Simple Summary
Seaweeds and probiotics are commonly utilized as additives in the diet of aquatic animals. However, limited information exists regarding the effects of Gracilaria lichenoides and Bacillus amyloliquefaciens, either alone or in combination, on Penaeus monodon. In our study, we examined the impact of dietary supplementation with G. lichenoides and B. amyloliquefaciens, both individually and in combination, on the growth performance, antioxidant capacity, and intestinal function of P. monodon. Our findings indicate a beneficial influence of dietary G. lichenoides and B. amyloliquefaciens on P. monodon. These findings offer a theoretical foundation for the judicious utilization of G. lichenoides and B. amyloliquefaciens in the cultivation of black tiger shrimp, as well as for future investigations into their metabolic mechanisms.
Abstract
This research sought to assess the effects of dietary supplements with Gracilaria lichenoides and Bacillus amyloliquefaciens, either individually or combined, on the growth performance, antioxidant capacity, and intestinal function of Penaeus monodon. A total of 840 shrimps were randomly assigned to 28 tanks with an average initial weight of (1.04 ± 0.03) g (30 shrimp per tank) with 7 different treatment groups and 4 replicates per treatment. The control treatment (C) consisted of a basal diet; in contrast, the experimental groups were complement with varying levels of G. lichenoides (3% or 8%), either alone (S3 and S8) or in combination with B.amyloliquefaciens at different concentrations (3% G. lichenoides and 109 CFU/g—S3B9; 8% G. lichenoides and 1011 CFU/g B. amyloliquefaciens—S8B11; 109 CFU/g B. amyloliquefaciens—S9; 1011 CFU/g B. amyloliquefaciens—B11). The results indicated that the maximum values of final body weight (FBW) (10.49 ± 0.90) g, weight gain rate (WGR) (908.94 ± 33.58) g, and specific growth rate (SGR) (4.20 ± 0.06) g were perceived in the 3% G. lichenoide diet treatment, and compared with the control group, the difference was significant (p < 0.05). The whole-body lipid content of shrimp in the B9 group was significantly higher than that in the B11 group (p < 0.05), but no significant difference was observed when compared with shrimp fed other diets (p > 0.05). The ash content of shrimp in the B9 group was found to be significantly higher than that in the S3B9 group (p < 0.05). Furthermore, the lipase activity in the stomach and intestines of the experimental groups exhibited a statistically significantly increase compared to the control (p < 0.05). In comparison to the control group, the hepatopancreas of the S3 group exhibited a significant increase in the activities of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), and antioxidant genes [SOD, catalase (CAT), GSH-Px, thioredoxin (Trx), Hippo, and NF-E2-related factor 2 (Nrf2)] expression levels (p < 0.05). Additionally, the activities of total antioxidant capacity (T-AOC), SOD, peroxidase (POD), and antioxidant genes (CAT, GSH-Px, Trx, and Hippo) in the S3B9 treatment of hepatopancreas showed significant improvement (p < 0.05). The inclusion of dietary G. lichenoides and B. amyloliquefaciens resulted in enhanced relative expression of intestinal lipid metabolism genes (fatty acid synthetase (FAS), lipophorin receptor (LR), fatty acid transport protein 1 (FATP1)) and suppressed the expression of the long-chain fatty acid-CoA ligase 4 (LCL4) gene. Analysis of microbiota sequencing indicated improvements in composition and structure, with notable increases in Firmicutes at the phylum level and Vibrio at the genus level in the S3 group, as well as an increase in Tenericutes at the genus level in the S8B11 group. Overall, the inclusion of dietary G. lichenoides and B. amyloliquefaciens positively impacted the growth, antioxidant capacity, and microbial composition of shrimp, with particular enhancement observed in shrimp fed a supplementary 3% G. lichenoides diet.
1. Introduction
Penaeus mondon, also known as black tiger shrimp, is the second largest shrimp species in China, with an annual production of over 100,000 tons [1]. P. monodon, as a traditional cultured shrimp, has the characteristics of strong adaptability, fast growth, strong disease resistance, delicious flavor, high nutritional value, and resistance to transportation [2]. However, diseases are still the most important constraints affecting the development of shrimp culture. Also, water quality deterioration and serious pollution leads to an increase of the pathogenic microorganisms. Antibiotic use in aquaculture is often applied to shrimp farms to overcome disease and stress problems. In response to these problems, antibiotics are often used in shrimp production and medicine. To combat the adverse effects of antibiotics, researchers have been looking for alternatives. Additionally, consumers demand growing levels of quality and safety for shrimp farming, as well as an absence of antibiotics and pollutants. As a result, shrimp farming strategies must accelerate the search for safe dietary supplements and additives that enhance their health, vital activities, and environmental resilience. Antioxidants and microbes added to feed have been considered a non-antibiotic approach that can improve health and performance [3].
There are several efforts underway to develop products or methods for treating or preventing shrimp farm stress [4]. Macroalgae (seaweeds) are rich in bioactive compounds that can potentially be exploited as functional ingredients in feed additives or supplements for animal health applications [5]. Gracilaria lichenoides, belonging to the division Rhodophyta, is an important aquaculture species with good bioremediation potential due to their fast growth rate, high nutrient uptake capacity, and high temperature tolerance [6,7]. A significant portion of the lipid content of seaweeds comes from polyunsaturated fatty acids (PUFAs), especially those of tropical origin [8]. Aside from its polysaccharides and dietary fibers, the seaweeds is known to contain incomparable amounts of minerals, vitamins, and polyphenols [9]. The utilization of seaweed as a dietary supplement for fish and shrimp at low concentrations (2.5–10%) has demonstrated beneficial outcomes, such as enhanced growth performance, increased feed utilization [10,11,12,13], and improved immune and stress responses [14,15]. Supplementation of Sciaenops ocellatus feed with Isochrysis galbana extract, resulted in significantly higher survival rates [16]. Feeding Oncorhynchus mykiss Gracilaria pygmaea polysaccharide led to significantly higher final average weight compared to the control group, with feed intake showing a tendency to increase with higher levels of supplementation [17]. Incorporating Gracilaria gracilis into the diet of Sparus aurata enhanced weight gain rate and feed conversion ratio [18]. The incorporation of Eucheuma denticulatum into the diet of Paralichthys olivaceus resulted in notable enhancement in the growth rate and feed efficiency of juvenile fish, with a concurrent increase in the accumulation of dorsal muscle omega-3 polyunsaturated fatty acid in experimental groups [19]. Additionally, feeding Gracilaria persica to Acipenser persicus led to elevated activities of superoxide dismutase (SOD) and catalase (CAT) [20]. Furthermore, supplementation of Sargassum aquifolium in the feed resulted in heightened activities of CAT, SOD, and glutathione (GSH), along with a reduction in malondialdehyde (MDA) content. Notable, SOD and CAT were significantly upregulated in Oreochromis niloticus [21]. Sargassum horneri was found to enhance hepatic total antioxidant capacity (T-AOC), SOD, and CAT activities in Acanthopagrus schlegelii, with a notable upregulation of CAT expression [22]. Similarly, the dietary supplementation of sulphated polysaccharide resulted in a significant increase in hepatic T-AOC, CAT, and SOD activities, as well as marked upregulated Cu2+/ZnSOD, CAT, and glutathione peroxidase (GSH-Px) antioxidant gene expression in Siganus canaliculatus [23]. In contrast, some researchers found that seaweeds and their extracts induced several morphological changes in fish guts, inhibiting digestion and nutrient absorption [24,25,26]. Gracilaria lemaneiformis increased the length and evenness of intestinal microvilli in L. vannamei [27]. Niu et al. [14] found that the addition of Undaria pinnatifida to the feed significantly increased the height and width of folds in shrimp, as well as the lipase activity. Extracts of Amphiroa Fragilissima promoted digestive enzyme (protease and amylase) activities in P. monodon [28].
Probiotics have been defined as “live microorganisms” which work by rebuilding the gastrointestinal trace (GIT)’s native microflora in response to ingestion of food or water [29]. Bacillus amyloliquefaciens, known as probiotic, which is closely related to Bacillus subtilits, can improve the intestinal microbial environment, facilitate aquatic animal growth, improve enzyme activity, and boost nonspecific immune responses [30,31]. In our earlier research, a specific probiotic B. amyloliquefaciens broke away the healthy L. vannamei intestine, which is an endogenous strain that has strong antagonistic effects against seven species of Vibrio spp. pathogens in aquatic animals. Besides, it can also improve the non-specific immune function, relieve oxidative stress, and strengthen the ability of shrimp to resist pathogenic bacterial infections. Similar results were also found when a strain of B. amyloliquefaciens was isolated from the intestine of healthy P. clarkia, exhibiting strong antimicrobial activity against different pathogens. B. amyloliquefaciens as a feed additive could improve the intestinal digestive enzyme activity, the innate immune enzyme activities, and the antiviral resistance of P. clarkia [32]. Bacillus also has beneficial effects on the microbial community and the development of L. vannamei. Wang et al. explained the effects of Bacillus (108~109 CFU/g) on the survival rate and molting of L. vannamei larvae [33]. B. amyloliquefaciens significantly improved the WG, SGR, and FCR of O. niloticus [34]. B. amyloliquefaciens LSG2-8 added to Rhynchocypris lagowskii feed significantly improved FBM, WGR, SGR, and FCR growth indices [35]. L. vannamei phenol oxidase activity, phagocytic activity, and scavenging efficiency were significantly increased by feeding higher concentrations of Bacillus subtilis E20 [36]. B. amyloliquefaciens R8 caused a significant increase in decreased mRNA expression of oxidative stress-related genes (SOD, Gpx, and Hsp70) in zebrafish liver [37].
The dietary role of G. lichenoides and B. amyloliquefaciens, either alone or in combination, on digestive and metabolic capacity regulatory in P. monodon remains uncertain. The potential benefits of incorporating G. lichenoides and B. amyloliquefaciens into shrimp diets were previously unidentified. Therefore, this study examined the impact of G. lichenoides and B. amyloliquefaciens, either individually or in combination, on the growth, antioxidant capacity, and intestinal health of P. mondon. The aim was to identify the most effective dosage and application method of G. lichenoides and B. amyloliquefaciens in the diet of P. mondon, optimize feed formulation, and enhance feed utilization efficiency and antioxidant stress response in P. mondon. Further, the effects of G. lichenoides and B. amyloliquefaciens on the intestinal microbiota and overall intestinal health of P. mondon was explored, with the goal of establishing a theoretical framework for research in black tiger shrimp aquaculture and elucidating their metabolic mechanisms.
2. Materials and Methods
2.1. Ethical Statement
This animal study was conducted under animal use protocol number NHDF2023-12 and was approved by the Laboratory Animal Welfare and Ethics Committee of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences.
2.2. Experimental Diets Preparation and Nutritional Composition
Basic feed ingredients were purchased from Shandong Wuzhoufeng Agricultural Science and Technology Co., Ltd (Heze, China). G. lichenoide was purchased from the breeding farm of Xigao Village, Nanri Town, Xiuyu District, Putian City, Fujian Province. B. amyloliquefacients was supplied by our laboratory, isolated from the intestinal tract of healthy L. vannamei (Patent No.: ZL201410319443.7). Seven experimental diets were prepared based on the basal diet of black tiger shrimp. The basal diet served as the control, while the other diets included the addition of 3% G. lichenoide as S3, the addition of 3% G. lichenoide and 109 CFU/g B. amyloliquefacients as S3B9, the addition of 109 CFU/g B.amyloliquefacients as S9, the addition of 8% G. lichenoide as S8, the addition of 8% G. lichenoide and 1011 CFU/g B. amyloliquefacients as S8B11, and the addition of 1011 CFU/g B. amyloliquefacients as B11. The feed formulation and composition are analyzed in Table 1. Feed preparation was done according to the method of feed preparation in our laboratory [38]. The feed was made into pellets with diameters of 1.0 mm and 1.5 mm and were heated in an oven at 90 °C for 90 min, and then air-dried to about 10% moisture and stored in a −20 °C freezer. Moisture, crude protein, crude lipid, and ash of the experimental diets were determined using standard methods [39].

Table 1.
Composition and nutrient contents of experimental diets (%).
2.3. Shrimp and Culture System
Juvenile black tiger shrimp were obtained from the Shenzhen Experimental Base of the South China Sea Fisheries Research Institute of the Chinese Academy of Fishery Sciences. Shrimp were domesticated with commercial feed for one week before the beginning of the experiment. Shrimp with an initial average weight of (1.04 ± 0.03) g were randomly assigned to 28 fiberglass tanks (500 L, 0.5 m2 round bottom area, 30 shrimp per tank) in 7 groups of 4 tanks each. During the feeding period, shrimp were fed at 8:00, 17:00, and 22:00 every day, and the daily feeding regimen consisted of providing shrimp with an amount equivalent to 5% of their body weight, closing the screen after each feeding to prevent escape, and adjusting the feeding quantity promptly based on shrimp behavior and prevailing weather conditions. After 1 h of feeding, the remaining shrimp feed in the bait dish was collected, rinsed 2 to 3 times with water, dried at 60 °C, and weighed. The numbers of shrimp deaths and body weight of each tank were recorded. Oxygen was connected to the oxygen supply system by a hose air stone, which was filled day and night. The breeding water was filtered seawater. The rearing water was siphoned off the bottom every day, and 1/3 of the rearing water was changed on a weekly basis. During the experiment, water parameters were maintained as follows: temperature 29.7 ± 0.9 °C, pH 8.0 ± 0.1, salinity 30, dissolved oxygen more than 6.0 mg/L. The feeding trial lasted for 56 days.
2.4. Sampling and Preservation
At the end of the feeding trial, shrimp were fasted for 24 h, and then weighed to determine growth performance. Five shrimp from each tank were randomly selected for whole-body composition analysis. Additionally, five shrimp from each tank were sampled for analysis of enzyme activity and gene expression in the hepatopancreas and intestines. The intestines from two shrimp per tank were extracted at 56 d and stored in Davidson’s solution for 24 h. The intestines from five shrimp per tank were extracted and quickly frozen in liquid nitrogen, followed by storage at −80 °C. To reduce the impact of interindividual differences, intestines from each treatment were mixed for the analysis of the intestinal microbiota.
