
LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by Acetylsalicylic Acid: Behavioral and Transcriptional Evidence
 , , ,
, , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
- -
- -
- the Molluscan Defense Molecule (LymMDM), an Ig-superfamily member, which allows mollusks to mount an effective immune response and ensure their survival [32],
- -
2. Materials and Methods
2.1. Animals
2.2. LPS Treatment
2.3. Acetylsalicylic Acid (ASA) Treatment
2.4. Behavioral Procedure to Induce a Garcia Effect
2.5. Experimental Design
2.5.1. Experiment 1: Transcriptional Effects of ASA, LPS, and Their Paired Exposure
2.5.2. Experiment 2: LPS-Induced Garcia Effect Procedure and Its Pharmacological Regulation Mediated by ASA (Behavioral Data)
- Snails of the ‘Saline_C group’ were first injected with snail saline, and 3 h later, were exposed to the novel food (carrot slurry) for 2 min, and the number of rasps was recorded. Thus, these snails were not exposed to the novel taste (i.e., carrot slurry) before the injection.
- Snails of the ‘Saline group’ were exposed to carrot slurry for 2 min and the number of rasps was counted. One hour later, snails were injected with snail saline. After 3 h, snails were re-exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was again recorded.
- Snails of the ‘LPS-C group’ were injected with LPS and 3 h later were exposed to the carrot slurry for the first time. The number of rasps elicited by the carrot slurry was again recorded for 2 min.
- Snails of the ‘LPS group’ were exposed to carrot slurry for 2 min, during which the number of rasps was counted. One hour later, snails were injected with LPS and, 3 h later, were re-exposed to carrot slurry for 2 min, and the number of rasps elicited by the carrot slurry was again recorded.
- Snails of the ‘ASA group’ were exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was counted. One hour later, snails were exposed to ASA for 1 h. Three hours later, snails were re-exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was again recorded.
- Snails of the ‘ASA_Sal group’ were exposed to carrot slurry for 2 min and the number of rasps was counted. One hour later, snails were exposed to ASA for 1 h and immediately after were injected with snail saline. After 3 h, snails were re-exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was again recorded.
- Snails of the ‘Garcia effect group’ were exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was counted. One hour later, snails were injected with LPS. Three hours later, snails were re-exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was again recorded.
- Snails of the ‘ASA_LPS group’ were exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was counted. One hour later, snails were exposed to ASA for 1 h and immediately after were injected with LPS. Three hours later, snails were re-exposed to carrot slurry for 2 min and the number of rasps elicited by the carrot slurry was again recorded.
2.5.3. Experiment 3: Transcriptional Effects Induced by the LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by ASA
2.6. Total RNA Extraction, Reverse Transcription, and Real-Time Quantitative PCR
2.7. Data Analysis
3. Results
