Waterbird Species Are Highly Sensitive to Wetland Traits: Simulation-Based Conservation Strategies for the Birds of the Sicilian Wetlands (Italy)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Selection
2.2. Field Surveys
2.3. Model Calibration
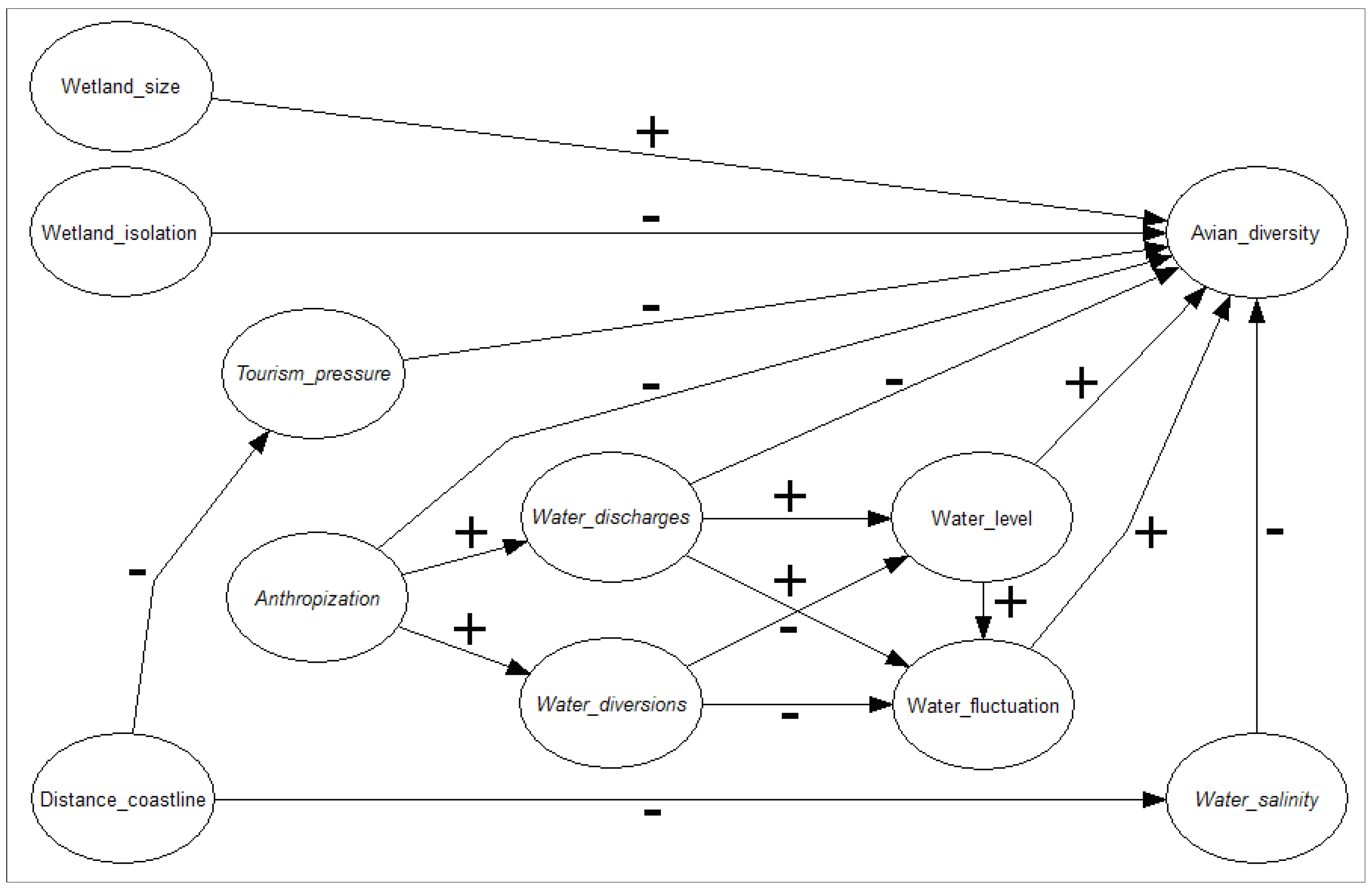
2.4. Disentangled Causal Effects on Avian Diversity
2.5. Model Validation
2.6. Simulations
3. Results
3.1. Sicilian Wetlands
3.2. Wetland Pantano Bruno
4. Discussion
4.1. Methodological Issues
4.2. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, R.C.; Barchiesi, S.; Beltrame, C.; Finlayson, C.M.; Galewski, T.; Harrison, I.J.; Paganini, M.; Perennou, C.; Rosenqvist, A.; Walpole, M.; et al. State of the World’s Wetlands and Their Services to People: A Compilation of Recent Analyses; Social Science Electronic Publishing: Gland, Switzerland, 2015. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; John Wiley & Sons: New York, NY, USA, 2015. [Google Scholar]
- Gardner, R.C.; Finlayson, C.M. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Ramsar Convention: Gland, Switzerland, 2018. [Google Scholar]
- Fraser, L.H.; Keddy, P.A. The World’s Largest Wetlands: Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Berthold, P. Bird Migration: A General Survey; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Keddy, P.A. Wetland Ecology: Principles and Conservation, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Costanza, R.; De Groot, R.; Sutton, P.; Van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Change 2014, 26, 152–158. [Google Scholar] [CrossRef]
- De Groot, R.; Brander, L.; Ploeg, R.; Costanza, F.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst. Serv. 2012, 1, 50–61. [Google Scholar] [CrossRef]
- Jensen, R.A.; Wisz, M.S.; Madsen, J. Prioritizing refuge sites for migratory geese to alleviate conflicts with agriculture. Biol. Conserv. 2008, 141, 1806–1818. [Google Scholar] [CrossRef]
- Mao, Q.; Liao, C.; Wu, Z.; Guan, W.; Yang, W.; Tang, Y.; Wu, G. Effects of land cover pattern along urban-rural gradient on bird diversity in wetlands. Diversity 2019, 11, 86. [Google Scholar] [CrossRef]