2.5. Measurement and Analysis
2.5.1. Growth Performance
The growth parameters were calculated according to the following formulas:
Weight gain rate (WGR, %) = 100 × [FBW (g) − IBW (g)]/IBW (g)
Specific growth rate (SGR, %) = 100 × [ln(FBW) − ln(IBW)]/56 days
Feed conversion ratio (FCR) = 100 × dry feed intake (g)/body weight gain (g)
Survival rate (SR, %) = 100 × final living numbers (shrimp)/initial numbers (shrimp)
Note: initial weight (IBW), final body weight (FBW)
2.5.2. Activity of Physiological and Biochemical Indexes
The hepatopancreas samples were homogenized in sterilized PBS buffer (0.86%, pH = 7.4; 1: 9, w/v) by a handheld homogenizer; then, the samples were centrifuged for 10 min (3000 r/min, 4 °C) and the supernatant was fractionated [40]. Hepatopancreas T-AOC, SOD, CAT, GSH-Px, peroxidase (POD) activities, and MDA and total protein contents were determined according to the kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The lipase activity in the stomach and intestine were also analyzed with commercial test kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.5.3. RNA Extraction and qPCR Measurement
RNA from hepatopancreas and intestine was extracted using TRIzol reagent (QIAGEN Cat: no. 74104, Guangzhou, China) according to the manufacturer’s instructions, and DNase I (QIAGEN Cat: no. 79254) was used to remove contaminated DNA. The primer sequences for the target gene and the internal reference gene EF-1α for real-time PCR are shown in Table 2 [41]. All primer pairs for real-time PCR were synthesized by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China). The mRNA expression levels of the target genes were quantified using the fluorescence quantitative PCR method described in a previous report. Each sample was tested three times using real-time PCR (n = 4). The gene expressions were quantified using the 2-ΔΔct calculation method with an internal reference gene [42].
Table 2.
The primers for real-time fluorescence quantification PCR.
2.5.4. HE Stain of the Intestinal Tissue
Two shrimp from each tank had their intestines sampled and preserved in Davidson’s solution and ethanol. The tissues were then rinsed, dehydrated, and prepared for staining and photography. Eight samples were randomly selected for measuring the height of the intestinal epithelial cell and intestinal folds height of shrimp.
2.5.5. Intestinal Microbiome Analysis
Genomic DNA was extracted from intestinal microbial samples using the OMEGA Soil DNA Kit (M5635-02) (Omega Bio-Tek, Norcross, GA, USA) and stored at −20 °C. The bacterial 16S rRNA genes V3-V4 region was amplified using primers 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′), and the resulting PCR products were purified and combined in equal amounts. After quantifying the individuals, the amplicons were pooled and sequenced using the Illumina NovaSeq platform with the NovaSeq 6000 SP Reagent Kit (500 cycles) at Suzhou PANOMIX Biomedical Tech Co., Ltd (Suzhou, China). Microbiota sequencing and analysis were conducted at the same facility. Microbiota sequencing and subsequent analyses were performed at Suzhou Panomix Biomedical Tech Co., Ltd (Suzhou, China).
2.6. Statistical Analysis
The mean ± SD of the data was presented. Statistical analysis was conducted using SPSS 26.0 software (IBM Corporation, Somers, NY, USA) for Windows. Prior to performing ANOVA, the normality and homogeneity of the data were tested using the Kolmogorov-Smirnov test and Levene’s test, respectively. Once the data passed the tests, a one-way ANOVA was performed. If there were notable differences, the group means were compared using Duncan’s multiple-range test, with a significance level of p < 0.05.
3. Results
3.1. Growth Performance
Growth performance of P. monodon is presented in Table 3. There were no significant differences in IBW, FCR, and SR among the groups (p > 0.05). However, shrimp fed diets with 3% G. lichenoide had significantly higher FBW, WGR, and SGR compared to the control group (p < 0.05).
Table 3.
Effects of different diet treatments on growth performance of shrimp after feeding for 56 days (n = 4).
3.2. Whole-Body Composition
Table 4 shows the composition of shrimps’ whole-body composition on different diets. There were no significant differences in moisture and protein content among the diet treatments (p > 0.05). The lipid content of shrimp in the B9 group was higher than that of shrimp in the B11 group (p < 0.05), but not significantly different from shrimp fed other diets (p > 0.05). The ash content of shrimp in the B9 group was higher than that of shrimp in the S3B9 group (p < 0.05).
Table 4.
Body composition of shrimp.
3.3. Change of Antioxidant Enzyme Activities in the Hepatopancreas
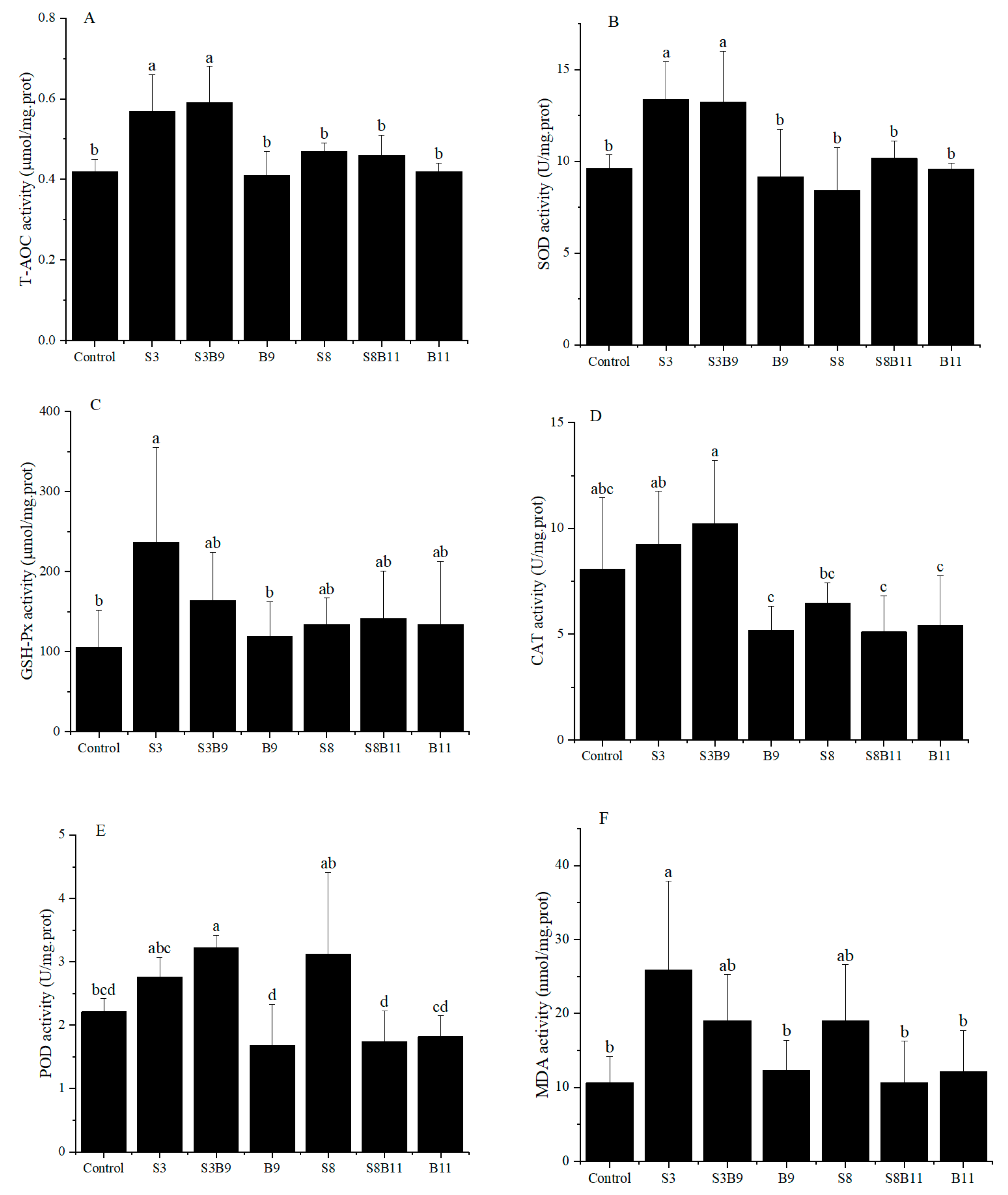
Activities of T-AOC, SOD, GSH-Px, CAT, POD, and MDA content in the hepatopancreas of shrimp for all diet treatments are shown in Figure 1. T-AOC and SOD activities were significantly higher in the S3 and S3B9 groups compared to the other groups (p < 0.05). CAT activity was significantly higher in the S3B9 group compared to the B9, S8B11, and B11 groups (p < 0.05), but not significantly different from the control, S3, and S8 groups (p > 0.05). The GSH-Px activity in shrimp from the S3 group was significantly higher than in the control and B9 groups (p < 0.05), but there was no significant difference with the other groups (p > 0.05). The POD activity in the S3B9 group was significantly higher than in the control, B9, S8B11, and B11 groups (p < 0.05). There was no significant difference in GSH-Px activity among the S3, S3B9, and S8 groups (p > 0.05). The MDA content in shrimp from S3 group was significantly higher than in the control, B9, S8B11, and B11 groups (p < 0.05). The antioxidant capacity in hepatopancreatic tissues of P. monodon was significantly increased in the 3% G. lichenoide group (S3) and the 3% G. lichenoides and 109 CFU/g B. amyloliquefaciens (S3B9).
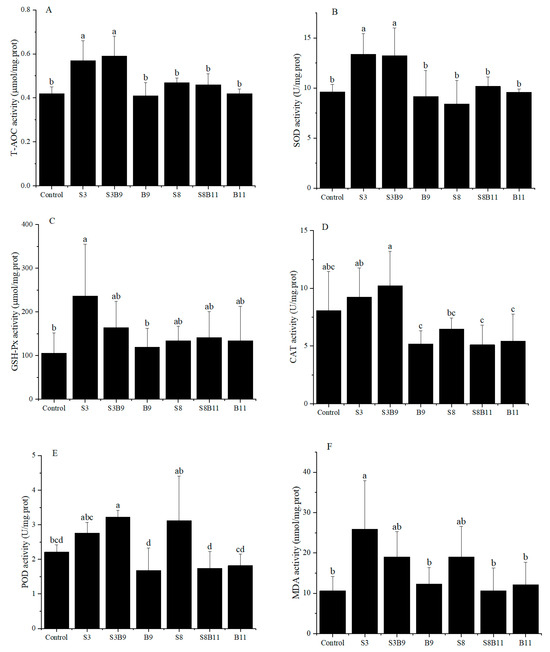
Figure 1.
(A) T-AOC, (B) SOD, (C) CAT, (D) GSH-Px, (E) POD, and (F) MDA content in the hepatopancreas of shrimp fed different diets for 56 days. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were significantly different (p < 0.05) among treatments, the same as below.
3.4. The Relative Expression of Antioxidant Genes in the Hepatopancreas of Shrimp
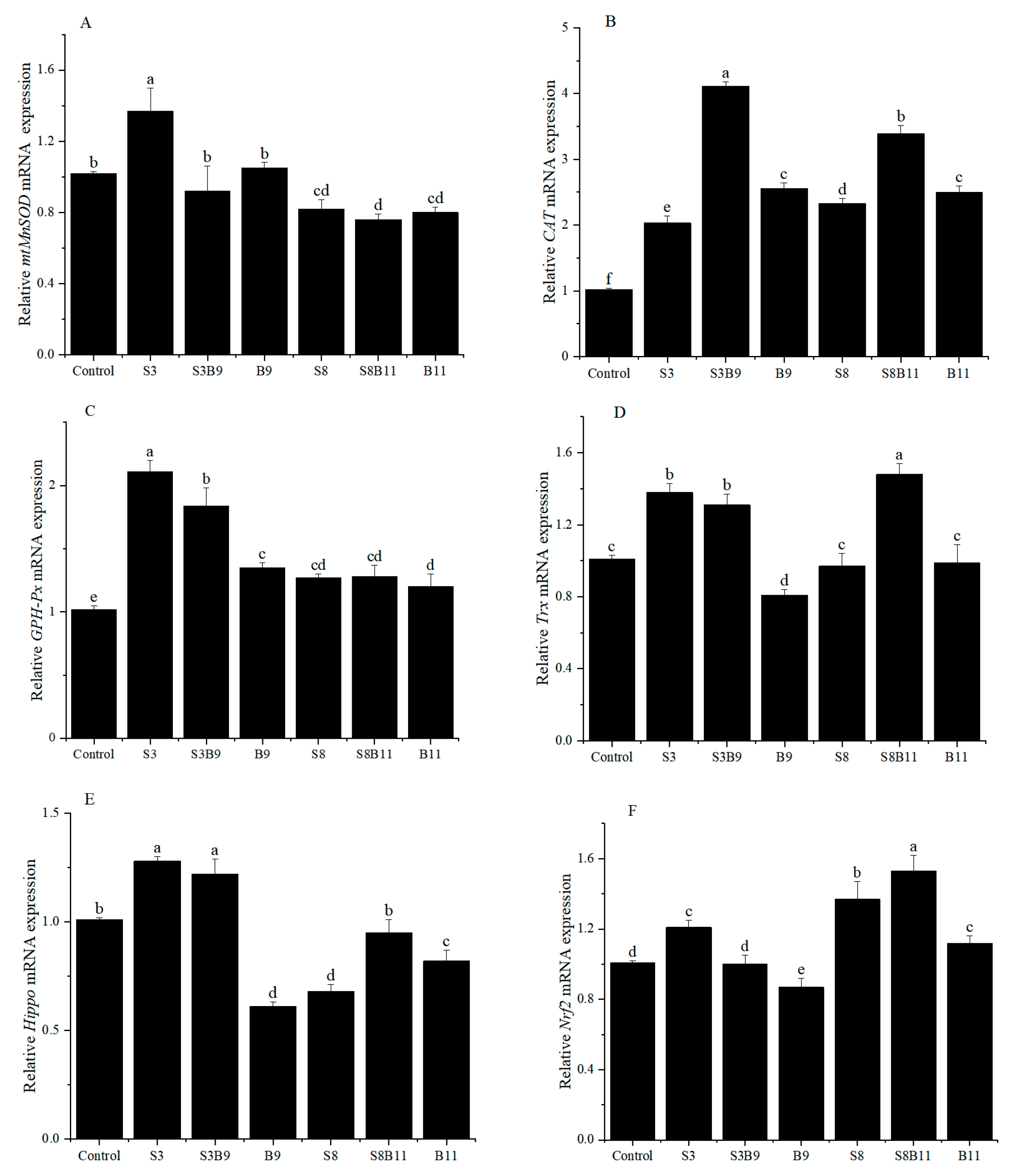
The relative expression of antioxidant genes in the hepatopancreas are showed in Figure 2. The relative expression level of mtMnSOD in the S3 group was significantly higher than that of shrimp in other groups (p < 0.05). However, the relative expression of SOD in the S8, S8B11, and B11 groups was significantly lower than that of shrimp in the control group (p < 0.05). Regardless of the supplementation of G. lichenoides and B. amyloliquefaciens, either individually or in combination in the diet, the relative expressions of CAT and GSH-Px in the hepatopancreas were significantly increased compared to the control group (p < 0.05). Furthermore, the CAT and GSH-Px genes had the highest expression in the S3B9 and S3 groups, respectively. The expression of Trx in shrimp increased significantly in the S3, S3B9, and S8B11 groups compared to the control (p < 0.05), with the highest levels observed in the S3 group. The expression of Hippo in shrimp was significantly higher in the S3 and S3B9 groups compared to the other groups (p < 0.05). The Nrf2 gene relative expression in shrimp was significantly higher in the S3, S8, and S8B11 groups compared to other groups (p < 0.05), with the highest levels observed in the S8B11 group. The results indicate a significant increase in antioxidant gene expression in the hepatopancreas of shrimp between the 3% G. lichenoide group and the 3% G. lichenoide + 109 CFU/g B. amyloliquefaciens group.
