
Comparative Study on the Effectiveness of Three Inoculation Methods for Valsa sordida in Populus alba var. pyramidalis

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Fungi
2.2. Punching, Burning, and Toothpick Inoculation Experiments on Six-Year-Old P. alba var. pyramidalis
2.2.1. Punching Inoculation
2.2.2. Burning Inoculation
2.2.3. Toothpick Inoculation
2.3. Burning Inoculation Experiment on Two-Year-Old P. alba var. pyramidalis
2.4. Disease Markers and Statistical Methods
2.4.1. Statistical Analysis and Calculation of Disease Incidence, Disease Severity Index, and Lesion Area at Each Inoculation Site
2.4.2. Observation and Measurement of Lesion Morphology and Spore Microstructure at Inoculation Sites
2.4.3. Observation and Statistical Analysis of Wounded Area of Xylem at Inoculation Sites
2.5. Data Processing and Statistical Analysis
3. Results and Analysis
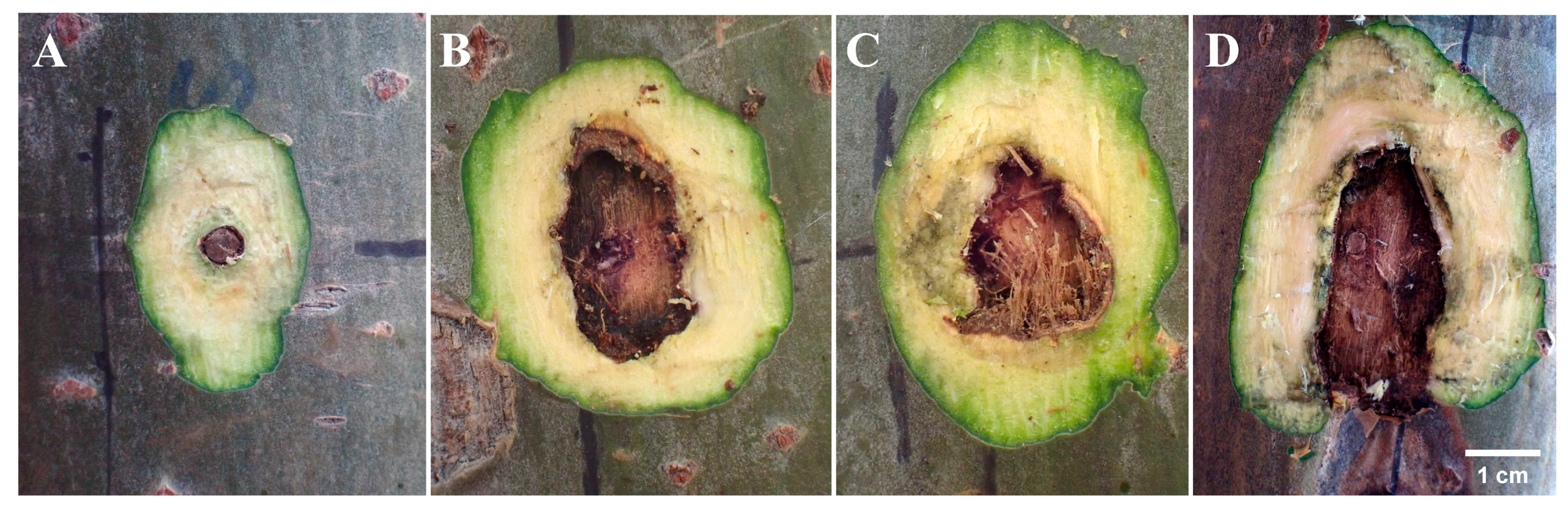
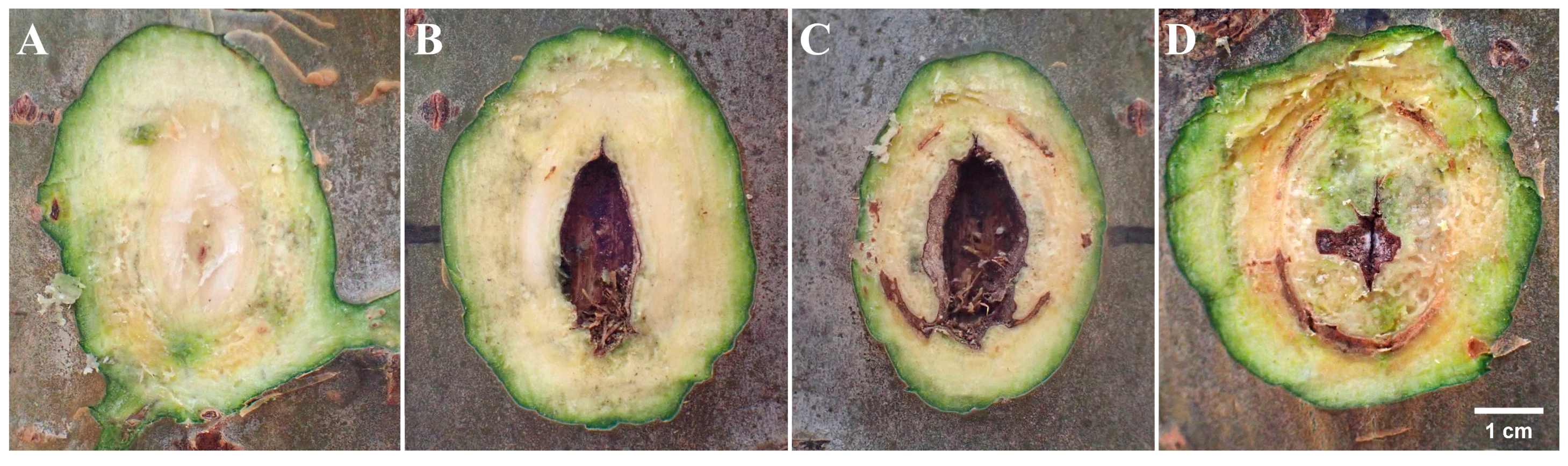
3.1. Comparative Analysis of Symptoms of Lesions at Inoculation Sites Wounded by Punching, Burning, and Toothpick Inoculation Techniques
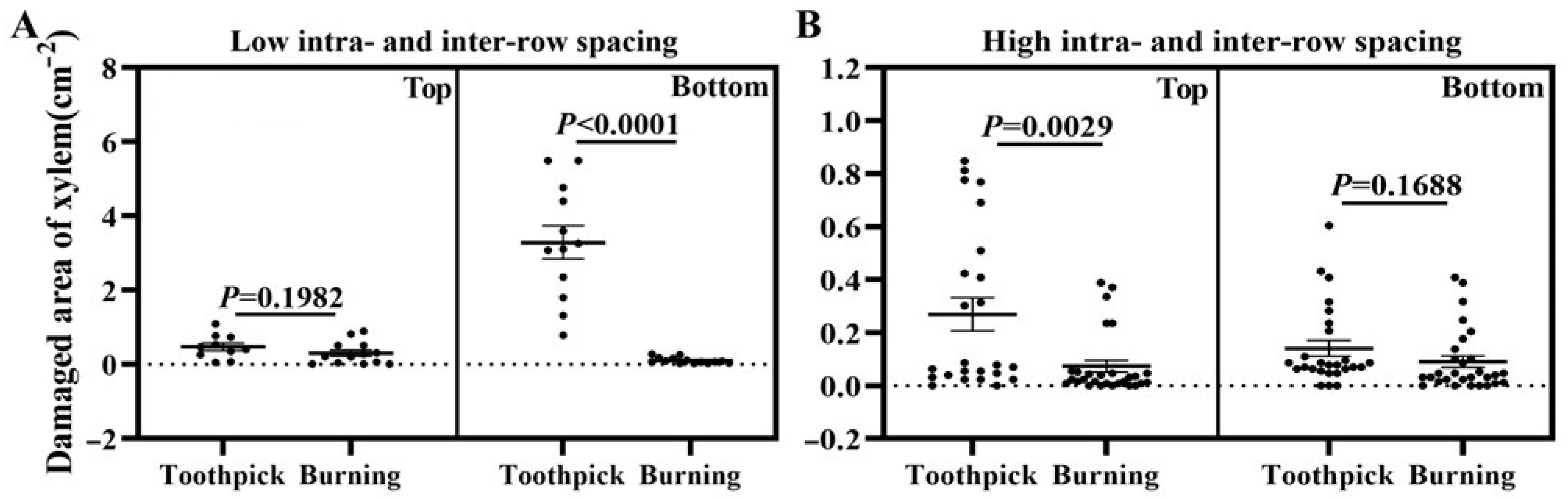
3.2. Phenomenon of Xylem Damage and Statistical Analysis of Xylem Damage Area at Inoculation Sites in Three Inoculation Methods
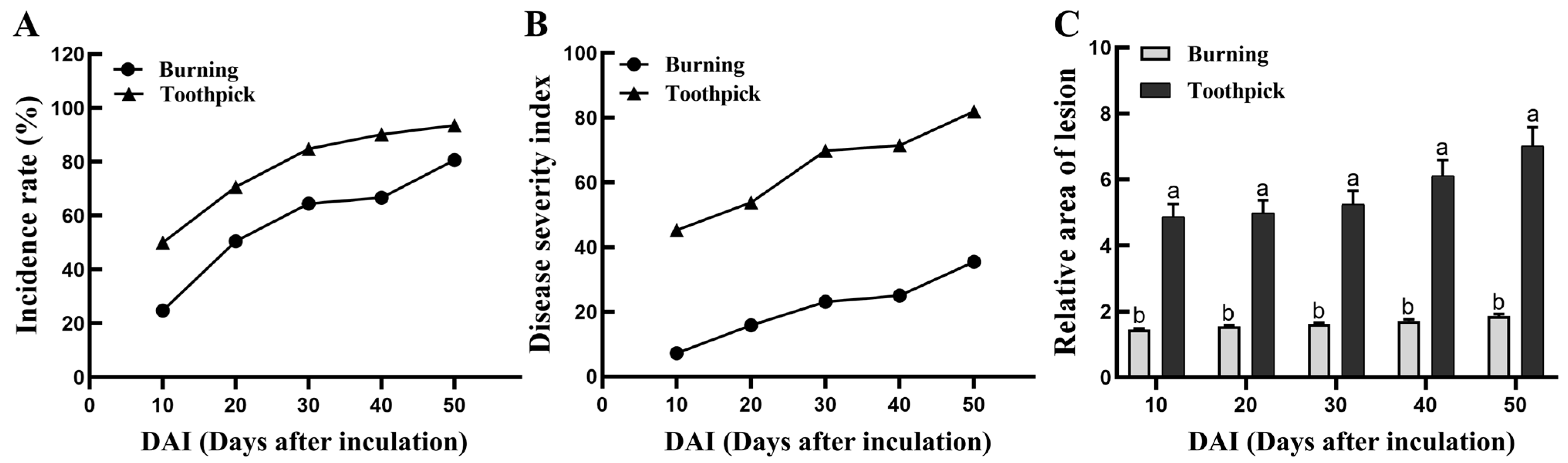
3.3. Comparative Analysis of Disease Incidence, Disease Severity Index, and Relative Area of Lesion at the Inoculation Sites in the Burning and Toothpick Inoculation Groups
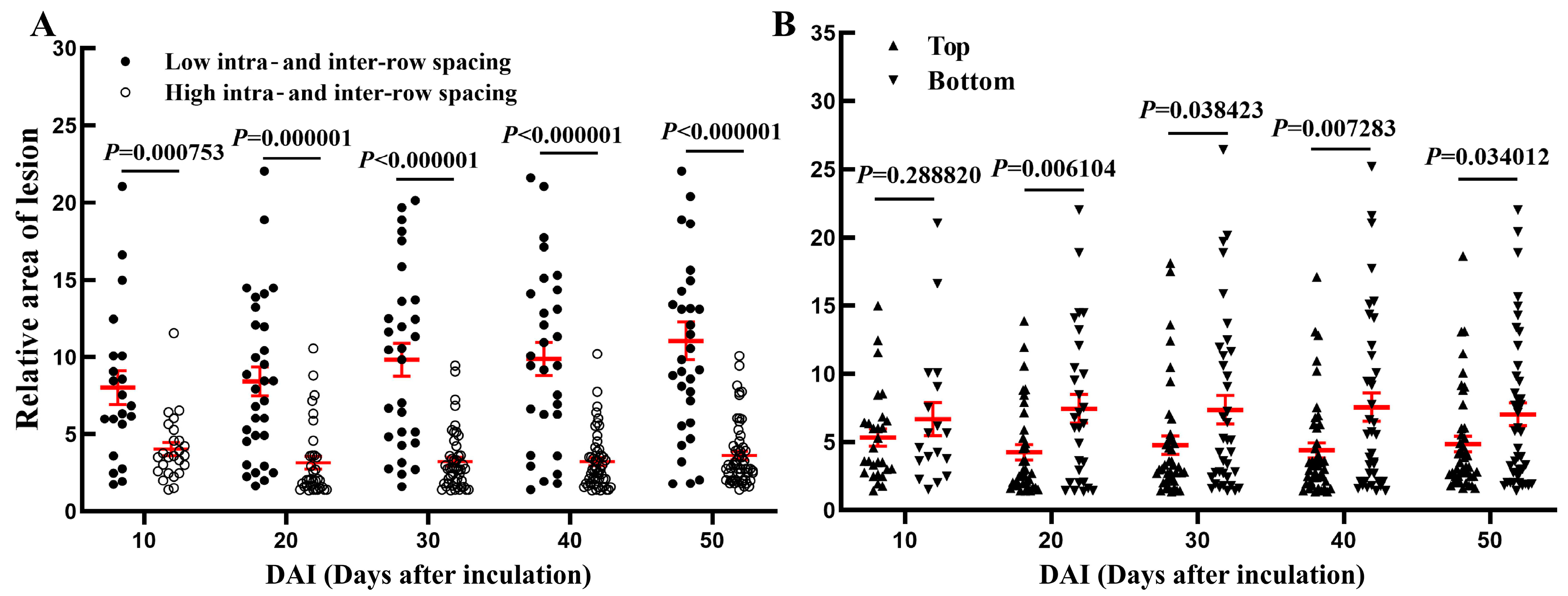
3.4. Effects of Intra- and Inter-Row Spacing as Well as Inoculation Positions on the Outcomes of Toothpick Inoculation
4. Conclusions and Discussion
4.1. Discussion on the Induction of Lesion by Punching, Burning, and Toothpick Inoculation of V. sordida
