
Bibliometric Overview on T-Cell Intracellular Antigens and Their Pathological Implications


Abstract
Simple Summary
Abstract
1. Introduction
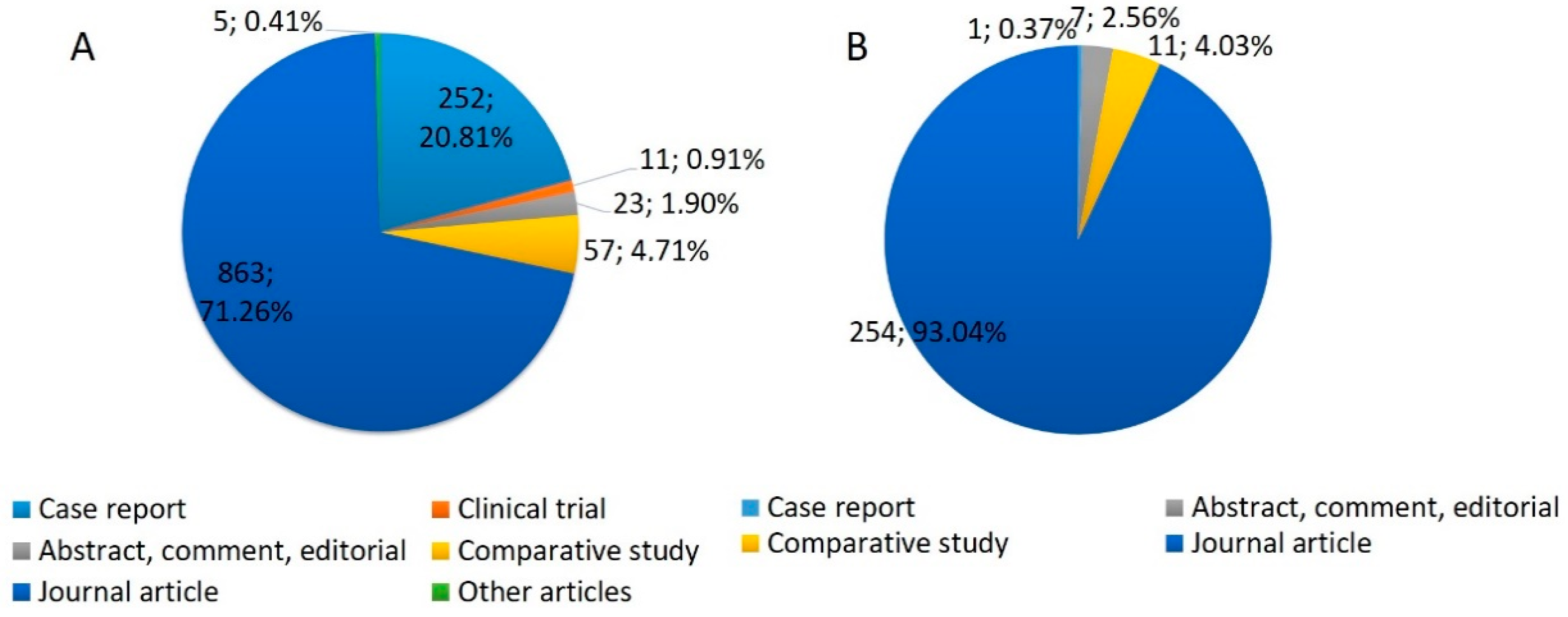
2. Features of Analyzed Publications
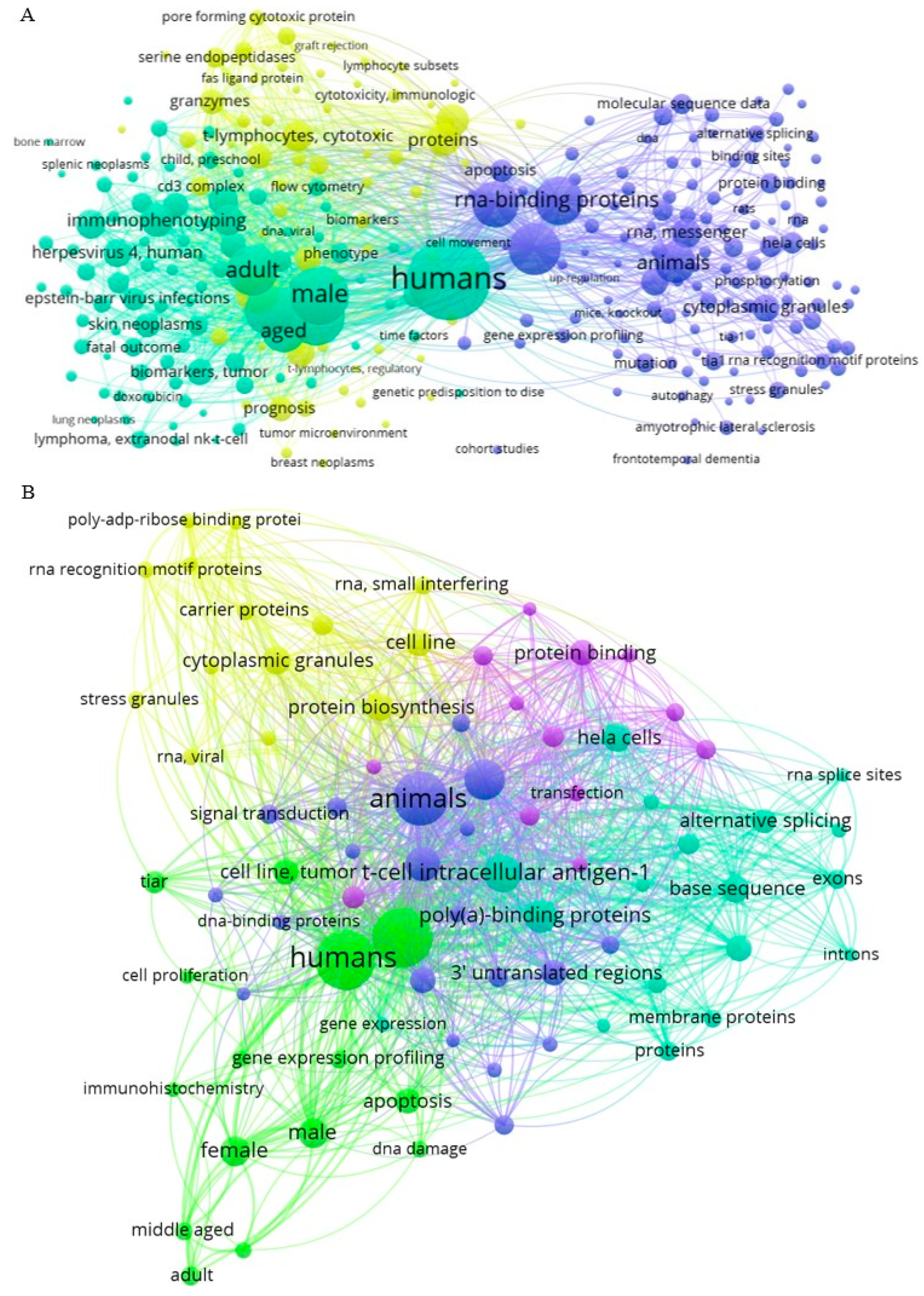
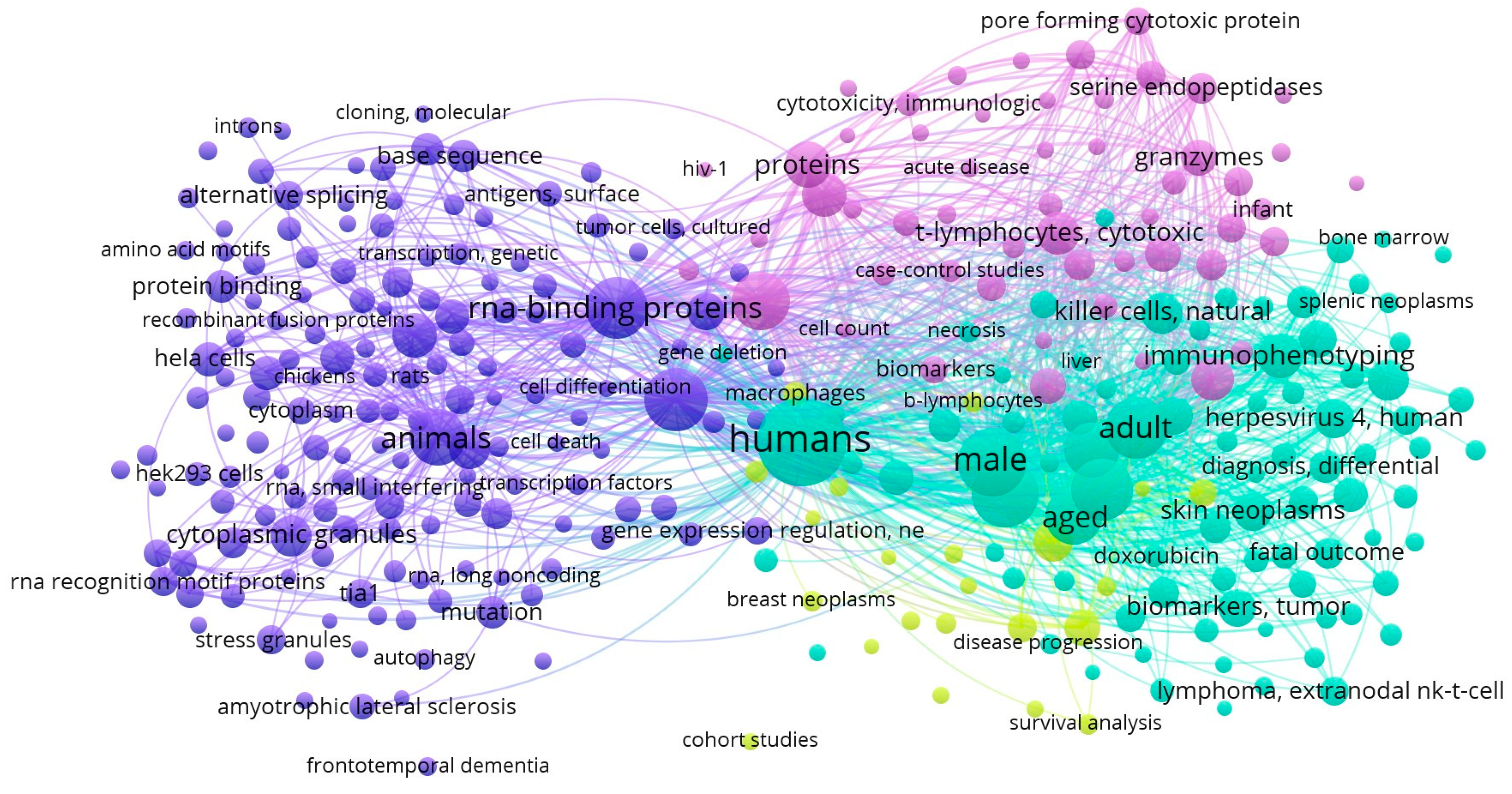
3. TIA1- and TIAR-Related Networking Analysis

4. TIA1- and TIAR-Associated Molecular Functions and Biological Processes
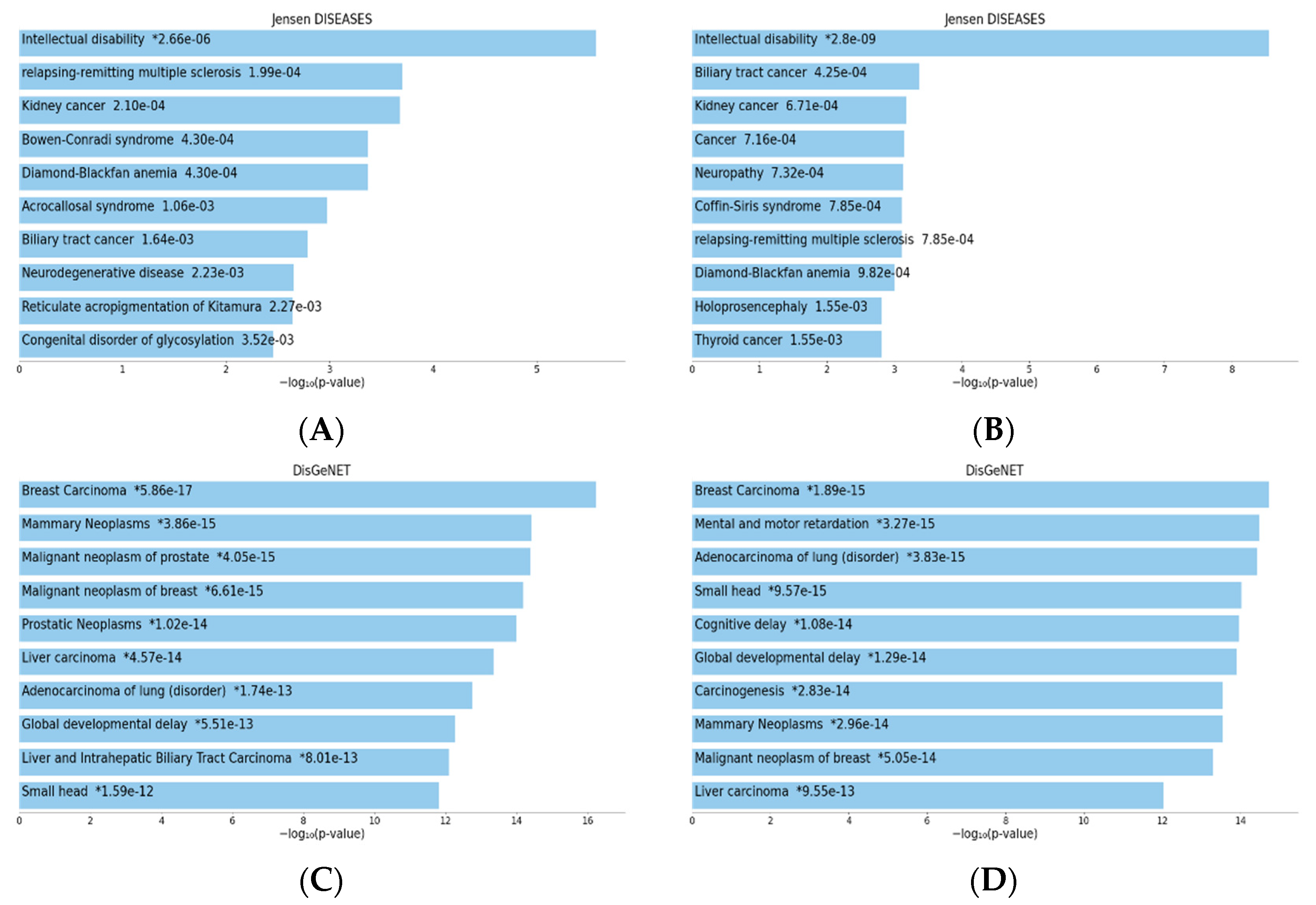
5. Implications of TIA1 and TIAR in Human Pathologies
6. Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, Q.; Streuli, M.; Saito, H.; Schlossman, S.F.; Anderson, P. A polyadenylate binding protein localized to the granules of cytolytic lymphocytes induces DNA fragmentation in target cells. Cell 1991, 67, 629–639. [Google Scholar] [CrossRef]
- Kawakami, A.; Tian, Q.; Streuli, M.; Poe, M.; Edelhoff, S.; Disteche, C.M.; Anderson, P. Intron-exon organization and chromosomal localization of the human TIA-1 gene. J. Immunol. Baltim. Md. 1950 1994, 152, 4937–4945. [Google Scholar] [CrossRef]
- Izquierdo, J.M.; Valcárcel, J. Two isoforms of the t-cell intracellular antigen 1 (TIA-1) splicing factor display distinct splicing regulation activities: Control of TIA-1 isoform ratio by TIA-1-related protein. J. Biol. Chem. 2007, 282, 19410–19417. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gómez, A.; Izquierdo, J.M. The multifunctional faces of T-cell intracellular antigen 1 in health and disease. Int. J. Mol. Sci. 2022, 23, 1400. [Google Scholar] [CrossRef]
- Kawakami, A.; Tian, Q.; Duan, X.; Streuli, M.; Schlossman, S.F.; Anderson, P. Identification and functional characterization of a TIA-1-related nucleolysin. Proc. Natl. Acad. Sci. USA 1992, 89, 8681–8685. [Google Scholar] [CrossRef] [PubMed]
- Velasco, B.R.; Izquierdo, J.M. T-cell intracellular antigen 1-like protein in physiology and pathology. Int. J. Mol. Sci. 2022, 23, 7836. [Google Scholar] [CrossRef]
- Aznarez, I.; Barash, Y.; Shai, O.; He, D.; Zielenski, J.; Tsui, L.-C.; Parkinson, J.; Frey, B.J.; Rommens, J.M.; Blencowe, B.J. A systematic analysis of intronic sequences downstream of 5′ splice sites reveals a widespread role for U-rich motifs and TIA1/TIAL1 proteins in alternative splicing regulation. Genome Res. 2008, 18, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Dember, L.M.; Kim, N.D.; Liu, K.Q.; Anderson, P. Individual RNA recognition motifs of TIA-1 and TIAR have different RNA binding specificities. J. Biol. Chem. 1996, 271, 2783–2788. [Google Scholar] [CrossRef]
