
Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem



Abstract
:Simple Summary
Abstract

1. Introduction
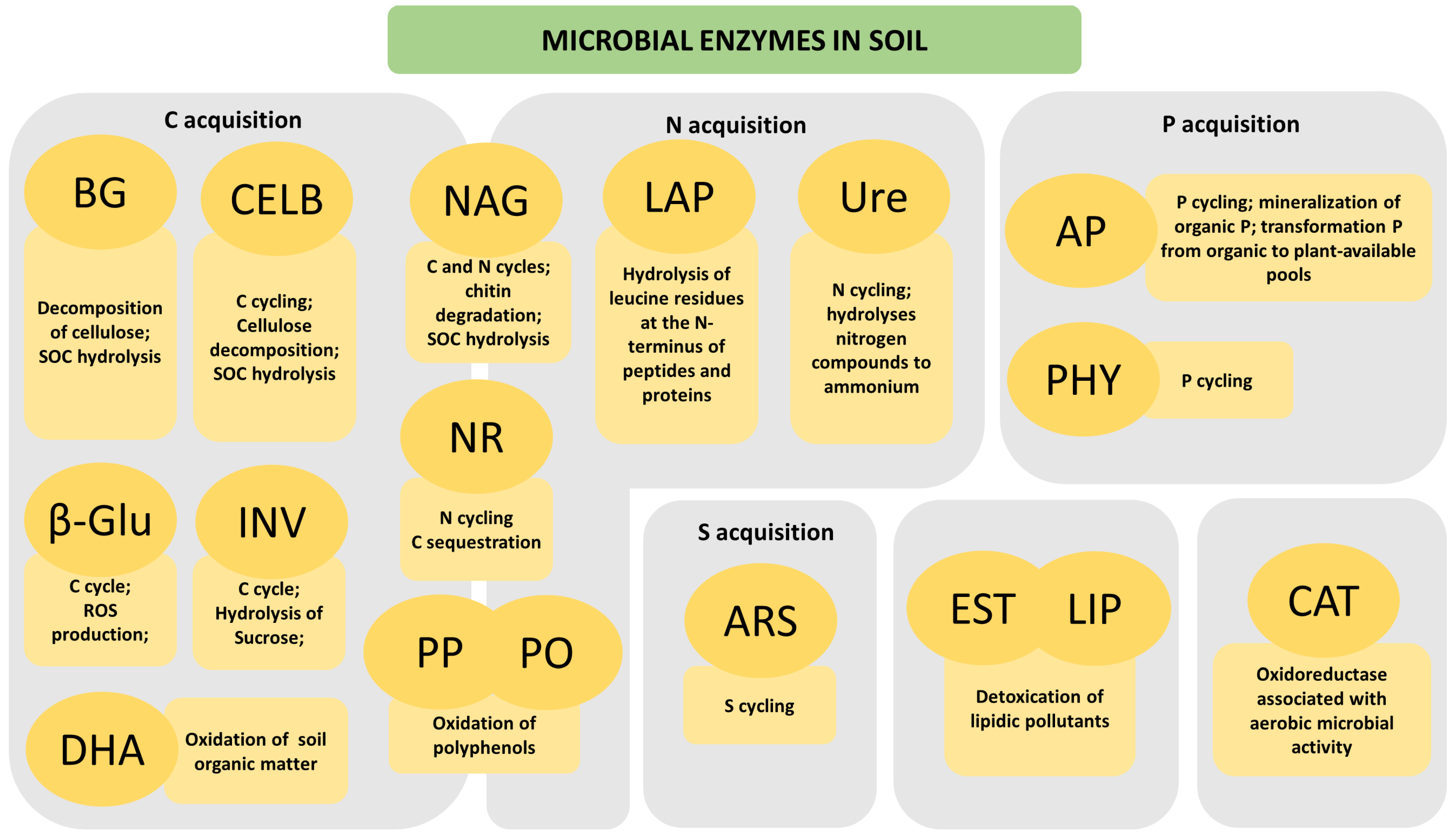
2. Main Microbial Enzymes in Soil
2.1. β-Glucosidases
2.2. β-1,4-N-Acetyl-Glucosaminidases
2.3. Invertases
2.4. Leucine Aminopeptidases
2.5. Ureases
2.6. Acid/Alkaline Phosphatases
2.7. Sulfatases
2.8. Dehydrogenases
2.9. Other Enzymes
2.9.1. Cellulases
2.9.2. Lipolytic Enzymes (Lipases, Carboxylesterases and Other Esterases)
2.9.3. Phenol Oxidases, Peroxidases
2.9.4. Catalases
2.9.5. Nitrate Reductases
3. Soil Microbial Enzyme Activities and Challenges in Different Soil Ecosystems
3.1. Forest and Grassland
3.2. Tropical and Subtropical Regions
3.3. Arid Lands/Desert
3.4. Saline Regions
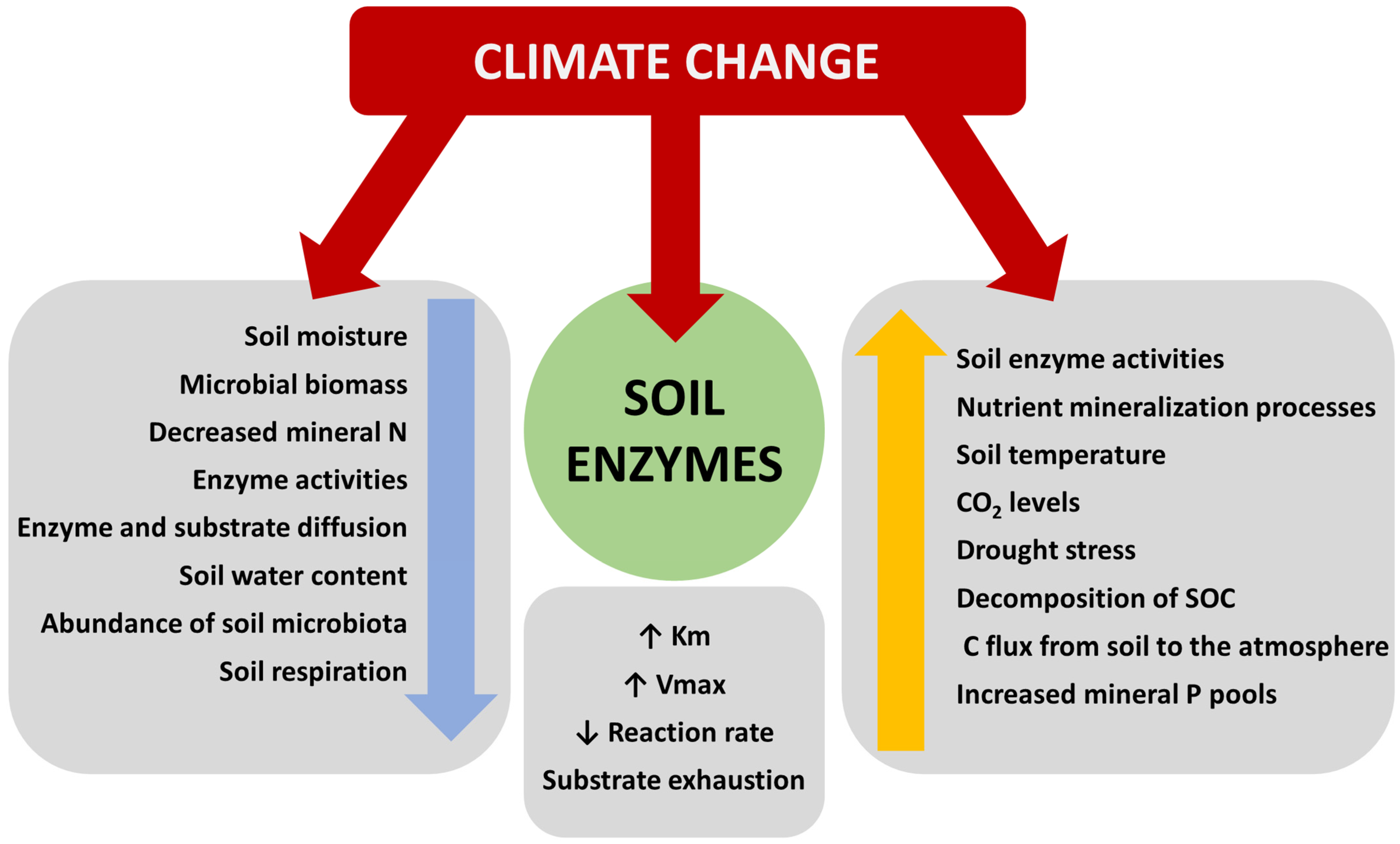
4. Role of Microbial Enzymes in C Sequestration and Enzyme Activity Shift through Climate Changes
4.1. Enzymes Activity at Different Climate Conditions
4.1.1. Arctic and Permafrost Regions
4.1.2. Tundra and Boreal Systems
4.1.3. Tropics and Subtropics
4.2. Strategies for C Sequestration and Enzymes Activities
4.2.1. Land Use/Conversion
4.2.2. Biochars
5. Influence of Intensive Agriculture on the Microbial Enzymatic Activity
5.1. Influence of N and/or P Addition to Soil on the Activity of Soil Microbial Enzymes
5.2. Influence of Herbicides and Other Agriculture Additivities on the Soil Enzymes
6. Current Challenges and Future Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Banerjee, S.; Van Der Heijden, M.G.A. Soil Microbiomes and One Health. Nat. Rev. Microbiol. 2023, 21, 6–20. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The Biomass Distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, B.A. Enzyme Activities as a Component of Soil Biodiversity: A Review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, Z.; Ma, P.; Wang, Z.; Niu, D.; Fu, H.; Elser, J.J. Effects of Grassland Degradation on Ecological Stoichiometry of Soil Ecosystems on the Qinghai-Tibet Plateau. Sci. Total Environ. 2020, 722, 137910. [Google Scholar] [CrossRef] [PubMed]
- Stone, M.M.; Weiss, M.S.; Goodale, C.L.; Adams, M.B.; Fernandez, I.J.; German, D.P.; Allison, S.D. Temperature Sensitivity of Soil Enzyme Kinetics under N-fertilization in Two Temperate Forests. Glob. Change Biol. 2012, 18, 1173–1184. [Google Scholar] [CrossRef]
- Allison, S.D.; Wallenstein, M.D.; Bradford, M.A. Soil-Carbon Response to Warming Dependent on Microbial Physiology. Nat. Geosci. 2010, 3, 336–340. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, X.; Ma, S.; Wu, Y.; Qiu, L.; Sun, P.; Li, Q. Widespread Increasing Ecosystem Water Limitation During the Past Three Decades in the Yellow River Basin, China. J. Geophys. Res. Biogeosci. 2023, 128, e2022JG007140. [Google Scholar] [CrossRef]
- Nandan, A.; Nampoothiri, K.M. Therapeutic and Biotechnological Applications of Substrate Specific Microbial Aminopeptidases. Appl. Microbiol. Biotechnol. 2020, 104, 5243–5257. [Google Scholar] [CrossRef]
- Farzadfar, S.; Knight, J.D.; Congreves, K.A. Soil Organic Nitrogen: An Overlooked but Potentially Significant Contribution to Crop Nutrition. Plant Soil 2021, 462, 7–23. [Google Scholar] [CrossRef]
- Vranova, V.; Rejsek, K.; Formanek, P. Proteolytic Activity in Soil: A Review. Appl. Soil Ecol. 2013, 70, 23–32. [Google Scholar] [CrossRef]
- Turner, B.L.; Joseph Wright, S. The Response of Microbial Biomass and Hydrolytic Enzymes to a Decade of Nitrogen, Phosphorus, and Potassium Addition in a Lowland Tropical Rain Forest. Biogeochemistry 2014, 117, 115–130. [Google Scholar] [CrossRef]
- Yan, B.; Sun, Y.; He, G.; He, R.; Zhang, M.; Fang, H.; Shi, L. Nitrogen Enrichment Affects Soil Enzymatic Stoichiometry via Soil Acidification in Arid and Hot Land. Pedobiologia 2020, 81–82, 150663. [Google Scholar] [CrossRef]
- Uwituze, Y.; Nyiraneza, J.; Fraser, T.D.; Dessureaut-Rompré, J.; Ziadi, N.; Lafond, J. Carbon, Nitrogen, Phosphorus, and Extracellular Soil Enzyme Responses to Different Land Use. Front. Soil Sci. 2022, 2, 814554. [Google Scholar] [CrossRef]
- Feng, H.; Sun, Y.; Zhi, Y.; Wei, X.; Luo, Y.; Mao, L.; Zhou, P. Identification and Characterization of the Nitrate Assimilation Genes in the Isolate of Streptomyces griseorubens JSD-1. Microb. Cell Factories 2014, 13, 174. [Google Scholar] [CrossRef]
- Giles, M.; Morley, N.; Baggs, E.M.; Daniell, T.J. Soil Nitrate Reducing Processes—Drivers, Mechanisms for Spatial Variation, and Significance for Nitrous Oxide Production. Front. Microbiol. 2012, 3, 00407. [Google Scholar] [CrossRef] [PubMed]
- Sibevieh, S.; Salehghamari, E.; Amoozegar, M.A.; Zolfaghari, M.R.; Soleimani, M.; Nasrollahzadeh, Z.; Eftekhari Yazdi, S. Bioassay and Molecular Screening of Nitrate Reductase Enzymes in Halophilic Bacteria. J. Cell Mol. Res. 2021, 13, 19–26. [Google Scholar] [CrossRef]
- Yokoyama, D.; Mori, T.; Wagai, R.; Hiradate, S.; Kitayama, K. Characteristics of Phosphorus Fractions in the Soils Derived from Sedimentary and Serpentinite Rocks in Lowland Tropical Rain Forests, Borneo. Soil Sci. Plant Nutr. 2018, 64, 218–221. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic Stoichiometry of Microbial Nutrient Acquisition in Tropical Soils. Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of Soil Physicochemical Properties and Enzyme Activities to Long-Term Reclamation of Coastal Saline Soil, Eastern China. Sci. Total Environ. 2017, 607–608, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Štursová, M.; Baldrian, P. Effects of Soil Properties and Management on the Activity of Soil Organic Matter Transforming Enzymes and the Quantification of Soil-Bound and Free Activity. Plant Soil 2011, 338, 99–110. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil Enzymes in a Changing Environment: Current Knowledge and Future Directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clément, J.-C.; Lavorel, S. Linkages between Extracellular Enzyme Activities and the Carbon and Nitrogen Content of Grassland Soils. Soil Biol. Biochem. 2016, 96, 198–206. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Wang, L.; Duan, Y.; Yao, B.; Chen, Y.; Cao, W. Soil Extracellular Enzyme Stoichiometry Reflects Microbial Metabolic Limitations in Different Desert Types of Northwestern China. Sci. Total Environ. 2023, 874, 162504. [Google Scholar] [CrossRef] [PubMed]
- Rosinger, C.; Rousk, J.; Sandén, H. Can Enzymatic Stoichiometry Be Used to Determine Growth-Limiting Nutrients for Microorganisms?—A Critical Assessment in Two Subtropical Soils. Soil Biol. Biochem. 2019, 128, 115–126. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F. Towards an Integration of Ecological Stoichiometry and the Metabolic Theory of Ecology to Better Understand Nutrient Cycling. Ecol. Lett. 2009, 12, 369–384. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad Shah, J.J. Ecoenzymatic Stoichiometry of Microbial Organic Nutrient Acquisition in Soil and Sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Zhao, J.; Xiao, K.; Wang, K. Effects of Nitrogen Addition on Activities of Soil Nitrogen Acquisition enzymes: A Meta-Analysis. Agric. Ecosyst. Environ. 2018, 252, 126–131. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Zhao, J.; Zhang, W.; Xiao, K.; Wang, K. Nitrogen Addition Aggravates Microbial Carbon Limitation: Evidence from Ecoenzymatic Stoichiometry. Geoderma 2018, 329, 61–64. [Google Scholar] [CrossRef]
- Dong, C.; Wang, W.; Liu, H.; Xu, X.; Zeng, H. Temperate Grassland Shifted from Nitrogen to Phosphorus Limitation Induced by Degradation and Nitrogen Deposition: Evidence from Soil Extracellular Enzyme Stoichiometry. Ecol. Indic. 2019, 101, 453–464. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector Analysis of Ecoenzyme Activities Reveal Constraints on Coupled C, N and P Dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Mori, T.; Imai, N.; Yokoyama, D.; Kitayama, K. Effects of Nitrogen and Phosphorus Fertilization on the Ratio of Activities of Carbon-Acquiring to Nitrogen-Acquiring Enzymes in a Primary Lowland Tropical Rainforest in Borneo, Malaysia. Soil Sci. Plant Nutr. 2018, 64, 554–557. [Google Scholar] [CrossRef]
- Tatariw, C.; MacRae, J.D.; Fernandez, I.J.; Gruselle, M.-C.; Salvino, C.J.; Simon, K.S. Chronic Nitrogen Enrichment at the Watershed Scale Does Not Enhance Microbial Phosphorus Limitation. Ecosystems 2018, 21, 178–189. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of Soil Microbial Community to Continuous Experimental Nitrogen Additions for 13 Years in a Nitrogen-Rich Tropical Forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, Y.; Gao, D.; Wang, X.; Liu, W.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Ecoenzymatic Stoichiometry and Nutrient Dynamics along a Revegetation Chronosequence in the Soils of Abandoned Land and Robinia pseudoacacia Plantation on the Loess Plateau, China. Soil Biol. Biochem. 2019, 134, 1–14. [Google Scholar] [CrossRef]
- Utobo, E.B.; Tewari, L. Soil Enzymes as Bioindicators of Soil Ecosystem Status. Appl. Ecol. Environ. Res. 2015, 13. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, W.; Dai, X.; Schaeffer, S.; Yang, F.; Radosevich, M.; Xu, L.; Liu, X.; Sun, X. Responses of Absolute and Specific Soil Enzyme Activities to Long Term Additions of Organic and Mineral Fertilizer. Sci. Total Environ. 2015, 536, 59–67. [Google Scholar] [CrossRef] [PubMed]
- De Deyn, G.B.; Kooistra, L. The Role of Soils in Habitat Creation, Maintenance and Restoration. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200170. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.R.; Powers, R.F. Forest Soils. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013; p. B9780124095489051691. ISBN 978-0-12-409548-9. [Google Scholar]
- Hatten, J.; Liles, G. A ‘Healthy’ Balance—The Role of Physical and Chemical Properties in Maintaining Forest Soil Function in a Changing World. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2019; Volume 36, pp. 373–396. ISBN 978-0-444-63998-1. [Google Scholar]
- Polley, H.W.; Aspinwall, M.J.; Collins, H.P.; Gibson, A.E.; Gill, R.A.; Jackson, R.B.; Jin, V.L.; Khasanova, A.R.; Reichmann, L.G.; Fay, P.A. CO2 Enrichment and Soil Type Additively Regulate Grassland Productivity. New Phytol. 2019, 222, 183–192. [Google Scholar] [CrossRef]
- Wang, L.; Hamel, C.; Lu, P.; Wang, J.; Sun, D.; Wang, Y.; Lee, S.-J.; Gan, G.Y. Using Enzyme Activities as an Indicator of Soil Fertility in Grassland—An Academic Dilemma. Front. Plant Sci. 2023, 14, 1175946. [Google Scholar] [CrossRef]
- Kimble, J.M. Cryosols; Springer: Berlin/Heidelberg, Germany, 2004; ISBN 978-3-642-05856-1. [Google Scholar]