4.2. Effects of Different Stand Densities and Inoculation Positions on Inoculation Outcomes
4.3. Application of Toothpick Inoculation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, Z.D. Research Methods on Plant Disease; China Agriculture Press: Beijing, China, 1998. [Google Scholar]
- Xu, Z.G. General Plant Pathology; Higher Education Press: Beijing, China, 2009. [Google Scholar]
- Reid, L.M.; Hamilton, R.I.; Mather, D.E. Effect of macroconidial suspension volume and concentration on expression of resistance to Fusarium graminearum in Maize. Plant Dis. 1995, 79, 461–466. [Google Scholar] [CrossRef]
- Ullstrup, A.J. Methods for inoculating corn ears with Gibberella zeae and Diplodia maydis. Plant Dis. 1970, 54, 658–662. [Google Scholar]
- Ye, J.R.; He, W. Forest Pathology; China Forestry Publishing House: Beijing, China, 2011. [Google Scholar]
- Li, P.; Liu, W.; Zhang, Y.; Xing, J.; Li, J.; Feng, J.; Su, X.; Zhao, J. Fungal canker pathogens trigger carbon starvation by inhibiting carbon metabolism in poplar stems. Sci. Rep. 2019, 9, 10111. [Google Scholar] [CrossRef] [PubMed]
- Bessho, H.; Komori, S.; Soejima, J. Simple excised twig assay of Malus species for determining resistance to Valsa canker. Acta Hortic. 2004, 658, 391–397. [Google Scholar] [CrossRef]
- Yang, C.H. A comparative study on different methods of inoculation of poplar canker. J. Bejing For. Univ. 1985, 2, 76–81. [Google Scholar]
- Ghimire, K.; Petrović, K.; Kontz, B.J.; Bradle, C.A.; Chilvers, M.I.; Mueller, D.S.; Smith, D.L.; Wise, K.A.; Mathew, F.M. Inoculation method impacts symptom development associated with Diaporthe aspalathi, D. caulivora, and D. longicolla on soybean (Glycine max). Plant Dis. 2019, 103, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wan, D.; Duan, B.; Bai, X.; Bai, Q.; Chen, N.; Ma, T. Genome