- Erwin, K.L. Wetlands and global climate change: The role of wetland restoration in a changing world. Wetl. Ecol. Manag. 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Gabler, C.A.; Osland, M.J.; Grace, J.B.; Stagg, C.L.; Day, R.B.; Hartley, S.B.; Enwright, N.M.; From, A.S.; McCoy, M.L.; McLeod, J.L. Macroclimatic change expected to transform coastal wetland ecosystems this century. Nat. Clim. Change 2017, 7, 142–147. [Google Scholar] [CrossRef]
- Osland, M.J.; Enwright, N.M.; Day, R.H.; Gabler, C.A.; Stagg, C.L.; Grace, J.B. Beyond just sea-level rise: Considering macroclimatic drivers within coastal wetland vulnerability assessments to climate change. Glob. Change Biol. 2016, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Barlow, P.M.; Reichard, E.G. Saltwater intrusion in coastal regions of North America. Hydrogeol. J. 2010, 18, 247–260. [Google Scholar] [CrossRef]
- Rodríguez-Santalla, I.; Navarro, N. Main Threats in Mediterranean Coastal Wetlands. The Ebro Delta Case. J. Mar. Sci. Eng. 2021, 9, 1190. [Google Scholar] [CrossRef]
- Spina, F.; Volponi, S. Atlante della Migrazione Degli Uccelli in Italia; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 2008. [Google Scholar]
- Bijlsma, R.G. Bottleneck Areas for Migratory Birds in the Mediterranean Region: An Assessment of the Problems and Recommendations for Action; International Council for Bird Preservation: Cambridge, UK, 1990. [Google Scholar]
- Heath, M.F.; Evans, M.I. (Eds.) Important Bird Areas in Europe. Priority Sites for Conservation; BirdLife International: Cambridge, UK, 2000; Volume 2. [Google Scholar]
- Ferrarini, A.; Celada, C.; Gustin, M. Preserving the Mediterranean bird flyways: Assessment and prioritization of 38 main wetlands under human and climate threats in Sardinia and Sicily (Italy). Sci. Total Environ. 2020, 751, 141556. [Google Scholar] [CrossRef]
- Ferrarini, A.; Celada, C.; Gustin, M. Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands. Diversity 2022, 14, 696. [Google Scholar] [CrossRef]