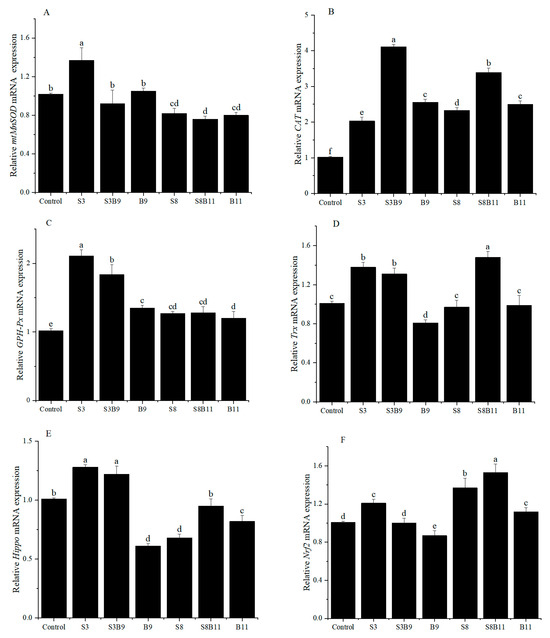
Figure 2.
Expression levels of antioxidant genes in the hepatopancreas of shrimp fed different diets for 56 days. (A) mtMnSOD, (B) CAT, (C) GSH-Px, (D) Trx, (E) Hippo, (F) Nrf2. The reference gene is EF-1α. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were sig-nificantly different (p < 0.05) among treatments.
3.5. Lipase Activity
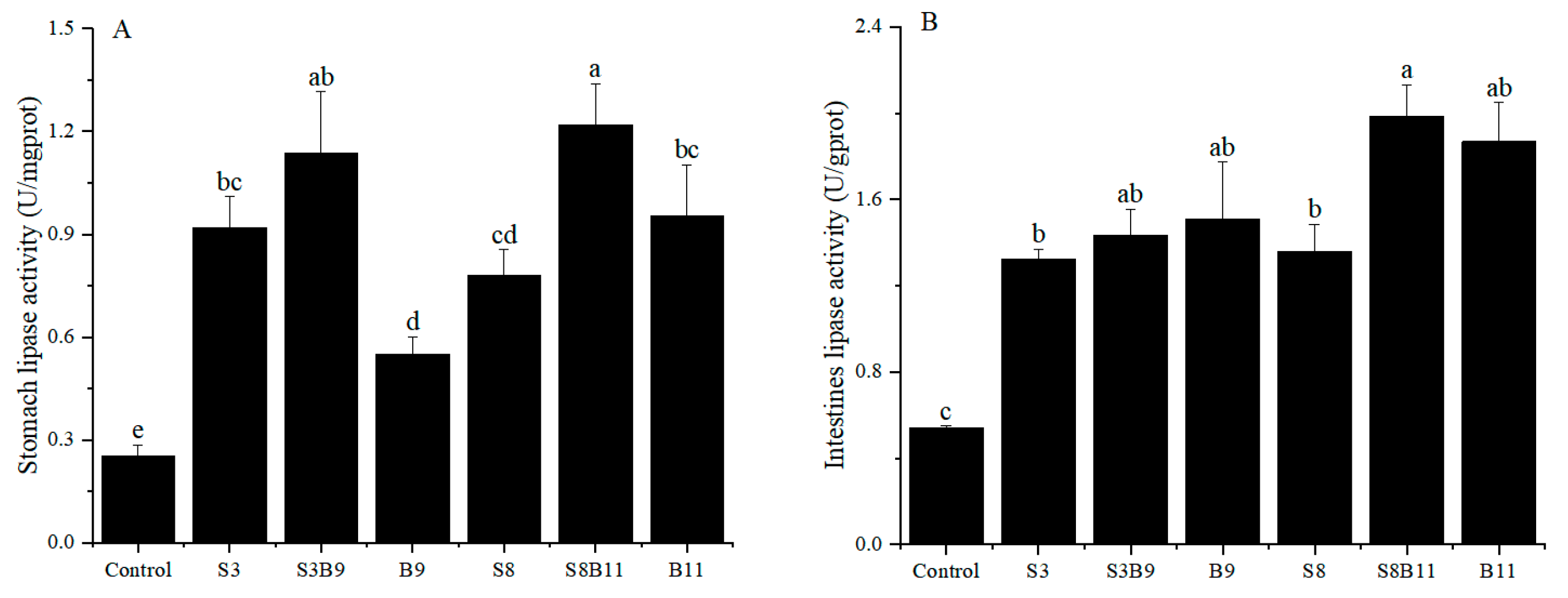
The results of stomach and intestinal lipase activity are shown in Figure 3. The lipase activity of intestine and stomach in the experimental groups were significantly higher than those of the control group (p < 0.05). Results showed that supplementing the diet with the G. lichenoide and B. amyloliquefacients improved the stomach and intestinal lipase activity of P. monodon.
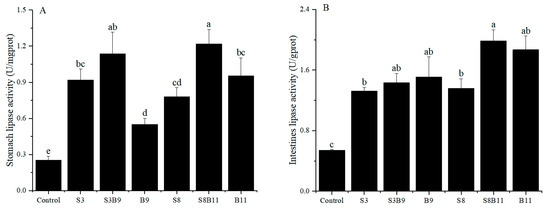
Figure 3.
Lipase activity in the stomach and intestine of shrimp fed different diets for 56 days. (A) Lipase activity of P. monodon in the stomach. (B) Lipase activity of P. monodon in the intestine. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were significantly different (p < 0.05) among treatments.
3.6. Intestinal Tissue Structure
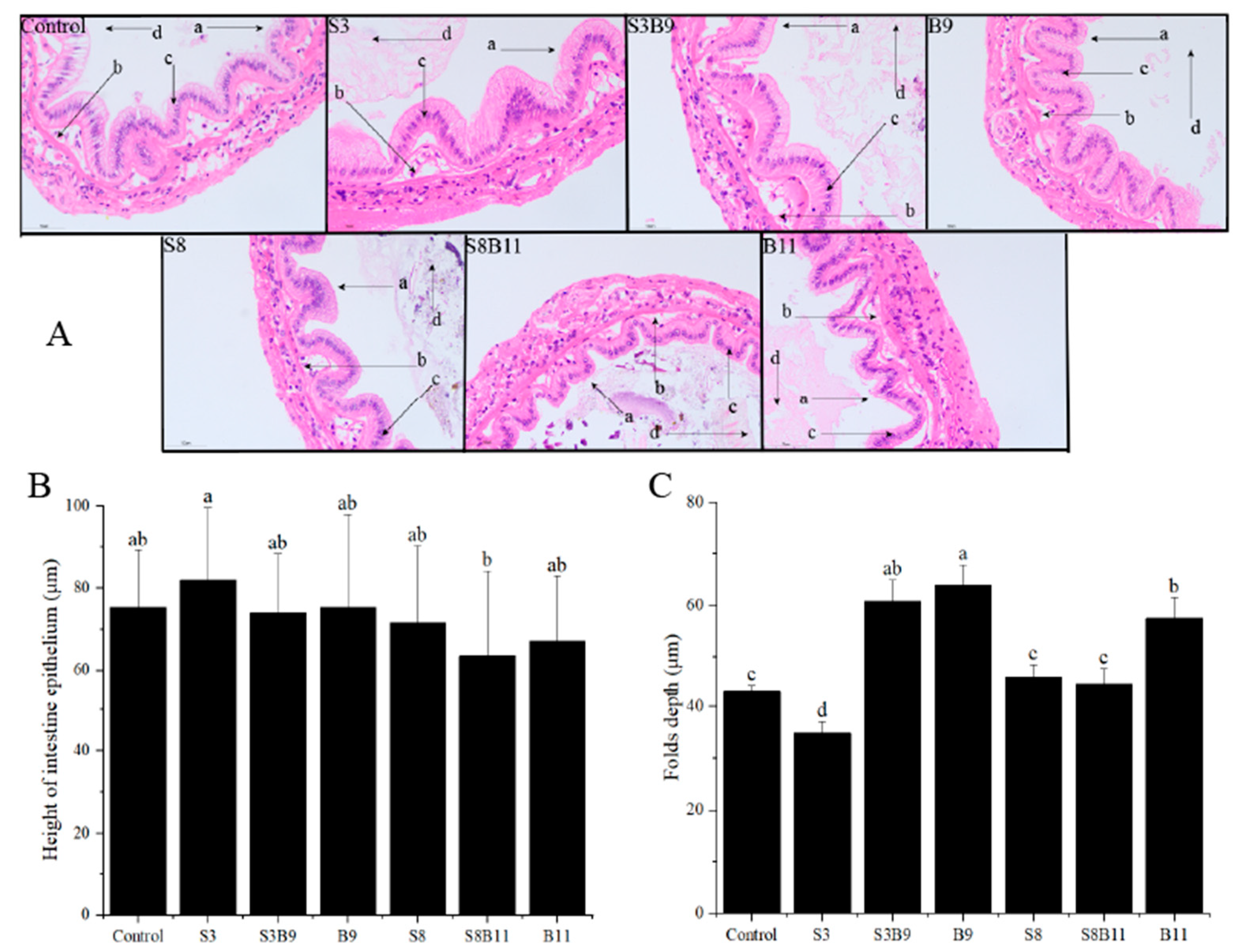
After staining with HE dye, the intestinal tissue of P. monodon showed a higher number of epithelium and neatly arranged brush borders in the experimental group compared to the control group (Figure 4). Vacuoles were almost absent and nuclei in S3 and S8 groups were closely arranged. The height of intestine epithelium in the S3 group was significantly higher than in the S8B11 group (p < 0.05). The folds depth in the S3B9, B9, and B11 groups was also significantly higher than in other groups (p < 0.05). Overall, supplementing the diet with B. amyloliquefaciens (109 CFU/g) improved the intestinal tissue structure of P. monodon.
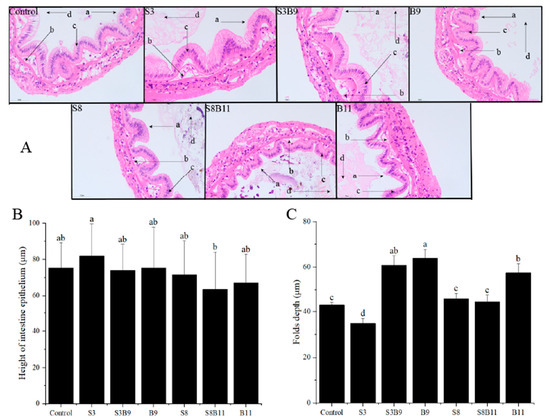
Figure 4.
Intestinal structure of P. monodon fed the different experimental diets for 56 days. (A) Intestine sections of shrimp fed different diets, ×400. (B) Height of intestine epithelium. (C) Folds depth of intestine in shrimp fed different diets. The letters in figure (A) indicate: (a) brush border, (b) epithelium, (c) nuclei, (d) lumen. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were significantly different (p < 0.05) among treatments.
3.7. Intestinal Lipid Metabolism Genes Expression
The expression of lipid metabolism genes in the intestine is illustrated in Figure 5. The relative expression levels of fatty acid FAS and LR genes in the shrimp intestine was significantly higher in the groups supplemented with G. lichenoides and B. amyloliquefaciens, either individually or in combination, compared to the control group (p < 0.05). The B9 and S8B11 groups had the highest expression levels of FAS and LR genes. The relative expression levels of intestinal fatty acid transport protein 1 (FATP1) were significantly higher in the B9 and S8B11 groups compared to other groups (p < 0.05), while the expression of FATP1 was lower in the S3, S3B9, S8, and B11 groups compared to the control group (p < 0.05). The expression of long-chain fatty acid-CoA ligase 4 (LCL4) was lower in all experimental groups with G. lichenoides and B. amyloliquefaciens supplementation compared to control (p < 0.05). The results reveal that adding B.amyloliquefaciens (109 CFU/g) and 8% G. lichenoide + B. amyloliquefaciens (1011 CFU/g) had a significant impact on the expression of genes involved in shrimp intestinal lipid metabolism.