- Meyer, C.; Garzia, A.; Mazzola, M.; Gerstberger, S.; Molina, H.; Tuschl, T. The TIA1 RNA-binding-protein family regulates EIF2AK2-mediated stress response and cell cycle progression. Mol. Cell 2018, 69, 622–635.e6. [Google Scholar] [CrossRef]
- López de Silanes, I.; Galbán, S.; Martindale, J.L.; Yang, X.; Mazan-Mamczarz, K.; Indig, F.E.; Falco, G.; Zhan, M.; Gorospe, M. Identification and functional outcome of mRNAs associated with RNA-binding protein TIA-1. Mol. Cell Biol. 2005, 25, 9520–9531. [Google Scholar] [CrossRef]
- Mazan-Mamczarz, K.; Lal, A.; Martindale, J.L.; Kawai, T.; Gorospe, M. Translational repression by RNA-binding protein TIAR. Mol. Cell Biol. 2006, 26, 2716–2727. [Google Scholar] [CrossRef]
- Sidali, A.; Teotia, V.; Solaiman, N.S.; Bashir, N.; Kanagaraj, R.; Murphy, J.J.; Surendranath, K. AU-rich element RNA binding proteins: At the crossroads of post-transcriptional regulation and genome integrity. Int. J. Mol. Sci. 2021, 23, 96. [Google Scholar] [CrossRef] [PubMed]
- Barron, V.A.; Lou, H. Alternative splicing of the neurofibromatosis type I pre-mRNA. Biosci. Rep. 2011, 32, 131–138. [Google Scholar] [CrossRef]
- Wang, Z.; Kayikci, M.; Briese, M.; Zarnack, K.; Luscombe, N.M.; Rot, G.; Zupan, B.; Curk, T.; Ule, J. iCLIP Predicts the dual splicing effects of TIA-RNA interactions. PLoS Biol. 2010, 8, e1000530. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhao, J.; Hou, M.; Wang, Y.; Zhang, Y.; Zhao, X.; Zhang, C.; Guo, D. HuR and TIA1/TIAL1 are involved in regulation of alternative splicing of SIRT1 pre-mRNA. Int. J. Mol. Sci. 2014, 15, 2946–2958. [Google Scholar] [CrossRef]
- Anderson, P.; Kedersha, N. Stressful initiations. J. Cell Sci. 2002, 115, 3227–3234. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Wheeler, J.R.; Walters, R.W.; Agrawal, A.; Barsic, A.; Parker, R. ATPase-modulated stress granules contain a diverse proteome and substructure. Cell 2016, 164, 487–498. [Google Scholar] [CrossRef]
- Kedersha, N.L.; Gupta, M.; Li, W.; Miller, I.; Anderson, P. RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2α to the assembly of mammalian stress granules. J. Cell Biol. 1999, 147, 1431–1442. [Google Scholar] [CrossRef]
- Waris, S.; Wilce, M.C.J.; Wilce, J.A. RNA recognition and stress granule formation by TIA proteins. Int. J. Mol. Sci. 2014, 15, 23377–23388. [Google Scholar] [CrossRef]
- Arimoto-Matsuzaki, K.; Saito, H.; Takekawa, M. TIA1 Oxidation inhibits stress granule assembly and sensitizes cells to stress-induced apoptosis. Nat. Commun. 2016, 7, 10252. [Google Scholar] [CrossRef]