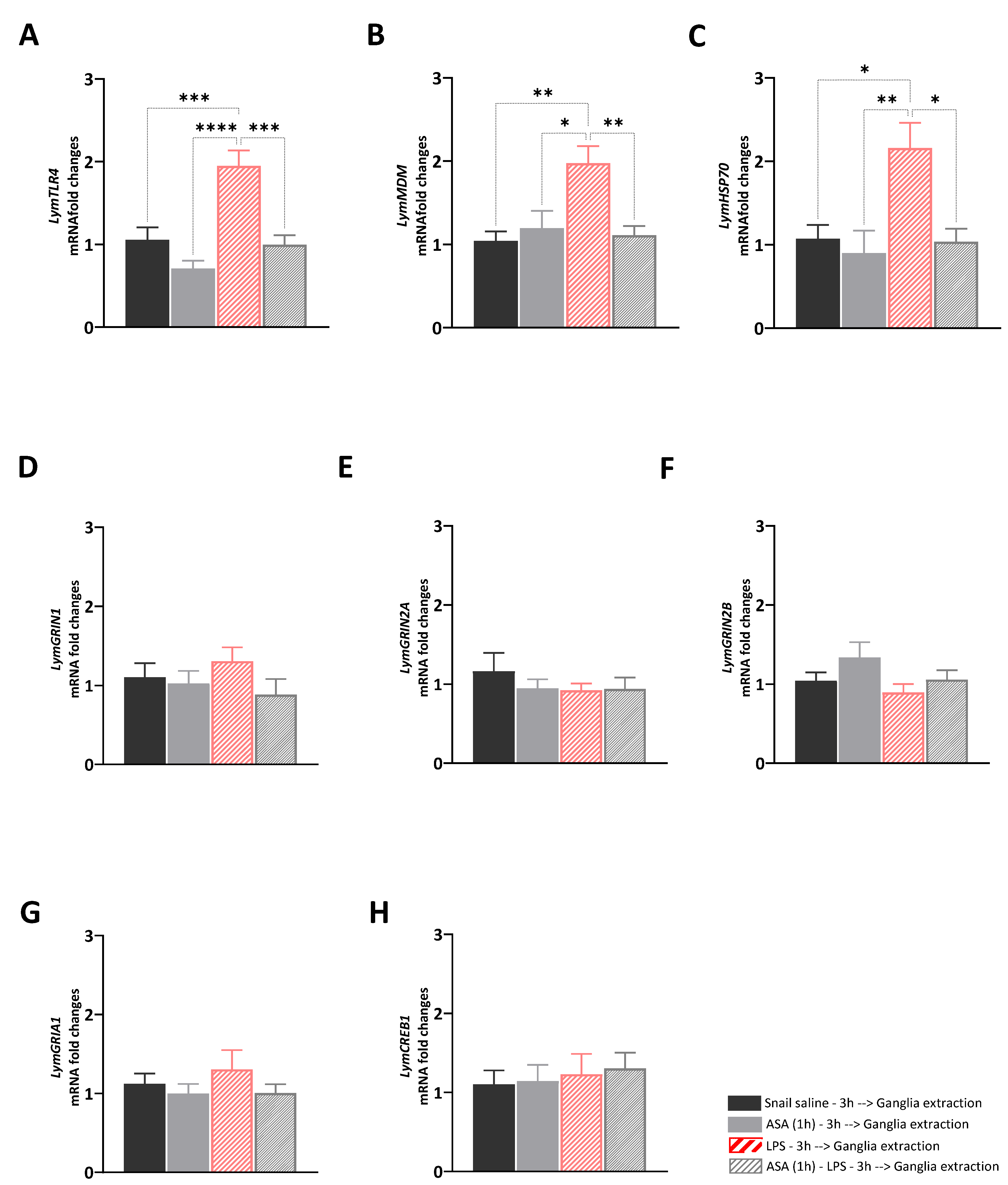
3.1. Experiment 1: Transcriptional Effects of ASA, LPS, and Their Paired Exposure
3.2. Experiment 2: LPS-Induced Garcia Effect Procedure and Its Pharmacological Regulation Mediated by ASA
3.3. Experiment 3: Transcriptional Effects Induced by the LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by ASA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rivi, V.; Benatti, C.; Lukowiak, K.; Colliva, C.; Alboni, S.; Tascedda, F.; Blom, J.M.C. What Can We Teach Lymnaea and What Can Lymnaea Teach Us? Biol. Rev. Camb. Philos. Soc. 2021, 96, 1590–1602. [Google Scholar] [CrossRef]
- Fodor, I.; Svigruha, R.; Kemenes, G.; Kemenes, I.; Pirger, Z. The Great Pond Snail (Lymnaea stagnalis) as a Model of Ageing and Age-Related Memory Impairment: An Overview. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 975–982. [Google Scholar] [CrossRef]
- Fodor, I.; Hussein, A.A.; Benjamin, P.R.; Koene, J.M.; Pirger, Z. The Unlimited Potential of the Great Pond Snail, Lymnaea stagnalis. eLife 2020, 9, e56962. [Google Scholar] [CrossRef]
- Ito, E.; Yamagishi, M.; Hatakeyama, D.; Watanabe, T.; Fujito, Y.; Dyakonova, V.; Lukowiak, K. Memory Block: A Consequence of Conflict Resolution. J. Exp. Biol. 2015, 218, 1699–1704. [Google Scholar] [CrossRef] [Green Version]
- Lukowiak, K.; Sangha, S.; Scheibenstock, A.; Parvez, K.; McComb, C.; Rosenegger, D.; Varshney, N.; Sadamoto, H. A Molluscan Model System in the Search for the Engram. J. Physiol. Paris 2003, 97, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Benatti, C.; Rigillo, G.; Blom, J.M.C. Invertebrates as Models of Learning and Memory: Investigating Neural and Molecular Mechanisms. J. Exp. Biol. 2023, 226, jeb244844. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Batabyal, A.; Benatti, C.; Tascedda, F.; Blom, J.M.C.; Lukowiak, K. Prey Populations with Different Predation Histories Show Differences in Behavioral and Transcriptional Effects under Acute Predation Threat. Neurobiol. Learn. Mem. 2023, 203, 107775. [Google Scholar] [CrossRef] [PubMed]
- Kagan, D.; Batabyal, A.; Rivi, V.; Lukowiak, K. A Change in Taste: The Role of MicroRNAs in Altering Hedonic Value. J. Exp. Biol. 2022, 225, jeb243840. [Google Scholar] [CrossRef]
- Kagan, D.; Rivi, V.; Benatti, C.; Tascedda, F.; Blom, J.M.C.; Lukowiak, K. No Food for Thought: An Intermediate Level of Food Deprivation Enhances Memory in Lymnaea stagnalis. J. Exp. Biol. 2023, 226, jeb245566. [Google Scholar] [CrossRef]
- Swinton, C.; Swinton, E.; Shymansky, T.; Hughes, E.; Zhang, J.; Rothwell, C.; Kakadiya, M.; Lukowiak, K. Configural Learning: A Higher Form of Learning in Lymnaea. J. Exp. Biol. 2019, 222, jeb190405. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, P.R.; Kemenes, G. Invertebrate Models to Study Learning and Memory: Lymnaea. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 197–204. ISBN 978-0-08-045046-9. [Google Scholar]
- Benjamin, P.R. Lymnaea. Scholarpedia 2008, 3, 4124. [Google Scholar] [CrossRef]