- Chen, S.-Y.; Wei, P.-J.; Wu, T.-H.; Wu, Q.-B.; Luo, F.-D. Effect of Permafrost Degradation on Carbon Sequestration of Alpine Ecosystems. Sci. Total Environ. 2023, 899, 165642. [Google Scholar] [CrossRef]
- Alsharif, W.; Saad, M.M.; Hirt, H. Desert Microbes for Boosting Sustainable Agriculture in Extreme Environments. Front. Microbiol. 2020, 11, 1666. [Google Scholar] [CrossRef]
- Cusack, D.F.; Marín-Spiotta, E. Wet Tropical Soils and Global Change. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2019; Volume 36, pp. 131–169. ISBN 978-0-444-63998-1. [Google Scholar]
- Propster, J.R.; Schwartz, E.; Hayer, M.; Miller, S.; Monsaint-Queeney, V.; Koch, B.J.; Morrissey, E.M.; Mack, M.C.; Hungate, B.A. Distinct Growth Responses of Tundra Soil Bacteria to Short-Term and Long-Term Warming. Appl. Environ. Microbiol. 2023, 89, e01543-22. [Google Scholar] [CrossRef]
- Viitamäki, S.; Pessi, I.S.; Virkkala, A.-M.; Niittynen, P.; Kemppinen, J.; Eronen-Rasimus, E.; Luoto, M.; Hultman, J. The Activity and Functions of Soil Microbial Communities in the Finnish Sub-Arctic Vary across Vegetation Types. FEMS Microbiol. Ecol. 2022, 98, fiac079. [Google Scholar] [CrossRef]
- Yang, F.; Huang, J.; He, Q.; Zheng, X.; Zhou, C.; Pan, H.; Huo, W.; Yu, H.; Liu, X.; Meng, L.; et al. Impact of Differences in Soil Temperature on the Desert Carbon Sink. Geoderma 2020, 379, 114636. [Google Scholar] [CrossRef]
- Ma, J.; Tsegaye, E.; Li, M.; Wu, B.; Jiang, X. Biodiversity of Trichoderma from Grassland and Forest Ecosystems in Northern Xinjiang, China. 3 Biotech 2020, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Liu, J.; Wang, J.; Han, W.; Shen, Z.; Muraina, T.O.; Chen, J.; Sun, D. Comparison of Soil Microbial Community between Reseeding Grassland and Natural Grassland in Songnen Meadow. Sci. Rep. 2020, 10, 16884. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Li, J.; Wang, P.; Wang, D.; Han, L.; Gao, X.; Shu, J. Response of Soil Bacteria on Habitat-Specialization and Abundance Gradient to Different Afforestation Types. Sci. Rep. 2023, 13, 18181. [Google Scholar] [CrossRef] [PubMed]
- Garaycochea, S.; Romero, H.; Beyhaut, E.; Neal, A.L.; Altier, N. Soil Structure, Nutrient Status and Water Holding Capacity Shape Uruguayan Grassland Prokaryotic Communities. FEMS Microbiol. Ecol. 2020, 96, fiaa207. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Chabot, C.L.; Liebner, S.; Holm, S.; Snyder, M.W.; Dillon, M.; Dudgeon, S.R.; Douglas, T.A.; Leewis, M.-C.; Walter Anthony, K.M.; et al. Permafrost Microbial Communities and Functional Genes Are Structured by Latitudinal and Soil Geochemical Gradients. ISME J. 2023, 17, 1224–1235. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The Biological Activities of Beta-Glucosidase, Phosphatase and Urease as Soil Quality Indicators: A Review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Sun, J.; Zhu, H.; Yang, X.; Zheng, Y.; Sun, T.; Xu, H.; Meng, J.; Zhang, A. Carboxylesterase and Lipase-Catalyzed Degradation of Phthalate Esters in Soil and Water: Congener Structure Selectivity and Specificity. Environ. Technol. Innov. 2022, 28, 102571. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Zaborowska, M.; Kucharski, J. Microbial Diversity and Enzyme Activity as Indicators of Permethrin-Exposed Soil Health. Molecules 2023, 28, 4756. [Google Scholar] [CrossRef]
- de Almeida, R.F.; Naves, E.R.; da Mota, R.P. Soil Quality: Enzymatic Activity of Soil β-Glucosidase. Glob. J. Agric. Res. Rev. 2015, 3, 146–450. [Google Scholar]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different Approaches to Evaluating Soil Quality Using Biochemical Properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Merino, C.; Godoy, R.; Matus, F. Soil Microorganisms and Enzyme Activity at Different Levels of Organic Matter Stability. J. Soil Sci. Plant Nutr. 2016, 16, 14–30. [Google Scholar] [CrossRef]
- Wang, X.-C.; Lu, Q. Beta-Glucosidase Activity in Paddy Soils of the Taihu Lake Region, China. Pedosphere 2006, 16, 118–124. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought Decreases Soil Enzyme Activity in a Mediterranean Quercus Ilex L. Forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Rietz, D.N.; Haynes, R.J. Effects of Irrigation-Induced Salinity and Sodicity on Soil Microbial Activity. Soil Biol. Biochem. 2003, 35, 845–854. [Google Scholar] [CrossRef]
- Krolicka, M.; Hinz, S.W.A.; Koetsier, M.J.; Eggink, G.; Van Den Broek, L.A.M.; Boeriu, C.G. β-N-Acetylglucosaminidase MthNAG from Myceliophthora thermophila C1, a Thermostable Enzyme for Production of N-Acetylglucosamine from Chitin. Appl. Microbiol. Biotechnol. 2018, 102, 7441–7454. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Meng, H.; Gu, J.-D. Microbial Extracellular Enzymes in Biogeochemical Cycling of Ecosystems. J. Environ. Manag. 2017, 197, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Martínez, V.; Cruz, L.; Sotomayor-Ramírez, D.; Pérez-Alegría, L. Enzyme Activities as Affected by Soil Properties and Land Use in a Tropical Watershed. Appl. Soil Ecol. 2007, 35, 35–45. [Google Scholar] [CrossRef]
- Li, C.; Jiang, S.; Du, C.; Lu, Z.; He, N.; Zhou, Y.; Jiang, S.S.; Zhang, G.G. High-Level Extracellular Expression of a New β-N-Acetylglucosaminidase in Escherichia coli for Producing GlcNAc. Front. Microbiol. 2021, 12, 648373. [Google Scholar] [CrossRef]
- Nadeem, H.; Rashid, M.H.; Siddique, M.H.; Azeem, F.; Muzammil, S.; Javed, M.R.; Ali, M.A.; Rasul, I.; Riaz, M. Microbial Invertases: A Review on Kinetics, Thermodynamics, Physiochemical Properties. Process Biochem. 2015, 50, 1202–1210. [Google Scholar] [CrossRef]
- Osiebe, O.; Adewale, I.O.; Omafuvbe, B.O. Production and Characterization of Intracellular Invertase from Saccharomyces cerevisiae (OL629078.1), Using Cassava-Soybean as a Cost-Effective Substrate. Sci. Rep. 2023, 13, 16295. [Google Scholar] [CrossRef]
- Lincoln, L.; More, S.S. Bacterial Invertases: Occurrence, Production, Biochemical Characterization, and Significance of Transfructosylation. J. Basic Microbiol. 2017, 57, 803–813. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Trasar-Cepeda, C.; Leirós, M.D.C.; Seoane, S.; Gil-Sotres, F. Intra-Annual Variation in Biochemical Properties and the Biochemical Equilibrium of Different Grassland Soils under Contrasting Management and Climate. Biol. Fertil. Soils 2011, 47, 633–645. [Google Scholar] [CrossRef]
- Wang, M.; Markert, B.; Shen, W.; Chen, W.; Peng, C.; Ouyang, Z. Microbial Biomass Carbon and Enzyme Activities of Urban Soils in Beijing. Environ. Sci. Pollut. Res. 2011, 18, 958–967. [Google Scholar] [CrossRef]
- Carroll, R.K.; Veillard, F.; Gagne, D.T.; Lindenmuth, J.M.; Poreba, M.; Drag, M.; Potempa, J.; Shaw, L.N. The Staphylococcus Aureus Leucine Aminopeptidase Is Localized to the Bacterial Cytosol and Demonstrates a Broad Substrate Range That Extends beyond Leucine. bchm 2013, 394, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Fowler, J.H.; Walling, L.L. Leucine Aminopeptidases: Diversity in Structure and Function. Biol. Chem. 2006, 387, 1535–1544. [Google Scholar] [CrossRef]
- Liew, S.M.; Tay, S.T.; Puthucheary, S.D. Enzymatic and Molecular Characterisation of Leucine Aminopeptidase of Burkholderia pseudomallei. BMC Microbiol. 2013, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, J.; Wang, J. Clay Minerals Change the Toxic Effect of Cadmium on the Activities of Leucine Aminopeptidase. Adsorpt. Sci. Technol. 2021, 2021, 1024085. [Google Scholar] [CrossRef]
- Zambelli, B.; Musiani, F.; Benini, S.; Ciurli, S. Chemistry of Ni 2+ in Urease: Sensing, Trafficking, and Catalysis. Acc. Chem. Res. 2011, 44, 520–530. [Google Scholar] [CrossRef]
- Yu, X.; Yang, H.; Wang, H. A Cleaner Biocementation Method of Soil via Microbially Induced Struvite Precipitation: A Experimental and Numerical Analysis. J. Environ. Manag. 2022, 316, 115280. [Google Scholar] [CrossRef]
- Mujah, D.; Shahin, M.A.; Cheng, L. State-of-the-Art Review of Biocementation by Microbially Induced Calcite Precipitation (MICP) for Soil Stabilization. Geomicrobiol. J. 2017, 34, 524–537. [Google Scholar] [CrossRef]
- Imran, M.A.; Nakashima, K.; Evelpidou, N.; Kawasaki, S. Durability Improvement of Biocemented Sand by Fiber-Reinforced MICP for Coastal Erosion Protection. Materials 2022, 15, 2389. [Google Scholar] [CrossRef]
- Zheng, X.; Lu, X.; Zhou, M.; Huang, W.; Zhong, Z.; Wu, X.; Zhao, B. Experimental Study on Mechanical Properties of Root–Soil Composite Reinforced by MICP. Materials 2022, 15, 3586. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, D.; Wang, C.; Wang, Y.; Li, C. Staphylococcus Derived from Desert and Its Solidified Ability on Aeolian Sandy Soil. Adv. Civ. Eng. 2020, 2020, 8877039. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C.-H.; Shin, Y.-J.; So, J.-S. Formations of Calcium Carbonate Minerals by Bacteria and Its Multiple Applications. SpringerPlus 2016, 5, 250. [Google Scholar] [CrossRef]
- Vaskevicius, L.; Malunavicius, V.; Jankunec, M.; Lastauskiene, E.; Talaikis, M.; Mikoliunaite, L.; Maneikis, A.; Gudiukaite, R. Insights in MICP Dynamics in Urease-Positive Staphylococcus Sp. H6 and Sporosarcina pasteurii Bacterium. Environ. Res. 2023, 234, 116588. [Google Scholar] [CrossRef]
- Baddam, R.; Reddy, G.B.; Raczkowski, C.; Cyrus, J.S. Activity of Soil Enzymes in Constructed Wetlands Treated with Swine Wastewater. Ecol. Eng. 2016, 91, 24–30. [Google Scholar] [CrossRef]
- Nim, Y.S.; Wong, K.-B. The Maturation Pathway of Nickel Urease. Inorganics 2019, 7, 85. [Google Scholar] [CrossRef]
- Corstanje, R.; Schulin, R.; Lark, R.M. Scale-dependent Relationships between Soil Organic Carbon and Urease Activity. Eur. J. Soil Sci. 2007, 58, 1087–1095. [Google Scholar] [CrossRef]
- García-Gil, J.C.; Plaza, C.; Soler-Rovira, P.; Polo, A. Long-Term Effects of Municipal Solid Waste Compost Application on Soil Enzyme Activities and Microbial Biomass. Soil Biol. Biochem. 2000, 32, 1907–1913. [Google Scholar] [CrossRef]
- Stone, M.M.; Plante, A.F. Changes in Phosphatase Kinetics with Soil Depth across a Variable Tropical Landscape. Soil Biol. Biochem. 2014, 71, 61–67. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Bünemann, E., Oberson, A., Frossard, E., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 215–243. ISBN 978-3-642-15270-2. [Google Scholar]
- Zhang, J.; Jin, K.; Luo, Y.; Du, L.; Tian, R.; Wang, S.; Shen, Y.; Zhang, J.; Li, N.; Shao, W.; et al. Responses of Soil Enzyme Activity to Long-Term Nitrogen Enrichment and Water Addition in a Typical Steppe. Agronomy 2023, 13, 1920. [Google Scholar] [CrossRef]
- Kai, M.; Takazumi, K.; Adachi, H.; Wasaki, J.; Shinano, T.; Osaki, M. Cloning and Characterization of Four Phosphate Transporter cDNAs in Tobacco. Plant Sci. 2002, 163, 837–846. [Google Scholar] [CrossRef]