- Bossowski, A.; Czarnocka, B.; Bardadin, K.; Moniuszko, A.; Łyczkowska, A.; Czerwinska, J.; Dadan, J.; Bossowska, A. Identification of chosen apoptotic (TIAR and TIA-1) markers expression in thyroid tissues from adolescents with immune and non-immune thyroid diseases. Folia Histochem. Cytobiol. 2010, 48, 178–184. [Google Scholar] [CrossRef][Green Version]
- Förch, P.; Puig, O.; Kedersha, N.; Martínez, C.; Granneman, S.; Séraphin, B.; Anderson, P.; Valcárcel, J. The apoptosis-promoting factor TIA-1 is a regulator of alternative pre-mRNA splicing. Mol. Cell 2000, 6, 1089–1098. [Google Scholar] [CrossRef]
- Reyes, R.; Alcalde, J.; Izquierdo, J.M. Depletion of T-cell intracellular antigen proteins promotes cell proliferation. Genome Biol. 2009, 10, R87. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.R.P.; Miller, I.J.; Anderson, P.; Streuli, M. RNA-binding protein TIAR is essential for primordial germ cell development. Proc. Natl. Acad. Sci. USA 1998, 95, 2331–2336. [Google Scholar] [CrossRef]
- Sánchez-Jiménez, C.; Izquierdo, J.M. T-cell intracellular antigen (TIA)-proteins deficiency in murine embryonic fibroblasts alters cell cycle progression and induces autophagy. PLoS ONE 2013, 8, e75127. [Google Scholar] [CrossRef] [PubMed]
- Geng, Z.; Li, P.; Tan, L.; Song, H. Targeted knockdown of RNA-binding protein TIAR for promoting self-renewal and attenuating differentiation of mouse embryonic stem cells. Stem Cells Int. 2015, 2015, e657325. [Google Scholar] [CrossRef][Green Version]
- Scheu, S.; Stetson, D.B.; Reinhardt, R.L.; Leber, J.H.; Mohrs, M.; Locksley, R.M. Activation of the integrated stress response during T helper cell differentiation. Nat. Immunol. 2006, 7, 644–651. [Google Scholar] [CrossRef]
- Barron, V.A.; Zhu, H.; Hinman, M.N.; Ladd, A.N.; Lou, H. The neurofibromatosis type I pre-mRNA is a novel target of CELF protein-mediated splicing regulation. Nucleic Acids Res. 2010, 38, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Rayman, J.B.; Hijazi, J.; Li, X.; Kedersha, N.; Anderson, P.J.; Kandel, E.R. Genetic Perturbation of TIA1 reveals a physiological role in fear memory. Cell Rep. 2019, 26, 2970–2983.e4. [Google Scholar] [CrossRef]
- Vanderweyde, T.; Youmans, K.; Liu-Yesucevitz, L.; Wolozin, B. Role of stress granules and RNA-binding proteins in neurodegeneration: A mini-review. Gerontology 2013, 59, 524–533. [Google Scholar] [CrossRef]