- Totani, Y.; Nakai, J.; Dyakonova, V.E.; Lukowiak, K.; Sakakibara, M.; Ito, E. Induction of LTM Following an Insulin Injection. eNeuro 2020, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totani, Y.; Aonuma, H.; Oike, A.; Watanabe, T.; Hatakeyama, D.; Sakakibara, M.; Lukowiak, K.; Ito, E. Monoamines, Insulin and the Roles They Play in Associative Learning in Pond Snails. Front. Behav. Neurosci. 2019, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Kemenes, G.; Benjamin, P.R. Appetitive Learning in Snails Shows Characteristics of Conditioning in Vertebrates. Brain Res. 1989, 489, 163–166. [Google Scholar] [CrossRef]
- Kemenes, I.; Straub, V.A.; Nikitin, E.S.; Staras, K.; O’Shea, M.; Kemenes, G.; Benjamin, P.R. Role of Delayed Nonsynaptic Neuronal Plasticity in Long-Term Associative Memory. Curr. Biol. 2006, 16, 1269–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivi, V.; Batabyal, A.; Wiley, B.; Benatti, C.; Tascedda, F.; Blom, J.M.C.; Lukowiak, K. Fluoride Affects Memory by Altering the Transcriptional Activity in the Central Nervous System of Lymnaea stagnalis. NeuroToxicology 2022, 92, 61–66. [Google Scholar] [CrossRef]
- Rivi, V.; Benatti, C.; Colliva, C.; Radighieri, G.; Brunello, N.; Tascedda, F.; Blom, J.M.C. Lymnaea stagnalis as Model for Translational Neuroscience Research: From Pond to Bench. Neurosci. Biobehav. Rev. 2020, 108, 602–616. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Batabyal, A.; Benatti, C.; Blom, J.M.; Tascedda, F.; Lukowiak, K. A Flavonoid, Quercetin, Is Capable of Enhancing Long-Term Memory Formation If Encountered at Different Times in the Learning, Memory Formation, and Memory Recall Continuum. J. Comp. Physiol. A 2021, 208, 253–265. [Google Scholar] [CrossRef]
- Benatti, C.; Rivi, V.; Colliva, C.; Radighieri, G.; Tascedda, F.; Blom, J.M.C. Redefining Operant Conditioning of Escape Behaviour in Lymnaea stagnalis. Invert. Sur. J. 2020, 17, 129–137. [Google Scholar] [CrossRef]
- Garcia, J.; Kimeldorf, D.J.; Koelling, R.A. Conditioned Aversion to Saccharin Resulting from Exposure to Gamma Radiation. Science 1955, 122, 157–158. [Google Scholar] [CrossRef]
- Garcia, J.; Lasiter, P.S.; Bermudez-Rattoni, F.; Deems, D.A. A General Theory of Aversion Learning. Ann. N. Y. Acad. Sci. 1985, 443, 8–21. [Google Scholar] [CrossRef]
- Garcia, J.; Koelling, R.A. Relation of Cue to Consequence in Avoidance Learning. Psychon. Sci. 1966, 4, 123–124. [Google Scholar] [CrossRef] [Green Version]
- Rivi, V.; Batabyal, A.; Juego, K.; Kakadiya, M.; Benatti, C.; Blom, J.M.C.; Lukowiak, K. To Eat or Not to Eat: A Garcia Effect in Pond Snails (Lymnaea stagnalis). J. Comp. Physiol. A 2021, 207, 479–495. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Batabyal, A.; Benatti, C.; Blom, J.M.C.; Lukowiak, K. Nature versus Nurture in Heat Stress-Induced Learning between Inbred and Outbred Populations of Lymnaea stagnalis. J. Therm. Biol. 2022, 103, 103170. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Benatti, C.; Actis, P.; Tascedda, F.; Blom, J.M.C. Behavioral and Transcriptional Effects of Short or Prolonged Fasting on the Memory Performances of Lymnaea stagnalis. Neuroendo 2022, 113, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Batabyal, A.; Benatti, C.; Blom, J.M.; Tascedda, F.; Lukowiak, K. Novel Taste, Sickness, and Memory: Lipopolysaccharide to Induce a Garcia-like Effect in Inbred and Wild Strains of Lymnaea stagnalis. Physiol. Behav. 2023, 263, 114137. [Google Scholar] [CrossRef] [PubMed]