- Ferrarini, A.; Gustin, M.; Celada, C. Twenty-three years of land-use changes induced considerable threats to the main wetlands of Sardinia and Sicily (Italy) along the Mediterranean bird flyways. Diversity 2021, 13, 240. [Google Scholar] [CrossRef]
- Evans, D. Building the European Union’s Natura 2000 network. Nat. Conserv. 2012, 1, 11–26. [Google Scholar] [CrossRef]
- Griffith, D.A. Spatial Autocorrelation: A Primer; Association of American Geographers: Washington, DC, USA, 1987. [Google Scholar]
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A fixed- radius point count method for nonbreeding and breeding season use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef]
- Cooke, R.M.; Kurowicka, D.; Hanea, A.M.; Morales Nápoles, O.; Ababei, D.A.; Ale, B.; Roelen, A. Continuous/Discrete Non-Parametric Bayesian Belief Nets with UNICORN and Uninet; Technical Report; Delft University: Delft, The Netherlands, 2017. [Google Scholar]
- Hanea, A.M.; Kurowicka, D.; Cooke, R.M.; Ababei, D.A. Mining and visualising ordinal data with non-parametric continuous BBNs. Comput. Stat. Data Anal. 2010, 54, 668–687. [Google Scholar] [CrossRef]
- Hanea, A.; Napoles, O.M.; Ababei, D. Non-parametric Bayesian networks: Improving theory and reviewing applications. Reliab. Eng. Syst. Saf. 2015, 144, 265–284. [Google Scholar] [CrossRef]
- Sklar, M. Fonctions de repartition an dimensions et leurs marges. Publ. Inst. Statist. Univ. Paris 1959, 8, 229–231. [Google Scholar]
- Zilko, A.A.; Kurowicka, D. Copula in a multivariate mixed discrete–continuous model. Comput. Stat. Data Anal. 2016, 103, 28–55. [Google Scholar] [CrossRef]
- Ferrarini, A.; Gustin, M.; Celada, C. Wetland attributes significantly affect patterns of bird species distribution in the Sardinian wetlands (Italy): An uncertain future for waterbird conservation. J. Appl. Ecol. 2023, 60, 650–660. [Google Scholar] [CrossRef]
