Candidates and Deep Characterization
3.5. Microcosms Evaluation
3.5.1. Soil Evaluation: EC, pH and Slaking Index Coefficient (SIC)
3.5.2. Microscopy and Granulometry Evaluation of Soil Aggregation Degree
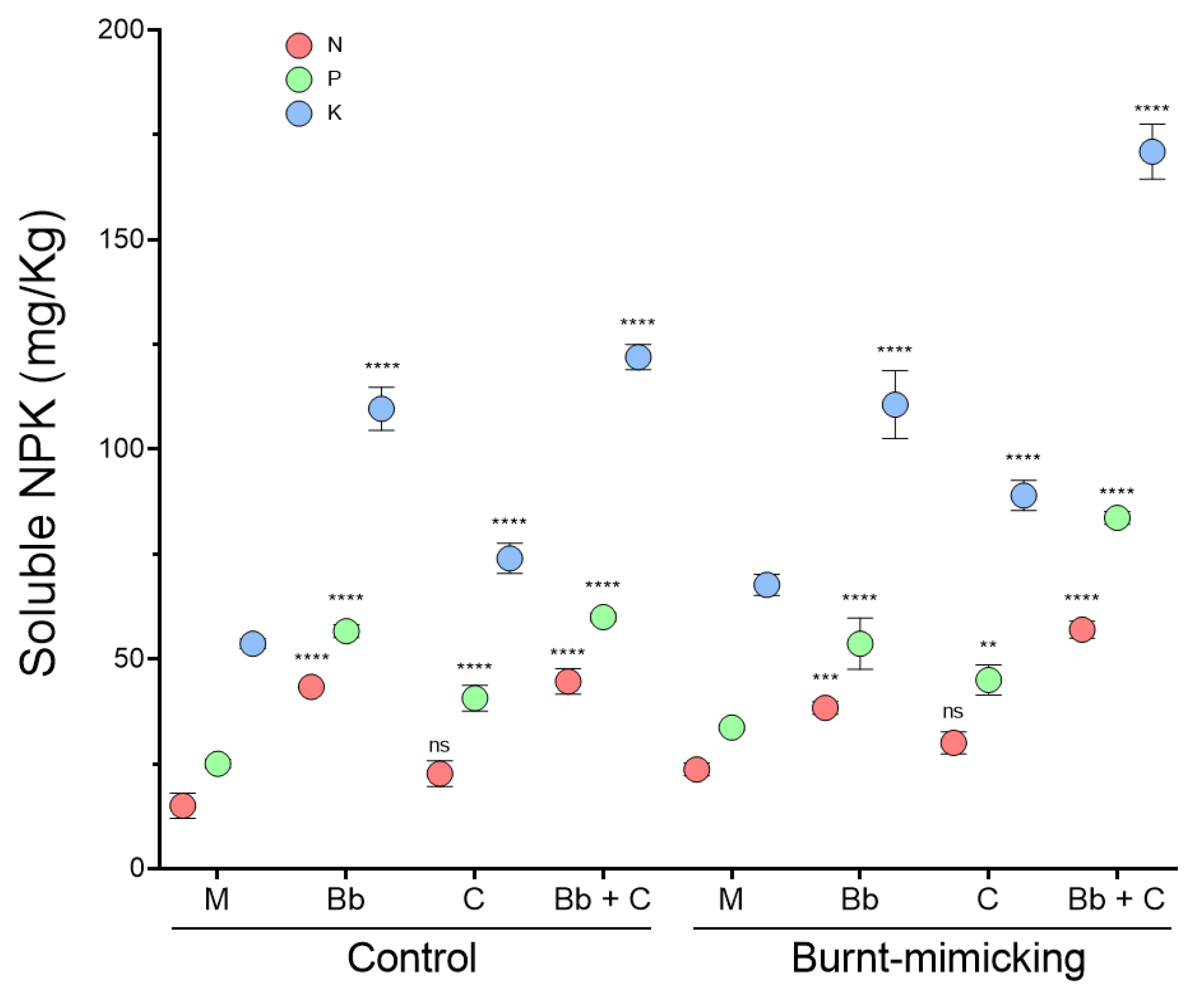
3.5.3. Main Nutrient Determination (N, P, and K)
3.5.4. Germination Rate
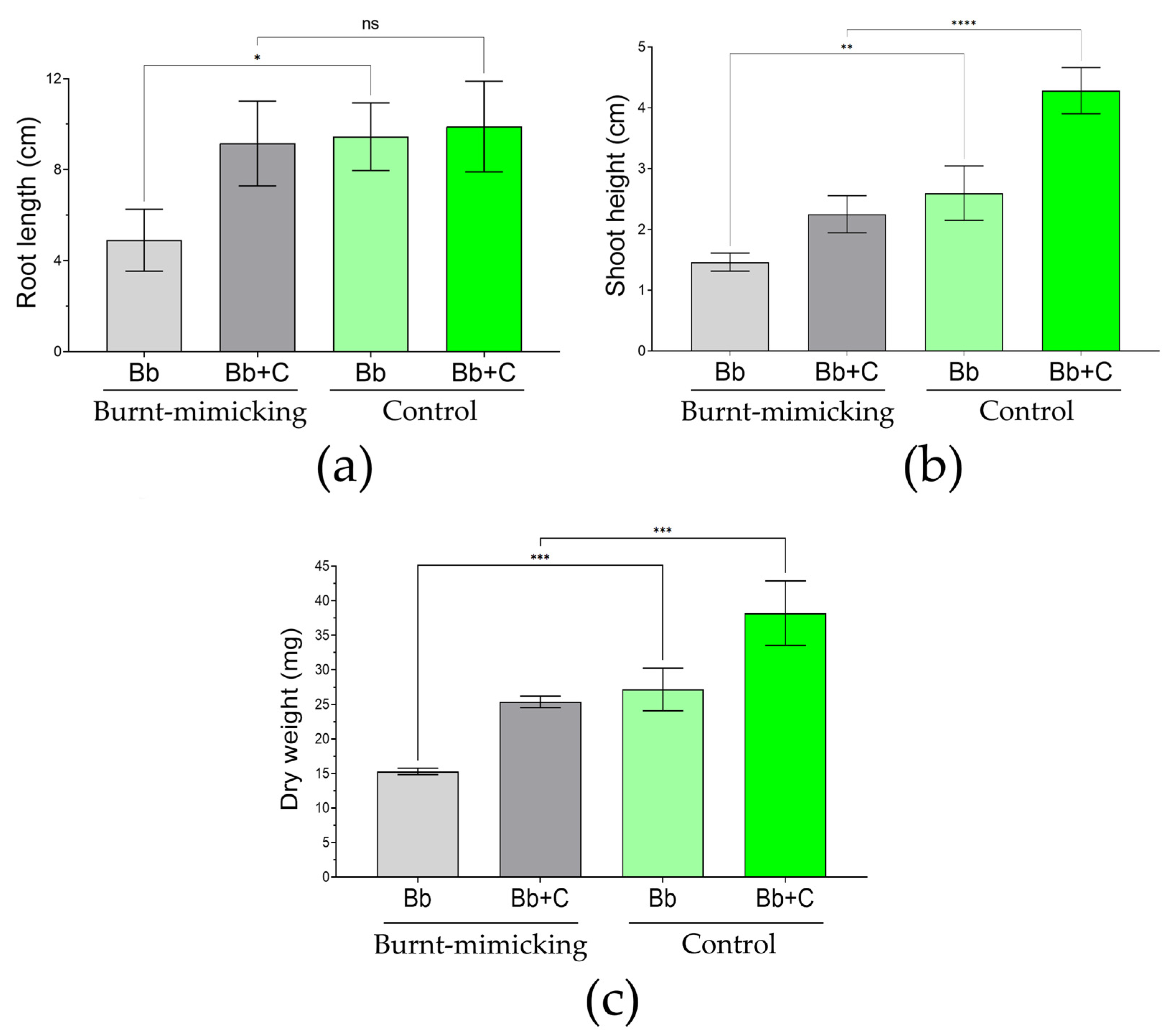
3.5.5. Plant Phenotyping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NOAA. State of the Climate: Global Climate Report for 2022; USA. Available online: https://www.ncei.noaa.gov/access/monitoring/monthly-report/global/202213 (accessed on 18 January 2023).
- Van den Besselaar, E.J.M.; Klein Tank, A.M.G.; Buishand, T.A. Trends in European precipitation extremes over 1951–2010. Int. J. Climatol. 2013, 33, 2682–2689. [Google Scholar] [CrossRef]
- EFFIS. Annual Fire Report. Available online: https://effis.jrc.ec.europa.eu/ (accessed on 16 January 2023).
- MITECO. Incendios Forestales. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/incendios-forestales/estadisticas-datos.aspx (accessed on 16 January 2023).
- Memoli, V.; Panico, S.C.; Santorufo, L.; Barile, R.; Di Natale, G.; Di Nunzio, A.; Toscanesi, M.; Trifuoggi, M.; De Marco, A.; Maisto, G. Do Wildfires Cause Changes in Soil Quality in the Short Term? Int. J. Environ. Res. Public Health 2020, 17, 5343. [Google Scholar] [CrossRef] [PubMed]
- Bodí, M.B.; Mataix-Solera, J.; Doerr, S.H.; Cerdà, A. The wettability of ash from burned vegetation and its relationship to Mediterranean plant species type, burn severity and total organic carbon content. Geoderma 2011, 160, 599–607. [Google Scholar] [CrossRef]
- Agbeshie, A.A.; Abugre, S.; Atta-Darkwa, T.; Awuah, R. A review of the effects of forest fire on soil properties. J. For. Res. 2022, 33, 1419–1441. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakesby, R.A. Post-wildfire soil erosion in the Mediterranean: Review and future research directions. Earth-Sci. Rev. 2011, 105, 71–100. [Google Scholar] [CrossRef]
- García-Carmona, M.; García-Orenes, F.; Arcenegui, V.; Mataix-Solera, J. The Recovery of Mediterranean Soils After Post-Fire Management: The Role of Biocrusts and Soil Microbial Communities. Span. J. Soil Sci. 2023, 13, 11388. [Google Scholar] [CrossRef]
- Navidi, M.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; Carra, B.G.; Parhizkar, M.; Zema, D.A. Mid-Term Changes in Soil Properties after Wildfire, Straw Mulching and Salvage Logging in Pinus halepensis Mill. Forests. Fire 2022, 5, 158. [Google Scholar] [CrossRef]
- Arocena, J.M.; Opio, C. Prescribed fire-induced changes in properties of sub-boreal forest soils. Geoderma 2003, 113, 1–16. [Google Scholar] [CrossRef]
- Neff, J.C.; Harden, J.W.; Gleixner, G. Fire effects on soil organic matter content, composition, and nutrients in boreal interior Alaska. Can. J. For. Res. 2005, 35, 2178–2187. [Google Scholar] [CrossRef]
- Kumar, M.; Sheikh, M.A.; Bhat, J.A.; Bussmann, R.W. Effect of fire on soil nutrients and under storey vegetation in Chir pine forest in Garhwal Himalaya, India. Acta Ecol. Sin. 2013, 33, 59–63. [Google Scholar] [CrossRef]
- Ferreira, A.J.D.; Coelho, C.O.A.; Ritsema, C.J.; Boulet, A.K.; Keizer, J.J. Soil and water degradation processes in burned areas: Lessons learned from a nested approach. Catena 2008, 74, 273–285. [Google Scholar] [CrossRef]
- Girona-García, A.; Vieira, D.C.S.; Silva, J.; Fernández, C.; Robichaud, P.R.; Keizer, J.J. Effectiveness of post-fire soil erosion mitigation treatments: A systematic review and meta-analysis. Earth-Sci. Rev. 2021, 217, 103611. [Google Scholar] [CrossRef]