- Makoi, J.; Ndakidemi, P. Selected Soil Enzymes: Examples of Their Potential Roles in the Ecosystem. Afr. J. Biotechnol. 2008, 7, 181–191. [Google Scholar]
- Park, Y.; Solhtalab, M.; Thongsomboon, W.; Aristilde, L. Strategies of Organic Phosphorus Recycling by Soil Bacteria: Acquisition, Metabolism, and Regulation. Environ. Microbiol. Rep. 2022, 14, 3–24. [Google Scholar] [CrossRef]
- Dodor, D.E.; Tabatabai, M.A. Effect of Cropping Systems on Phosphatases in Soils. J. Plant Nutr. Soil Sci. 2003, 166, 7–13. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Tabatabai, M.A. Enzyme Activities in a Limed Agricultural Soil. Biol. Fertil. Soils 2000, 31, 85–91. [Google Scholar] [CrossRef]
- Kandeler, F.; Kampichler, C.; Horak, O. Influence of Heavy Metals on the Functional Diversity of Soil Microbial Communities. Biol. Fertil. Soils 1996, 23, 299–306. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Li, D.; Xiao, K.; Wang, K. Controls on Soil Arylsulfatase Activity at a Regional Scale. Eur. J. Soil Biol. 2019, 90, 9–14. [Google Scholar] [CrossRef]
- Kunito, T.; Kurita, H.; Kumori, M.; Sakaguchi, K.; Nishizawa, S.; Fujita, K.; Moro, H.; Sawada, K.; Miyabara, Y.; Toda, H.; et al. Microbial Synthesis of Arylsulfatase Depends on the Soluble and Adsorbed Sulfate Concentration in Soils. Eur. J. Soil Biol. 2022, 111, 103418. [Google Scholar] [CrossRef]
- Chen, G.; Lu, Q.; Bai, J.; Wen, L.; Zhang, G.; Wang, W.; Wang, C.; Liu, Z. Organic Sulfur Mineralization in Surface Soils from Coastal Wetlands with Different Flooding Periods Affected by the Flow-Sediment Regulation in the Yellow River Delta, China. CATENA 2022, 215, 106343. [Google Scholar] [CrossRef]
- Kertesz, M.A. The Role of Soil Microbes in Plant Sulphur Nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef]
- Zaborowska, M.; Kucharski, J.; Wyszkowska, J. Brown Algae and Basalt Meal in Maintaining the Activity of Arylsulfatase of Soil Polluted with Cadmium. Water. Air. Soil Pollut. 2017, 228, 267. [Google Scholar] [CrossRef]
- Ujjainiya, P.; Choudhary, M.; Jatav, H.S.; Tokala, V.Y.; Rajput, V.D.; Minkina, T. Impact of Weed Management Practices on Soil Microflora and Dehydrogenase Enzyme Activity Under Varying Levels of Nitrogen in Winter Season Onion (Allium cepa L.). Bull. Environ. Contam. Toxicol. 2022, 108, 430–436. [Google Scholar] [CrossRef]
- Subhani, A.; Changyong, H.; Zhengmiao, X.; Min, L.; El-ghamry, A.M. Impact of Soil Environment and Agronomic Practices on Microbial/Dehydrogenase Enzyme Activity in Soil. A Review. Pak. J. Biol. Sci. 2001, 4, 333–338. [Google Scholar] [CrossRef]
- Silva, J.C.; Gouveia, E.R. Some Properties of Endoglucanases Produced by Streptomyces Spp. in Medium Based on Sugar Cane Bagasse. Rev. Bras. Tecnol. Agroind. 2008, 2. [Google Scholar] [CrossRef]
- Margesin, R.; Zimmerbauer, A.; Schinner, F. Soil Lipase Activity—A Useful Indicator of Oil Biodegradation. Biotechnol. Tech. 1999, 13, 859–863. [Google Scholar] [CrossRef]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic Mulching in Agriculture. Trading Short-Term Agronomic Benefits for Long-Term Soil Degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari, M.; Drogui, P.; Seyhi, B.; Brar, S.K.; Buelna, G.; Dubé, R. Occurrence, Fate and Effects of Di (2-Ethylhexyl) Phthalate in Wastewater Treatment Plants: A Review. Environ. Pollut. 2014, 194, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, G.; Christie, P.; Zhang, M.; Luo, Y.; Teng, Y. Occurrence and Risk Assessment of Phthalate Esters (PAEs) in Vegetables and Soils of Suburban Plastic Film Greenhouses. Sci. Total Environ. 2015, 523, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Fan, X.; Qiu, Y.-J.; Li, C.-Y.; Xing, S.; Zheng, Y.-T.; Xu, J.-H. Newly Identified Thermostable Esterase from Sulfobacillus acidophilus: Properties and Performance in Phthalate Ester Degradation. Appl. Environ. Microbiol. 2014, 80, 6870–6878. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, A.; Fang, L.; Wang, J.; Liu, W. Levels and Distribution of Dechlorane Plus and Related Compounds in Surficial Sediments of the Qiantang River in Eastern China: The Results of Urbanization and Tide. Sci. Total Environ. 2013, 443, 194–199. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, J.; Zheng, C.; Zhang, X.; Zhang, A.; Qi, H. Phthalate Pollution Driven by the Industrial Plastics Market: A Case Study of the Plastic Market in Yuyao City, China. Environ. Sci. Pollut. Res. 2019, 26, 11224–11233. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xu, W.; Zhang, W.; Guang, C.; Mu, W. Microbial Elimination of Pyrethroids: Specific Strains and Involved Enzymes. Appl. Microbiol. Biotechnol. 2022, 106, 6915–6932. [Google Scholar] [CrossRef]
- Birolli, W.G.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Biodegradation of the Pyrethroid Pesticide Esfenvalerate by Marine-Derived Fungi. Mar. Biotechnol. 2016, 18, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L. Phenol Oxidase, Peroxidase and Organic Matter Dynamics of Soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Sinsabaugh, R.L.; Repert, D.A.; Parkhurst, D.F. Microbial Enzyme Shifts Explain Litter Decay Responses to Simulated Nitrogen Deposition. Ecology 2000, 81, 2359–2365. [Google Scholar] [CrossRef]
- Wu, X.D.; Zhao, L.; Fang, H.B.; Chen, J.; Pang, Q.Q.; Wang, Z.W.; Chen, M.J.; Ding, Y.J. Soil Enzyme Activities in Permafrost Regions of the Western Qinghai-Tibetan Plateau. Soil Sci. Soc. Am. J. 2012, 76, 1280–1289. [Google Scholar] [CrossRef]
- Chabot, M.; Morales, E.; Cummings, J.; Rios, N.; Giatpaiboon, S.; Mogul, R. Simple Kinetics, Assay, and Trends for Soil Microbial Catalases. Anal. Biochem. 2020, 610, 113901. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, J.; Mehandia, S.; Singh, G.; Raina, A.; Arya, S.K. Catalase Enzyme: Application in Bioremediation and Food Industry. Biocatal. Agric. Biotechnol. 2018, 16, 192–199. [Google Scholar] [CrossRef]
- Philippot, L.; Piutti, S.; Martin-Laurent, F.; Hallet, S.; Germon, J.C. Molecular Analysis of the Nitrate-Reducing Community from Unplanted and Maize-Planted Soils. Appl. Environ. Microbiol. 2002, 68, 6121–6128. [Google Scholar] [CrossRef]
- Guan, P.; Yang, J.; Yang, Y.; Wang, W.; Zhang, P.; Wu, D. Land Conversion from Cropland to Grassland Alleviates Climate Warming Effects on Nutrient Limitation: Evidence from Soil Enzymatic Activity and Stoichiometry. Glob. Ecol. Conserv. 2020, 24, e01328. [Google Scholar] [CrossRef]
- Zi, H.B.; Hu, L.; Wang, C.T.; Wang, G.X.; Wu, P.F.; Lerdau, M.; Ade, L.J. Responses of Soil Bacterial Community and Enzyme Activity to Experimental Warming of an Alpine Meadow. Eur. J. Soil Sci. 2018, 69, 429–438. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Zhang, Y.; Li, P.; Zhang, X. Ecoenzymatic Stoichiometry and Microbial Nutrient Limitation in Rhizosphere Soil in the Arid Area of the Northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Chen, H.; Zheng, M.; Mao, Q.; Xiao, K.; Wang, K.; Li, D. Cropland Conversion Changes the Status of Microbial Resource Limitation in Degraded Karst Soil. Geoderma 2019, 352, 197–203. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Zhang, J.; Chen, Y.; Yang, L.; Li, H.; Wang, L. Factors Influencing Soil Enzyme Activity in China’s Forest Ecosystems. Plant Ecol. 2018, 219, 31–44. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil Extracellular Enzyme Activities, Soil Carbon and Nitrogen Storage under Nitrogen Fertilization: A Meta-Analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil Moisture Is the Major Factor Influencing Microbial Community Structure and Enzyme Activities across Seven Biogeoclimatic Zones in Western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; He, N.; Wang, Q.; Wang, S.; Wang, R.; Zhao, N.; Jia, Y.; Wang, C. Soil Enzyme Activity and Stoichiometry in Forest Ecosystems along the North-South Transect in Eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Treseder, K.K. Soil Extracellular Enzyme Activities Correspond with Abiotic Factors More than Fungal Community Composition. Biogeochemistry 2014, 117, 23–37. [Google Scholar] [CrossRef]
- Li, Q.; Dong, Y.; Zhang, Q.; Jia, W.; Cheng, X. Determinants of Soil Carbon- and Nitrogen-Hydrolyzing Enzymes within Different Afforested Lands in Central China. Environ. Sci. Pollut. Res. 2022, 29, 18868–18881. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Taylor, P.; Alvarez-Clare, S.; Bustamante, M.M.C.; Chuyong, G.; Dobrowski, S.Z.; Grierson, P.; Harms, K.E.; Houlton, B.Z.; et al. Relationships among Net Primary Productivity, Nutrients and Climate in Tropical Rain Forest: A Pan-tropical Analysis. Ecol. Lett. 2011, 14, 939–947. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, Phosphorus, or Nitrogen Limit Root Allocation, Tree Growth, or Litter Production in a Lowland Tropical Forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef]
- Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Controls over Leaf Litter Decomposition in Wet Tropical Forests. Ecology 2009, 90, 3333–3341. [Google Scholar] [CrossRef]
- Mori, T.; Lu, X.; Aoyagi, R.; Mo, J. Reconsidering the Phosphorus Limitation of Soil Microbial Activity in Tropical Forests. Funct. Ecol. 2018, 32, 1145–1154. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of Soil Extracellular Enzyme Activity along a Climatic Transect in Temperate Grasslands of Northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Deng, L.; Guo, X.; Han, F.; Ju, W.; Wang, X.; Chen, H.; Tan, W.; Zhang, X. Patterns of Soil Microbial Nutrient Limitations and Their Roles in the Variation of Soil Organic Carbon across a Precipitation Gradient in an Arid and Semi-Arid Region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef]
- Pointing, S.B.; Belnap, J. Microbial Colonization and Controls in Dryland Systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef]
- Zhou, J.; Shen, R. Dictionary of Soil Science; Science Press: Beijing, China, 2013. [Google Scholar]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial Stress-Response Physiology and Its Implications for Ecosystem Function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Tapia-Torres, Y.; Elser, J.J.; Souza, V.; García-Oliva, F. Ecoenzymatic Stoichiometry at the Extremes: How Microbes Cope in an Ultra-Oligotrophic Desert Soil. Soil Biol. Biochem. 2015, 87, 34–42. [Google Scholar] [CrossRef]
- Schaeffer, A.; Amelung, W.; Hollert, H.; Kaestner, M.; Kandeler, E.; Kruse, J.; Miltner, A.; Ottermanns, R.; Pagel, H.; Peth, S.; et al. The Impact of Chemical Pollution on the Resilience of Soils under Multiple Stresses: A Conceptual Framework for Future Research. Sci. Total Environ. 2016, 568, 1076–1085. [Google Scholar] [CrossRef]