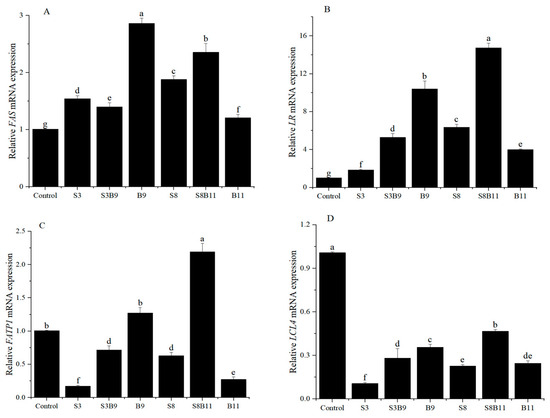
Figure 5.
FAS (A), LR (B), FATP1 (C), and LCL4 (D) gene expression level in the intestine of shrimp fed different diets for 56 days. The reference gene is EF-1α. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were significantly different (p < 0.05) among treatments.
3.8. Intestinal Microbiota Changes
3.8.1. Richness and Diversity
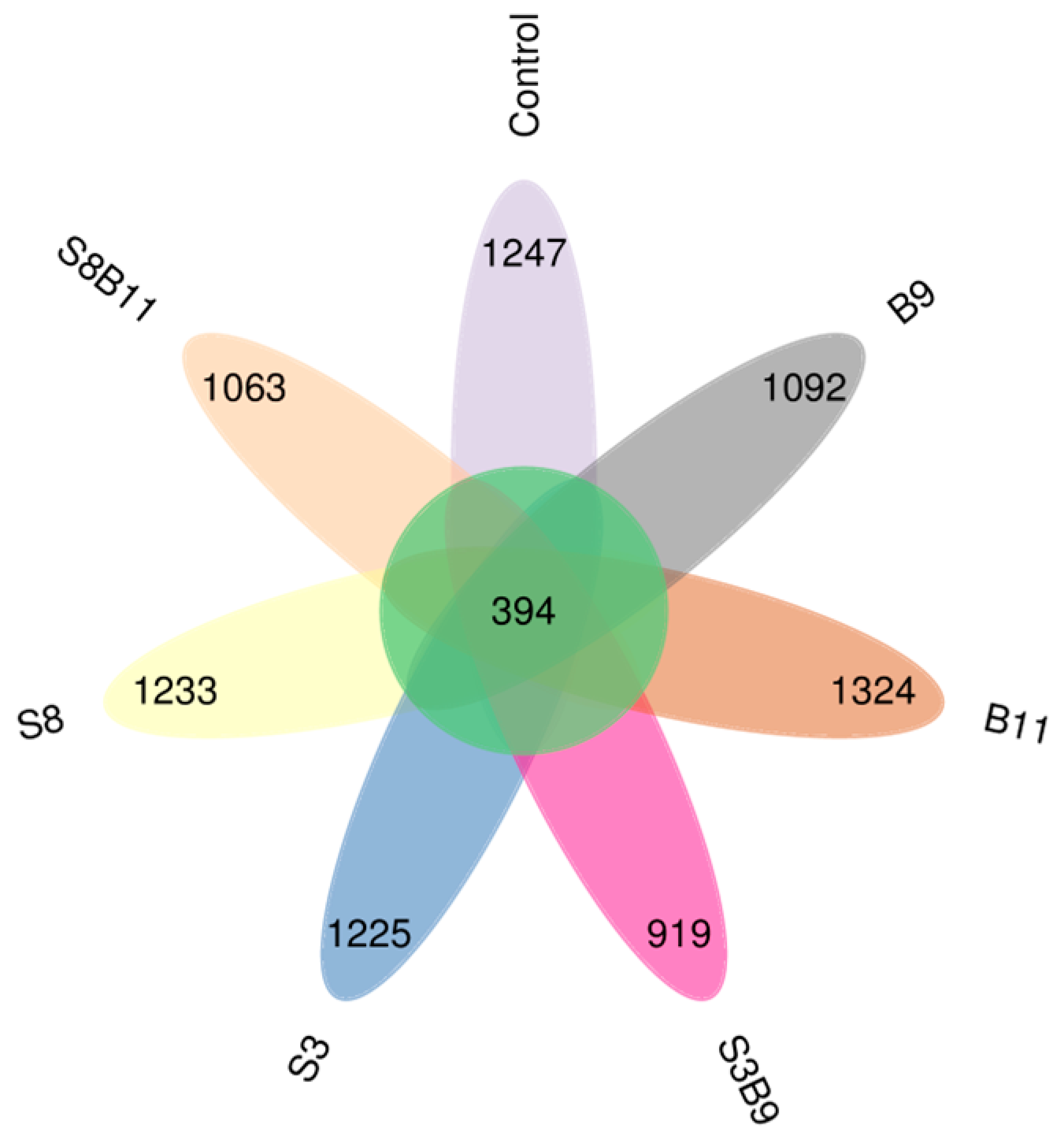
A total of 2,535,118 high-quality sequences were obtained from the intestine microbiota of P. monodon. The sequence length was from 159 to 437 bp. The core OUT (operational taxonomic unit) accounted for 394 (Figure 6). The microbiota coverage of each group was all close to 99%, which suggests that the sequencing depth of this intestinal microbiota analysis was sufficient. Thus, the experimental results were representative. The bacterial richness analysis showed that Chao1 index was increased in the S8, B9 and B11 groups compared with the control group, while there was no significant difference among the groups (p > 0.05) (Table 5).

Figure 6.
Shared OUTs Venn analysis.
Table 5.
Intestinal microbial diversity of P. monodon fed the different experimental diets for 56 days.
3.8.2. Analysis of Microbial Community Composition
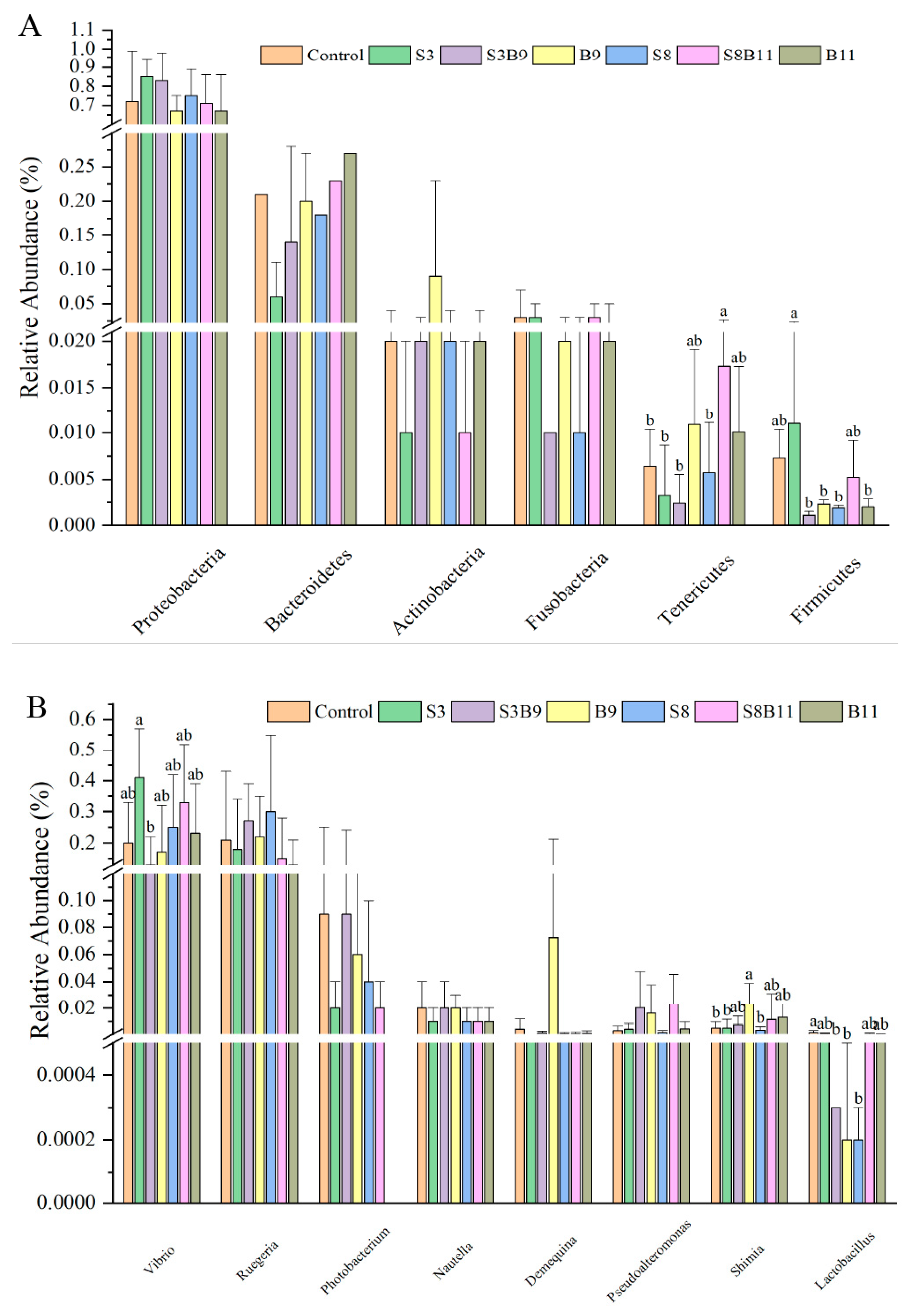
A total of 43 bacterial phyla were identified in the seven groups. Of the dominant phylum, the relative abundances of Tenericutes in the S8B11 groups were increased compared to the control, S3, S3B9, and S8 groups (p < 0.05), yet the relative abundance of Firmicutes in the S3 group was significantly higher than that in the S3B9, B9, S8, and B11 groups (p < 0.05) (Figure 7A). As shown in Figure 7B, at the genus level, the relative abundance of Vibrio in the S3 group were significantly higher than that in the S3B9 (p < 0.05) and had no significant difference with other groups (p > 0.05). Moreover, the relative abundance of Shimia in the B9 group was significantly higher than that of the control, S3, and S8 groups (p < 0.05). However, the relative abundances of Lactobacillus in the S3B9, B9, and S8 groups were significantly lower than that of the control group (p < 0.05).
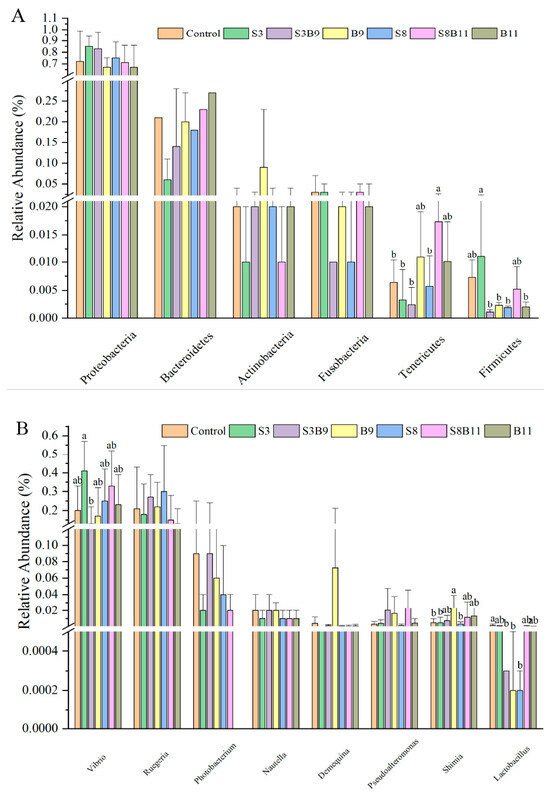
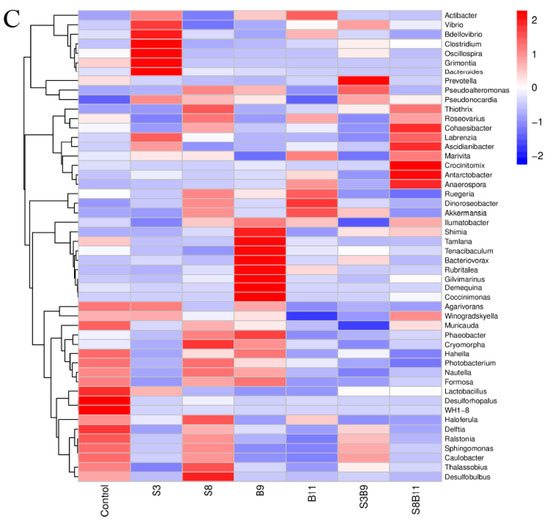
Figure 7.