- Li, F.; Liang, X.; Li, H.; Jin, Y.; Jin, J.; He, M.; Klumpp, E.; Bol, R. Enhanced soil aggregate stability limits colloidal phosphorus loss potentials in agricultural systems. Environ. Sci. Eur. 2020, 32, 17. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, L.; Nunes, A.N.; Bento-Gonçalves, A.; Vieira, A. Soil Erosion after Wildfires in Portugal: What Happens When Heavy Rainfall Events Occur? In Research on Soil Erosion; Danilo, G., Silvia, S., Eds.; IntechOpen: Rijeka, Croatia, 2012; Chapter 4. [Google Scholar]
- Chungu, D.; Ng’andwe, P.; Mubanga, H.; Chileshe, F. Fire alters the availability of soil nutrients and accelerates growth of Eucalyptus grandis in Zambia. J. For. Res. 2020, 31, 1637–1645. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Elberling, B.; Ambus, P.L. Fire increases soil nitrogen retention and alters nitrogen uptake patterns among dominant shrub species in an Arctic dry heath tundra. Sci. Total Environ. 2022, 807, 150990. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ruiz, C.; Sandoval, M.; Stolpe, N.; Sanchez-Hernandez, J.C. Fire impacts on soil and post fire emergency stabilization treatments in Mediterranean-climate regions. Chil. J. Agric. Res. 2022, 82, 335–347. [Google Scholar] [CrossRef]
- Bodí, M.B.; Martin, D.A.; Balfour, V.N.; Santín, C.; Doerr, S.H.; Pereira, P.; Cerdà, A.; Mataix-Solera, J. Wildland fire ash: Production, composition and eco-hydro-geomorphic effects. Earth-Sci. Rev. 2014, 130, 103–127. [Google Scholar] [CrossRef]
- Sanborn, P.T.; Ballard, T.M. Combustion Losses of Sulphur from Conifer Foliage: Implications of Chemical Form and Soil Nitrogen Status. Biogeochemistry 1991, 12, 129–134. [Google Scholar] [CrossRef]
- Gómez-Sánchez, E.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; González-Romero, J.; Sagra, J.; Moya, D.; De Las Heras, J. Effects of post-fire hillslope stabilisation techniques on chemical, physico-chemical and microbiological soil properties in mediterranean forest ecosystems. J. Environ. Manag. 2019, 246, 229–238. [Google Scholar] [CrossRef]
- Datta, R. To extinguish or not to extinguish: The role of forest fire in nature and soil resilience. J. King Saud Univ. Sci. 2021, 33, 101539. [Google Scholar] [CrossRef]
- Oyonarte, C.; Aranda, V.; Durante, P. Soil surface properties in Mediterranean mountain ecosystems: Effects of environmental factors and implications of management. For. Ecol. Manag. 2008, 254, 156–165. [Google Scholar] [CrossRef]
- Barreiro, A.; Díaz-Raviña, M. Fire impacts on soil microorganisms: Mass, activity, and diversity. Curr. Opin. Environ. Sci. Health 2021, 22, 100264. [Google Scholar] [CrossRef]
- Nelson, A.R.; Narrowe, A.B.; Rhoades, C.C.; Fegel, T.S.; Daly, R.A.; Roth, H.K.; Chu, R.K.; Amundson, K.K.; Young, R.B.; Steindorff, A.S.; et al. Wildfire-dependent changes in soil microbiome diversity and function. Nat. Microbiol. 2022, 7, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Moya, D.; Fonturbel, T.; Peña, E.; Alfaro-Sanchez, R.; Plaza-Álvarez, P.A.; González-Romero, J.; Lucas-Borja, M.E.; de Las Heras, J. Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron). Forests 2022, 13, 1118. [Google Scholar] [CrossRef]
- Singh, D.; Sharma, P.; Kumar, U.; Daverey, A.; Arunachalam, K. Effect of forest fire on soil microbial biomass and enzymatic activity in oak and pine forests of Uttarakhand Himalaya, India. Ecol. Process. 2021, 10, 29. [Google Scholar] [CrossRef]
- Maquia, I.S.A.; Fareleira, P.; Videira e. Castro, I.; Soares, R.; Brito, D.R.A.; Mbanze, A.A.; Chaúque, A.; Máguas, C.; Ezeokoli, O.T.; Ribeiro, N.S.; et al. The Nexus between Fire and Soil Bacterial Diversity in the African Miombo Woodlands of Niassa Special Reserve, Mozambique. Microorganisms 2021, 9, 1562. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.-A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Bouskill, N.J.; Mekonnen, Z.; Zhu, Q.; Grant, R.; Riley, W.J. Microbial contribution to post-fire tundra ecosystem recovery over the 21st century. Commun. Earth Environ. 2022, 3, 26. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef] [Green Version]