sequence and genetic transformation of a widely distributed and cultivated poplar. Plant Biotechnol. J. 2019, 17, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kang, X.; Wang, S.; Wang, S.D.; Dai, L.L.; Chen, H.W. Pollen development and multi-nucleate microspores of Populus bolleana Lauche. For. Stud. China 2008, 10, 107–111. [Google Scholar] [CrossRef]
- Zhang, J.G. Studies on Populus in Xinjiang; Forestry Publishing House: Beijing, China, 2014. [Google Scholar]
- Kepley, J.B.; Reeves, F.B.; Jacobi, W.R.; Adams, G. Species associated with cyospora canker on Populus tremuloides. Mycotaxon 2015, 130, 1124–1131. [Google Scholar] [CrossRef]
- Qu, H.H.; Yang, X.Q.; Jiang, L.Q.; Wang, P.; Yan, P.; Wang, Q.J.; Li, S. Researches of V. sordida Nits. Heilongjiang Agric. Sci. 2012, 5, 145–149. [Google Scholar]
- Abbasi, K.; Abbasi, S.; Forouhifar, K.B.; Zebarjadi, A.R. Study of genetic diversity in Cytospora chrysosperma isolates obtained from walnut trees in Iran using inter simple sequence repeat (ISSR) markers. Arch. Phytopathol. Plant Prot. 2015, 48, 327–335. [Google Scholar] [CrossRef]
- Shao, Y.H.; Ge, W.H.; Qin, X.B.; Wang, J.X.; Su, Y. Relations of Fungus Quantity in Poplar Saplings to V. sordida Infection. J. Shandong For. Sci. Technol. 2003, 6, 22–23. [Google Scholar]
- Wang, Z.M.; Liu, G.R.; Chen, Y.Q. Cytospora canker of main poplar varieties in Jilin and the management. Plant Prot. 2008, 34, 102–106. [Google Scholar]
- Jiao, L.M.; Liang, T.M.; Ji, C.Y.; Li, X.Y.; Zhang, Y. Research on pathogenicity of Cytospora chrysosperma (Pers.) Fr. on Poplar and bacteriostasis of Ambrosia artemisiifolia L. Hunan For. Sci. Technol. 2020, 47, 77–81. [Google Scholar]