- Raiesi, F.; Sadeghi, E. Interactive Effect of Salinity and Cadmium Toxicity on Soil Microbial Properties and Enzyme Activities. Ecotoxicol. Environ. Saf. 2019, 168, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: London, UK; San Diego, CA, USA, 1995; ISBN 978-0-12-513840-6. [Google Scholar]
- Zheng, L.; Zhang, M.; Xiao, R.; Chen, J.; Yu, F. Impact of Salinity and Pb on Enzyme Activities of a Saline Soil from the Yellow River Delta: A Microcosm Study. Phys. Chem. Earth Parts ABC 2017, 97, 77–87. [Google Scholar] [CrossRef]
- Raiesi, F.; Razmkhah, M.; Kiani, S. Salinity Stress Accelerates the Effect of Cadmium Toxicity on Soil N Dynamics and Cycling: Does Joint Effect of These Stresses Matter? Ecotoxicol. Environ. Saf. 2018, 153, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Du, C.; Jian, J.; Hou, L.; Wang, Z.; Wang, Q.; Geng, Z. The Interplay of Labile Organic Carbon, Enzyme Activities and Microbial Communities of Two Forest Soils across Seasons. Sci. Rep. 2021, 11, 5002. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, J.; Chen, G.; Guo, J.; Li, Y. Enzyme Stoichiometry Indicates the Variation of Microbial Nutrient Requirements at Different Soil Depths in Subtropical Forests. PLoS ONE 2020, 15, e0220599. [Google Scholar] [CrossRef] [PubMed]
- Boeddinghaus, R.S.; Nunan, N.; Berner, D.; Marhan, S.; Kandeler, E. Do General Spatial Relationships for Microbial Biomass and Soil Enzyme Activities Exist in Temperate Grassland Soils? Soil Biol. Biochem. 2015, 88, 430–440. [Google Scholar] [CrossRef]
- Chen, H.; Luo, P.; Wen, L.; Yang, L.; Wang, K.; Li, D. Determinants of Soil Extracellular Enzyme Activity in a Karst Region, Southwest China. Eur. J. Soil Biol. 2017, 80, 69–76. [Google Scholar] [CrossRef]
- Xu, H.; Liu, G.; Wu, X.; Smoak, J.M.; Mu, C.; Ma, X.; Zhang, X.; Li, H.; Hu, G. Soil Enzyme Response to Permafrost Collapse in the Northern Qinghai-Tibetan Plateau. Ecol. Indic. 2018, 85, 585–593. [Google Scholar] [CrossRef]
- Stark, S.; Männistö, M.K.; Eskelinen, A. Nutrient Availability and pH Jointly Constrain Microbial Extracellular Enzyme Activities in Nutrient-Poor Tundra Soils. Plant Soil 2014, 383, 373–385. [Google Scholar] [CrossRef]
- Climate Change 2021: The Physical Science Basis; IPCC: Geneva, Switzerland, 2021.
- Sim, J.X.F.; Drigo, B.; Doolette, C.L.; Vasileiadis, S.; Karpouzas, D.G.; Lombi, E. Impact of Twenty Pesticides on Soil Carbon Microbial Functions and Community Composition. Chemosphere 2022, 307, 135820. [Google Scholar] [CrossRef]
- Wieder, W.R.; Bonan, G.B.; Allison, S.D. Global Soil Carbon Projections Are Improved by Modelling Microbial Processes. Nat. Clim. Chang. 2013, 3, 909–912. [Google Scholar] [CrossRef]
- Wieder, W.R.; Grandy, A.S.; Kallenbach, C.M.; Taylor, P.G.; Bonan, G.B. Representing Life in the Earth System with Soil Microbial Functional Traits in the MIMICS Model. Geosci. Model Dev. 2015, 8, 1789–1808. [Google Scholar] [CrossRef]
- Shah, A.M.; Khan, I.M.; Shah, T.I.; Bangroo, S.A.; Kirmani, N.A.; Nazir, S.; Malik, A.R.; Aezum, A.M.; Mir, Y.H.; Hilal, A.; et al. Soil Microbiome: A Treasure Trove for Soil Health Sustainability under Changing Climate. Land 2022, 11, 1887. [Google Scholar] [CrossRef]
- Steinweg, J.M.; Dukes, J.S.; Wallenstein, M.D. Modeling the Effects of Temperature and Moisture on Soil Enzyme Activity: Linking Laboratory Assays to Continuous Field Data. Soil Biol. Biochem. 2012, 55, 85–92. [Google Scholar] [CrossRef]
- Rillig, M.C.; Ryo, M.; Lehmann, A.; Aguilar-Trigueros, C.A.; Buchert, S.; Wulf, A.; Iwasaki, A.; Roy, J.; Yang, G. The Role of Multiple Global Change Factors in Driving Soil Functions and Microbial Biodiversity. Science 2019, 366, 886–890. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Wan, S. Predominant Role of Water in Regulating Soil and Microbial Respiration and Their Responses to Climate Change in a Semiarid Grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A.; Zhang, K.; Li, X.; Tan, H.; Gao, Y.; Li, G. Leaf Nitrogen and Phosphorus of Temperate Desert Plants in Response to Climate and Soil Nutrient Availability. Sci. Rep. 2014, 4, 6932. [Google Scholar] [CrossRef]
- Deslippe, J.R.; Hartmann, M.; Simard, S.W.; Mohn, W.W. Long-Term Warming Alters the Composition of Arctic Soil Microbial Communities. FEMS Microbiol. Ecol. 2012, 82, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of Warming and Drought on Grassland Microbial Communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Chen, Y.; Zhang, J.; Li, H.; Wang, L.; Chen, Q. Short-Term Warming Shifts Microbial Nutrient Limitation without Changing the Bacterial Community Structure in an Alpine Timberline of the Eastern Tibetan Plateau. Geoderma 2020, 360, 113985. [Google Scholar] [CrossRef]
- Feng, C.; Sun, H.; Zhang, Y. The Magnitude and Direction of Priming Were Driven by Soil Moisture and Temperature in a Temperate Forest Soil of China. Pedobiologia 2021, 89, 150769. [Google Scholar] [CrossRef]
- Yoshitake, S.; Tabei, N.; Mizuno, Y.; Yoshida, H.; Sekine, Y.; Tatsumura, M.; Koizumi, H. Soil Microbial Response to Experimental Warming in Cool Temperate Semi-natural Grassland in Japan. Ecol. Res. 2015, 30, 235–245. [Google Scholar] [CrossRef]
- Fang, X.; Zhou, G.; Li, Y.; Liu, S.; Chu, G.; Xu, Z.; Liu, J. Warming Effects on Biomass and Composition of Microbial Communities and Enzyme Activities within Soil Aggregates in Subtropical Forest. Biol. Fertil. Soils 2016, 52, 353–365. [Google Scholar] [CrossRef]
- Stark, S.; Ylänne, H.; Tolvanen, A. Long-Term Warming Alters Soil and Enzymatic N:P Stoichiometry in Subarctic Tundra. Soil Biol. Biochem. 2018, 124, 184–188. [Google Scholar] [CrossRef]
- Zuccarini, P.; Asensio, D.; Ogaya, R.; Sardans, J.; Peñuelas, J. Effects of Seasonal and Decadal Warming on Soil Enzymatic Activity in a P-deficient Mediterranean Shrubland. Glob. Chang. Biol. 2020, 26, 3698–3714. [Google Scholar] [CrossRef]
- Koch, O.; Tscherko, D.; Kandeler, E. Temperature Sensitivity of Microbial Respiration, Nitrogen Mineralization, and Potential Soil Enzyme Activities in Organic Alpine Soils. Glob. Biogeochem. Cycles 2007, 21, 2007GB002983. [Google Scholar] [CrossRef]
- Fraser, F.C.; Hallett, P.D.; Wookey, P.A.; Hartley, I.P.; Hopkins, D.W. How Do Enzymes Catalysing Soil Nitrogen Transformations Respond to Changing Temperatures? Biol. Fertil. Soils 2013, 49, 99–103. [Google Scholar] [CrossRef]
- Allison, S.D. Cheaters, Diffusion and Nutrients Constrain Decomposition by Microbial Enzymes in Spatially Structured Environments. Ecol. Lett. 2005, 8, 626–635. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, C.; Wang, Y.; Xu, Z.; Han, H.; Li, L.; Wan, S. Warming and Increased Precipitation Have Differential Effects on Soil Extracellular Enzyme Activities in a Temperate Grassland. Sci. Total Environ. 2013, 444, 552–558. [Google Scholar] [CrossRef]
- Razavi, B.S.; Blagodatskaya, E.; Kuzyakov, Y. Temperature Selects for Static Soil Enzyme Systems to Maintain High Catalytic Efficiency. Soil Biol. Biochem. 2016, 97, 15–22. [Google Scholar] [CrossRef]
- Yun, J.; Jung, J.Y.; Kwon, M.J.; Seo, J.; Nam, S.; Lee, Y.K.; Kang, H. Temporal Variations Rather than Long-Term Warming Control Extracellular Enzyme Activities and Microbial Community Structures in the High Arctic Soil. Microb. Ecol. 2022, 84, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zang, H.; Freeman, B.; Musarika, S.; Evans, C.D.; Chadwick, D.R.; Jones, D.L. Microbial Utilization of Low Molecular Weight Organic Carbon Substrates in Cultivated Peats in Response to Warming and Soil Degradation. Soil Biol. Biochem. 2019, 139, 107629. [Google Scholar] [CrossRef]
- Keuper, F.; Dorrepaal, E.; Van Bodegom, P.M.; Van Logtestijn, R.; Venhuizen, G.; Van Hal, J.; Aerts, R. Experimentally Increased Nutrient Availability at the Permafrost Thaw Front Selectively Enhances Biomass Production of Deep-rooting Subarctic Peatland Species. Glob. Chang. Biol. 2017, 23, 4257–4266. [Google Scholar] [CrossRef]
- Aerts, R. The Freezer Defrosting: Global Warming and Litter Decomposition Rates in Cold Biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Melle, C.; Wallenstein, M.; Darrouzet-Nardi, A.; Weintraub, M.N. Microbial Activity Is Not Always Limited by Nitrogen in Arctic Tundra Soils. Soil Biol. Biochem. 2015, 90, 52–61. [Google Scholar] [CrossRef]
- Baker, N.R.; Allison, S.D. Extracellular Enzyme Kinetics and Thermodynamics along a Climate Gradient in Southern California. Soil Biol. Biochem. 2017, 114, 82–92. [Google Scholar] [CrossRef]
- Marx, J.-C.; Collins, T.; D’Amico, S.; Feller, G.; Gerday, C. Cold-Adapted Enzymes from Marine Antarctic Microorganisms. Mar. Biotechnol. 2007, 9, 293–304. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford Univ. Press: New York, NY, USA; Oxford, UK, 2002; ISBN 978-0-19-511703-5. [Google Scholar]
- Chang, R.; Liu, S.; Chen, L.; Li, N.; Bing, H.; Wang, T.; Chen, X.; Li, Y.; Wang, G. Soil Organic Carbon Becomes Newer under Warming at a Permafrost Site on the Tibetan Plateau. Soil Biol. Biochem. 2021, 152, 108074. [Google Scholar] [CrossRef]
- Mu, C.; Abbott, B.W.; Norris, A.J.; Mu, M.; Fan, C.; Chen, X.; Jia, L.; Yang, R.; Zhang, T.; Wang, K.; et al. The Status and Stability of Permafrost Carbon on the Tibetan Plateau. Earth-Sci. Rev. 2020, 211, 103433. [Google Scholar] [CrossRef]
- Chen, H.; Zhu, Q.; Peng, C.; Wu, N.; Wang, Y.; Fang, X.; Gao, Y.; Zhu, D.; Yang, G.; Tian, J.; et al. The Impacts of Climate Change and Human Activities on Biogeochemical Cycles on the Q Inghai-T Ibetan P Lateau. Glob. Chang. Biol. 2013, 19, 2940–2955. [Google Scholar] [CrossRef]
- Hicks Pries, C.E.; Schuur, E.A.G.; Natali, S.M.; Crummer, K.G. Old Soil Carbon Losses Increase with Ecosystem Respiration in Experimentally Thawed Tundra. Nat. Clim. Chang. 2016, 6, 214–218. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Bockheim, J.; Canadell, J.G.; Euskirchen, E.; Field, C.B.; Goryachkin, S.V.; Hagemann, S.; Kuhry, P.; Lafleur, P.M.; Lee, H.; et al. Vulnerability of Permafrost Carbon to Climate Change: Implications for the Global Carbon Cycle. BioScience 2008, 58, 701–714. [Google Scholar] [CrossRef]
- Wang, X.; Dong, S.; Gao, Q.; Zhou, H.; Liu, S.; Su, X.; Li, Y. Effects of Short-Term and Long-Term Warming on Soil Nutrients, Microbial Biomass and Enzyme Activities in an Alpine Meadow on the Qinghai-Tibet Plateau of China. Soil Biol. Biochem. 2014, 76, 140–142. [Google Scholar] [CrossRef]
- Allison, S.D.; Treseder, K.K. Warming and Drying Suppress Microbial Activity and Carbon Cycling in Boreal Forest Soils. Glob. Chang. Biol. 2008, 14, 2898–2909. [Google Scholar] [CrossRef]