- Santini, G.; Ruggiero, A.G.; Ceccherini, M.T.; Santorufo, L.; Memoli, V.; Pietramellara, G.; De Marco, A.; Giarra, A.; Di Natale, G.; Trifuoggi, M.; et al. N cycle in burnt and unburnt soils under different vegetation covers in the Mediterranean region. Catena 2022, 214, 106234. [Google Scholar] [CrossRef]
- Bright, B.C.; Hudak, A.T.; Kennedy, R.E.; Braaten, J.D.; Henareh Khalyani, A. Examining post-fire vegetation recovery with Landsat time series analysis in three western North American forest types. Fire Ecol. 2019, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Tangney, R.; Paroissien, R.; Le Breton, T.D.; Thomsen, A.; Doyle, C.A.T.; Ondik, M.; Miller, R.G.; Miller, B.P.; Ooi, M.K.J. Success of post-fire plant recovery strategies varies with shifting fire seasonality. Commun. Earth Environ. 2022, 3, 126. [Google Scholar] [CrossRef]
- Prior, L.D.; Bowman, D.M.J.S. Classification of Post-Fire Responses of Woody Plants to include Pyrophobic Communities. Fire 2020, 3, 15. [Google Scholar] [CrossRef]
- Abella, S.R. Disturbance and plant succession in the Mojave and Sonoran deserts of the American Southwest. Int. J. Environ. Res. Public Health 2010, 7, 1248–1284. [Google Scholar] [CrossRef] [Green Version]
- Buhk, C.; Meyn, A.; Jentsch, A. The Challenge of Plant Regeneration after Fire in the Mediterranean Basin: Scientific Gaps in Our Knowledge on Plant Strategies and Evolution of Traits. Plant Ecol. 2007, 192, 1–19. [Google Scholar] [CrossRef]
- Auld, T.D.; Denham, A.J. How Much Seed Remains in the Soil after a Fire? Plant Ecol. 2006, 187, 15–24. [Google Scholar] [CrossRef]
- Fournier, T.; Fèvre, J.; Carcaillet, F.; Carcaillet, C. For a few years more: Reductions in plant diversity 70 years after the last fire in Mediterranean forests. Plant Ecol. 2020, 221, 559–576. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef]
- Dove, N.C.; Klingeman, D.M.; Carrell, A.A.; Cregger, M.A.; Schadt, C.W. Fire alters plant microbiome assembly patterns: Integrating the plant and soil microbial response to disturbance. New Phytol. 2021, 230, 2433–2446. [Google Scholar] [CrossRef] [PubMed]
- Naydenova, G.; Radkova, M.; Iantcheva, A. Legumes in natural post-fire successions of forest meadows and pastures in Northern Bulgaria. Thaiszia J. Bot. 2022, 32, 67–79. [Google Scholar] [CrossRef]
- Bekdouche, F.; Sahnoune, M.; Krouchi, F.; Achour, S.; Guemati, N.; Arezki, D. The contribution of legumes to post-fire regeneration of Quercus suber and Pinus halepensis forests in Northeastern Algeria. Rev. D’ecologie (Terre Vie) 2011, 66, 29–42. [Google Scholar] [CrossRef]
- Niza-Costa, M.; Rodríguez-dos Santos, A.S.; Rebelo-Romão, I.; Ferrer, M.V.; Sequero López, C.; Vílchez, J.I. Geographically Disperse, Culturable Seed-Associated Microbiota in Forage Plants of Alfalfa (Medicago sativa L.) and Pitch Clover (Bituminaria bituminosa L.): Characterization of Beneficial Inherited Strains as Plant Stress-Tolerance Enhancers. Biology 2022, 11, 1838. [Google Scholar] [CrossRef]
- Salis, M.; Del Giudice, L.; Jahdi, R.; Alcasena-Urdiroz, F.; Scarpa, C.; Pellizzaro, G.; Bacciu, V.; Schirru, M.; Ventura, A.; Casula, M.; et al. Spatial Patterns and Intensity of Land Abandonment Drive Wildfire Hazard and Likelihood in Mediterranean Agropastoral Areas. Land 2022, 11, 1942. [Google Scholar] [CrossRef]
- Mantero, G.; Morresi, D.; Marzano, R.; Motta, R.; Mladenoff, D.J.; Garbarino, M. The influence of land abandonment on forest disturbance regimes: A global review. Landsc. Ecol. 2020, 35, 2723–2744. [Google Scholar] [CrossRef]