- Xing, J.; Li, P.; Zhang, Y.; Li, J.; Liu, Y.; Lachenbruch, B.; Su, X.; Zhao, J. Fungal pathogens of canker disease trigger canopy dieback in poplar saplings by inducing functional failure of the phloem and cambium and carbon starvation in the xylem. Physiol. Mol. Plant Pathol. 2020, 112, 101523. [Google Scholar] [CrossRef]
- Wang, X.L.; Zang, R.; Wang, L.; Kang, Z.S.; Huang, L. The occurrence of Valsa malicola on apple trees and its pathogenicity. Sci. Silvae Sin. 2007, 43, 23–26. [Google Scholar]
- Balendres, M.A.; De, T.R.; Dela, C.F. Culture storage age and fungal re-isolation from host-tissue influence Colletotrichum spp. virulence to pepper fruits. J. Phytopathol. 2019, 167, 510–515. [Google Scholar] [CrossRef]
- Scholthof, K.B. The disease triangle: Pathogens, the environment and society. Nat. Rev. Microbiol. 2007, 5, 152–156. [Google Scholar] [CrossRef]
- Seiwa, K.; Miwa, Y.; Sahashi, N.; Kanno, H.; Tomita, M.; Ueno, N.; Yamazaki, M. Pathogen attack and spatial patterns of juvenile mortality and growth in a temperate tree, Prunus grayana. Can. J. For. Res. 2008, 38, 2445–2454. [Google Scholar] [CrossRef]
- Yamazaki, M.; Iwamoto, S.; Seiwa, K. Distance- and density-dependent seedling mortality caused by several diseases in eight tree species co-occurring in a temperate forest. Plant Ecol. 2009, 201, 181–196. [Google Scholar] [CrossRef]
- Bagchi, R.; Swinfield, T.; Gallery, R.E.; Lewis, O.T.; Gripenberg, S.; Narayan, L.; Freckleton, R.P. Testing the Janzen-Connell mechanism: Pathogens cause overcompensating density dependence in a tropical tree. Ecol. Lett. 2010, 13, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Benítez, M.S.; Hersh, M.H.; Vilgalys, R.; Clark, J.S. Pathogen regulation of plant diversity via effective specialization. Trends Ecol. Evol. 2013, 28, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Ares, A.; Neill, A.R.; Puettmann, K.J. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. For. Ecol. Manag. 2010, 260, 1104–1113. [Google Scholar] [CrossRef]