- Rivi, V.; Batabyal, A.; Benatti, C.; Tascedda, F.; Blom, J.M.C.; Lukowiak, K. Aspirin Reverts Lipopolysaccharide-Induced Learning and Memory Impairment: First Evidence from an Invertebrate Model System. Naunyn Schmiedeberg’s Arch. Pharmacol. 2022, 395, 1573–1585. [Google Scholar] [CrossRef]
- Juhász, A.; Lawton, S.P. Toll like Receptors and Their Evolution in the Lymnaeid Freshwater Snail Species Radix auricularia and Lymnaea stagnalis, Key Intermediate Hosts for Zoonotic Trematodes. Dev. Comp. Immunol. 2022, 127, 104297. [Google Scholar] [CrossRef]
- Nie, L.; Cai, S.-Y.; Shao, J.-Z.; Chen, J. Toll-Like Receptors, Associated Biological Roles, and Signaling Networks in Non-Mammals. Front. Immunol. 2018, 9, 1523. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 Signal Transduction Pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Hoek, R.M.; Smit, A.B.; Frings, H.; Vink, J.M.; de Jong-Brink, M.; Geraerts, W.P. A New Ig-Superfamily Member, Molluscan Defence Molecule (MDM) from Lymnaea stagnalis, Is down-Regulated during Parasitosis. Eur. J. Immunol. 1996, 26, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Bedford, T.B.C.; Albright, D.R.; Michalak, P. Evolvability of HSP70 Expression under Artificial Selection for Inducible Thermotolerance in Independent Populations of Drosophila melanogaster. Physiol. Biochem. Zool. 2002, 75, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G. Heat Shock Protein 70 (Hsp70): Membrane Location, Export and Immunological Relevance. Methods 2007, 43, 229–237. [Google Scholar] [CrossRef]
- Yu, E.; Yoshinaga, T.; Jalufka, F.L.; Ehsan, H.; Mark Welch, D.B.; Kaneko, G. The Complex Evolution of the Metazoan HSP70 Gene Family. Sci. Rep. 2021, 11, 17794. [Google Scholar] [CrossRef]
- Frank, D.A.; Greenberg, M.E. CREB: A Mediator of Long-Term Memory from Mollusks to Mammals. Cell 1994, 79, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Korneev, S.A.; Vavoulis, D.V.; Naskar, S.; Dyakonova, V.E.; Kemenes, I.; Kemenes, G. A CREB2-Targeting MicroRNA Is Required for Long-Term Memory after Single-Trial Learning. Sci. Rep. 2018, 8, 3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadamoto, H.; Sato, H.; Kobayashi, S.; Murakami, J.; Aonuma, H.; Ando, H.; Fujito, Y.; Hamano, K.; Awaji, M.; Lukowiak, K.; et al. CREB in the Pond Snail Lymnaea stagnalis: Cloning, Gene Expression, and Function in Identifiable Neurons of the Central Nervous System. J. Neurobiol. 2004, 58, 455–466. [Google Scholar] [CrossRef]
- Batabyal, A.; Rivi, V.; Benatti, C.; Blom, J.M.C.; Lukowiak, K. Long-Term Memory of Configural Learning Is Enhanced via CREB Upregulation by the Flavonoid Quercetin in Lymnaea stagnalis. J. Exp. Biol. 2021, 224, jeb242761. [Google Scholar] [CrossRef]
- Dong, N.; Bandura, J.; Zhang, Z.; Wang, Y.; Labadie, K.; Noel, B.; Davison, A.; Koene, J.; Sun, H.-S.; Coutellec, M.-A.; et al. Ion Channel Profiling of the Lymnaea stagnalis Ganglia via Transcriptome Analysis. BMC Genom. 2021, 22, 18. [Google Scholar] [CrossRef]