- Fernández-Raga, M.; Martins, M.; Porras, E.; Fraile, R.; Keizer, J. Splash Erosion on Terraces, Does It Make a Difference If the Terracing Is Done before or after a Fire? Hydrology 2021, 8, 180. [Google Scholar] [CrossRef]
- Martins, M.; Serpa, D.; Machado, A.I.; Lenne, R.F.H.; Faria, S.; Ferreira, R.S.V.; Skulska, I.; Prats; Varela, M.E.; Keizer, J.J. Effects of terrace construction on runoff and erosion in a recently burnt forest area in north-central Portugal. Flamma 2013, 4, 81–84. [Google Scholar]
- Materechera, S.A.; Dexter, A.R.; Alston, A.M. Formation of aggregates by plant roots in homogenised soils. Plant Soil 1992, 142, 69–79. [Google Scholar] [CrossRef]
- Batista, A.M.; Libardi, P.L.; Giarola, N.F.B. Evaluation of the soil aggregation induced by the plant roots in an Oxisol by turbidimetry and water percolation. Rhizosphere 2020, 16, 100265. [Google Scholar] [CrossRef]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root Exudates Induce Soil Macroaggregation Facilitated by Fungi in Subsoil. Front. Environ. Sci. 2018, 6, 140. [Google Scholar] [CrossRef]
- Naveed, M.; Brown, L.K.; Raffan, A.C.; George, T.S.; Bengough, A.G.; Roose, T.; Sinclair, I.; Koebernick, N.; Cooper, L.; Hallett, P.D. Rhizosphere-Scale Quantification of Hydraulic and Mechanical Properties of Soil Impacted by Root and Seed Exudates. Vadose Zone J. 2018, 17, 170083. [Google Scholar] [CrossRef] [Green Version]
- Cania, B.; Vestergaard, G.; Krauss, M.; Fliessbach, A.; Schloter, M.; Schulz, S. A long-term field experiment demonstrates the influence of tillage on the bacterial potential to produce soil structure-stabilizing agents such as exopolysaccharides and lipopolysaccharides. Environ. Microbiome 2019, 14, 1. [Google Scholar] [CrossRef]
- Lehmann, A.; Zheng, W.; Ryo, M.; Soutschek, K.; Roy, J.; Rongstock, R.; Maaß, S.; Rillig, M.C. Fungal Traits Important for Soil Aggregation. Front. Microbiol. 2020, 10, 2904. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Song, D.; Liu, D.; Sun, J.; Wang, X.; Zhou, W.; Liang, G. Soil Aggregation Shaped the Distribution and Interaction of Bacterial-Fungal Community Based on a 38-Year Fertilization Experiment in China. Front. Microbiol. 2022, 13, 824681. [Google Scholar] [CrossRef]
- Cai, P.; Sun, X.; Wu, Y.; Gao, C.-H.; Mortimer, M.; Holden, P.; Redmile-Gordon, M.; Huang, Q. Soil biofilms: Microbial interactions, challenges, and advanced techniques for ex-situ characterization. Soil Ecol. Lett. 2019, 1, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Yang, Y.; Yi, D.; Zhang, H. Measurements of Root Colonized Bacteria Species. Bio-Protocol 2021, 11, e3976. [Google Scholar] [CrossRef]
- Breakwell, D.; Woolverton, C.; MacDonald, B.; Smith, K.; Robison, R. Colony Morphology Protocol. In Proceedings of the 14th American Society for Microbiology Conference for Undergraduate Educators (ASMCUE), Buffalo, NY, USA, 29 September 2007. [Google Scholar]
- Dashti, A.; Jadaon, M.; Abdulsamad, A.; Dashti, H. Heat Treatment of Bacteria: A Simple Method of DNA Extraction for Molecular Techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Sulistiyani, T.; Meliah, S. Isolation and Characterization of Nitrogen Fixing Endophytic Bacteria Associated with Sweet Sorghum (Sorghum bicolor). In Proceedings of the 1st SATREPS Conference, Bogor, Indonesia, 14 November 2016. [Google Scholar]
- Iwata, K.; Azlan, A.; Yamakawa, H.; Omori, T. Ammonia accumulation in culture broth by the novel nitrogen-fixing bacterium, Lysobacter sp. E4. J. Biosci. Bioeng. 2010, 110, 415–418. [Google Scholar] [CrossRef]
- Hartono, H.; Nurfitriani; Asnawati, F.; Citra, H.; Handayani, N.I.; Junda, M.; Ali, A.; Hala, Y.; Jumadi, O. Ability of ammonium excretion, indol acetic acid production, and phosphate solubilization of nitrogen-fixing bacteria isolated from crop rhizosphere and their effect on plant growth. ARPN J. Eng. Appl. Sci. 2016, 11, 11735–11741. [Google Scholar]