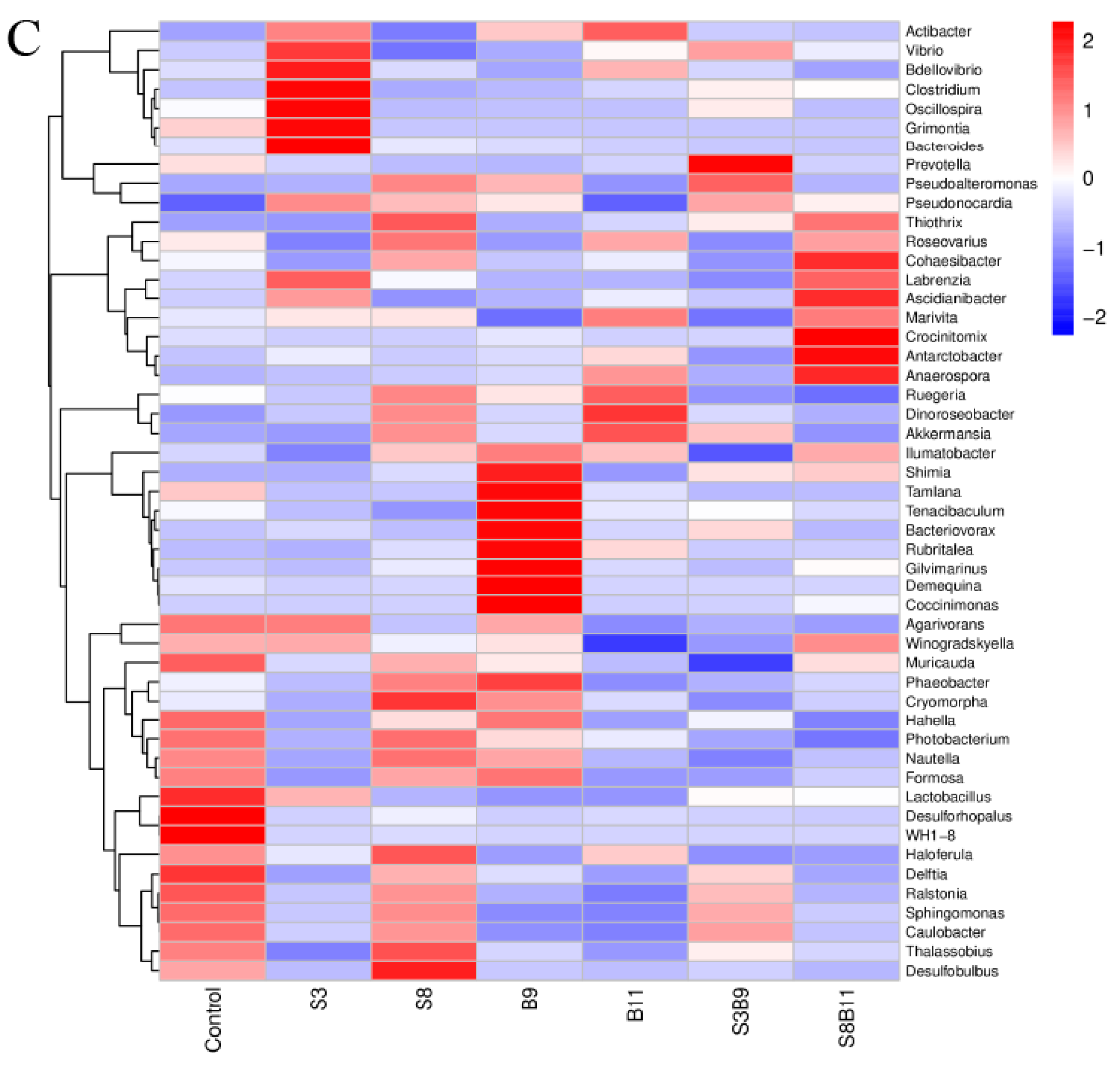
Intestine microbe composition of P. monodon that were fed the different experimental diets for 56 days. (A) Relative abundance of major bacterial phyla in P. monodon fed with the different experimental diets for 56 days. (B) Relative abundance of major bacterial genera in P. monodon fed with the different experimental diets for 56 days. (C) Heat map analysis of intestine microbial on the top 50 genera. Vertical bars represented the mean ± SD (n = 4) and data indicated with different letters were significantly different (p < 0.05) among treatments.
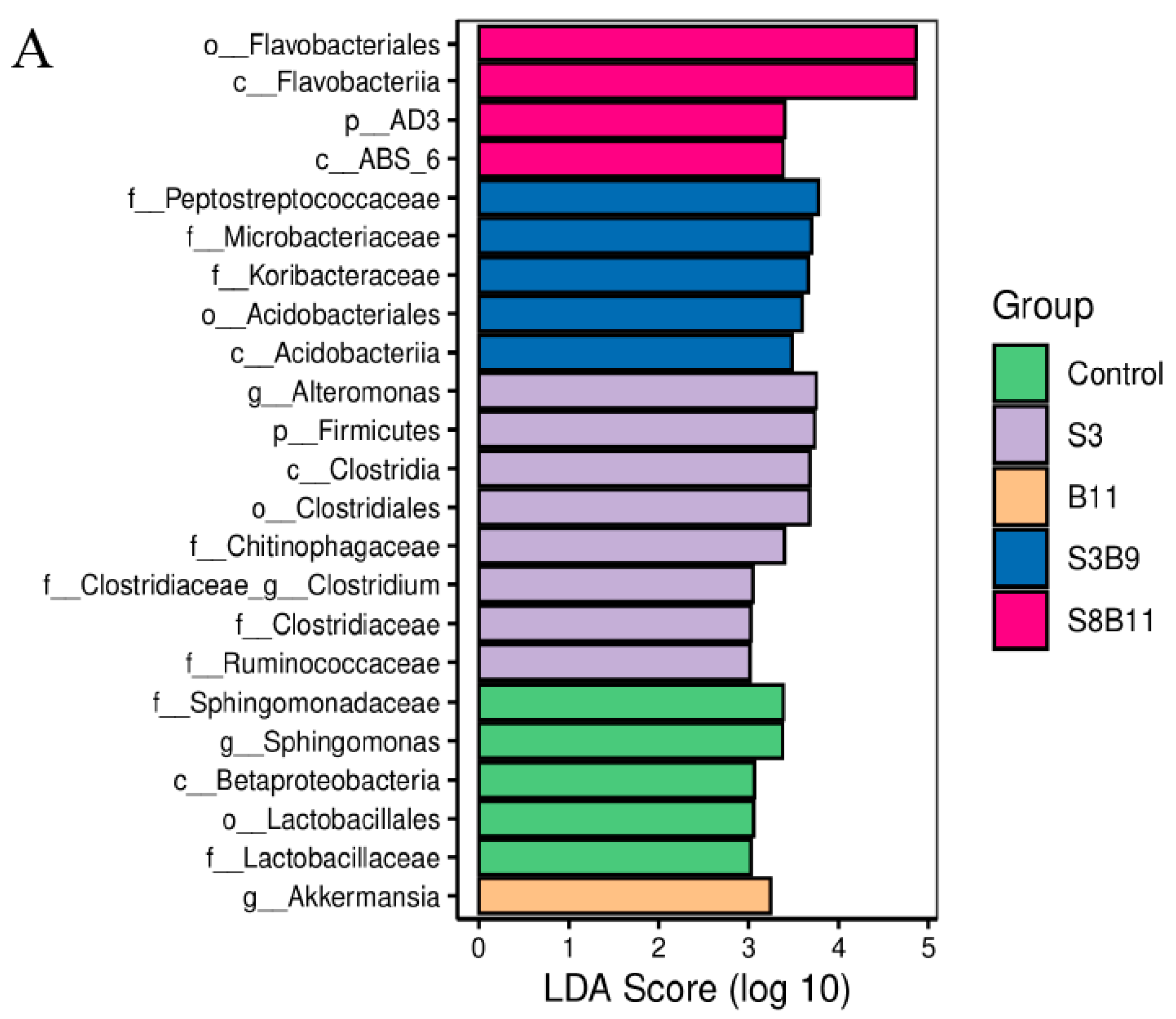
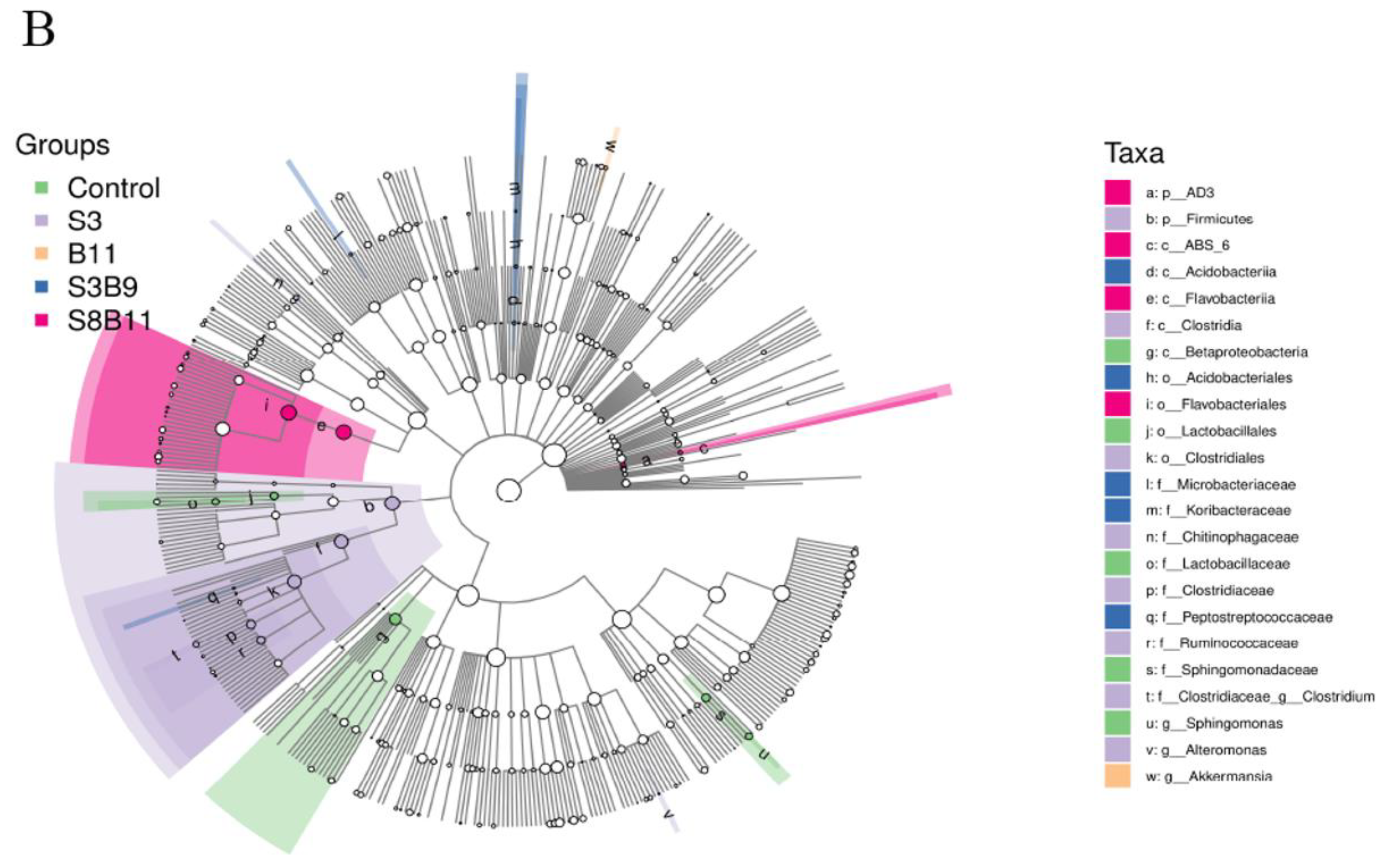
The linear discriminant analysis (LDA) effect size (Lefse) package was used to determine the differential abundances of microbiota taxa among the groups. There are 23 bacteria taxa distinguishing among the experimental groups, except the S8 and B9 groups. The relative abundance of 5, 8, 1, 5, and 4 biomarkers (LDA score > 2.0) was higher in the control, S3, B11, S3B9, and S8B11 groups, respectively. LDA scores of Lefse showed that 8 taxa were increased in the S3 group and there were two groups only enriched at genus level, including Altomonas and Clostridium (Figure 7A). From the results of Lefse cladogram (Figure 8B), 1 phylum, 1 class, 1 order, 4 families, and 2 genera were enriched in the S3, including Firmicutes (from phylum to genus), while 1 phylum, 2 class, and 1 order were enriched in the S8B11 group, including Flavobacteriia (from class to order). At the order level, Acidobacteriales is the possible biomarker for the S3B9 group, while Lactrobacillales was found at the control group. However, no suitable biomarker was found in the S8 and B9 groups.
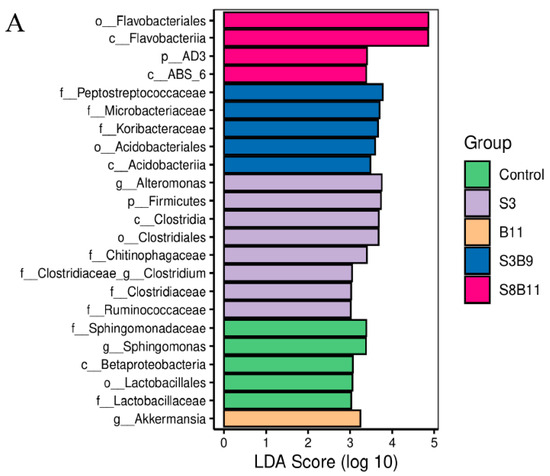
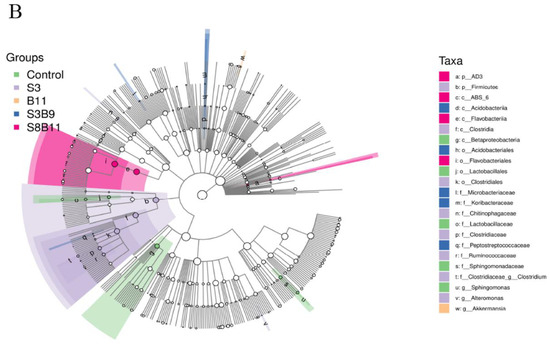
Figure 8.
Indicator species analysis of intestinal microbiota of P. monodon fed the different experimental diets for 56 days. (A) LDA score of Lefse-PICRUSt. The length of the column represents the effect size of bacterial lineages. (B) Lefse cladogram. The cladogram was obtained by mapping differences onto a known hierarchical classification tree. Green: bacterial taxa enriched in the control; Purple: bacterial taxa enriched in the S3 group; Orange: bacterial taxa enriched in the B11 group; Blue: bacterial taxa enriched in the S3B9 group; Pink: bacterial taxa enriched in the S8B11; White: no significant differences. Only the taxa that had a linear discriminate analysis (LDA) value above 2.0 are shown.