- Vanderweyde, T.; Yu, H.; Varnum, M.; Liu-Yesucevitz, L.; Citro, A.; Ikezu, T.; Duff, K.; Wolozin, B. Contrasting pathology of the stress granule proteins TIA-1 and G3BP in tauopathies. J. Neurosci. 2012, 32, 8270–8283. [Google Scholar] [CrossRef]
- Waelter, S.; Boeddrich, A.; Lurz, R.; Scherzinger, E.; Lueder, G.; Lehrach, H.; Wanker, E.E. Accumulation of mutant huntingtin fragments in aggresome-like inclusion bodies as a result of insufficient protein degradation. Mol. Biol. Cell 2001, 12, 1393–1407. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiao, B.; Hou, L.; Xiao, T.; Liu, X.; Wang, J.; Xu, J.; Zhou, L.; Yan, X.; Tang, B.; et al. Mutation analysis of the TIA1 gene in chinese patients with amyotrophic lateral sclerosis and frontotemporal dementia. Neurobiol. Aging. 2018, 64, 160.e9–160.e12. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, J.M.; Alcalde, J.; Carrascoso, I.; Reyes, R.; Ludeña, M.D. Knockdown of T-cell intracellular antigens triggers cell proliferation, invasion and tumour growth. Biochem. J. 2011, 435, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, R.; Yang, F.; Cheng, R.; Chen, X.; Cui, S.; Gu, Y.; Sun, W.; You, C.; Liu, Z.; et al. miR-19a Promotes colorectal cancer proliferation and migration by targeting TIA1. Mol. Cancer 2017, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Jiménez, C.; Ludeña, M.D.; Izquierdo, J.M. T-cell intracellular antigens function as tumor suppressor genes. Cell Death Dis. 2015, 6, e1669. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, M.; Lin, B.; Yao, D.; Li, J.; Tang, X.; Li, S.; Liu, Y.; Xie, R.; Yu, S. miR-487a promotes progression of gastric cancer by targeting TIA1. Biochimie 2018, 154, 119–126. [Google Scholar] [CrossRef]
- Liu, J.; Cao, X. RBP–RNA interactions in the control of autoimmunity and autoinflammation. Cell Res. 2023, 33, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Khan, R.A.; Giddaluru, J.; Battu, S.; Vishwakarma, S.K.; Subahan, M.; Satti, V.; Khan, N.; Khan, A.A. Transcriptome meta-analysis identifies immune signature comprising of RNA binding proteins in ulcerative colitis patients. Cell Immunol. 2018, 334, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Dinh, P.X.; Beura, L.K.; Das, P.B.; Panda, D.; Das, A.; Pattnaik, A.K. Induction of stress granule-like structures in vesicular stomatitis virus-infected cells. J. Virol. 2013, 87, 372–383. [Google Scholar] [CrossRef]
- Le Sage, V.; Cinti, A.; McCarthy, S.; Amorim, R.; Rao, S.; Daino, G.L.; Tramontano, E.; Branch, D.R.; Mouland, A.J. Ebola virus VP35 blocks stress granule assembly. Virology 2017, 502, 73–83. [Google Scholar] [CrossRef]