- Reyes, I.; Valery, A.; Valduz, Z. Phosphate-solubilizing microorganisms isolated from rhizospheric and bulk soils of colonizer plants at an abandoned rock phosphate mine. Plant Soil 2006, 287, 69–75. [Google Scholar] [CrossRef]
- Zheng, B.-X.; Ibrahim, M.; Zhang, D.-P.; Bi, Q.-F.; Li, H.-Z.; Zhou, G.-W.; Ding, K.; Peñuelas, J.; Zhu, Y.-G.; Yang, X.-R. Identification and characterization of inorganic-phosphate-solubilizing bacteria from agricultural fields with a rapid isolation method. AMB Express 2018, 8, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajawat, M.V.S.; Singh, S.; Tyagi, S.P.; Saxena, A.K. A Modified Plate Assay for Rapid Screening of Potassium-Solubilizing Bacteria. Pedosphere 2016, 26, 768–773. [Google Scholar] [CrossRef]
- Mohd Yusof, H.; Halimi, M.; Samsudin, A.A. Isolation and Characterisation of Sulphur Oxidizing Bacteria Isolated from Hot Spring in Malaysia for Biological Deodorisation of Hydrogen Sulphide in Chicken Manure. Media Peternak. 2017, 40, 178–187. [Google Scholar] [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K.; Verma, M. Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. 3 Biotech 2017, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Coffey, B.M.; Anderson, G.G. Biofilm formation in the 96-well microtiter plate. Methods Mol. Biol. 2014, 1149, 631–641. [Google Scholar] [CrossRef]
- Ambrosini, A.; Passaglia, L.M.P. Plant Growth–Promoting Bacteria (PGPB): Isolation and Screening of PGP Activities. Curr. Protoc. Plant Biol. 2017, 2, 190–209. [Google Scholar] [CrossRef]
- Li, Z.; Chang, S.; Lin, L.; Li, Y.; An, Q. A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Lett. Appl. Microbiol. 2011, 53, 178–185. [Google Scholar] [CrossRef]
- Takao, T.; Kitatani, F.; Watanabe, N.; Yagi, A.; Sakata, K. A Simple Screening Method for Antioxidants and Isolation of Several Antioxidants Produced by Marine Bacteria from Fish and Shellfish. Biosci. Biotechnol. Biochem. 1994, 58, 1780–1783. [Google Scholar] [CrossRef] [Green Version]
- Shafekh, E.S.; Rohin, M.A.K.; Catherine, C.C.W.; Syakiroh, S.Z.A.; Habibah, U.A.; Abd Hadi, N.; Farhanah, N.M.Y.; Husna, N.Z.; Nafizah, S.M.B.; Azlina, M.; et al. Total phenolic content and In vitro antioxidant activity of Vigna sinensis. Int. Food Res. J. 2012, 19, 1393–1400. [Google Scholar]
- Schmitz, L.; Yan, Z.; Schneijderberg, M.; de Roij, M.; Pijnenburg, R.; Zheng, Q.; Franken, C.; Dechesne, A.; Trindade, L.M.; van Velzen, R.; et al. Synthetic bacterial community derived from a desert rhizosphere confers salt stress resilience to tomato in the presence of a soil microbiome. ISME J. 2022, 16, 1907–1920. [Google Scholar] [CrossRef]
- Flynn, K.D.; Bagnall, D.K.; Morgan, C.L.S. Evaluation of SLAKES, a smartphone application for quantifying aggregate stability, in high-clay soils. Soil Sci. Soc. Am. J. 2020, 84, 345–353. [Google Scholar] [CrossRef]
- Fajardo, M.; Jones, E.; Wittig, R. Assessing soil aggregate stability with mobile phones. In Proceedings of the 22nd EGU General Assembly, Online, 4–8 May 2020; p. 3240. [Google Scholar]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.-X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Vilchez Morillas, J.I. Soil Characterization of Fire-Affected Places in Los Güájares Wildfire. 2023. Available online: https://figshare.com/articles/dataset/Soil_characterization_of_fire-affected_places_in_Los_G_jares_wildfire/22789820/1 (accessed on 10 May 2023). [CrossRef]
- Vilchez Morillas, J.I. Wildfire Strains Characterization. 2023. Available online: https://figshare.com/articles/dataset/Wildfire_strains_characterization/22815908/1 (accessed on 10 May 2023).