4. Discussion
The findings of this study demonstrate that the inclusion of a small quantity of seaweed power in the diet resulted in significantly higher FBW, WGR, and SGR in the S3 group compared to the other groups, suggesting a positive impact on the growth of prawns. Niu et al. [14] conducted an experiment in which various concentrations of wakame were administered to spotted prawns, resulting in a significant increase in WG and SGR at the 2% concentration level. Similarly, Jumah et al. [43] supplemented prawn feeds with k-carrageenan, resulting in a significant increase in the FBW, WG, and SGR in the experimental group. Anil et al. [44] demonstrated that the inclusion of Solieriaceae in the diet of shrimp resulted in increased average daily gain and total weight, suggesting that seaweed have the potential to enhance the growth performance of shrimp. The beneficial effects of seaweeds on shrimp growth can be attributed to their rich content of vitamins, mineral contents, polyunsaturated fatty acids, and amino acids [45]. The active compounds have the ability to modulate the composition of bacterial flora, leading to enhanced intestinal function that facilitates the absorption of carbohydrates and fats in the diet [45]. The growth-promoting impact of seaweeds in the diet can be linked to the digestion and absorption of key nutrients in shrimp feed, which is facilitated by the release of hydrolytic enzymes from shrimps’ digestive glands [46]. The results indicate that the S3 group exhibited significantly higher lipase activity compared to the control group, implying that the inclusion of a small quantity of G. lichenoides may enhance feed digestion and absorption in shrimp, leading to improved growth performance.
The process of lipid metabolism plays a crucial role in the growth and development of shrimp. Lipid transport, lipolysis, and lipid synthesis collectively govern lipid metabolism [47]. The synthesis of lipids, specifically triglycerides, necessitates the catalytic activity of FAS within the fatty acid synthesis pathway, thereby exerting a significant influence on lipid synthesis [47,48]. Furthermore, a decrease in adipogenesis is indicated by the downregulation of FAS [49]. FATPs are essential for lipid transport and synthesis, as they exhibit lipo acyl-coenzyme-A synthetase activity and facilitate the transmembrane transport of long-chain fatty acid, ultimately contributing to fat deposition [50]. LRs are integral membrane proteins that facilitate the binding of lipoproteins and their corresponding ligands, thereby facilitating the cellular uptake and metabolism of lipoproteins [51]. LCL, as a superfamily of membrane proteins, catalyze the activation of fatty acids and the formation of acyl-cofactor thioesters, serving as crucial components in both anabolic and catabolic pathways involving fatty acids [52]. In conclusion, upregulation of the genes FAS, FATP, LR, and LCL has been found to be advantageous for lipid synthesis. The findings of this research indicate that the expression of genes related to fat synthesis, specifically FAS, FATP1, and LR, was increased in the intestines of shrimp in the B9 group compared to the B11 group. This difference in gene expression may account for the significantly higher lipid content observed in the whole body of shrimp of the B9 group compared to the B11 group, suggesting a potential dose-response relationship between B. amyloliquefaciens and lipid metabolism. Similar results indicated a rise in muscle and hepatopancreatic fat content in L. vannamei following the addition of B. subtilis in diet [53], as well as a notable increase in the expression levels of genes associated with lipid synthesis in grass carp after the supplementation B. subtilis H2 in diets [54].
The hepatopancreas plays a crucial role as an antioxidant tissue organ in shrimp, as animals generate significant quantities of reactive oxygen species (ROS) under stress conditions, leading to oxidative damage to proteins, lipids, and DNA [49,55]. The measurement of MDA content in tissues and cells serves as an indicator of the level of oxidative damage [56,57]. Following oxidative damage, the antioxidant enzymes within the biological antioxidant enzyme system (SOD, GSH-PX, CAT, POD) work to eliminate excess ROS and safeguard the organism from harm [58]. The magnitude of T-AOC is indicative of the organism’s ability to compensate for external stimuli through its antioxidant enzyme system and non-enzymatic system, as well as its metabolic state of free radicals [59]. This measurement can serve as a reflection of the organism’s overall antioxidant capacity. Therefore, it is imperative to reduce oxidative damage and enhance the antioxidant capacity of the organism. In the current study, there was a significant increase in T-AOC and SOD enzyme activities in the S3 and S3B9 groups, an increase in MDA and GSH-Px enzyme activities in the S3 group, and a significant increase in POD enzyme activity in the S3B9 group. These results align with the findings of Chen et al. [60], who observed a significant increase in GSH-Px and SOD activities in the hepatopancreas of shrimp fed with Rhodotorula mucilaginosa and B. licheniformis. Ghasem et al. [61] demonstrated that the activities of GSH-Px and SOD in Nile tilapia were significantly enhanced following the addition of PHDP and Pediococcus acidilactici to their diet. Similarly, feeding Gracilaria tenuistipitata extract to L. vannamei resulted in an increase in SOD activity [62]. Furthermore, the supplementation of dietary with 1 g/kg of seaweed polysaccharides led to elevated levels of T-AOC, SOD, and GSH-Px in the hemolymph of Fenneropenaeus merguiensis [63]. The observed phenomenon can be ascribed to the abundance of active biological compounds such as polyphenols, glycosides, anthocyanins, tannins, and thiocarbamates in seaweed. These compounds contribute to the high levels of natural antioxidants that effectively scavenge free radicals, stimulate the production of active antioxidant enzymes, and inhibit oxidative enzymes, thereby mitigating the detrimental effects of free radicals [64]. Additionally, the synthesis of extracellular polysaccharides by prebiotic bacteria plays a role in inhibiting ROS and modulating antioxidant-related genes, ultimately enhancing the antioxidant capacity of the system [65]. The antioxidant capabilities of algae have been evidenced in prior research through a variety of mechanisms, including the reduction of hydrogen peroxide radicals and their conversion into oxygen and water [66]. Furthermore, the observed trends in SOD and GSH-Px activities in the hepatopancreas of shrimp were found to align with their respective mRNA levels.
The Inclusion of 3% G. lichenoides in the feed resulted in a significant upregulation of mtMnSOD, GSH-Px, Trx, Hippo, and Nrf2 genes. Furthermore, the addition of 3% G. lichenoides in combination with B. amyloliquefaciens (109 CFU/g) led to an increase in the relative expression levels of CAT, GSH-Px, Trx, and Hippo genes. This study further examined antioxidant-related genes’ expression levels of hepatopancreas in shrimp supplemented with additives, based on the analysis of physiological and biochemical indices. Under conditions of oxidative stress, Nrf2 levels are maintained through a reduction in ubiquitination and degradation, as well as activation of downstream antioxidant factors [67]. Nrf2 is regulated by multiple pathways at various stages, forming a dimer with sMAF proteins and controlling the expression of genes containing antioxidant response elements (ARE), such as GPx, Trx, Prx, TrxR, CAT, and SOD [68]. This process mediates the activation of a diverse array of antioxidant genes through the Keap1-Nrf2- ARE signaling pathway [67,69]. The SOD and CAT genes in zebrafish was found to be significantly increased by addition of G. gracilis [70]. Furthermore, Chen et al. [59] conducted a study which demonstrated that the inclusion of mannan oligosaccharide (MOS) and B. lincheniformis in the diet resulted in a significant upregulation of the expression levels of CAT, GPx, and SOD in L. vannamei. The inclusion of 3% G. lichenoides in the diet led to an upregulation of Nrf2, SOD, and GSH-Px gene expression, aligning with the observed antioxidant enzyme activity levels. These findings indicate that even a modest quantity of algal meal can enhance antioxidant enzymes through the stimulation of gene expression. Curcumin was observed to enhance the activities of antioxidant enzyme SOD, CAT, and GPx by upregulating the expression of Cu/Zn SOD, MnSOD, CAT, and GPx mRNA, which was associated with the Keap1/Nrf2 signaling pathway in grass carp following infection with Aeromonas hydrophila [59]. Keap1 is capable of regulating the expression of antioxidant genes, such as CAT and GSH-Px, through modulation of the Nrf2 signaling pathway. Moreove, a fraction of Keap1 may potentially increase the efficacy of hepatopancreatic antioxidant enzymes by activating genes through the Nrf2 signaling pathway [69]. The findings indicated elevated levels of Trx expression in the S3, S3B9, and S8B11 groups, aligning with previous research demonstrating the ability of aspalathin to mitigate lead-induced renal oxidative stress in mice through modulation of the Trx and Nrf2 signaling pathways [71]. This mechanism involves the binding of Nrf2 to the antioxidant ARE and subsequent upregulation of Trx and TrxR genes via the Trx signaling pathway, ultimately reducing oxidative stress-induced damage [71]. The inclusion of 8% G. lichenoides in combination with B. amyloliquefaciens (1011 CFU/g) in the diet resulted in an upregulation of Nrf2, CAT, and Trx gene levels. However, there was no notable impact on antioxidant enzyme activities, indicating the necessity for further exploration into the specific mechanisms underlying this effect. The study revealed that the supplementation of B. amyloliquefaciens (109 CFU/g) in the diet did not have significant alterations in the antioxidant enzyme activities and expression levels of relevant antioxidant genes in P. monodon. Conversely, the concurrent administration of modest quantities of algal meal and B. amyloliquefaciens induced the activation of the antioxidant response factor Nrf2, thereby facilitating the expression of diverse antioxidant enzymes and enhancing the transcription of downstream genes including Trx, SOD, CAT, and GPx. It is hypothesized that the coexistence of low levels of G. lichenoides and B. amyloliquefaciens may result in a heightened presence of natural antioxidants.
In the process of nutrient assimilation, the intestines are essential due to the increased surface area for absorption provided by the folds within them [14]. In the present study, the experimental group demonstrated a greater abundance of intestinal tissue epithelia with well-defined brush borders and densely packed nuclei compared to the control group. Additionally, the study found that the height of intestinal epithelial cells in P. monodon fed a diet containing G. lichenoides was significantly greater than those in the S8B11 group, indicating that the inclusion of G. lichenoides in the diet may enhance the nutrient absorption capacity of the shrimp. Previous research has shown that seaweed (Enteromorpha) can increase villus width and lead to a larger villus surface area [63]. Paul et al. [72] illustrated that the inclusion of dietary K. alvarezii resulted in a notable enhancement of villus width and crypt depth within the intestine of broiler chicken. The supplementation of seaweeds in cultured animals has been shown to augment intestinal microvilli and increase absorptive surface area, thereby potentially enhancing nutrient utilization and promoting growth. In the current study, the highest values of intestine epithelium in P. monodon were observed in the S3 group. Additionally, elevated values of fold depth were noted in the S3B9, B9, and B11 groups, which were significantly greater than those in the control group. Previous research has shown that B. subtilis can enhance broiler villus height, villus surface area, and absorbing epithelial cell area [53], while the supplementation of B. licheniformis has been found to promote the development of intestinal villi in common carp, with well-developed and well-arranged villi [73]. These findings suggest that Bacillus have the potential to improve the structure integrity of intestinal tissue.
The intestinal microbiota serves as the primary locus for organ digestion and absorption, supplying essential nutrients and energy to the host organism while also exerting a significant influence on the growth, health, and developmental processes of shrimp [38]. In this study, there were no significant differences in the diversity and richness of microbial communities. Diversity and richness are comprehensive indicators composed of a large number of bacteria, indicating that the type of feed added does not change the total type and number of intestinal florae [38]. But there are differences in microbial community composition: at the phylum level, Proteobacteria, Bacteroidetes, Actinobacteria, and Fusobacteria were identified as the predominant bacterial taxa in the intestinal microbiota of P. monodon. Previous studies have shown that Proteobacteria, whose microbiota is dominated by Gram-negative bacteria, have a high risk of disease and reduce the health of the host [74]. In the study by Wang et al. [75], the microbial community of Proteobacteria in the intestine of L. vannabaeus was increased after white spot syndrome virus (WSSV) infection. Actinobacteria have been shown to degrade organic matter, synthesize antibacterial compounds [76], and modulate inflammatory and autoimmune responses through the induction of regulatory T cells, potentially enhancing disease resistance and immunity in shrimp [77,78]. Firmicutes and Bacteroidetes are generally the major microbiota in the gut of humans and other mammals, participating in the digestion and metabolism of food by the host [79], and are generally thought to contribute to improved digestion in aquatic animals [80]. The relative abundance of Firmicutes in the S3 group was significantly higher than that in the S3B9, S8, and B11 groups. Firmicutes is a commonly predominant microbial taxa in the gastrointestinal tract of humans and other mammals, playing a significant role in food digestion and metabolism within the host organism [73,81]. In aquatic animals, these microbial groups are believed to enhance digestive processes. Firmicutes have been shown to facilitate the absorption of fatty acids in host organisms and promote the synthesis of short-chain fatty acids [79], a process closely associated with the development of fatty liver [38]. In light of the alterations observed in the levels of FBW, WGR, SGR, and intestinal lipase activity within the S3 group, the findings indicate that the supplementation of G. lichenoides at a low concentration may enhance digestive enzyme activities and growth performance through modulation of the Firmicutes composition. At the taxonomic level of genus, we identified several dominant genera were identified that displayed significant differences among the groups, including Vibrio, Shimia, and Lactobacillus. Vibrio, a facultative pathogen, exhibited high enrichment in the intestinal microbiota of aquatic animals [82]. Our findings indicated a significantly relative abundance of Vibrio in the S3 group compared to the S3B9. Previous research by Duan et al. [83] supported the enrichment of Vibrio in the intestinal microbiota of the fast-growing shrimp, aligning with the results of our study. The growth performance of shrimp in the S3 group was significantly highest.