- Lloyd, R.E. Nuclear proteins hijacked by mammalian cytoplasmic plus strand RNA viruses. Virology 2015, 479–480, 457–474. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.; Khaperskyy, D.A. Translation inhibition and stress granules in the antiviral immune response. Nat. Rev. Immunol. 2017, 17, 647–660. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dong, L.; Yu, S.; Wang, X.; Zheng, H.; Zhang, P.; Meng, C.; Zhan, Y.; Tan, L.; Song, C.; et al. Newcastle disease virus induces stable formation of bona fide stress granules to facilitate viral replication through manipulating host protein translation. FASEB J. 2017, 31, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- LeBlang, C.J.; Medalla, M.; Nicoletti, N.W.; Hays, E.C.; Zhao, J.; Shattuck, J.; Cruz, A.L.; Wolozin, B.; Luebke, J.I. Reduction of the RNA binding protein TIA1 exacerbates neuroinflammation in tauopathy. Front. Neurosci. 2020, 14, 285. [Google Scholar] [CrossRef] [PubMed]
- Piecyk, M.; Wax, S.; Beck, A.R.; Kedersha, N.; Gupta, M.; Maritim, B.; Chen, S.; Gueydan, C.; Kruys, V.; Streuli, M.; et al. TIA-1 is a translational silencer that selectively regulates the expression of TNF-alpha. EMBO J. 2000, 19, 4154–4163. [Google Scholar] [CrossRef] [PubMed]
- Simarro, M.; Giannattasio, G.; Xing, W.; Lundequist, E.-M.; Stewart, S.; Stevens, R.L.; Orduña, A.; Boyce, J.A.; Anderson, P.J. The translational repressor T-cell intracellular antigen-1 (TIA-1) is a key modulator of Th2 and Th17 responses driving pulmonary inflammation induced by exposure to house dust mite. Immunol. Lett. 2012, 146, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; York, B.; Wang, S.; Feng, Q.; Xu, J.; O’Malley, B.W. An essential function of the SRC-3 coactivator in suppression of cytokine mRNA translation and inflammatory response. Mol. Cell 2007, 25, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Maziuk, B.; Ballance, H.I.; Wolozin, B. Dysregulation of RNA binding protein aggregation in neurodegenerative disorders. Front. Mol. Neurosci. 2017, 10, 89. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Frater, J.L.; Klein, J.; Chen, L.; Bartlett, N.L.; Foyil, K.V.; Kreisel, F.H. Expression of TIA1 and PAX5 in classical hodgkin lymphoma at initial diagnosis may predict clinical outcome. Appl. Immunohistochem. Mol. Morphol. AIMM 2016, 24, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Hu, Y.; Fan, Z.; Gao, R.; Yang, R.; Bi, J.; Hou, J. In silico identification of EP400 and TIA1 as critical transcription factors involved in human hepatocellular carcinoma relapse. Oncol. Lett. 