- Martínez-Fernández, D. Physiological responses of Bituminaria bituminosa to drought and heavy metals. J. Plant Physiol. 2012, 168, 2206–2211. [Google Scholar] [CrossRef]
- Correal, E.; Hoyos, A.; Ríos Ríos, S.; Méndez, P.; Real, D.; Snowball, R.; Costa, J. Seed production of Bituminaria bituminosa: Size, production, retention and germination capacity of the legumes. Options Mediterr. 2008, 79, 379–384. [Google Scholar]
- Kuzina, E.; Mukhamatdyarova, S.; Sharipova, Y.; Makhmutov, A.; Belan, L.; Korshunova, T. Influence of Bacteria of the Genus Pseudomonas on Leguminous Plants and Their Joint Application for Bioremediation of Oil Contaminated Soils. Plants 2022, 11, 3396. [Google Scholar] [CrossRef]
- Costa, A.; Corallo, B.; Amarelle, V.; Stewart, S.; Pan, D.; Tiscornia, S.; Fabiano, E. Paenibacillus sp. Strain UY79, Isolated from a Root Nodule of Arachis villosa, Displays a Broad Spectrum of Antifungal Activity. Appl. Environ. Microbiol. 2022, 88, e0164521. [Google Scholar] [CrossRef]
- Sherpa, M.T.; Bag, N.; Das, S.; Haokip, P.; Sharma, L. Isolation and characterization of plant growth promoting rhizobacteria isolated from organically grown high yielding pole type native pea (Pisum sativum L.) variety Dentami of Sikkim, India. Curr. Res. Microb. Sci. 2021, 2, 100068. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Dejene, T.; Benucci, G.M.N.; Mediavilla, O.; Hernández-Rodríguez, M.; Geml, J.; Baldrian, P.; Sanz-Benito, I.; Olaizola, J.; Bonito, G.; et al. Co-responses of bacterial and fungal communities to fire management treatments in Mediterranean pyrophytic ecosystems. Sci. Total Environ. 2023, 875, 162676. [Google Scholar] [CrossRef]
- Rai, D.; Silveira, M.L.; Strauss, S.L.; Meyer, J.L.; Castellano-Hinojosa, A.; Kohmann, M.M.; Brandani, C.B.; Gerber, S. Short-term prescribed fire-induced changes in soil microbial communities and nutrients in native rangelands of Florida. Appl. Soil Ecol. 2023, 189, 104914. [Google Scholar] [CrossRef]
- Soria, R.; Tortosa, A.; Rodríguez-Berbel, N.; Lucas-Borja, M.E.; Ortega, R.; Miralles, I. Short-Term Response of Soil Bacterial Communities after Prescribed Fires in Semi-Arid Mediterranean Forests. Fire 2023, 6, 145. [Google Scholar] [CrossRef]
- Pérez Matos, A.E.; Bacci, G.; Borruso, L.; Landolfi, M.; Petrocchi, D.; Renzi, S.; Perito, B. Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico. Microorganisms 2023, 11, 1130. [Google Scholar] [CrossRef] [PubMed]
- Hrenović, J.; Kisić, I.; Delač, D.; Durn, G.; Bogunović, I.; Mikulec, M.; Pereira, P. Short-Term Effects of Experimental Fire on Physicochemical and Microbial Properties of a Mediterranean Cambisol. Fire 2023, 6, 155. [Google Scholar] [CrossRef]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Adu-Gyamfi, J.J., Ed.; Springer: Dordrecht, The Netherlands, 2002; pp. 201–213. [Google Scholar]
- Carvalhais, L.C.; Dennis, P.G.; Fedoseyenko, D.; Hajirezaei, M.-R.; Borriss, R.; von Wirén, N. Root exudation of sugars, amino acids, and organic acids by maize as affected by nitrogen, phosphorus, potassium, and iron deficiency. J. Plant Nutr. Soil Sci. 2011, 174, 3–11. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Pantigoso, H.A.; Manter, D.K.; Fonte, S.J.; Vivanco, J.M. Root exudate-derived compounds stimulate the phosphorus solubilizing ability of bacteria. Sci. Rep. 2023, 13, 4050. [Google Scholar] [CrossRef]
- Singh, S.K.; Wu, X.; Shao, C.; Zhang, H. Microbial enhancement of plant nutrient acquisition. Stress Biol. 2022, 2, 3. [Google Scholar] [CrossRef]
- Yeager, C.M.; Northup, D.E.; Grow, C.C.; Barns, S.M.; Kuske, C.R. Changes in nitrogen-fixing and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire. Appl. Environ. Microbiol. 2005, 71, 2713–2722. [Google Scholar] [CrossRef] [Green Version]
- Kardol, P.; Yang, T.; Arroyo, D.N.; Teste, F.P. Plant-soil feedback in the ‘real world’: How does fire fit into all of this? Plant Soil 2023, 485, 91–102. [Google Scholar] [CrossRef]
- Hart, S.C.; DeLuca, T.H.; Newman, G.S.; MacKenzie, M.D.; Boyle, S.I. Post-fire vegetative dynamics as drivers of microbial community structure and function in forest soils. For. Ecol. Manag. 2005, 220, 166–184. [Google Scholar] [CrossRef]