- Li, Y.; Qing, Y.; Lyu, M.; Chen, S.; Yang, Z.; Lin, C.; Yang, Y. Effects of Artificial Warming on Different Soil Organic Carbon and Nitrogen Pools in a Subtropical Plantation. Soil Biol. Biochem. 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Feng, C.; Ma, Y.; Jin, X.; Wang, Z.; Ma, Y.; Fu, S.; Chen, H.Y.H. Soil Enzyme Activities Increase Following Restoration of Degraded Subtropical Forests. Geoderma 2019, 351, 180–187. [Google Scholar] [CrossRef]
- Bai, W.; Wang, G.; Shang, G.; Xu, L.; Wang, Z. Effects of Experimental Warming on Soil Enzyme Activities in an Alpine Swamp Meadow on the Qinghai-Tibetan Plateau. Pedobiologia 2023, 101, 150910. [Google Scholar] [CrossRef]
- Cao, R.; Yang, W.; Chang, C.; Wang, Z.; Wang, Q.; Li, H.; Tan, B. Differential Seasonal Changes in Soil Enzyme Activity along an Altitudinal Gradient in an Alpine-Gorge Region. Appl. Soil Ecol. 2021, 166, 104078. [Google Scholar] [CrossRef]
- Razavi, B.S.; Liu, S.; Kuzyakov, Y. Hot Experience for Cold-Adapted Microorganisms: Temperature Sensitivity of Soil Enzymes. Soil Biol. Biochem. 2017, 105, 236–243. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Ganjurjav, H.; Wang, X.; Su, X.; Wu, X. Soil Bacterial and Fungal Diversity Differently Correlated with Soil Biochemistry in Alpine Grassland Ecosystems in Response to Environmental Changes. Sci. Rep. 2017, 7, 43077. [Google Scholar] [CrossRef]
- Machuca, Á.; Córdova, C.; Stolpe, N.B.; Barrera, J.A.; Chávez, D.; Almendras, K.; Bonilla, A.M. In Vitro Sensitivity of Forest Soil Enzymes to Temperature Increase in Western Patagonia. J. Soil Sci. Plant Nutr. 2018. [Google Scholar] [CrossRef]
- Zhao, Z.; Wu, Y.; Chen, W.; Sun, W.; Wang, Z.; Liu, G.; Xue, S. Soil Enzyme Kinetics and Thermodynamics in Response to Long-Term Vegetation Succession. Sci. Total Environ. 2023, 882, 163542. [Google Scholar] [CrossRef]
- Song, Y.; Sun, L.; Song, C.; Li, M.; Liu, Z.; Zhu, M.; Chen, S.; Yuan, J.; Gao, J.; Wang, X.; et al. Responses of Soil Microbes and Enzymes to Long-Term Warming Incubation in Different Depths of Permafrost Peatland Soil. Sci. Total Environ. 2023, 900, 165733. [Google Scholar] [CrossRef]
- Pan, F.; Zhang, W.; Liang, Y.; Liu, S.; Wang, K. Increased Associated Effects of Topography and Litter and Soil Nutrients on Soil Enzyme Activities and Microbial Biomass along Vegetation Successions in Karst Ecosystem, Southwestern China. Environ. Sci. Pollut. Res. 2018, 25, 16979–16990. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Priyadarshi, M.; Said, S.; Negi, S. Effect of Wastewater on the Soil and Irrigation Process: A Laboratory Study. J. Geogr. Stud. 2017, 1, 46–55. [Google Scholar] [CrossRef]
- Climate Change 2022: Impacts, Adaptation and Vulnerability; IPCC: Geneva, Switzerland, 2022.
- Kandeler, E.; Tscherko, D.; Bardgett, R.D.; Hobbs, P.J.; Kampichler, C.; Jones, T.H. The Response of Soil Microorganisms and Roots to Elevated CO2 and Temperature in a Terrestrial Model Ecosystem. Plant Soil 1998, 202, 251–262. [Google Scholar] [CrossRef]
- Elzobair, K.A.; Stromberger, M.E.; Ippolito, J.A.; Lentz, R.D. Contrasting Effects of Biochar versus Manure on Soil Microbial Communities and Enzyme Activities in an Aridisol. Chemosphere 2016, 142, 145–152. [Google Scholar] [CrossRef]
- Morgan, J.A.; Follett, R.F.; Allen, L.H.; Del Grosso, S.; Derner, J.D.; Dijkstra, F.; Franzluebbers, A.; Fry, R.; Paustian, K.; Schoeneberger, M.M. Carbon Sequestration in Agricultural Lands of the United States. J. Soil Water Conserv. 2010, 65, 6A–13A. [Google Scholar] [CrossRef]
- Lal, R.; Negassa, W.; Lorenz, K. Carbon Sequestration in Soil. Curr. Opin. Environ. Sustain. 2015, 15, 79–86. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic Enzyme Activities in Agricultural and Forest Soils. Some Implications for Their Use as Indicators of Soil Quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef]
- Finzi, A.C.; Austin, A.T.; Cleland, E.E.; Frey, S.D.; Houlton, B.Z.; Wallenstein, M.D. Responses and Feedbacks of Coupled Biogeochemical Cycles to Climate Change: Examples from Terrestrial Ecosystems. Front. Ecol. Environ. 2011, 9, 61–67. [Google Scholar] [CrossRef]
- Wall, D.H.; Nielsen, U.N.; Six, J. Soil Biodiversity and Human Health. Nature 2015, 528, 69–76. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Hai, X.; Shangguan, Z.; Deng, L. Dynamics of Soil Microbial C:N:P Stoichiometry and Its Driving Mechanisms Following Natural Vegetation Restoration after Farmland Abandonment. Sci. Total Environ. 2019, 693, 133613. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Fang, J.; Ji, C.; Shen, H.; Zheng, C.; Zhu, B. Soil Microbial Carbon and Nutrient Constraints Are Driven More by Climate and Soil Physicochemical Properties than by Nutrient Addition in Forest Ecosystems. Soil Biol. Biochem. 2020, 141, 107657. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, Y.; Duan, C.; Wang, X.; Zhang, X.; Ju, W.; Chen, H.; Yue, S.; Wang, Y.; Li, S.; et al. Ecoenzymatic Stoichiometry Reveals Microbial Phosphorus Limitation Decreases the Nitrogen Cycling Potential of Soils in Semi-Arid Agricultural Ecosystems. Soil Tillage Res. 2020, 197, 104463. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, C.; Wang, Y.; Cheng, H.; An, S.; Chang, S.X. Soil Extracellular Enzyme Stoichiometry Reflects the Shift from P- to N-Limitation of Microorganisms with Grassland Restoration. Soil Biol. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Luo, L.; Gu, J.-D. Alteration of Extracellular Enzyme Activity and Microbial Abundance by Biochar Addition: Implication for Carbon Sequestration in Subtropical Mangrove Sediment. J. Environ. Manag. 2016, 182, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.W.W.; Liao, J.X.; Lau, S.Y.; So, P.S.; Hau, B.C.H.; Peprah-manu, D. Coupled Effects of Elevated CO2 and Biochar on Microbial Communities of Vegetated Soil. J. Environ. Manag. 2023, 342, 118136. [Google Scholar] [CrossRef]
- Feng, J.; Yu, D.; Sinsabaugh, R.L.; Moorhead, D.L.; Andersen, M.N.; Smith, P.; Song, Y.; Li, X.; Huang, Q.; Liu, Y.; et al. Trade-offs in Carbon-degrading Enzyme Activities Limit Long-term Soil Carbon Sequestration with Biochar Addition. Biol. Rev. 2023, 98, 1184–1199. [Google Scholar] [CrossRef]
- Babin, D.; Leoni, C.; Neal, A.L.; Sessitsch, A.; Smalla, K. Editorial to the Thematic Topic “Towards a More Sustainable Agriculture through Managing Soil Microbiomes. ” FEMS Microbiol. Ecol. 2021, 97, fiab094. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural Sustainability and Intensive Production Practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Wittwer, R.A.; Bender, S.F.; Hartman, K.; Hydbom, S.; Lima, R.A.A.; Loaiza, V.; Nemecek, T.; Oehl, F.; Olsson, P.A.; Petchey, O.; et al. Organic and Conservation Agriculture Promote Ecosystem Multifunctionality. Sci. Adv. 2021, 7, eabg6995. [Google Scholar] [CrossRef]
- Liu, Z.; Rong, Q.; Zhou, W.; Liang, G. Effects of Inorganic and Organic Amendment on Soil Chemical Properties, Enzyme Activities, Microbial Community and Soil Quality in Yellow Clayey Soil. PLoS ONE 2017, 12, e0172767. [Google Scholar] [CrossRef] [PubMed]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of Forest Soil Respiration in Response to Nitrogen Deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Liu, L.; Greaver, T.L. A Global Perspective on Belowground Carbon Dynamics under Nitrogen Enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef]
- Yayi, N.; Yulong, D.; Yuqiang, L.; Xuyang, W.; Yun, C.; Lilong, W. Soil Microbial Community Responses to Short-Term Nitrogen Addition in China’s Horqin Sandy Land. PLoS ONE 2021, 16, e0242643. [Google Scholar] [CrossRef]
- Ma, Y.; Zhu, B.; Sun, Z.; Zhao, C.; Yang, Y.; Piao, S. The Effects of Simulated Nitrogen Deposition on Extracellular Enzyme Activities of Litter and Soil among Different-Aged Stands of Larch. J. Plant Ecol. 2014, 7, 240–249. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Van Groenigen, K.J.; Hungate, B.A.; Cao, J.; Zhou, X.; Wang, R. A Keystone Microbial Enzyme for Nitrogen Control of Soil Carbon Storage. Sci. Adv. 2018, 4, eaaq1689. [Google Scholar] [CrossRef] [PubMed]
- Keeler, B.L.; Hobbie, S.E.; Kellogg, L.E. Effects of Long-Term Nitrogen Addition on Microbial Enzyme Activity in Eight Forested and Grassland Sites: Implications for Litter and Soil Organic Matter Decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Li, J.; Zhou, X.; Cao, J.; Wang, R.; Wang, Y.; Shelton, S.; Jin, Z.; Walker, L.M.; et al. Costimulation of Soil Glycosidase Activity and Soil Respiration by Nitrogen Addition. Glob. Chang. Biol. 2017, 23, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Lajtha, K.; Crawley, M.J. Chronic Nitrogen Fertilization and Carbon Sequestration in Grassland Soils: Evidence of a Microbial Enzyme Link. Biogeochemistry 2015, 126, 301–313. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between Roots and Microorganisms for Nitrogen: Mechanisms and Ecological Relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Augusto, L.; Achat, D.L.; Jonard, M.; Vidal, D.; Ringeval, B. Soil Parent Material—A Major Driver of Plant Nutrient Limitations in Terrestrial Ecosystems. Glob. Chang. Biol. 2017, 23, 3808–3824. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Chen, X.; Jing, X.; Zhu, B. A Meta-Analysis of Soil Extracellular Enzyme Activities in Response to Global Change. Soil Biol. Biochem. 2018, 123, 21–32. [Google Scholar] [CrossRef]
- Ma, W.; Li, J.; Gao, Y.; Xing, F.; Sun, S.; Zhang, T.; Zhu, X.; Chen, C.; Li, Z. Responses of Soil Extracellular Enzyme Activities and Microbial Community Properties to Interaction between Nitrogen Addition and Increased Precipitation in a Semi-Arid Grassland Ecosystem. Sci. Total Environ. 2020, 703, 134691. [Google Scholar] [CrossRef]
- Gong, S.; Zhang, T.; Guo, R.; Cao, H.; Shi, L.; Guo, J.; Sun, W. Response of Soil Enzyme Activity to Warming and Nitrogen Addition in a Meadow Steppe. Soil Res. 2015, 53, 242. [Google Scholar] [CrossRef]
- Hewins, D.B.; Broadbent, T.; Carlyle, C.N.; Bork, E.W. Extracellular Enzyme Activity Response to Defoliation and Water Addition in Two Ecosites of the Mixed Grass Prairie. Agric. Ecosyst. Environ. 2016, 230, 79–86. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil Enzymology: Classical and Molecular Approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Tang, M.; Ding, Z.; Jiang, L.; Li, P.; Ma, S.; Tian, D.; Xu, L.; Zhu, J.; et al. Nitrogen Deposition Has Minor Effect on Soil Extracellular Enzyme Activities in Six Chinese Forests. Sci. Total Environ. 2017, 607–608, 806–815. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, M.; Jin, G.; Sui, X.; Zhang, T.; Song, F. Effects of Nitrogen Deposition on Nitrogen-Mineralizing Enzyme Activity and Soil Microbial Community Structure in a Korean Pine Plantation. Microb. Ecol. 2021, 81, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Liangjie, S.; Yunshe, D.; Yuchun, Q.; Yating, H.; Qin, P.; Xinchao, L.; Junqiang, J.; Shufang, G.; Congcong, C. Intra- and Inter-Annual Variation of Soil Microbial and Enzymatic Response to Water and Nitrogen Addition in a Chinese Semi-Arid Steppe. J. Pure Appl. Microbiol. 2014, 8, 1339–1351. [Google Scholar]