2020, 19, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Hamada, J.; Shoda, K.; Masuda, K.; Fujita, Y.; Naruto, T.; Kohmoto, T.; Miyakami, Y.; Watanabe, M.; Kudo, Y.; Fujiwara, H.; et al. Tumor-promoting function and prognostic significance of the RNA-binding protein T-cell intracellular antigen-1 in esophageal squamous cell carcinoma. Oncotarget 2016, 7, 17111–17128. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Liu, W.; Li, G.; Li, F.; Zhang, S. Epstein-Barr virus infection and expression of T-cell intracellular antigen-1 (TIA-1) in intestinal T-cell lymphoma. Zhonghua Bing. Li Xue Za Zhi 1999, 28, 348–351. [Google Scholar]
- Zlobec, I.; Karamitopoulou, E.; Terracciano, L.; Piscuoglio, S.; Iezzi, G.; Muraro, M.G.; Spagnoli, G.; Baker, K.; Tzankov, A.; Lugli, A. TIA-1 Cytotoxic granule-associated RNA binding protein improves the prognostic performance of CD8 in mismatch repair-proficient colorectal cancer. PLoS ONE 2010, 5, e14282. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Murakami, Y.I.; Shimada, S.; Iwamizu-Watanabe, S.; Yamashita, Y.; Hasegawa, Y.; Kojima, H.; Nagasawa, T. TIA-1 expression in hairy cell leukemia. Mod. Pathol. 2004, 17, 840–846. [Google Scholar] [CrossRef]
- Boulland, M.L.; Kanavaros, P.; Wechsler, J.; Casiraghi, O.; Gaulard, P. Cytotoxic protein expression in natural killer cell lymphomas and in alpha beta and gamma delta peripheral T-cell lymphomas. J. Pathol. 1997, 183, 432–439. [Google Scholar] [CrossRef]
- Kim, H.S.; Kuwano, Y.; Zhan, M.; Pullmann, R.; Mazan-Mamczarz, K.; Li, H.; Kedersha, N.; Anderson, P.; Wilce, M.C.J.; Gorospe, M.; et al. Elucidation of a C-rich signature motif in target mRNAs of RNA-binding protein TIAR. Mol. Cell Biol. 2007, 27, 6806–6817. [Google Scholar] [CrossRef]
- Taupin, J.L.; Tian, Q.; Kedersha, N.; Robertson, M.; Anderson, P. The RNA-binding protein TIAR is translocated from the nucleus to the cytoplasm during Fas-mediated apoptotic cell death. Proc. Natl. Acad. Sci. USA 1995, 92, 1629–1633. [Google Scholar] [CrossRef]
- Kedersha, N.; Cho, M.R.; Li, W.; Yacono, P.W.; Chen, S.; Gilks, N.; Golan, D.E.; Anderson, P. Dynamic shuttling of TIA-1 accompanies the recruitment of mRNA to mammalian stress granules. J. Cell Biol. 2000, 151, 1257–1268. [Google Scholar] [CrossRef]
- Kedersha, N.; Chen, S.; Gilks, N.; Li, W.; Miller, I.J.; Stahl, J.; Anderson, P. Evidence that ternary complex (eIF2-GTP-tRNA(i)(Met))-deficient preinitiation complexes are core constituents of mammalian stress granules. Mol. Biol. Cell 2002, 13, 195–210. [Google Scholar] [CrossRef]