- Kandel, E.R. The Molecular Biology of Memory: CAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Kandel, E.R.; Dudai, Y.; Mayford, M.R. The Molecular and Systems Biology of Memory. Cell 2014, 157, 163–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberini, C.M. The Role of Reconsolidation and the Dynamic Process of Long-Term Memory Formation and Storage. Front. Behav. Neurosci. 2011, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberini, C.M. Transcription Factors in Long-Term Memory and Synaptic Plasticity. Physiol. Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Benatti, C.; Colliva, C.; Blom, J.; Ottaviani, E.; Tascedda, F. Transcriptional Effect of Serotonin in the Ganglia of Lymnaea stagnalis. Inv. Surv. J. 2017, 14, 251–258. [Google Scholar] [CrossRef]
- Aonuma, H.; Kaneda, M.; Hatakeyama, D.; Watanabe, T.; Lukowiak, K.; Ito, E. Relationship between the Grades of a Learned Aversive-Feeding Response and the Dopamine Contents in Lymnaea. Biol. Open 2016, 5, 1869–1873. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, P.R. Gastropod Feeding: Behavioural and Neural Analysis of a Complex Multicomponent System. Symp. Soc. Exp. Biol. 1983, 37, 159–193. [Google Scholar]
- Benjamin, P.R. Distributed Network Organization Underlying Feeding Behavior in the Mollusk Lymnaea. Neural Syst. Circuits 2012, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, E.; Franceschi, C. The Neuroimmunology of Stress from Invertebrates to Man. Prog. Neurobiol. 1996, 48, 421–440. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 Trafficking and Its Influence on LPS-Induced pro-Inflammatory Signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Dantzer, R.; Kelley, K.W. Twenty Years of Research on Cytokine-Induced Sickness Behavior. Brain Behav. Immun. 2007, 21, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.; Berk, M.; Goehler, L.; Song, C.; Anderson, G.; Gałecki, P.; Leonard, B. Depression and Sickness Behavior Are Janus-Faced Responses to Shared Inflammatory Pathways. BMC Med. 2012, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatsepina, O.G.; Nikitina, E.A.; Shilova, V.Y.; Chuvakova, L.N.; Sorokina, S.; Vorontsova, J.E.; Tokmacheva, E.V.; Funikov, S.Y.; Rezvykh, A.P.; Evgen’ev, M.B. HSP70 Affects Memory Formation and Behaviorally Relevant Gene Expression in Drosophila melanogaster. Cell Stress. Chap 2021, 26, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [Green Version]
- Porto, R.R.; Dutra, F.D.; Crestani, A.P.; Holsinger, R.M.D.; Quillfeldt, J.A.; Homem de Bittencourt, P.I.; de Oliveira Alvares, L. HSP70 Facilitates Memory Consolidation of Fear Conditioning through MAPK Pathway in the Hippocampus. Neuroscience 2018, 375, 108–118. [Google Scholar] [CrossRef]
- Seppälä, O.; Walser, J.-C.; Cereghetti, T.; Seppälä, K.; Salo, T.; Adema, C.M. Transcriptome Profiling of a Multiuse Model Species Lymnaea stagnalis (Gastropoda) for Ecoimmunological Research. bioRxiv 2020, 22, 144. [Google Scholar] [CrossRef]