- Jensen, M.; Michelsen, A.; Menassie, G. Responses in Plant, Soil Inorganic and Microbial Nutrient Pools to Experimental Fire, Ash and Biomass Addition in a Woodland Savanna. Oecologia 2001, 128, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Flores-Duarte, N.J.; Pajuelo, E.; Mateos-Naranjo, E.; Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S.; Carrasco López, J.A. A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2023, 24, 7003. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Hagerty, C.H.; Paulitz, T.C. Synthetic microbial consortia derived from rhizosphere soil protect wheat against a soilborne fungal pathogen. Front. Microbiol. 2022, 13, 908981. [Google Scholar] [CrossRef] [PubMed]
- Harvey, H.J.; Wildman, R.D.; Mooney, S.J.; Avery, S.V. Soil aggregates by design: Manufactured aggregates with defined microbial composition for interrogating microbial activities in soil microhabitats. Soil Biol. Biochem. 2020, 148, 107870. [Google Scholar] [CrossRef]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil Aggregate Microbial Communities: Towards Understanding Microbiome Interactions at Biologically Relevant Scales. Appl. Environ. Microbiol. 2019, 85, e00324-19. [Google Scholar] [CrossRef] [Green Version]
- Galloway, A.F.; Pedersen, M.J.; Merry, B.; Marcus, S.E.; Blacker, J.; Benning, L.G.; Field, K.J.; Knox, J.P. Xyloglucan is released by plants and promotes soil particle aggregation. New Phytol. 2018, 217, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Stone, M.; Emelko, M.B.; Droppo, I.G.; Silins, U. Biostabilization and erodibility of cohesive sediment deposits in wildfire-affected streams. Water Res. 2011, 45, 521–534. [Google Scholar] [CrossRef]
- Seneviratne, G.; Jayasekara, A.P.D.A.; De Silva, M.S.D.L.; Abeysekera, U.P. Developed microbial biofilms can restore deteriorated conventional agricultural soils. Soil Biol. Biochem. 2011, 43, 1059–1062. [Google Scholar] [CrossRef]
- Bogino, P.C.; Oliva Mde, L.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [Green Version]
- Qurashi, A.W.; Sabri, A.N. Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz. J. Microbiol. 2012, 43, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Navas, A.; González-López, J.; Arcos, S.C.; Manzanera, M. Biosafety Test for Plant Growth-Promoting Bacteria: Proposed Environmental and Human Safety Index (EHSI) Protocol. Front. Microbiol. 2015, 6, 1514. [Google Scholar] [CrossRef] [Green Version]
- Schoebitz, M.; Vidal, G. Microbial consortium and pig slurry to improve chemical properties of degraded soil and nutrient plant uptake (Schoebitz and Vidal, 2016). J. Soil Sci. Plant Nutr. 2016, 16, 226–236. [Google Scholar]
- Jacques, R.J.S.; Okeke, B.C.; Bento, F.M.; Teixeira, A.S.; Peralba, M.C.R.; Camargo, F.A.O. Microbial consortium bioaugmentation of a polycyclic aromatic hydrocarbons contaminated soil. Bioresour. Technol. 2008, 99, 2637–2643. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gnecco, G.; Covacevich, F.; Consolo, V.F.; Behr, J.H.; Sommermann, L.; Moradtalab, N.; Maccario, L.; Sørensen, S.J.; Deubel, A.; Schellenberg, I.; et al. Effect of Long-Term Agricultural Management on the Soil Microbiota Influenced by the Time of Soil Sampling. Front. Soil Sci. 2022, 2, 837508. [Google Scholar] [CrossRef]
- Dove, N.C.; Taş, N.; Hart, S.C. Ecological and genomic responses of soil microbiomes to high-severity wildfire: Linking community assembly to functional potential. ISME J. 2022, 16, 1853–1863. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | Location | GPS | Altitude (mamsl) | Geological Contex |
|---|---|---|---|---|---|
| 1 | Burnt (B1) | Cerro Lobera | 36°51′01.5″ N 3°36′25.4″ W | 377 | Marble with biotite 1 |
| Unburnt (UB1) | 36°51′04.5″ N 3°36′40.1″ W | 417 | Marble with biotite 1 | ||
| 2 | Burnt (B2) | Barranco del Girón | 36°52′11.1″ N 3°38′12.9″ W | 736 | Marble with tremolite 1, squists |
| Unburnt (UB2) | 36°52′14.8″ N 3°38′35.1″ W | 726 | Quartz-squists and squists 2 | ||
| 3 | Burnt (B3) | Alto de la Hoya de las Bolinas-Venta de la Cebada | 36°51′50.5″ N 3°32′32.2″ W | 745 | Dolomites, limestones, marbles |
| Unburnt (UB3) | 36°51′27.4″ N 3°32′38.6″ W | 731 | Alluvial sediment, dolomites, marbles |
| Sample a | Deep (cm) | Field Texture | pH | EC b | OM c (%) | Total N (%) | P (P2O5, mg/kg) | K (K2O, mg/kg) | Mg (mg/kg) | Ca (mg/kg) | Fe (mg/kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | 0–5 | Coarse | 8.5 | 0.19 | 6.70 VH | 0.298 H | 478 VH | 434 VH | 947 VH | 4308 VH | 234 VH |
| UB1 | 0–5 | Coarse | 8.5 | 0.01 | 1.05 L | 0.087 L | <23 VL | 80 A | 168 VH | 2964 H | 55 H |