- Dean, J.L.E.; Sully, G.; Clark, A.R.; Saklatvala, J. The involvement of AU-rich element-binding proteins in p38 mitogen-activated protein kinase pathway-mediated mRNA stabilisation. Cell Signal. 2004, 16, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Duttagupta, R.; Tian, B.; Wilusz, C.J.; Khounh, D.T.; Soteropoulos, P.; Ouyang, M.; Dougherty, J.P.; Peltz, S.W. Global analysis of pub1p targets reveals a coordinate control of gene expression through modulation of binding and stability. Mol. Cell Biol. 2005, 25, 5499–5513. [Google Scholar] [CrossRef][Green Version]
- García-Mauriño, S.M.; Rivero-Rodríguez, F.; Velázquez-Cruz, A.; Hernández-Vellisca, M.; Díaz-Quintana, A.; De la Rosa, M.A.; Díaz-Moreno, I. RNA binding protein regulation and cross-talk in the control of AU-rich mRNA fate. Front. Mol. Biosci. 2017, 4, 71. [Google Scholar] [CrossRef] [PubMed]
- Stoecklin, G.; Stubbs, T.; Kedersha, N.; Wax, S.; Rigby, W.F.C.; Blackwell, T.K.; Anderson, P. MK2-induced Tristetraprolin:14-3-3 complexes prevent stress granule association and ARE-mRNA decay. EMBO J. 2004, 23, 1313–1324. [Google Scholar] [CrossRef]
- Katsanou, V.; Papadaki, O.; Milatos, S.; Blackshear, P.J.; Anderson, P.; Kollias, G.; Kontoyiannis, D.L. HuR as a negative posttranscriptional modulator in inflammation. Mol. Cell 2005, 19, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Maeda, K. Control of RNA stability in immunity. Annu. Rev. Immunol. 2021, 39, 481–509. [Google Scholar] [CrossRef]
- van Eck, N.J.; Waltman, L. Visualizing bibliometric networks. In Measuring Scholarly Impact: Methods and Practice; Ding, Y., Rousseau, R., Wolfram, D., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 285–320. ISBN 978-3-319-10377-8. [Google Scholar]
- Carrascoso, I.; Alcalde, J.; Tabas-Madrid, D.; Oliveros, J.C.; Izquierdo, J.M. transcriptome-wide analysis links the short-term expression of the b isoforms of TIA proteins to protective proteostasis-mediated cell quiescence response. PLoS ONE 2018, 13, e0208526. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene set knowledge discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Horste, E.L.; Fansler, M.M.; Cai, T.; Chen, X.; Mitschka, S.; Zhen, G.; Lee, F.C.Y.; Ule, J.; Mayr, C. Subcytoplasmic location of translation controls protein output. Mol. Cell 2023, 83, 4509–4523.e11. [Google Scholar] [CrossRef]
- Van Nostrand, E.L.; Freese, P.; Pratt, G.A.; Wang, X.; Wei, X.; Xiao, R.; Blue, S.M.; Chen, J.Y.; Cody, N.A.L.; Dominguez, D.; et al. A large-scale binding and functional map of human RNA-binding proteins. Nature 2020, 583, 711–719. [Google Scholar] [CrossRef]