- Ferrarini, A.; Gustin, M.; Celada, C. Simulation modeling unveils the unalike effects of alternative strategies for waterbird conservation in the coastal wetlands of Sardinia (Italy). Biology 2023, 12, 1440. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhou, L.; Xia, S.; Zhou, J. Impact of Urbanisation Intensity on Bird Diversity in River Wetlands around Chaohu Lake, China. Animals 2022, 12, 473. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.M.; Adam, P. Climate Change and Intertidal Wetlands. Biology 2013, 2, 445–480. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhu, S.; Gao, J.; Jiang, H.; Deng, G.; Sheng, L.; Cao, Y.; Li, L.; Lin, B. The Influence of Ecological Engineering on Waterbird Diversity in Different Habitats within the Xianghai Nature Reserve. Diversity 2022, 14, 1016. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P. Ecological Assembly Rules: Perspectives, Advances, Retreats; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- European Environment Agency. Protected Areas in Europe—An Overview; EEA: Copenhagen, Denmark, 2012. [Google Scholar]
- Kirby, J.; Davidson, N.; Giles, N.; Owen, M.; Spray, C. Waterbirds and Wetland Recreation Handbook. A Review of Issues and Management Practice; The Wildfowl and Wetlands Trust Publisher: Gloucester, UK, 2004. [Google Scholar]
- Rubega, M.A.; Robinson, J.A. Water salinization and shorebirds: Emerging issues. Int. Wader Stud. 1997, 9, 45–54. [Google Scholar]
- Dezvareh, G.A.; Nabavi, E.; Shamskilani, M.; Darban, A.K. Water salinity reduction using the phytoremediation method by three plant species and analyzing their behavior. Water Air Soil Pollut. 2023, 234, 90. [Google Scholar] [CrossRef]
- Enwright, N.M.; Griffith, K.T.; Osland, M.J. Barriers to and opportunities for landward migration of coastal wetlands with sea-level rise. Front. Ecol. Environ. 2016, 14, 307–316. [Google Scholar] [CrossRef]
- Taft, O.W.; Colwell, M.A.; Isola, C.R.; Safran, R.J. Waterbird responses to experimental drawdown: Implications for the multispecies management of wetland mosaics. J. Appl. Ecol. 2002, 39, 987–1001. [Google Scholar] [CrossRef]