- Roberts, M.R.; Paul, N.D. Seduced by the dark side: Integrating molecular and ecological perspectives on the influence of light on plant defence against pests and pathogens. New Phytol. 2006, 170, 677–699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhou, G.Y.; Dong, W.; Yang, L.; Wu, Y. Impact of community structure of Dalbergia odorifera plantations on foliage diseases occurrence. Sci. Silvae Sin. 2015, 51, 121–127. [Google Scholar]
- Wang, B.; Tian, C.; Liang, Y. Landscape and stand-scale factors drive the infestation of an endemic fungal pathogen: The role of leaf traits. For. Ecol. Manag. 2022, 514, 120213. [Google Scholar] [CrossRef]
- Patton, R.F. The effect of age upon susceptibility of eastern white pine to infection by Cronartium ribicola. Phytopathology 1961, 51, 429–434. [Google Scholar]
- Ma, Y.C. Occurrence regularity and triggering factor of canker of Carya cathayensis. For. Pest Dis. 2021, 40, 21–25. [Google Scholar]
- Xiang, Y.Y.; Guo, S.G. Study on the Control Index of Dothiorella gregaria Canker of Poplar. For. Res. 1996, 9, 409–412. [Google Scholar]
- Lin, Y.L.; Huang, L.L.; Suolang, L.M.; Gao, X.N.; Chen, Y.C.; Kang, Z.S. A rapid laboratory evaluation system for apple ring rot. Acta Phytophylacica Sin. 2011, 38, 37–41. [Google Scholar]
- Yin, Y.X.; Liu, Y.M.; Ma, R.; Zhao, L.; He, M.R.; Cai, G.F. Laboratory evaluation method of walnut Canker pathogenicity. Chin. Agric. Sci. Bull. 2016, 32, 5. [Google Scholar]
- Yue, C.Y.; Zhang, X.P.; MA, P.P.; Jiao, S.P.; Ke, R.M.; Wang, C.X. Resistance of different poplars in the initial afforestation stages on Cytospora chrysosperma in Xinjiang. J. Northwest For. Univ. 2011, 26, 113–118. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Lesion Area in the Treatment Group/Area of Inoculation Sites in the Control Group (Srelative area) |