5. Conclusions
The inclusion of dietary G. lichenoides and B. amyloliquefaciens positively impacted the growth, antioxidant capacity, and microbial composition of shrimp. Adding 3% G. lichenoide had significantly higher FBW, WGR, and SGR; the antioxidant capacity in hepatopancreatic tissues of P. monodon was significantly increased in the 3% G. lichenoide group and the 3% G. lichenoide + 109 CFU/g B. amyloliquefaciens group; antioxidant gene expression was significantly increased in the hepatopancreas of shrimp between the 3% G. lichenoide group and the 3% G. lichenoide + 109 CFU/g B. amyloliquefaciens group; supplementing the diet with the G. lichenoide and B. amyloliquefacients improved the stomach and intestinal lipase activity of P. monodon; supplementing the diet with B. amyloliquefaciens (109 CFU/g) improved the intestinal tissue structure of P. monodon; adding B. amyloliquefaciens (109 CFU/g) and 8% G. lichenoide + B. amyloliquefaciens (1011 CFU/g) had a significant impact on the expression of genes involved in shrimp intestinal lipid metabolism; and the relative abundance of intestinal microbiome in the control, S3, B11, S3B9, and S8B11 groups had a significant impact. The results indicate that the inclusion of 3% G. lichenoides led to specific improvements in shrimp. This research investigated the impacts of G. lichenoide and B. amyloliquefaciens on growth, antioxidant capacity, and intestinal microbiome, offering a theoretical foundation for further study in this area.
Author Contributions
J.T.: designing and performing experiments and writing the manuscript; Y.W.: designing experiments and revising the manuscript; J.H.: review and revising the manuscript. H.Y.: collecting samples and statistical analyses; Y.D.: review and revising the manuscript; J.W.: review and revising the manuscript; C.Z.: review and revising the manuscript. Z.H.: review and revising the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded the Finance science and technology project of Hainan province (323MS126), Guangdong province Rural Science and Technology Commissioners Project, Guangdong Natural Science Foundation (2019A1515012159), Financial Special Project of the Ministry of Agriculture and Rural Areas (NHYYSWZZZYKZX2020), Central Public-interest Scientific Institution Basal Research Fund, CAFS (2020KX03, 2023TD97), Rural Revitalization Strategy Special Fund Seed Industry Revitalization Project of Guangdong Province (2022SPY02001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank the editor and anonymous reviewers for their comments that helped improve this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yang, W.Y.; Jiang, S.; Yang, Q.B.; Jiang, S.G.; Huang, J.H.; Yang, L.S.; Chen, X.; Li, Y.D.; Zhou, F.L. Effects of vitamin C on transport of Penaeus monodon. Isr. J. Aquac.-Bamidgeh 2023, 75, 1–10. [Google Scholar] [CrossRef]
- Tu, H.T.; Silvestre, F.; Wang, N.; Thome, J.P.; Phuong, N.T.; Kestemont, P. A multi-biomarker approach to assess the impact of farming systems on black tiger shrimp (Penaeus monodon). Chemosphere 2010, 81, 1204–1211. [Google Scholar] [CrossRef]
- Xie, J.J.; Chen, X.; Guo, T.Y.; Xie, S.W.; Fang, H.H.; Liu, Z.L.; Zhang, Y.M.; Tian, L.X.; Liu, Y.J.; Niu, J. Dietary values of Forsythia suspensa extract in Penaeus monodon under normal rearing and Vibrio parahaemolyticus 3HP (VP3HP) challenge conditions: Effect on growth, intestinal barrier function, immune response and immune related gene expression. Fish Shellfish Immunol. 2018, 75, 316–326. [Google Scholar] [CrossRef]
- Schleder, D.D.; Peruch, L.G.B.; Poli, M.A.; Ferreira, T.H.; Silva, C.P.; Andreatta, E.R.; Hayashi, L.; do Nascimento Vieira, F. Effect of brown seaweeds on pacific white shrimp growth performance, gut morphology, digestive enzymes activity and resistance to white spot virus. Aquaculture 2018, 495, 359–365. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Yang, Y.F.; Fei, X.G.; Song, J.M.; Hu, H.Y.; Wang, G.C.; Chung, I.K. Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters. Aquaculture 2006, 254, 248–255. [Google Scholar] [CrossRef]
- Xu, Y.; Fang, J.; Wei, W. Application of Gracilaria lichenoides (Rhodophyta) for alleviating excess nutrients in aquaculture. J. Appl. Phycol. 2008, 20, 199–203. [Google Scholar] [CrossRef]
- MacArtain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweed. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef]
- Burtin, P. Nutritional value of seaweeds. J. Agric. Food Chem. 2003, 2, 498–503. [Google Scholar]
- Moutinho, S.; Linares, F.; Rodríguez, J.L.; Sousa, V.; Valente, L.M.P. Inclusion of 10% seaweed meal in diets for juvenile and on-growing lifestages of senegalese sole (Solea senegalensis). J. Appl. Phycol. 2018, 30, 3589–3601. [Google Scholar] [CrossRef]
- Marinho, G.; Nunes, C.; Sousa-Pinto, I.; Pereira, R.; Rema, P.; Valente, L.M.P. The IMTA-cultivated Chlorophyta Ulva spp. as a sustainable ingredient in nile tilapia (Oreochromis niloticus) diets. J. Appl. Phycol. 2013, 25, 1359–1367. [Google Scholar] [CrossRef]
- Wassef, E.A.; El-Sayed, A.F.M.; Kandeel, K.M.; Sakr, E.M. Evaluation of Pterocladia (Rhodophyta) and Ulva (Chlorophyta) meals as additives to gilthead seabream Sparus aurata diets. Egypt. J. Aquat. Res. 2005, 31, 321–332. [Google Scholar]
- Mustafa, G.; Wakamatsu, S.; Takeda, T.; Umino, T.; Nakagawa, H. Effects of algae meal as feed additive on growth, feed efficiency, andbody composition in red sea bream. Fish. Sci. 1995, 61, 25–28. [Google Scholar] [CrossRef]
- Niu, J.; Chen, X.; Lu, X.; Jiang, S.-G.; Lin, H.-Z.; Liu, Y.-J.; Huang, Z.; Wang, J.; Wang, Y.; Tian, L.-X. Effects of different levels of dietary wakame (Undaria pinnatifida) on growth, immunity and intestinal structure of juvenile Penaeus monodon. Aquaculture 2015, 435, 78–85. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Araújo, M.; Batista, S.; Peixoto, M.J.; Sousa-Pinto, I.; Brotas, V.; Cunha, L.M.; Rema, P. Carotenoid deposition, flesh quality and immunological response of nile tilapia fed increasing levels of IMTA-cultivated Ulva spp. J. Appl. Phycol. 2016, 28, 691–701. [Google Scholar] [CrossRef]
- Lazo, J.P.; Dinis, M.T.; Holt, G.J.; Faulk, C.; Arnold, C.R. Co-Feeding microparticulate diets with algae: Toward eliminating the need of zooplankton at first feeding in larval red drum (Sciaenops ocellatus). Aquaculture 2000, 188, 339–351. [Google Scholar] [CrossRef]
- Sotoudeh, E.; Jafari, M. Effects of dietary supplementation with Red Seaweed, Gracilaria pygmaea, on growth, carcass composition and hematology of juvenile rainbow trout, Oncorhynchus Mykiss. Aquacult. Int. 2017, 25, 1857–1867. [Google Scholar] [CrossRef]
- Passos, R.; Correia, A.P.; Ferreira, I.; Pires, P.; Pires, D.; Gomes, E.; Do Carmo, B.; Santos, P.; Simões, M.; Afonso, C.; et al. Effect on health status and pathogen resistance of gilthead seabream (Sparus aurata) fed with diets supplemented with Gracilaria gracilis. Aquaculture 2021, 531, 735888. [Google Scholar] [CrossRef]
- Ragaza, J.A.; Koshio, S.; Mamauag, R.E.; Ishikawa, M.; Yokoyama, S.; Villamor, S.S. Dietary supplemental effects of red seaweed Eucheuma denticulatum on growth performance, carcass composition and blood chemistry of juvenile flounder, Paralichthys olivaceus. Aquac. Res. 2015, 46, 647–657. [Google Scholar] [CrossRef]
- Adel, M.; Omidi, A.H.; Dawood, M.A.O.; Karimi, B.; Shekarabi, S.P.H. Dietary Gracilaria persica mediated the growth performance, fillet colouration, and immune response of persian sturgeon (Acipenser persicus). Aquaculture 2021, 530, 735950. [Google Scholar] [CrossRef]
- Negm, S.S.; Ismael, N.E.M.; Ahmed, A.I.; Asely, A.M.E.; Naiel, M.A.E. The efficiency of dietary Sargassum aquifolium on the performance, innate immune responses, antioxidant activity, and intestinal microbiota of nile tilapia (Oreochromis niloticus) raised at high stocking density. J. Appl. Phycol. 2021, 33, 4067–4082. [Google Scholar] [CrossRef]
- Shi, Q.; Yu, C.; Zhu, D.; Li, S.; Wen, X. Effects of dietary Sargassum horneri on resisting hypoxia stress, which changes blood biochemistry, antioxidant status, and Hepatic HSP mRNA expressions of juvenile Black sea bream Acanthopagrus Schlegelii. J. Appl. Phycol. 2020, 32, 3457–3466. [Google Scholar] [CrossRef]
- Bakky, M.A.H.; Tran, N.T.; Zhang, Y.; Hu, H.; Lin, H.; Zhang, M.; Liang, H.; Zhang, Y.; Li, S. Effects of dietary supplementation of Gracilaria lemaneiformis-derived sulfated polysaccharides on the growth, antioxidant capacity, and innate immunity of rabbitfish (Siganus canaliculatus). Fish Shellfish Immunol. 2023, 139, 108933. [Google Scholar] [CrossRef]
- Heidarieh, M.; Mirvaghefi, A.R.; Akbari, M.; Farahmand, H.; Sheikhzadeh, N.; Shahbazfar, A.A.; Behgar, M. Effect of dietary ergosan on growth performance, digestive enzymes, intestinal histology, hematological parameters and body composition of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2012, 38, 1169–1174. [Google Scholar] [CrossRef]
- Silva, D.M.; Valente, L.M.P.; Sousa-Pinto, I.; Pereira, R.; Pires, M.A.; Seixas, F.; Rema, P. Evaluation of IMTA-produced seaweeds (Gracilaria, Porphyra, and Ulva) as dietary ingredients in Nile tilapia, Oreochromis niloticus L., juveniles. Effects on growth performance and gut histology. J. Appl. Phycol. 2015, 27, 1671–1680. [Google Scholar] [CrossRef]
- Araújo, M.; Rema, P.; Sousa-Pinto, I.; Cunha, L.M.; Peixoto, M.J.; Pires, M.A.; Seixas, F.; Brotas, V.; Beltrán, C.; Valente, L.M.P. Dietary inclusion of IMTA-cultivated Gracilaria vermiculophylla in rainbow trout (Oncorhynchus mykiss) diets: Effects on growth, intestinal morphology, tissue pigmentation, and immunological response. J. Appl. Phycol. 2016, 28, 679–689. [Google Scholar] [CrossRef]
- Yu, Y.-Y.; Chen, W.-D.; Liu, Y.-J.; Niu, J.; Chen, M.; Tian, L.-X. Effect of different dietary levels of Gracilaria lemaneiformis dry power on growth performance, hematological parameters and intestinal structure of juvenile pacific White shrimp (Litopenaeus vannamei). Aquaculture 2016, 450, 356–362. [Google Scholar] [CrossRef]
- Muttharasi, C.; Gayathri, V.; Muralisankar, T.; Mohan, K.; Uthayakumar, V.; Radhakrishnan, S.; Kumar, P.; Palanisamy, M. Growth Performance, Digestive enzymes and antioxidants activities in the shrimp Litopenaeus vannamei fed with Amphiroa fragilissima crude polysaccharides encapsulated Artemia nauplii. Aquaculture 2021, 545, 737263. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics: Table 1. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef]
- Kumar, V.; Roy, S.; Meena, D.K.; Sarkar, U.K. Application of probiotics in shrimp aquaculture: Importance, mechanisms of action, and methods of administration. Rev. Fish Sci. Aquac. 2016, 24, 342–368. [Google Scholar] [CrossRef]
- Xu, L.; Yuan, J.; Chen, X.; Zhang, S.; Xie, M.; Chen, C.; Wu, Z. Screening of intestinal probiotics and the effects of feeding probiotics on the digestive enzyme activity, immune, intestinal flora and WSSV resistance of Procambarus clarkii. Aquaculture 2021, 540, 736748. [Google Scholar] [CrossRef]
- Wang, R.; Guo, Z.; Tang, Y.; Kuang, J.; Duan, Y.; Lin, H.; Jiang, S.; Shu, H.; Huang, J. Effects on development and microbial community of shrimp Litopenaeus vannamei larvae with probiotics treatment. AMB Expr. 2020, 10, 109. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, Y.; Zhang, D.; Wang, Q.; Wang, G.; Elsadek, M.; Yao, Q.; Chen, Y.; Guo, Z. The effect of adding Bacillus amyloliquefaciens LSG2-8 in diets on the growth, immune function, antioxidant capacity, and disease resistance of Rhynchocypris lagowskii. Fish Shellfish Immunol. 2022, 125, 258–265. [Google Scholar] [CrossRef]
- Truong Thy, H.T.; Tri, N.N.; Quy, O.M.; Fotedar, R.; Kannika, K.; Unajak, S.; Areechon, N. Effects of the dietary supplementation of mixed probiotic spores of Bacillus amyloliquefaciens 54A, and Bacillus pumilus 47B on growth, innate immunity and stress responses of striped catfish (Pangasianodon hypophthalmus). Fish Shellfish Immunol. 2017, 60, 391–399. [Google Scholar] [CrossRef]