- Elphick, C.S. Experimental approaches to shorebird habitat management. Int. Wader Stud. 1997, 9, 20–28. [Google Scholar]
- Elphick, C.S.; Oring, L.W. Winter management of Californian rice fields for waterbirds. J. Appl. Ecol. 1998, 35, 95–108. [Google Scholar] [CrossRef]
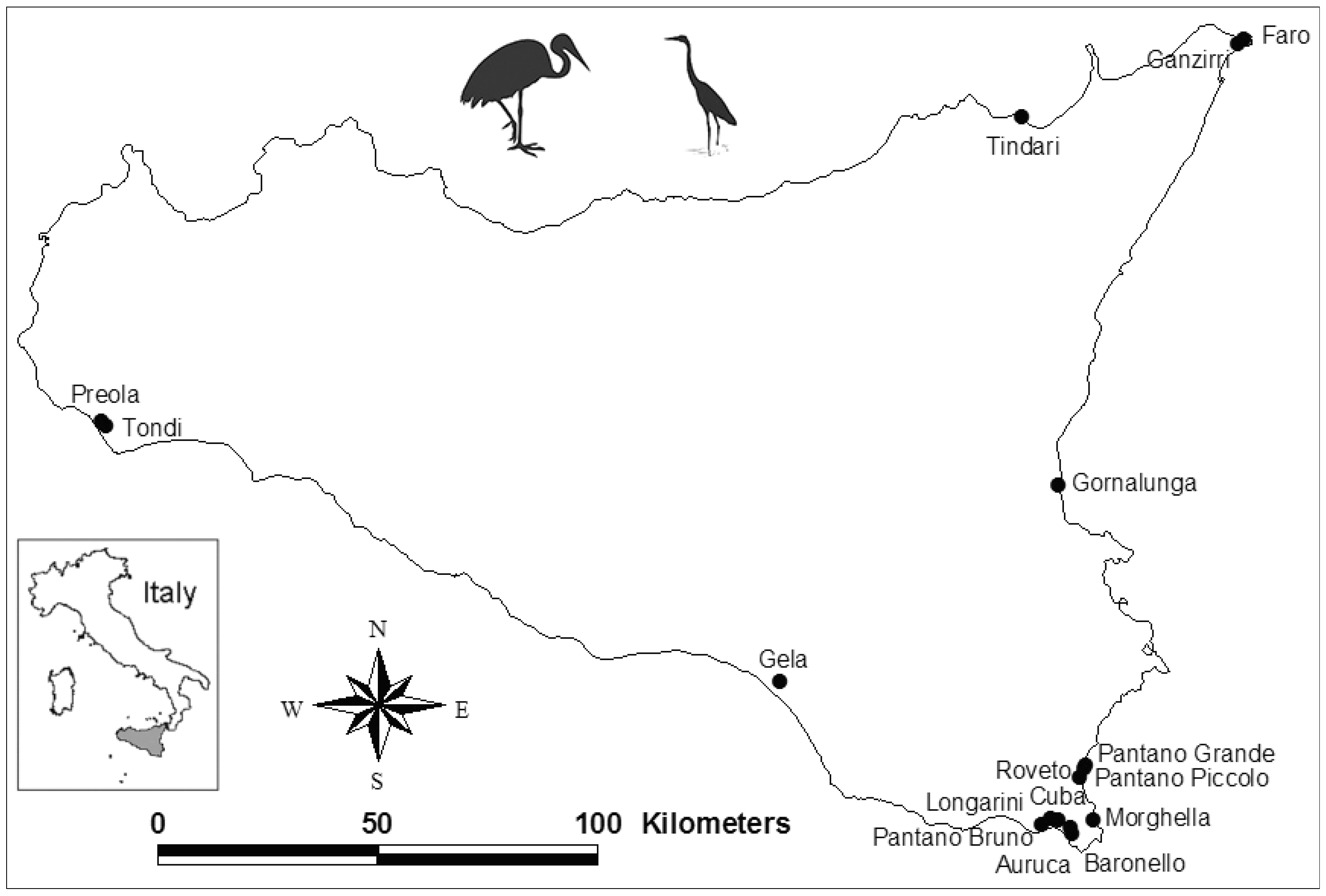



{kind=link}
{kind=link}
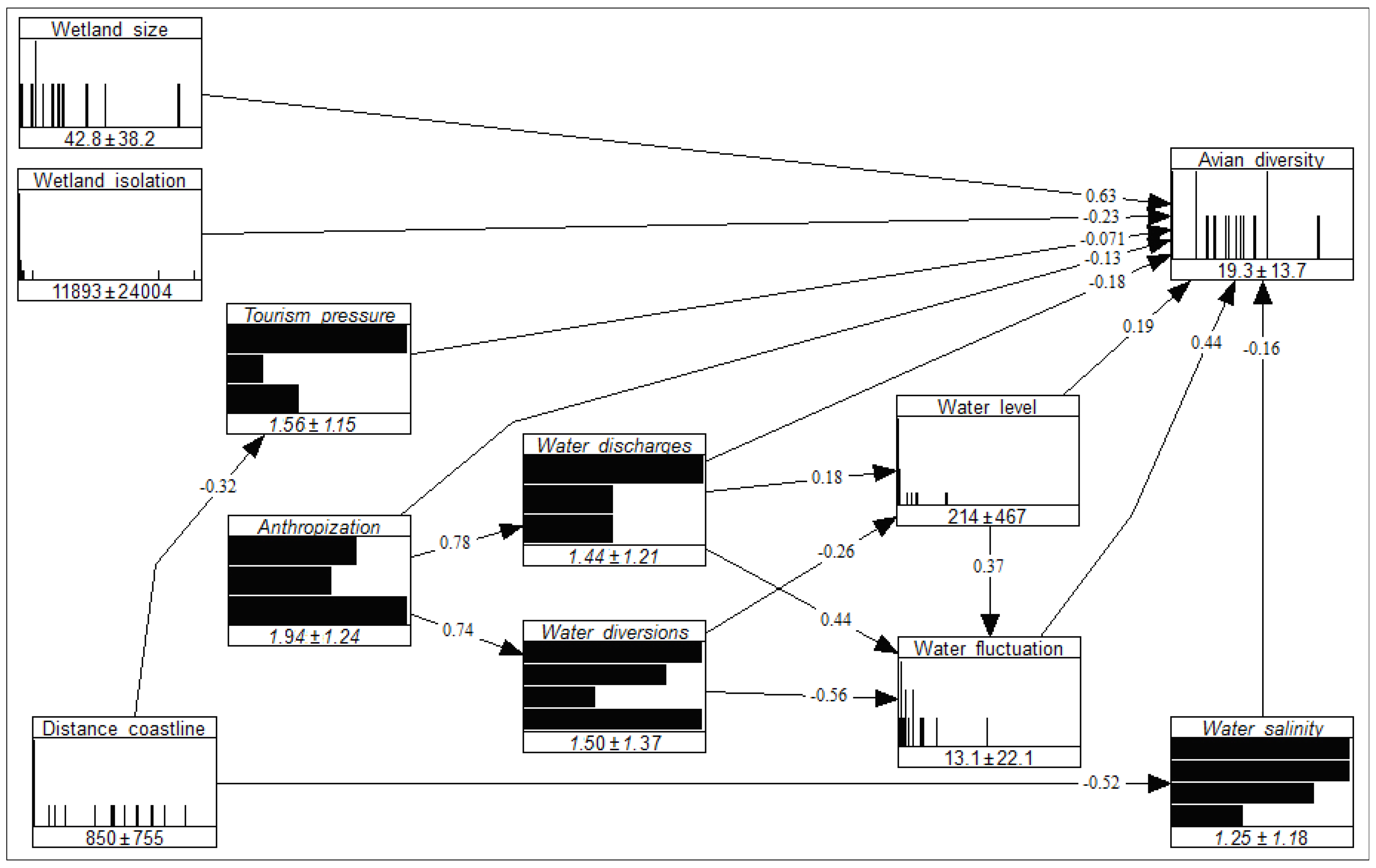{kind=link}
| Wetland Trait | Direct Effect on Alpha Avian Diversity | Indirect Effect on Alpha Avian Diversity | Total Effect on Alpha Avian Diversity |
|---|---|---|---|
| Anthropization | −0.13 | −0.02 | −0.15 |
| Distance to the coastline | 0.00 | 0.11 | 0.11 |
| Mean water level | 0.19 | 0.16 | 0.35 |
| Tourism pressure | −0.07 | 0.00 | −0.07 |
| Water discharges | −0.18 | 0.23 | 0.05 |
| Water diversions | 0.00 | −0.30 | −0.30 |
| Water level fluctuations | 0.44 | 0.00 | 0.44 |
| Water salinity | −0.16 | 0.00 | −0.16 |
| Wetland isolation | −0.23 | 0.00 | −0.23 |
| Wetland size | 0.63 | 0.00 | 0.63 |
| Code | Counterfactuality | Conditionalization | Avian Diversity (Mean ± S.D.) |
|---|---|---|---|
| S0 | none | none | 19.3 ± 13.7 |
| S1 | tourism pressure is widespread in all wetlands | tourism pressure = 3 | 18.2 ± 12.8 |
| S2 | water salinity is widespread in all wetlands | water salinity = 3 | 16.2 ± 12.1 |
| S3 | water discharges are null in all wetlands | water discharges = 0 | 17.7 ± 12.8 |
| S4 | anthropization is widespread in all wetlands | anthropization = 3 | 17.9 ± 12.6 |
| S5 | water diversions are widespread in all wetlands | water diversions = 3 | 15.5 ± 11.7 |
| S6 | water level fluctuations are null in all wetlands | water level fluctuations = 0 cm | 10.3 ± 8.7 |
| S7 | all conditions deteriorate (worst possible scenario) | scenarios from S1 to S6 together | 6.1 ± 6.6 |
| S8 | tourism pressure is null in all wetlands | tourism pressure = 0 | 19.8 ± 13.3 |
| S9 | water salinity is null in all wetlands | water salinity = 0 | 21.5 ± 13.6 |
| S10 | water discharges are widespread in all wetlands | water discharges = 3 | 20.4 ± 13.4 |