|---|---|
| 0 | No infection in the treatment group (Srelative area ≤ 1.25) |
| 1 | The lesion area in the treatment group is 1.25 to 1.75 times that of the control inoculation sites (1.25 < Srelative area ≤ 1.75) |
| 2 | The lesion area in the treatment group is 1.75 to 2.25 times that of control inoculation sites (1.75 < Srelative area ≤ 2.25) |
| 3 | The lesion area in the treatment group is 2.25 to 2.75 times that of the control inoculation sites (2.25 < Srelative area ≤ 2.75) |
| 4 | The lesion area in the treatment group is over 2.75 times that of the control inoculation sites (Srelative area > 2.75) |
| Treatment | Incidence Rate, % | Disease Severity Index | Relative Lesion Area | |
|---|---|---|---|---|
| 1 | CZA | 18.75 | 30.08 | 1.65 ± 0.40 |
| 2 | CZC | 100 | 87.50 | 3.58 ± 1.38 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, W.; Pan, L.; Fu, Y.; Suo, Y.; Zhang, Y.; Liu, H.; Su, X.; Zhao, J. Comparative Study on the Effectiveness of Three Inoculation Methods for Valsa sordida in Populus alba var. pyramidalis. Biology 2024, 13, 251. https://doi.org/10.3390/biology13040251
Shen W, Pan L, Fu Y, Suo Y, Zhang Y, Liu H, Su X, Zhao J. Comparative Study on the Effectiveness of Three Inoculation Methods for Valsa sordida in Populus alba var. pyramidalis. Biology. 2024; 13(4):251. https://doi.org/10.3390/biology13040251
Chicago/Turabian StyleShen, Wanna, Long Pan, Yuchen Fu, Yutian Suo, Yinan Zhang, Huixiang Liu, Xiaohua Su, and Jiaping Zhao. 2024. "Comparative Study on the Effectiveness of Three Inoculation Methods for Valsa sordida in Populus alba var. pyramidalis" Biology 13, no. 4: 251. https://doi.org/10.3390/biology13040251