- Tseng, D.-Y.; Ho, P.-L.; Huang, S.-Y.; Cheng, S.-C.; Shiu, Y.-L.; Chiu, C.-S.; Liu, C.-H. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish Shellfish Immunol. 2009, 26, 339–344. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Saputra, F.; Chen, Y.-C.; Hu, S.-Y. Dietary administration of Bacillus amyloliquefaciens R8 reduces hepatic oxidative stress and enhances nutrient metabolism and immunity against Aeromonas hydrophila and Streptococcus agalactiae in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 86, 410–419. [Google Scholar] [CrossRef]
- Xu, P.W. Optimum Dietary Lipid Requirement and Short-Chain Fatty Acids Nutritional Function for Juvenile Golden Pompano (Trachinotus ovatus). Ph.D. Thesis, Shanghai Ocean University, Shanghai, China, 2022. [Google Scholar]
- Baur, F.J.; Ensminger, L.G. The Association of Official Analytical Chemists (AOAC). J. Am. Oil Chem. Soc. 1977, 54, 171–172. [Google Scholar] [CrossRef]
- Ming, J.; Ye, J.; Zhang, Y.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Dietary optimal reduced glutathione improves innate immunity, oxidative stress resistance and detoxification function of grass carp (Ctenopharyngodon idella) against Microcystin-LR. Aquaculture 2019, 498, 594–605. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Liu, Y.; Tian, L.; Niu, J. Dietary fish oil levels modulated lipid metabolism, immune response, intestinal health and salinity stress resistance of juvenile Penaeus monodon fed a low fish-meal diet. Anim. Feed Sci. Technol. 2022, 289, 115321. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sun, M.M.; Huang, J.H.; Yang, Q.B.; Zhou, F.L.; Wen, W.G.; Chen, X.; Jiang, S.G. Comparison on characteristics of growth and resistance to ammonia among 13 families of Penaeus monodon. J. Shanghai Ocean. Univ. 2011, 20, 510–516. [Google Scholar]
- Anil, K.S.; Balakrishanan, G.; Kanji, J.L.; Jitesh, S.B.; Kumaran, R. Comparison of Penaeus monodon (Crustacea, Penaeidae) growth between commercial feed vs. commercial shrimp feed supplemented with Kappaphycus alvarezii (Rhodophyta, Solieriaceae) seaweed sap. AACL Bioflux. 2011, 4, 292–300. [Google Scholar]
- Omont, A.; Quiroz-Guzman, E.; Tovar-Ramirez, D.; Peña-Rodríguez, A. Effect of diets supplemented with different seaweed extracts on growth performance and digestive enzyme activities of juvenile white shrimp Litopenaeus vannamei. J. Appl. Phycol. 2019, 31, 1433–1442. [Google Scholar] [CrossRef]
- Traifalgar, R.F.; Serrano, A.E.; CoRRE, V.; Kira, H.; TuNG, H.T.; Michael, F.R.; Koshio, S. Evaluation of dietary fucoidan supplementation effects on growth performance and vibriosis resistance of Penaeus monodon postlarvae. Aquacult. Sci. 2009, 57, 167–174. [Google Scholar]
- Guo, J.; Fu, Y.; Wu, Z.; Yu, X.; Guo, Y.; Liu, J.; Zhang, W.; Mai, K. Effects of dietary carbohydrate levels on growth performance, body composition, glucose/lipid metabolism and insulin signaling pathway in abalone Haliotis discus hannai. Aquaculture 2022, 557, 738284. [Google Scholar] [CrossRef]
- Bai, T. Study on Early Lipid Characteristics and Key Lipid Metabolism Enzymes of Schizothorax grahami. Master’s Thesis, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- Lu, L.; Luo, X.G.; Ji, C.; Liu, B.; Yu, S.X. Effect of manganese supplementation and source on carcass traits, meat quality, and lipid oxidation in broilers. J. Anim. Sci. 2007, 85, 812–822. [Google Scholar] [CrossRef]
- Zhao, L.L.; Wang, D.X.; Liao, Z.B.; Bi, Q.Z.; Ma, Q.; Wei, Y.L.; Liang, M.Q.; Qiao, X.T.; Cheng, Z.Y.; Xu, H.G. Tissue distribution and nutritional regulation of fatty acid transport protein Scopthalmus maximus and Takifugu rubripes. Chin. J. Anim. Nutr. 2022, 34, 6620–6633. [Google Scholar]
- Li, H.B.; Liu, F.; Fu, J.H.; Feng, L.; Dai, C.G.; Hu, Y. Cloning and spatio-temporal expression of lipophorin receptors genes related to reproduction in the oriental armyworm Mythimna separata. J. Plant Prot. 2022, 49, 741–748. [Google Scholar]
- Knights, K.M. Long-chain-fatty-acid coA ligases: The key to fatty acid activation, formation of xenobiotic acyl-coA thioesters and lipophilic xenobiotic conjugates. Curr. Med. Chem. 2003, 3, 235–244. [Google Scholar] [CrossRef]
- Al-Fataftah, A.-R.; Abdelqader, A. Effects of dietary Bacillus subtilis on heat-stressed broilers performance, intestinal morphology and microflora composition. Anim. Feed Sci. Technol. 2014, 198, 279–285. [Google Scholar] [CrossRef]
- Shi, F.; Zi, Y.; Lu, Z.; Li, F.; Yang, M.; Zhan, F.; Li, Y.; Li, J.; Zhao, L.; Lin, L.; et al. Bacillus subtilis H2 modulates immune response, fat metabolism and bacterial flora in the gut of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2020, 106, 8–20. [Google Scholar] [CrossRef]
- Zheng, J.; Duan, Y.; Yu, J.; Li, F.; Guo, Q.; Li, T.; Yin, Y. Effects of Long-term protein restriction on meat quality and muscle metabolites of shaziling pigs. Animals 2022, 12, 2007. [Google Scholar] [CrossRef]
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free. Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Wang, P.; Geng, J.; Gao, J.; Zhao, H.; Li, J.; Shi, Y.; Yang, B.; Xiao, C.; Linghu, Y.; Sun, X.; et al. Macrophage achieves self-protection against oxidative stress-induced ageing through the Mst-Nrf2 axis. Nat. Commun. 2019, 10, 755. [Google Scholar] [CrossRef]
- Abbas, H.H.H.; Authman, M.M.N. Effects of accumulated selenium on some physiological parameters and oxidative stress indicators in tilapia fish (Oreochromis spp.). Am. -Eurasian J. Agric. Environ. Sci. 2009, 5, 219–222. [Google Scholar]
- Chen, M.; Chen, X.-Q.; Tian, L.-X.; Liu, Y.-J.; Niu, J. Enhanced intestinal health, immune responses and ammonia resistance in Pacific white shrimp (Litopenaeus vannamei) fed dietary hydrolyzed yeast (Rhodotorula mucilaginosa) and Bacillus licheniformis. Aquac. Rep. 2020, 17, 100385. [Google Scholar] [CrossRef]
- Mohammadi, G.; Hafezieh, M.; Karimi, A.A.; Azra, M.N.; Van Doan, H.; Tapingkae, W.; Abdelrahman, H.A.; Dawood, M.A.O. The synergistic effects of plant polysaccharide and Pediococcus acidilactici as a synbiotic additive on growth, antioxidant status, immune response, and resistance of nile tilapia (Oreochromis niloticus) against Aeromonas hydrophila. Fish Shellfish Immunol. 2022, 120, 304–313. [Google Scholar] [CrossRef]
- Liu, P.-C.; Lin, P.-W.; Huang, C.-L.; Hsu, C.-H.; Chen, J.-C. Long-term administration of diets containing Gracilaria tenuistipitata extract induce the expression of immune-related genes and increase the immune response and resistance against Vibrio harveyi in white shrimp Litopenaeus vannamei. Gene Rep. 2019, 15, 100378. [Google Scholar] [CrossRef]
- Liu, W.-C.; Zhou, S.-H.; Balasubramanian, B.; Zeng, F.-Y.; Sun, C.-B.; Pang, H.-Y. Dietary seaweed (Enteromorpha) polysaccharides improves growth performance involved in regulation of immune responses, intestinal morphology and microbial community in banana shrimp Fenneropenaeus merguiensis. Fish Shellfish Immunol. 2020, 104, 202–212. [Google Scholar] [CrossRef]
- Arjeh, E.; Akhavan, H.-R.; Barzegar, M.; Carbonell-Barrachina, Á.A. Bio-active compounds and functional properties of pistachio hull: A review. Trends Food Sci. 2020, 97, 55–64. [Google Scholar] [CrossRef]
- Akbary, P.; Ajdari, A.; Ajang, B. Growth, survival, nutritional value and phytochemical, and antioxidant state of Litopenaeus vannamei shrimp fed with premix extract of brown Sargassum ilicifolium, Nizimuddinia zanardini, Cystoseira indica, and Padina australis macroalgae. Aquac. Int. 2023, 31, 681–701. [Google Scholar] [CrossRef]
- Mohammadi, G.; Adorian, T.J.; Rafiee, G. Beneficial effects of Bacillus subtilis on water quality, growth, immune responses, endotoxemia and protection against lipopolysaccharide-induced damages in oreochromis niloticus under biofloc technology system. Aquac. Nutr. 2020, 26, 1476–1492. [Google Scholar] [CrossRef]
- Tong, H.D.; Wang, J.M.; Song, Y. The role of Keap1-Nrf2-ARE in organismal defence against oxidative stress injury. Carcinog.,Teratog. Mutagen. 2013, 25, 71–75. [Google Scholar]
- Morilla, M.J.; Ghosal, K.; Romero, E.L. More than pigments: The potential of astaxanthin and bacterioruberin-based nanomedicines. Pharmaceutics 2023, 15, 1828. [Google Scholar] [CrossRef]
- Niemcharoen, S. Effects of Microplastics on Gene Expression to Nonspecific Immune System in Pacific White Shrimp (Litopenaeus vannamei). Master’s Thesis, Chulalongkorn University, Bangkok, Thailand, 2022. [Google Scholar]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalili, M.; Tabarraei, A.; Van Doan, H.; Spanò, N.; Faggio, C. Mucosal immune parameters, immune and antioxidant defence related genes expression and growth performance of zebrafish (Danio rerio) fed on Gracilaria gracilis powder. Fish Shellfish Immunol. 2018, 83, 232–237. [Google Scholar] [CrossRef]
- Ji, K.; Liang, H.; Ren, M.; Ge, X.; Mi, H.; Pan, L.; Yu, H. The immunoreaction and antioxidant capacity of juvenile blunt snout bream (Megalobrama amblycephala) involves the PI3K/Akt/Nrf2 and NF-κB signal pathways in response to dietary methionine levels. Fish Shellfish Immunol. 2020, 105, 126–134. [Google Scholar] [CrossRef]
- Paul, S.S.; Vantharam Venkata, H.G.R.; Raju, M.V.; Rama Rao, S.V.; Nori, S.S.; Suryanarayan, S.; Kumar, V.; Perveen, Z.; Prasad, C.S. Dietary supplementation of extracts of red sea weed (Kappaphycus alvarezii) improves growth, intestinal morphology, expression of intestinal genes and immune responses in broiler chickens. J. Sci. Food Agric. 2021, 101, 997–1008. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, M.; Feng, J.; Chen, Y.; Li, M.; Chang, X. Effects of dietary Bacillus licheniformis on growth performance, intestinal morphology, intestinal microbiome, and disease resistance in common carp (Cyprinus carpio L.). Aquac. Int. 2021, 29, 1343–1358. [Google Scholar] [CrossRef]
- Hasyimi, W.; Widanarni, W.; Yuhana, M. Growth performance and intestinal microbiota diversity in pacific white shrimp Litopenaeus vannamei fed with a probiotic bacterium, honey prebiotic, and synbiotic. Curr. Microbiol. 2020, 77, 2982–2990. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Xu, K.; Zhang, X.; Sun, H.; Fan, L.; Yan, M. White spot syndrome virus (WSSV) infection impacts intestinal microbiota composition and function in Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 84, 130–137. [Google Scholar] [CrossRef]
- Liu, L. Effects of Lactobacillus Pentosus Combined with Arthrospira platensis on the Growth Performance, Digestive Ability, Immunity, Intestinal Microbiota and Disease Resistance of Litopenaeus vannamei. Master’s Thesis, Hainan University, Haikou, China, 2022. [Google Scholar]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Sha, Y.; Liu, M.; Wang, B.; Jiang, K.; Qi, C.; Wang, L. Bacterial population in intestines of Litopenaeus vannamei fed different probiotics or probiotic supernatant. J. Microbiol. Biotechnol. 2016, 26, 1736–1745. [Google Scholar] [CrossRef]
- Costello, E.K.; Gordon, J.I.; Secor, S.M.; Knight, R. Postprandial remodeling of the gut microbiota in burmese pythons. ISME J. 2010, 4, 1375–1385. [Google Scholar] [CrossRef]
- Yuan, H.; Song, W.; Tan, J.; Zheng, Y.; Wang, H.; Shi, L.; Zhang, S. The effects of dietary protein level on the growth performance, body composition, intestinal digestion and microbiota of Litopenaeus vannamei fed Chlorella Sorokiniana as the main protein source. Animals 2023, 13, 2881. [Google Scholar] [CrossRef]
- Tian, Z.-K.; Zhang, Y.-J.; Feng, Z.-J.; Jiang, H.; Cheng, C.; Sun, J.-M.; Liu, C.-M. Nephroprotective effect of gastrodin against lead-induced oxidative stress and inflammation in mice by the GSH, Trx, Nrf2 antioxidant system, and the HMGB1 pathway. Toxicol. Res. 2021, 10, 249–263. [Google Scholar] [CrossRef]
- Xiong, J.B.; Wang, K.; Wu, J.F.; Qiuqian, L.L.; Yang, K.L.; Qian, Y.X.; Zhang, D. Changs in intestinal bacterial communities are closely associated with shrimp disease severity. Appl. Microbiol. Biotechnol. 2015, 99, 6911–6919. [Google Scholar] [CrossRef]
- Duan, Y.; Huang, J.; Wang, Y.; Zhang, J. Characterization of bacterial community in intestinal and rearing water of Penaeus monodon differing growth performances in outdoor and indoor ponds. Aquac. Res. 2020, 51, 4279–4289. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).