- Mobbs, D.; Hagan, C.C.; Dalgleish, T.; Silston, B.; Prévost, C. The Ecology of Human Fear: Survival Optimization and the Nervous System. Front. Neurosci. 2015, 9, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snijders, L.; Thierij, N.M.; Appleby, R.; St. Clair, C.C.; Tobajas, J. Conditioned Taste Aversion as a Tool for Mitigating Human-Wildlife Conflicts. Front. Conserv. Sci. 2021, 2, 744704. [Google Scholar] [CrossRef]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdère, P.; Delbecque, J.-P.; Cattaert, D. Comparative Behavior. Anxiety-like Behavior in Crayfish Is Controlled by Serotonin. Science 2014, 344, 1293–1297. [Google Scholar] [CrossRef]
- Benatti, C.; Rivi, V.; Alboni, S.; Grilli, A.; Castellano, S.; Pani, L.; Brunello, N.; Blom, J.M.C.B.; Bicciato, S.; Tascedda, F. Identification and Characterization of the Kynurenine Pathway in the Pond Snail Lymnaea stagnalis. Sci. Rep. 2022, 12, 15617. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Gene Bank Accession | Target | Product Length (bp) | Type Sequence |
|---|---|---|---|
| X15542.1 | Snail, beta-tubulin LymbTUB | 100 bp (92–192) | FW: GAAATAGCACCGCCATCC |
| RV: CGCCTCTGTGAACTCCATCT | |||
| DQ278441.1 | Lymnaea stagnalis elongation factor 1-alpha, LymEF1α | 150 bp (7–157) | FW: GTGTAAGCAGCCCTCGAACT |
| RV: TTCGCTCATCAATACCACCA | |||
| AY577328.1 | Lymnaea stagnalis Toll-like receptor 4 LymTLR4 | 100 bp (74–174) | FW: GGAGGGTCAAGCATAAAGTGT |
| RV: CATCAAGGTCAACGCCAAT | |||
| U58769.1 | Lymnaea stagnalis molluscan defense molecule precursor LymMDM | 104 bp (1614–1718) | FW: CGGGTACACACACAGATGGA |
| RV: TGACTGAACATTGGGCACAC | |||
| DQ206432.1 | Lymnaea stagnalis heat-shock protein 70 LymHSP70 | 199 bp (134–333) | FW: AGGCAGAGATTGGCAGGAT |
| RV: CCATTTCATTGTGTCGTTGC | |||
| AY571900.1 | Lymnaea stagnalis NMDA-type glutamate receptor subunit 1 LymGRIN1 | 140 bp (831–917) | FW: AGAGGATGCATCTACAATTT |
| RV: CCATTTACTAGGTGAACTCC | |||
| FX180835 | Lymnaea stagnalis NMDA-type glutamate receptor subunit 2A LymGRIN2A | 129 bp (3454–3583) | FW: GATCACCAAGGATGATTACT |
| RV: CTTGGCTATATTCAAGTCTGT | |||
| FX180839 | Lymnaea stagnalis NMDA-type glutamate receptor subunit 2B LymGRIN2B | 126 bp (4147–4273) | FW: GACTCCTCTGTTTTGGAATA |
| RV: GGTTCCTTGATGGTTTATTA | |||
| FX183516.1 | Lymnaea stagnalis AMPA-type glutamate receptor subunit 1 LymGRIA1 | 111 bp (1205–1316) | FW: AGACTGTTGTAGCTGTCCTT |
| RV: ATAGCTATTGGATTTCTTGC | |||
| AB041522.1 | Lymnaea stagnalis cAMP responsive element binding protein LymCREB1 | 180 bp (49–229) | FW: GTCAGCAGGGAATGGTCCTG |
| RV: ACCGCAGCAACCCTAACAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivi, V.; Batabyal, A.; Lukowiak, K.; Benatti, C.; Rigillo, G.; Tascedda, F.; Blom, J.M.C. LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by Acetylsalicylic Acid: Behavioral and Transcriptional Evidence. Biology 2023, 12, 1100. https://doi.org/10.3390/biology12081100
Rivi V, Batabyal A, Lukowiak K, Benatti C, Rigillo G, Tascedda F, Blom JMC. LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by Acetylsalicylic Acid: Behavioral and Transcriptional Evidence. Biology. 2023; 12(8):1100. https://doi.org/10.3390/biology12081100
Chicago/Turabian StyleRivi, Veronica, Anuradha Batabyal, Ken Lukowiak, Cristina Benatti, Giovanna Rigillo, Fabio Tascedda, and Joan M. C. Blom. 2023. "LPS-Induced Garcia Effect and Its Pharmacological Regulation Mediated by Acetylsalicylic Acid: Behavioral and Transcriptional Evidence" Biology 12, no. 8: 1100. https://doi.org/10.3390/biology12081100