- Cho, N.H.; Cheveralls, K.C.; Brunner, A.D.; Kim, K.; Michaelis, A.C.; Raghavan, P.; Kobayashi, H.; Savy, L.; Li, J.Y.; Canaj, H.; et al. OpenCell: Endogenous tagging for the cartography of human cellular organization. Science 2022, 375, eabi6983. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Journals | Country | Documents | IF |
|---|---|---|---|---|
| TIA1 | Zhonghua Bing Li Xue Za Zhi = Chinese Journal of Pathology | China | 38 | 8.0 |
| The American Journal of Surgical Pathology | USA | 35 | 6.298 | |
| Histopathology | UK | 24 | 7.778 | |
| Journal of Virology | USA | 24 | 6.549 | |
| PLoS One | USA | 24 | 3.752 | |
| Journal of Cutaneous Pathology | Denmark | 21 | 1.458 | |
| Molecular and Cellular Biology | USA | 21 | 1.68 | |
| Virchows Archive: An International Journal of Pathology | Germany | 20 | 1.709 | |
| The American Journal of Dermatopathology | USA | 19 | 1.391 | |
| Pathology | USA | 18 | 3.526 | |
| TIAR | The Journal of Biological Chemistry | USA | 17 | 5.486 |
| Molecular and Cellular Biology | USA | 14 | 5.094 | |
| Journal of Virology | USA | 12 | 10.1 | |
| PLoS One | USA | 10 | 3.752 | |
| Nucleic Acids Research | UK | 9 | 19.160 | |
| Biochemical and Biophysical Research Communications | USA | 6 | 3.575 | |
| International Journal of Molecular Sciences | Switzerland | 5 | 6.208 | |
| Molecular Biology of the Cell | USA | 5 | 3.612 | |
| Molecular Cell | USA | 5 | 19.328 | |
| Proceedings of the National Academy of Sciences (USA) | USA | 5 | 12.779 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Velasco, B.; Naranjo, R.; Izquierdo, J.M. Bibliometric Overview on T-Cell Intracellular Antigens and Their Pathological Implications. Biology 2024, 13, 195. https://doi.org/10.3390/biology13030195
Ramos-Velasco B, Naranjo R, Izquierdo JM. Bibliometric Overview on T-Cell Intracellular Antigens and Their Pathological Implications. Biology. 2024; 13(3):195. https://doi.org/10.3390/biology13030195
Chicago/Turabian StyleRamos-Velasco, Beatriz, Rocío Naranjo, and José M. Izquierdo. 2024. "Bibliometric Overview on T-Cell Intracellular Antigens and Their Pathological Implications" Biology 13, no. 3: 195. https://doi.org/10.3390/biology13030195
APA StyleRamos-Velasco, B., Naranjo, R., & Izquierdo, J. M. (2024). Bibliometric Overview on T-Cell Intracellular Antigens and Their Pathological Implications. Biology, 13(3), 195. https://doi.org/10.3390/biology13030195