- Chen, X.; Hao, B.; Jing, X.; He, J.-S.; Ma, W.; Zhu, B. Minor Responses of Soil Microbial Biomass, Community Structure and Enzyme Activities to Nitrogen and Phosphorus Addition in Three Grassland Ecosystems. Plant Soil 2019, 444, 21–37. [Google Scholar] [CrossRef]
- Kátai, J.; Zsuposné, Á.O.; Tállai, M.; Alshaal, T. Would Fertilization History Render the Soil Microbial Communities and Their Activities More Resistant to Rainfall Fluctuations? Ecotoxicol. Environ. Saf. 2020, 201, 110803. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S. Nitrogen Addition Shapes Soil Enzyme Activity Patterns by Changing pH Rather than the Composition of the Plant and Microbial Communities in an Alpine Meadow Soil. Plant Soil 2019, 440, 11–24. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, Y.; Shi, Y.; He, N.; Wen, X.; Yu, Q.; Zheng, C.; Sun, X.; Qiu, W. Responses of Soil Hydrolytic Enzymes, Ammonia-Oxidizing Bacteria and Archaea to Nitrogen Applications in a Temperate Grassland in Inner Mongolia. Sci. Rep. 2016, 6, 32791. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mori, T.; Mao, Q.; Zhou, K.; Wang, Z.; Zhang, Y.; Mo, H.; Lu, X.; Mo, J. Long-Term Phosphorus Addition Downregulates Microbial Investments on Enzyme Productions in a Mature Tropical Forest. J. Soils Sediments 2020, 20, 921–930. [Google Scholar] [CrossRef]
- DeForest, J.L.; Moorhead, D.L. Effects of Elevated pH and Phosphorus Fertilizer on Soil C, N and P Enzyme Stoichiometry in an Acidic Mixed Mesophytic Deciduous Forest. Soil Biol. Biochem. 2020, 150, 107996. [Google Scholar] [CrossRef]
- Feng, L.; Cao, B.; Wang, X. Response of Soil Extracellular Enzyme Activity and Stoichiometry to Short-Term Warming and Phosphorus Addition in Desert Steppe. PeerJ 2023, 11, e16227. [Google Scholar] [CrossRef] [PubMed]
- Touhami, D.; Condron, L.M.; McDowell, R.W.; Moss, R. Effects of Long-term Phosphorus Fertilizer Inputs and Seasonal Conditions on Organic Soil Phosphorus Cycling under Grazed Pasture. Soil Use Manag. 2023, 39, 385–401. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, H.; Zhao, H.; Xia, H.; Sun, M.; Li, Z.; Li, P.; Zheng, C.; Dong, H.; Liu, J. Phosphorus Affects Enzymatic Activity and Chemical Properties of Cotton Soil. Plant Soil Environ. 2019, 65, 361–368. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Zeeshan, M.; Farooq, S.; Ali, I.; Khan, A.; Zhou, X.B. Irrigation and Nitrogen Fertilization Alter Soil Bacterial Communities, Soil Enzyme Activities, and Nutrient Availability in Maize Crop. Front. Microbiol. 2022, 13, 833758. [Google Scholar] [CrossRef]
- Shi, B.; Zhang, J.; Wang, C.; Ma, J.; Sun, W. Responses of Hydrolytic Enzyme Activities in Saline-Alkaline Soil to Mixed Inorganic and Organic Nitrogen Addition. Sci. Rep. 2018, 8, 4543. [Google Scholar] [CrossRef]
- Aziz, M.A.; Hazra, F.; Salma, S.; Nursyamsi, D. Soil Enzyme Activities and Their Relationship to Total Soil Bacteria, Soil Microbial Biomass and Soil Chemical Characteristics of Organic and Conventional Farming. J. Trop. Soils 2018, 23, 133–141. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic Farming Enhances Soil Microbial Abundance and Activity—A Meta-Analysis and Meta-Regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef] [PubMed]
- Durrer, A.; Gumiere, T.; Rumenos Guidetti Zagatto, M.; Petry Feiler, H.; Miranda Silva, A.M.; Henriques Longaresi, R.; Homma, S.K.; Cardoso, E.J.B.N. Organic Farming Practices Change the Soil Bacteria Community, Improving Soil Quality and Maize Crop Yields. PeerJ 2021, 9, e11985. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, H.; Wu, J.; Zhang, S.; Gao, C.; Zhang, S.; Tang, D.W.S. Soil Enzyme Activities, Soil Physical Properties, Photosynthetic Physical Characteristics and Water Use of Winter Wheat after Long-Term Straw Mulch and Organic Fertilizer Application. Front. Plant Sci. 2023, 14, 1186376. [Google Scholar] [CrossRef]
- Sacco, D.; Moretti, B.; Monaco, S.; Grignani, C. Six-Year Transition from Conventional to Organic Farming: Effects on Crop Production and Soil Quality. Eur. J. Agron. 2015, 69, 10–20. [Google Scholar] [CrossRef]
- Chen, X.; Henriksen, T.M.; Svensson, K.; Korsaeth, A. Long-Term Effects of Agricultural Production Systems on Structure and Function of the Soil Microbial Community. Appl. Soil Ecol. 2020, 147, 103387. [Google Scholar] [CrossRef]
- Chang, E.-H.; Chung, R.-S.; Tsai, Y.-H. Effect of Different Application Rates of Organic Fertilizer on Soil Enzyme Activity and Microbial Population. Soil Sci. Plant Nutr. 2007, 53, 132–140. [Google Scholar] [CrossRef]
- Ghosh, A.; Singh, A.B.; Kumar, R.V.; Manna, M.C.; Bhattacharyya, R.; Rahman, M.M.; Sharma, P.; Rajput, P.S.; Misra, S. Soil Enzymes and Microbial Elemental Stoichiometry as Bio-Indicators of Soil Quality in Diverse Cropping Systems and Nutrient Management Practices of Indian Vertisols. Appl. Soil Ecol. 2020, 145, 103304. [Google Scholar] [CrossRef]
- Ouyang, Y.; Reeve, J.R.; Norton, J.M. Soil Enzyme Activities and Abundance of Microbial Functional Genes Involved in Nitrogen Transformations in an Organic Farming System. Biol. Fertil. Soils 2018, 54, 437–450. [Google Scholar] [CrossRef]
- Lal, B.; Sharma, S.C.; Meena, R.L.; Sarkar, S.; Sahoo, A.; Balai, R.C.; Gautam, P.; Meena, B.P. Utilization of Byproducts of Sheep Farming as Organic Fertilizer for Improving Soil Health and Productivity of Barley Forage. J. Environ. Manag. 2020, 269, 110765. [Google Scholar] [CrossRef]
- Yang, L.; Muhammad, I.; Chi, Y.X.; Liu, Y.X.; Wang, G.Y.; Wang, Y.; Zhou, X.B. Straw Return and Nitrogen Fertilization Regulate Soil Greenhouse Gas Emissions and Global Warming Potential in Dual Maize Cropping System. Sci. Total Environ. 2022, 853, 158370. [Google Scholar] [CrossRef]
- Elser, J.J. Phosphorus: A Limiting Nutrient for Humanity? Curr. Opin. Biotechnol. 2012, 23, 833–838. [Google Scholar] [CrossRef]
- Hou, E.; Luo, Y.; Kuang, Y.; Chen, C.; Lu, X.; Jiang, L.; Luo, X.; Wen, D. Global Meta-Analysis Shows Pervasive Phosphorus Limitation of Aboveground Plant Production in Natural Terrestrial Ecosystems. Nat. Commun. 2020, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Baumann, F.; Song, C.; Zhang, M.; Shi, Y.; Kühn, P.; Scholten, T.; He, J.-S. Increasing Temperature Reduces the Coupling between Available Nitrogen and Phosphorus in Soils of Chinese Grasslands. Sci. Rep. 2017, 7, 43524. [Google Scholar] [CrossRef] [PubMed]
- Hou, E.; Chen, C.; Luo, Y.; Zhou, G.; Kuang, Y.; Zhang, Y.; Heenan, M.; Lu, X.; Wen, D. Effects of Climate on Soil Phosphorus Cycle and Availability in Natural Terrestrial Ecosystems. Glob. Chang. Biol. 2018, 24, 3344–3356. [Google Scholar] [CrossRef] [PubMed]
- Achat, D.L.; Pousse, N.; Nicolas, M.; Brédoire, F.; Augusto, L. Soil Properties Controlling Inorganic Phosphorus Availability: General Results from a National Forest Network and a Global Compilation of the Literature. Biogeochemistry 2016, 127, 255–272. [Google Scholar] [CrossRef]
- Lu, J.; Jia, P.; Feng, S.; Wang, Y.; Zheng, J.; Ou, S.; Wu, Z.; Liao, B.; Shu, W.; Liang, J.; et al. Remarkable Effects of Microbial Factors on Soil Phosphorus Bioavailability: A Country-scale Study. Glob. Chang. Biol. 2022, 28, 4459–4471. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, K.; Mori, T.; Mo, J.; Zhang, W. Effects of Phosphorus and Nitrogen Fertilization on Soil Arylsulfatase Activity and Sulfur Availability of Two Tropical Plantations in Southern China. For. Ecol. Manag. 2019, 453, 117613. [Google Scholar] [CrossRef]
- Prashar, P.; Shah, S. Impact of Fertilizers and Pesticides on Soil Microflora in Agriculture. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Sustainable Agriculture Reviews; Springer International Publishing: Cham, Switzerland, 2016; Volume 19, pp. 331–361. ISBN 978-3-319-26776-0. [Google Scholar]
- Gundi, V.A.K.B.; Viswanath, B.; Chandra, M.S.; Kumar, V.N.; Reddy, B.R. Activities of Cellulase and Amylase in Soils as Influenced by Insecticide Interactions. Ecotoxicol. Environ. Saf. 2007, 68, 278–285. [Google Scholar] [CrossRef]
- Yang, F.; Yang, S.; Xu, J.; Wang, Y.; Gao, M.; Zhang, M.; Dai, W.; Fu, H.; Zeng, W.; Liu, C. Dynamic Response of Soil Enzymes and Microbial Diversity to Continuous Application of Atrazine in Black Soil of a Cornfield without Rotation in Northeast China. Diversity 2021, 13, 259. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, C.; Rong, Q.; Li, C.; Mao, J.; Liu, Y.; Chen, J.; Liu, X. Effect of Two Organic Amendments on Atrazine Degradation and Microorganisms in Soil. Appl. Soil Ecol. 2020, 152, 103564. [Google Scholar] [CrossRef]
- Imfeld, G.; Vuilleumier, S. Measuring the Effects of Pesticides on Bacterial Communities in Soil: A Critical Review. Eur. J. Soil Biol. 2012, 49, 22–30. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Huang, B.; Song, Z.; Ren, L.; Hao, B.; Liu, J.; Zhu, J.; Fang, W.; Yan, D.; et al. Organic Fertilizer Improves Soil Fertility and Restores the Bacterial Community after 1,3-Dichloropropene Fumigation. Sci. Total Environ. 2020, 738, 140345. [Google Scholar] [CrossRef]
- Du, Z.; Zhu, Y.; Zhu, L.; Zhang, J.; Li, B.; Wang, J.; Wang, J.; Zhang, C.; Cheng, C. Effects of the Herbicide Mesotrione on Soil Enzyme Activity and Microbial Communities. Ecotoxicol. Environ. Saf. 2018, 164, 571–578. [Google Scholar] [CrossRef]
- Pose-Juan, E.; Sánchez-Martín, M.J.; Herrero-Hernández, E.; Rodríguez-Cruz, M.S. Application of Mesotrione at Different Doses in an Amended Soil: Dissipation and Effect on the Soil Microbial Biomass and Activity. Sci. Total Environ. 2015, 536, 31–38. [Google Scholar] [CrossRef]
- Bamaga, O.A.A.; Mahdy, M.A.K.; Lim, Y.A.L. Frequencies Distribution of Dihydrofolate Reductase and Dihydropteroate Synthetase Mutant Alleles Associated with Sulfadoxine–Pyrimethamine Resistance in Plasmodium Falciparum Population from Hadhramout Governorate, Yemen. Malar. J. 2015, 14, 516. [Google Scholar] [CrossRef]
- Floch, C.; Chevremont, A.-C.; Joanico, K.; Capowiez, Y.; Criquet, S. Indicators of Pesticide Contamination: Soil Enzyme Compared to Functional Diversity of Bacterial Communities via Biolog® Ecoplates. Eur. J. Soil Biol. 2011, 47, 256–263. [Google Scholar] [CrossRef]
- Liu, R.; Liang, J.; Yang, Y.; Jiang, H.; Tian, X. Effect of Polylactic Acid Microplastics on Soil Properties, Soil Microbials and Plant Growth. Chemosphere 2023, 329, 138504. [Google Scholar] [CrossRef]