| S11 | anthropization is null in all wetlands | anthropization = 0 | 20.3 ± 13.4 |
| S12 | water diversions are null in all wetlands | water diversion = 0 | 23.2 ± 13.9 |
| S13 | water level fluctuations are 30 cm in all wetlands | water level fluctuations = 30 cm | 29.7 ± 14.0 |
| S14 | all conditions improve (best possible scenario) | scenarios from S8 to S13 together | 38.5 ± 13.6 |
| S15 | all conditions deteriorate but tourism pressure is null | same as S7 but tourism pressure = 0 | 6.6 ± 6.8 |
| S16 | all conditions deteriorate but water salinity is null | same as S7 but water salinity = 0 | 8.4 ± 8.2 |
| S17 | all conditions deteriorate but water discharges are widespread | same as S7 but water discharges = 3 | 8.1 ± 8.4 |
| S18 | all conditions deteriorate but anthropization is null | same as S7 but anthropization = 0 | 7.9 ± 6.1 |
| S19 | all conditions deteriorate but water diversions are null | same as S7 but water diversions = 0 | 9.7 ± 7.3 |
| S20 | all conditions deteriorate but water level fluctuations equal 30 cm | same as S7 but water level fluctuations = 30 cm | 20.6 ± 11.8 |
| Code | Counterfactuality | Water Level Fluctuations (B) | Water Salinity (C) | Water Diversions (C) | Water Discharges (B) | Tourism Pressure (C) | No. of Waterbird Species |
|---|---|---|---|---|---|---|---|
| S0 | none | 6.8 | 0 | 2 | 2 | 1 | 19 |
| S21 | increase in water salinity | 6.8 | 3 | 2 | 2 | 1 | 14 |
| S22 | increase in tourism pressure | 6.8 | 0 | 2 | 2 | 3 | 18 |
| S23 | decrease in water discharges | 6.8 | 0 | 2 | 0 | 1 | 15 |
| S24 | increase in water diversions | 6.8 | 0 | 3 | 2 | 1 | 12 |
| S25 | water level fluctuations = 5 cm | 5.0 | 0 | 2 | 2 | 1 | 16 |
| S26 | water level fluctuations = 0 cm | 0.0 | 0 | 2 | 2 | 1 | 8 |
| S27 | S21 and S22 together | 6.8 | 3 | 2 | 2 | 3 | 13 |
| S28 | S21 and S23 together | 6.8 | 3 | 2 | 0 | 1 | 11 |
| S29 | S21 and S24 together | 6.8 | 3 | 3 | 2 | 1 | 9 |
| S30 | S21 and S25 together | 5.0 | 3 | 2 | 2 | 1 | 12 |
| S31 | S21 and S26 together | 0.0 | 3 | 2 | 2 | 1 | 4 |