| B2 | 0–5 | Coarse | 8.4 | 0.09 | 6.00 VH | 0.252 H | 446 VH | 799 VH | 1007 VH | 4972 VH | 659 VH |
| UB2 | 0–5 | Coarse | 8.1 | 0.02 | 6.70 VH | 0.279 H | 24 VL | 165 H | 183 VH | 5323 VH | 108 VH |
| B3 | 0–5 | Coarse | 8.2 | 0.06 | 3.80 H | 0.230 H | 530 VH | 412 VH | 479 VH | 5910 VH | 414 VH |
| UB3 | 0–5 | Coarse | 8.1 | 0.03 | 5.80 VH | 0.227 H | 645 VH | 424 VH | 210 VH | 4701 VH | 141 VH |
| B1 | 5–10 | Coarse | 8.3 | 0.03 | 3.90 H | 0.166 A | 53 A | 120 H | 508 VH | 3833 H | 116 VH |
| UB1 | 5–10 | Coarse | 8.7 | 0.01 | 0.75 L | 0.035 VL | <23 VL | 73 A | 186 VH | 3036 H | 42 H |
| B2 | 5–10 | Coarse | 8.3 | 0.06 | 8.00 VH | 0.295 H | 50 L | 353 VH | 930 VH | 3763 H | 412 VH |
| UB2 | 5–10 | Coarse | 8.1 | 0.01 | 4.00 H | 0.189 A | 24 VL | 130 H | 143 VH | 4069 VH | 91 VH |
| B3 | 5–10 | Coarse | 8.2 | 0.04 | 2.90 A | 0.105 A | 63 A | 155 H | 212 VH | 3589 VH | 142 VH |
| UB3 | 5–10 | Coarse | 8.1 | 0.01 | 2.20 A | 0.126 A | 190 H | 187 H | 116 H | 1675 A | 70 H |
| Location 1—Cerro Lobera | |||
|---|---|---|---|
| Burnt | Unburnt | ||
| Codename | Scientific Name | Codename | Scientific Name |
| A | Peribacillus frigoritolerans | AA | Unidentified |
| B | Bacillus cereus | AB | Pseudomonas koreensis |
| C | Paenibacillus lautus | AC | Pseudomonas koreensis |
| D | Bacillus mycoides | AD | Unidentified |
| E | Unidentified | AE | Paenarthrobacter nitroguajacolicus |
| F | Unidentified | AG | Unable to grow |
| E-alfa (EALPHA) | Pseudomonas koreensis | AF | Unidentified |
| AH | Pseudomonas lini | ||
| AI | Paenarthrobacter nitroguajacolicus | ||
| AJ | Peribacillus frigoritolerans | ||
| AK | Peribacillus frigoritolerans | ||
| Location 2—Barranco del Girón | |||
| Burnt | Unburnt | ||
| Codename | Scientific Name | Codename | Scientific Name |
| BA | Achromobacter spanius | CA | Peribacillus frigoritolerans/simplex |
| BB | Peribacillus frigoritolerans | CB | Peribacillus frigoritolerans/simplex |
| BC | Peribacillus frigoritolerans | CC | Unidentified |
| BD | Exiguobacterium sp. | CD | Bacillus toyonensis |
| BE | Pseudomonas fluorescens | CE | Peribacillus frigoritolerans |
| BF | Unidentified | CF | Pseudomonas fluorescens |
| BG | Exiguobacterium mexicanum | CG | Unidentified |
| CH | Flavobacterium sp. | ||
| CI | Paenarthrobacter nitroguajacolicus | ||
| Location 3—Alto de la Hoya de las Bolinas—Venta de la Cebada | |||
| Burnt | Unburnt | ||
| Codename | Scientific Name | Codename | Scientific Name |
| DA | Unidentified | EA | Paenarthrobacter nitroguajacolicus |
| DB | Unidentified | EB | Peribacillus frigoritolerans |
| DC | Pseudomonas fluorescens | EC | Unidentified |
| DD | Pseudomonas fluorescens | ED | Priestia megaterium |
| DE | Pseudomonas granadensis | EE | Unidentified |
| DF | Unable to grow | EF | Pseudomonas brassicacearum |
| DG | Unable to grow | ||
| DH | Unable to grow | ||
| Mix of Ashes | |||
|---|---|---|---|
| Codename | Scientific Name | Codename | Scientific Name |
| Ashes1 (ASH1) | Acinetobacter calcoaceticus | Ashes5 (ASH5) | Sphingobacterium sp. |
| Ashes2 (ASH2) | Unidentified | Ashes6 (ASH6) | Sphingobacterium sp. |
| Ashes3 (ASH3) | Pantoea agglomerans | Ashes7 (ASH7) | Acidovorax sp. |
| Ashes4 (ASH4) | Pseudomonas sp. | Ashes8 (ASH8) | Unidentified |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niza Costa, M.; Gil, T.; Teixeira, R.; Rodrígues dos Santos, A.S.; Rebelo Romão, I.; Sequero López, C.; Vílchez, J.I. Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guájares (Granada, Spain) Wildfire. Biology 2023, 12, 1093. https://doi.org/10.3390/biology12081093
Niza Costa M, Gil T, Teixeira R, Rodrígues dos Santos AS, Rebelo Romão I, Sequero López C, Vílchez JI. Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guájares (Granada, Spain) Wildfire. Biology. 2023; 12(8):1093. https://doi.org/10.3390/biology12081093
Chicago/Turabian StyleNiza Costa, Marla, Tatiana Gil, Raquel Teixeira, Ana Sofía Rodrígues dos Santos, Inês Rebelo Romão, Cristina Sequero López, and Juan Ignacio Vílchez. 2023. "Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guájares (Granada, Spain) Wildfire" Biology 12, no. 8: 1093. https://doi.org/10.3390/biology12081093
APA StyleNiza Costa, M., Gil, T., Teixeira, R., Rodrígues dos Santos, A. S., Rebelo Romão, I., Sequero López, C., & Vílchez, J. I. (2023). Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guájares (Granada, Spain) Wildfire. Biology, 12(8), 1093. https://doi.org/10.3390/biology12081093