- Janczak, K.; Hrynkiewicz, K.; Znajewska, Z.; Dąbrowska, G. Use of Rhizosphere Microorganisms in the Biodegradation of PLA and PET Polymers in Compost Soil. Int. Biodeterior. Biodegrad. 2018, 130, 65–75. [Google Scholar] [CrossRef]
- Liu, H.; Yang, X.; Liu, G.; Liang, C.; Xue, S.; Chen, H.; Ritsema, C.J.; Geissen, V. Response of Soil Dissolved Organic Matter to Microplastic Addition in Chinese Loess Soil. Chemosphere 2017, 185, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Huang, S.; Zhang, H.; Tong, Y.; Wen, D.; Xia, X.; Wang, H.; Luo, Y.; Barceló, D. Response of Soil Enzyme Activities and Bacterial Communities to the Accumulation of Microplastics in an Acid Cropped Soil. Sci. Total Environ. 2020, 707, 135634. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Feng, X.; Liu, Y.; Cui, W.; Sun, Y.; Zhang, S.; Wang, F. Effects of Microplastics and Carbon Nanotubes on Soil Geochemical Properties and Bacterial Communities. J. Hazard. Mater. 2022, 433, 128826. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Q.; Sun, Y.; Zhang, S.; Wang, F. Microplastics Change Soil Properties, Heavy Metal Availability and Bacterial Community in a Pb-Zn-Contaminated Soil. J. Hazard. Mater. 2022, 424, 127364. [Google Scholar] [CrossRef]
- Cardoso, E.J.B.N.; Vasconcellos, R.L.F.; Bini, D.; Miyauchi, M.Y.H.; Santos, C.A.D.; Alves, P.R.L.; Paula, A.M.D.; Nakatani, A.S.; Pereira, J.D.M.; Nogueira, M.A. Soil Health: Looking for Suitable Indicators. What Should Be Considered to Assess the Effects of Use and Management on Soil Health? Sci. Agric. 2013, 70, 274–289. [Google Scholar] [CrossRef]
- Squires, V.R. The Role of Food, Agriculture, Forestry and Fisheries in Human Nutrition; EOLSS Publications: Abu Dhabi, United Arab Emirates, 2011; Volume 3, ISBN 1-84826-136-5. [Google Scholar]
- Lazicki, P.; Mazza Rodrigues, J.L.; Geisseler, D. Sensitivity and Variability of Soil Health Indicators in a California Cropping System. Soil Sci. Soc. Am. J. 2021, 85, 1827–1842. [Google Scholar] [CrossRef]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil Aggregate Microbial Communities: Towards Understanding Microbiome Interactions at Biologically Relevant Scales. Appl. Environ. Microbiol. 2019, 85, e00324-19. [Google Scholar] [CrossRef]
- Gyawali, A.J.; Neely, H.; Foster, J.; Neely, C.; Lewis, K.; Pintar, J.; Bekewe, P.; Smith, A.P. Sampling for Biological Indicators of Soil Health: How Does Sampling Methodology Affect Research Results? Geoderma 2023, 435, 116513. [Google Scholar] [CrossRef]
- Smith, G.R.; Crowther, T.W.; Eisenhauer, N.; van den Hoogen, J. Building a Global Database of Soil Microbial Biomass and Function: A Call for Collaboration. Soil Org. 2019, 91, 139. [Google Scholar] [CrossRef]
- Yang, S.; Zheng, Q.; Yuan, M.; Shi, Z.; Chiariello, N.R.; Docherty, K.M.; Dong, S.; Field, C.B.; Gu, Y.; Gutknecht, J.; et al. Long-Term Elevated CO2 Shifts Composition of Soil Microbial Communities in a Californian Annual Grassland, Reducing Growth and N Utilization Potentials. Sci. Total Environ. 2019, 652, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Igalavithana, A.D.; Shaheen, S.M.; Park, J.N.; Lee, S.S.; Ok, Y.S. Potentially Toxic Element Contamination and Its Impact on Soil Biological Quality in Urban Agriculture: A Critical Review. In Heavy Metal Contamination of Soils; Sherameti, I., Varma, A., Eds.; Soil Biology; Springer International Publishing: Cham, Switzerland, 2015; Volume 44, pp. 81–101. ISBN 978-3-319-14525-9. [Google Scholar]
- Wei, B.; Yang, L. A Review of Heavy Metal Contaminations in Urban Soils, Urban Road Dusts and Agricultural Soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Rebello, S.; Nathan, V.K.; Sindhu, R.; Binod, P.; Awasthi, M.K.; Pandey, A. Bioengineered Microbes for Soil Health Restoration: Present Status and Future. Bioengineered 2021, 12, 12839–12853. [Google Scholar] [CrossRef]
- Yue, K.; Fornara, D.A.; Yang, W.; Peng, Y.; Li, Z.; Wu, F.; Peng, C. Effects of Three Global Change Drivers on Terrestrial C:N:P Stoichiometry: A Global Synthesis. Glob. Chang. Biol. 2017, 23, 2450–2463. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Soil Ecosystem/Location | Enzyme Activities and Important Remarks | Ref. |
|---|---|---|
| China’s forest ecosystems | The activity of CAT, PO, AcP, AlP, and proteases varied significantly across forest types. In particular, primosols, cambisols, and argosols have higher CAT and Ure activity than ferrosols. Enzyme activities decreased with increasing soil depth but increased with SOM. Both PO and Ure had a negative connection with MAT, whereas CAT, INV, and protease activities showed a complex pattern: they reduced at temperatures below 2.5 °C, increased between 2.5 °C and 17.5 °C, and then fell again at temperatures over 17.5 °C. Protease activity was somewhat positively correlated with MAP, but CAT, PO, and Ure activities were negatively correlated. The activity of CAT, INV, AcP, AlP, Ure, and proteases increased and then decreased with altitude. | [127] |
| Two afforested lands (coniferous woodland and leguminous shrubland), Wulongchi Research Station, Hubei Province, China | The C:N ratio of enzymes in afforested areas was much greater than in open areas. This ratio was found to be lower in forests compared to shrublands. | [132] |
| Two forests, Betula albosinensis (Ba) and Picea asperata Mast. (Pa); Qinling Mountains, China | The average seasonal enzyme activities of BG and BX in Ba forest soils were 30.0% and 32.3% greater, respectively, than those in Pa soils, whereas CBH activity was 19.7% lower. Local organic C in the soil had a substantial positive connection with CBH, BG, and BX enzyme activity. Pa soil had a lower SOC content and lower BG and BX enzyme activity than Ba soil. This was largely owing to differences in litterfall and root exudates between Pa and Ba. During the summer and autumn seasons, CBH, BG, and BX enzyme activity increased in both Pa and Ba forest soils. | [148] |
| Different soil depths in subtropical forests; soil layers (0–10, 10–20, 20–40, 40–60 cm) in a natural secondary evergreen broad-leaved forest and a Chinese fir (Cunninghamia lanceolata) plantation forest in subtropical China | Microbial C and P limitation changed with soil depth, with microorganisms in soil below 20 cm in both forests requiring greater N. The activity of C-acquiring (BX + CBH + BG), N-acquiring (NAG + LAP), and P-acquiring enzymes decreased as soil depth increased. However, the regularity of enzyme activity across the soil profile indicates an imbalance in microbial nutrient demand at different soil depths. | [149] |
| The 18 independent grassland sites differing in their land-use intensity in two geographic regions: the Hainich National Park in the middle of Germany and the Swabian Alb in south-west Germany | Enzyme activities related to C-acquiring and N-acquiring (BG, BX, and chitinase), as well as organic C, total N, extractable organic C, and mineral N, were found to be higher in the Swabian Alb (Leptosols) than in the Hainich National Park (mostly Stagnosols). Bulk density was found to be negatively correlated with microbial biomass, Ure activity, organic C, and total N. The activities of BG, chitinase, BX, AP, and Ure were impacted by local abiotic soil characteristics but showed little geographical association. | [150] |
| Regional-scale karst area, southwest China; secondary forest, shrubland, grassland and cropland underlain by either dolomite or limestone | The activity patterns of extracellular enzymes involved in C, N, and P cycling varied significantly between dolomite and limestone, as well as across the four types of land use. These variations in enzyme activity were impacted by changes in land use. | [151] |
| The collapsing, collapsed, and an unaffected site of a thermokarst feature on the Northern Qinghai–Tibetan Plateau | In the top 0–20 cm layer, collapsing soils had significantly lower INV activity than control and collapsed soils. At a soil depth of 0–10 cm, collapsing soils had the highest CAT activities and the lowest Ure activities among the three circumstances. Light fraction C content, C:N ratios, and moisture content emerged as important indicators of enzyme activity. Among the six enzyme activities measured, four showed significant differences in the upper 10 cm of soil. | [152] |
| Permafrost regions of the middle and western Qinghai–Tibetan Plateau; cold, arid steppe, with an active layer thicker than 2 m | The activities of enzymes such as INV, CAT, amylase, cellulase, Ure, and AlAP were measured. Soil enzyme activity was observed to be higher in Stipa roborowskyi Roshev vegetation communities than in Carex moorcroftii Falconer ex Boott communities. The alpine cold desert had the lowest soil enzyme activity. | [119] |
| Tundra soils, which contain low concentrations of soil nutrients, low pH, store a large proportion of the global soil C pool | The potential activity of BG was discovered to rise with increasing nutritional levels. In contrast, as soil pH increased, BG activity decreased. When nutritional restrictions were corrected through fertilization, microbial biomass and enzymatic capacity for cellulose decomposition increased, presumably improving SOM decomposition. However, increasing soil pH was found to reduce the enzymatic capacity for cellulose degradation, presumably due to changes in the bioavailability of organic substrates. | [153] |
| Soil Ecosystem/Location/Climate Zone | Effect on Soil Enzyme Activity and Soil | Ref. |
|---|---|---|
| Alpine | ||
| Alpine meadow, northwestern Sichuan, China | At a soil depth of 0–10 cm, experimental warming enhanced AcP, INV, and Ure activities, as well as accessible nutrients, while lowering CAT activity and SOM levels. Warming at 10–20 cm deep enhanced CAT activity, SOM, accessible N, and K+ while decreasing INV activity. | [124] |
| Alpine swamp meadow, Qinghai–Tibetan Plateau, alpine grasslands, permafrost regions, a 3-year experiment with two warming levels (2.7 °C and 5.3 °C) | NO3−N and SM were critical in explaining large differences in soil enzyme activity. Warming increased INV and amylase activity throughout the growing season while decreasing Ure activity, but had no significant influence on CAT or cellulase activity. | [193] |
| 3-year in situ soil core incubation experiment, a 2431-m altitudinal gradient in an alpine-gorge region, the eastern Qinghai–Tibet Plateau | Between 2013 and 2017, subalpine coniferous forests and alpine meadows had higher INV, Ure, and AcP activities than dry valley shrubland and valley-mountain ecotone forests. EEA’s sensitivity to seasons reduced with altitude. | [194] |