| S32 | S21, S23, and S25 together | 5.0 | 3 | 2 | 0 | 1 | 10 |
| S33 | worst possible scenario | 0.0 | 3 | 3 | 0 | 3 | 3 |
| S34 | decrease in tourism pressure | 6.8 | 0 | 2 | 2 | 0 | 20 |
| S35 | increase in water discharges | 6.8 | 0 | 2 | 3 | 1 | 21 |
| S36 | decrease in water diversions | 6.8 | 0 | 0 | 2 | 1 | 24 |
| S37 | water level fluctuations = 10 cm | 10.0 | 0 | 2 | 2 | 1 | 20 |
| S38 | water level fluctuations = 15 cm | 15.0 | 0 | 2 | 2 | 1 | 25 |
| S39 | water level fluctuations = 20 cm | 20.0 | 0 | 2 | 2 | 1 | 25 |
| S40 | water level fluctuations = 25 cm | 25.0 | 0 | 2 | 2 | 1 | 25 |
| S41 | water level fluctuations = 30 cm | 30.0 | 0 | 2 | 2 | 1 | 29 |
| S42 | S35 and S37 together | 10.0 | 0 | 2 | 3 | 1 | 23 |
| S43 | S35 and S41 together | 30.0 | 0 | 2 | 3 | 1 | 31 |
| S44 | S34 and S37 together | 10.0 | 0 | 2 | 2 | 0 | 21 |
| S45 | best possible scenario | 30.0 | 0 | 0 | 3 | 0 | 34 |
| S46 | S21, S35, and S36 together | 6.8 | 3 | 0 | 3 | 1 | 17 |
| S47 | S25, S35, and S36 together | 5.0 | 0 | 0 | 3 | 1 | 20 |
| S48 | S26, S35, and S36 together | 0.0 | 0 | 0 | 3 | 1 | 11 |
| S49 | S22, S24, S35, and S41 together | 30.0 | 0 | 3 | 3 | 3 | 22 |
| S50 | S21 and S38 together | 15.0 | 3 | 2 | 2 | 1 | 19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrarini, A.; Celada, C.; Gustin, M. Waterbird Species Are Highly Sensitive to Wetland Traits: Simulation-Based Conservation Strategies for the Birds of the Sicilian Wetlands (Italy). Biology 2024, 13, 242. https://doi.org/10.3390/biology13040242
Ferrarini A, Celada C, Gustin M. Waterbird Species Are Highly Sensitive to Wetland Traits: Simulation-Based Conservation Strategies for the Birds of the Sicilian Wetlands (Italy). Biology. 2024; 13(4):242. https://doi.org/10.3390/biology13040242
Chicago/Turabian StyleFerrarini, Alessandro, Claudio Celada, and Marco Gustin. 2024. "Waterbird Species Are Highly Sensitive to Wetland Traits: Simulation-Based Conservation Strategies for the Birds of the Sicilian Wetlands (Italy)" Biology 13, no. 4: 242. https://doi.org/10.3390/biology13040242