| Old-adapted alpine grassland of the Tibetan Plateau | Enzymes that degrade low-quality polymers remained temperature sensitive above 25 °C. Several enzymes’ substrate affinity rose up to 20 °C, but their Km increased rapidly at 25 °C, lowering catalytic effectiveness. | [195] |
| Alpine meadow, alpine steppe and cultivated grassland, Qinghai–Tibetan plateau; 3-year warming, enhanced precipitation and yak overgrazing | Despite various treatments being applied, the activity levels of sucrose and AP remained consistent. In contrast, overgrazing in cultivated grasslands led to an increase in Ure activity and microbial biomass (N). | [196] |
| Grasslands | ||
| Temperate grasslands of northern China during the growing season of 2013 | Temperate grasslands had lower enzyme C:N and C:P ratios than tropical soils. The enzyme ratios changed with soil depth, and log-transformed enzyme ratios differed from global ratios, indicating a greater investment in N-acquiring enzymes in temperate grasslands. | [137] |
| Temperate grassland of northern China at two depths of 0–10 and 10–20 cm | Due to warming AcP activity increased at 0–10 cm depth, as did NAG at 10–20 cm depth, whereas BG and AcP activity declined in the subsurface. Increased precipitation boosted NAG, LAP, and AlP activity in both surface and deep soils. | [174] |
| Forest | ||
| Forest soils from the fragile cold ecosystems, Western Patagonia, Chile | NAG activity, like microbiological activity, was more temperature sensitive than BG. Soil total nutrients had a greater influence on enzyme Kcat than accessible nutrients throughout vegetation succession, with BG being more sensitive to severe temperatures. | [197] |
| Ziwuling forest region of the Loess Plateau | During extended vegetation succession, total soil nutrients had a greater impact on enzyme Kcat than accessible nutrients. The kinetic characteristics of soil enzymes changed dramatically over this succession. BG was more responsive to severe temperatures than NAG or AlP. At both low (5 °C) and high (35 °C) temperatures, the Vmax, half-saturation constant (Km), and other kinetic parameters of BG were disconnected. | [198] |
| Other | ||
| Permafrost peatland near the Tuqiang Forestry Bureau in the Great Xing’an Mountain, Heilongjiang Province, northeast China | PO demonstrated a greater response to temperature fluctuations compared to enzymes like BG, NAG, and AcP. The combined effects of rising temperatures and water flooding resulted in a synergistic impact, leading to an increase in both bacterial and fungal populations, as well as the activity levels of various soil enzymes. | [199] |
| High Arctic dry tundra, continuous permafrost zone, Cambridge Bay, Nunavut, Canada | The activities of BG, cellobiase, NAG, LAP, and PO peaked in June and dropped throughout the summer. Environmental conditions have a major impact on hydrolase activity fluctuations, influencing EEA and the structure of the Arctic microbial community. | [176] |
| Mediterranean climate gradient in southern California | Vmax of most enzymes was more sensitive to temperature in cooler environments, particularly during the dry season. Km was more sensitive in warmer areas, indicating enzyme build up in drier regions, which influenced respiration following rewetting occurrences. | [181] |
| Karst region of southwestern China | As vegetation succession progressed, AlP activity increased and Ure dropped. Ure was positively connected with rock outcrop cover but negatively with litter N, soil accessible N, and pH, whereas AlP showed the opposite correlation. | [200] |
| Soil Ecosystem/Location and Used Fertilizers (If Used) | Enzyme Activities and Other Important Remarks | Ref. |
|---|---|---|
| N addition | ||
| Soils from hardwood forests at Bear Brook, Maine, and Fernow Forest, West Virginia. | Vmax and Km for AG, BG, BX, CBH, and NAG increased with N addition, especially at Fernow. N fertilization reduced Km at Bear Brook, but had varied effects at Fernow. Both Vmax and Km were temperature sensitive, with BX demonstrating a substantial relationship between N and temperature for Km in hardwood forest soils. | [7] |
| Agricultural field, yellow clayey soil, located in Jingshan county, Hubei, China. NPK, NPK plus green manure (NPKG), NPK plus pig manure (NPKM), and NPK plus straw (NPKS) were used for fertilization. | NPKM treatment increased ARS, BG, AG, NAG, and CBH activities compared to the unfertilized control. Except for phosphomonoesterase and NAG, the NPKG and NPKS treatments had equal or lower activity levels. Low PO activity may result in soluble phenolic build-up, which inhibits hydrolytic enzymes. | [220] |
| The effect of simulated N deposition in six forest ecosystems in eastern China. Soil samples from three blocks × four soil depths (0–10 cm, 10–20 cm, 20–40 cm and 40–60 cm) were collected. | Four to five years of N addition exhibited little effect on BG, CBH, PO, PP, NAG, LAP, and AcP activities and ratios, with very minimal site-specific responses for AcP. | [236] |
| Soil of a Korean pine plantation in which different concentrations (0, 20, 40, 80 kg N ha−1 year−1) of ammonium nitrate were applied for 5 consecutive years. | Moderate N addition (40 kg N ha−1 year−1) significantly reduced Ure activity, with all three treatments exhibiting lower protease activity than control. There was no connection discovered between microbial community structure and four mineralizing enzymes, and N concentrations had no effect on soil pH. | [237] |
| Nash’s Field long-term grassland experiment established on acidic soils at Silwood Park, Berkshire, UK. 19 years of chronic N-only addition to permanent grassland was tested. | Chronic N addition over 19 years improved C storage and BG activity in thick soils. N fertilizer decreased root C:N ratios, which increased microbial demand for root C. Lime application reduced BG activity and root mass in high-pH soils. | [228] |
| 16-year experiment conducted in a typical grassland in northern China. | N addition inhibited BG and AcP, while H2O addition had no effect on BG but lowered AcP. Soil enzyme activity was mostly affected by soil microbial biomass C. | [93] |
| Short-term N addition (NH4NO3) in a sandy grassland and semi-fixed sandy land in the Horqin Sandy Land, northern China. | NAG activity and soil microbial features remained constant across N levels and locales. N addition increased BG activity in sandy grassland and semi-fixed sandy land. | [223] |
| Semi-arid grassland in China | N addition increased C-acquiring enzyme activity but lowered N-acquiring enzyme activity in low-precipitation years, while it stimulated all enzymes in high-precipitation years. | [238] |
| Temperate and alpine grassland ecosystems in China | Although N and P additions had little effect on SOC concentration, they did change soil pH, total N, and total P content. Only AcP was inhibited by P addition at the temperate meadow site; other EEA and stoichiometric ratios were unaffected. | [239] |
| Typical meadow soil (Vertisols) near to Görbeháza, Debrecen, Hungary. The field is cultivated by a rain-fed maize monoculture and fertilized continuously at different doses of NPK | Long-term NPK fertilization improved microbial tolerance to fluctuations in SM content. High rainfall decreased soil NO3− and nitrification rates. The EEA responded more to SM than NPK, with the highest AP, DHA, and INV activity in the drier year and the highest Ure activity in the wettest year. High NPK rates lowered soil DHA activity. | [240] |
| Five-year field fertilization experiment to study how N addition affected soil enzyme activity patterns in the topsoil (0–20 cm) and subsoil (20–40 cm) in a Tibetan alpine meadow | N addition altered soil EEA via pH variations. At greater N rates, N-induced soil acidification enhanced BG and Ure activities while maintaining AcP and decreasing PO activity. | [241] |
| Typical steppe ecosystem in Inner Mongolia | N additions reduced soil N-related hydrolytic enzyme activity. | [242] |
| P addition | ||
| Subtropical/tropical moist forest in Dinghushan Biosphere Reserve (DHSBR) which is an UNESCO/MAB site located in the middle Guangdong Province in southern China | P addition reduced AcP activity while increasing LAP activity but had no effect on LAP specific activity and lowered NAG specific activity. CBH, AG, and BG exhibited no significant reaction, however P addition reduced BX activity. It also lowered PO and PP activity, indicating a decrease in microbial enzyme synthesis in P-poor tropical forests. | [243] |
| Mesophytic deciduous forest soil on the unglaciated portion of the Allegheny Plateau, southeast Ohio, USA | All treatments lowered extracellular AP activity across both soil horizons. The reduction in AP resulted in a relative increase in C acquisition compared to N and P acquiring enzymes, affecting overall ecoenzymatic stoichiometry. | [244] |
| Desert steppe in Eastern Yanchi County, Ningxia Hui Autonomous Region, Northwest China | The enzyme stoichiometry was 1.2:1:1.5. Soil BG activity declined with heat and P addition, while AIP was reduced by warming, P addition, and warming combined with P addition. | [245] |
| Topsoil (0–75 mm) from a grazed pasture receiving contrasting P inputs. The field study was situated at Winchmore, New Zealand | Long-term P input decreased AcP while increasing AlP activity, which peaked in the summer and dropped in the winter. AcP and AlP linked positively with soil temperature but negatively with SM. | [246] |
| Two cotton cultivars and three phosphorus (P) levels. A pot experiment was conducted in 2017 at the Baibi station, Anyang, Henan, China | The activities of INV, cellulase, and urea in cotton soil decreased significantly after P addition. | [247] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daunoras, J.; Kačergius, A.; Gudiukaitė, R. Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem. Biology 2024, 13, 85. https://doi.org/10.3390/biology13020085
Daunoras J, Kačergius A, Gudiukaitė R. Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem. Biology. 2024; 13(2):85. https://doi.org/10.3390/biology13020085
Chicago/Turabian StyleDaunoras, Jokūbas, Audrius Kačergius, and Renata Gudiukaitė. 2024. "Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem" Biology 13, no. 2: 85. https://doi.org/10.3390/biology13020085
APA StyleDaunoras, J., Kačergius, A., & Gudiukaitė, R. (2024). Role of Soil Microbiota Enzymes in Soil Health and Activity Changes Depending on Climate Change and the Type of Soil Ecosystem. Biology, 13(2), 85. https://doi.org/10.3390/biology13020085